
Protein Arginine Methyltransferases as Therapeutic Targets in Hematological Malignancies

 , , and
, , and
Abstract
Simple Summary
Abstract
1. Introduction
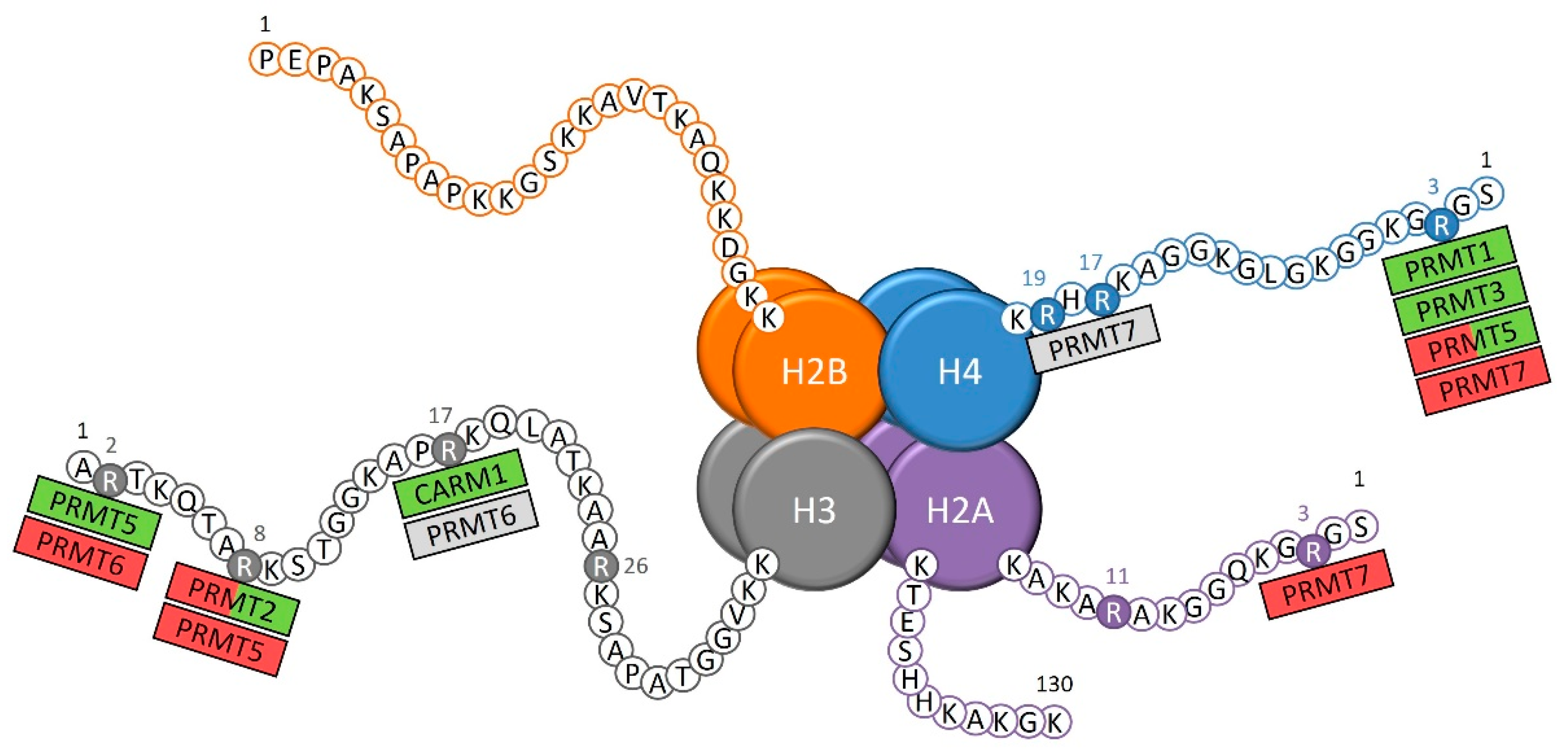
1.1. Arginine Methylation
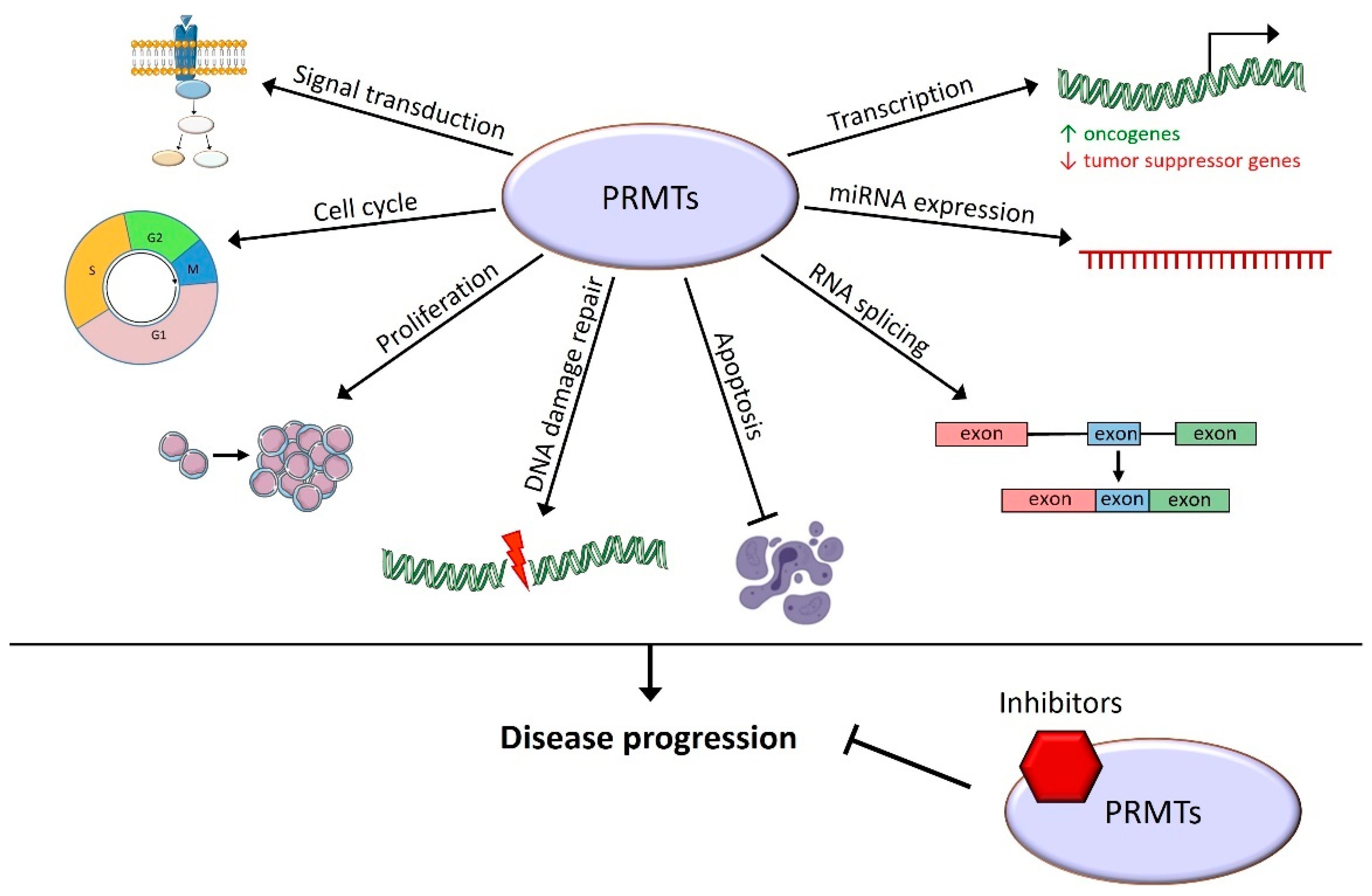
1.2. Epigenetic and Cellular Roles of PRMTs
1.3. PRMTs and Cancer
2. Involvement of PRMTs in Hematopoiesis and Hematological Malignancies
2.1. PRMT1
2.2. CARM1 (PRMT4)
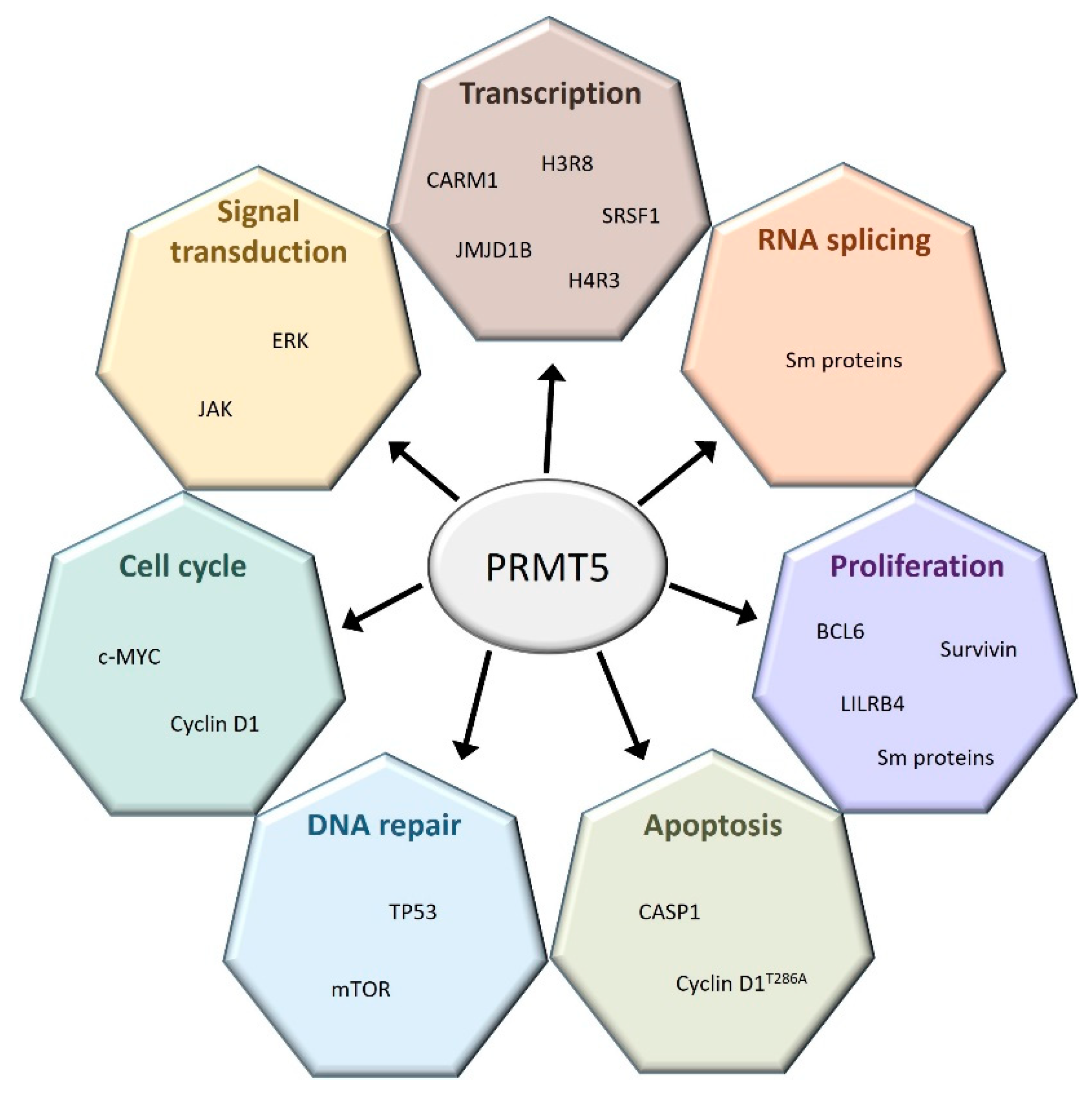
2.3. PRMT5
2.4. PRMT6
2.5. PRMT7
2.6. Other PRMTs
3. Pharmacological Inhibition of PRMTs in Hematological Malignancies
4. Clinical Trials
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Paik, W.K.; Kim, S. Enzymatic Methylation of Protein Fractions from Calf Thymus Nuclei. Biochem. Biophys. Res. Commun. 1967, 29, 14–20. [Google Scholar] [CrossRef]
- Bedford, M.T.; Clarke, S.G. Protein Arginine Methylation in Mammals: Who, What, and Why. Mol. Cell 2009, 33, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Blanc, R.S.; Richard, S. Arginine Methylation: The Coming of Age. Mol. Cell 2017, 65, 8–24. [Google Scholar] [CrossRef]
- Guccione, E.; Bassi, C.; Casadio, F.; Martinato, F.; Cesaroni, M.; Schuchlautz, H.; Lüscher, B.; Amati, B. Methylation of Histone H3R2 by PRMT6 and H3K4 by an MLL Complex Are Mutually Exclusive. Nature 2007, 449, 933–937. [Google Scholar] [CrossRef]
- Wu, Q.; Schapira, M.; Arrowsmith, C.H.; Barsyte-Lovejoy, D. Protein Arginine Methylation: From Enigmatic Functions to Therapeutic Targeting. Nat. Rev. Drug Discov. 2021, 20, 509–530. [Google Scholar] [CrossRef] [PubMed]
- Lorton, B.M.; Shechter, D. Cellular Consequences of Arginine Methylation. Cell Mol. Life Sci. 2019, 76, 2933–2956. [Google Scholar] [CrossRef]
- Lorenzo, A.D.; Bedford, M.T. Histone Arginine Methylation. FEBS Lett. 2011, 585, 2024–2031. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Teng, Y.; Wang, L.; Zhang, Z.; Chen, C.; Wang, Y.; Zhang, X.; Xiang, P.; Song, X.; Lu, J.; et al. LINC01431 Promotes Histone H4R3 Methylation to Impede HBV Covalently Closed Circular DNA Transcription by Stabilizing PRMT1. Adv. Sci. 2022, 9, 2103135. [Google Scholar] [CrossRef]
- Min, Z.; Xiaomeng, L.; Zheng, L.; Yangge, D.; Xuejiao, L.; Longwei, L.; Xiao, Z.; Yunsong, L.; Ping, Z.; Yongsheng, Z. Asymmetrical Methyltransferase PRMT3 Regulates Human Mesenchymal Stem Cell Osteogenesis via MiR-3648. Cell Death Dis. 2019, 10, 581. [Google Scholar] [CrossRef]
- Cheng, D.; Gao, G.; Lorenzo, A.D.; Jayne, S.; Hottiger, M.O.; Richard, S.; Bedford, M.T. Genetic Evidence for Partial Redundancy between the Arginine Methyltransferases CARM1 and PRMT6. J. Biol. Chem. 2020, 295, 17060–17070. [Google Scholar] [CrossRef]
- Kim, J.K.; Lim, Y.; Lee, J.O.; Lee, Y.-S.; Won, N.H.; Kim, H.; Kim, H.S. PRMT4 Is Involved in Insulin Secretion via the Methylation of Histone H3 in Pancreatic β Cells. J. Mol. Endocrinol. 2015, 54, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Chiang, K.; Zielinska, A.E.; Shaaban, A.M.; Sanchez-Bailon, M.P.; Jarrold, J.; Clarke, T.L.; Zhang, J.; Francis, A.; Jones, L.J.; Smith, S.; et al. PRMT5 Is a Critical Regulator of Breast Cancer Stem Cell Function via Histone Methylation and FOXP1 Expression. Cell Rep. 2017, 21, 3498–3513. [Google Scholar] [CrossRef] [PubMed]
- Karkhanis, V.; Wang, L.; Tae, S.; Hu, Y.-J.; Imbalzano, A.N.; Sif, S. Protein Arginine Methyltransferase 7 Regulates Cellular Response to DNA Damage by Methylating Promoter Histones H2A and H4 of the Polymerase δ Catalytic Subunit Gene, POLD1. J. Biol. Chem. 2012, 287, 29801–29814. [Google Scholar] [CrossRef] [PubMed]
- Ge, S.; Zhang, Q.; Chen, Y.; Tian, Y.; Yang, R.; Chen, X.; Li, F.; Zhang, B. Ribavirin Inhibits Colorectal Cancer Growth by Downregulating PRMT5 Expression and H3R8me2s and H4R3me2s Accumulation. Toxicol. Appl. Pharm. 2021, 415, 115450. [Google Scholar] [CrossRef] [PubMed]
- Okuno, K.; Akiyama, Y.; Shimada, S.; Nakagawa, M.; Tanioka, T.; Inokuchi, M.; Yamaoka, S.; Kojima, K.; Tanaka, S. Asymmetric Dimethylation at Histone H3 Arginine 2 by PRMT6 in Gastric Cancer Progression. Carcinogenesis 2018, 40, 15–26. [Google Scholar] [CrossRef]
- Fulton, M.D.; Brown, T.; Zheng, Y.G. Mechanisms and Inhibitors of Histone Arginine Methylation. Chem. Rec. 2018, 18, 1792–1807. [Google Scholar] [CrossRef]
- Cai, S.; Wang, P.; Xie, T.; Li, Z.; Li, J.; Lan, R.; Ding, Y.; Lu, J.; Ye, J.; Wang, J.; et al. Histone H4R3 Symmetric Di-Methylation by Prmt5 Protects against Cardiac Hypertrophy via Regulation of Filip1L/β-Catenin. Pharmacol. Res. 2020, 161, 105104. [Google Scholar] [CrossRef]
- Tarighat, S.S.; Santhanam, R.; Frankhouser, D.; Radomska, H.S.; Lai, H.; Anghelina, M.; Wang, H.; Huang, X.; Alinari, L.; Walker, A.; et al. The Dual Epigenetic Role of PRMT5 in Acute Myeloid Leukemia: Gene Activation and Repression via Histone Arginine Methylation. Leukemia 2016, 30, 789–799. [Google Scholar] [CrossRef]
- Dong, F.; Li, Q.; Yang, C.; Huo, D.; Wang, X.; Ai, C.; Kong, Y.; Sun, X.; Wang, W.; Zhou, Y.; et al. PRMT2 Links Histone H3R8 Asymmetric Dimethylation to Oncogenic Activation and Tumorigenesis of Glioblastoma. Nat. Commun. 2018, 9, 4552. [Google Scholar] [CrossRef]
- Hu, G.; Yan, C.; Xie, P.; Cao, Y.; Shao, J.; Ge, J. PRMT2 Accelerates Tumorigenesis of Hepatocellular Carcinoma by Activating Bcl2 via Histone H3R8 Methylation. Exp. Cell Res. 2020, 394, 112152. [Google Scholar] [CrossRef]
- Li, J.; Pan, X.; Ren, Z.; Li, B.; Liu, H.; Wu, C.; Dong, X.; Vos, P.; Pan, L.; Sun, J. Protein Arginine Methyltransferase 2 (PRMT2) Promotes Dextran Sulfate Sodium-induced Colitis by Inhibiting the SOCS3 Promoter via Histone H3R8 Asymmetric Dimethylation. Br. J. Pharmacol. 2022, 179, 141–158. [Google Scholar] [CrossRef] [PubMed]
- Blythe, S.A.; Cha, S.-W.; Tadjuidje, E.; Heasman, J.; Klein, P.S. β-Catenin Primes Organizer Gene Expression by Recruiting a Histone H3 Arginine 8 Methyltransferase, Prmt2. Dev. Cell 2010, 19, 220–231. [Google Scholar] [CrossRef] [PubMed]
- Cura, V.; Cavarelli, J. Structure, Activity and Function of the PRMT2 Protein Arginine Methyltransferase. Life 2021, 11, 1263. [Google Scholar] [CrossRef] [PubMed]
- Jain, K.; Jin, C.Y.; Clarke, S.G. Epigenetic Control via Allosteric Regulation of Mammalian Protein Arginine Methyltransferases. Proc. Natl. Acad. Sci. USA 2017, 114, 10101–10106. [Google Scholar] [CrossRef]
- Yang, Y.; Bedford, M.T. Protein Arginine Methyltransferases and Cancer. Nat. Rev. Cancer 2013, 13, 37–50. [Google Scholar] [CrossRef]
- Hwang, J.W.; Cho, Y.; Bae, G.-U.; Kim, S.-N.; Kim, Y.K. Protein Arginine Methyltransferases: Promising Targets for Cancer Therapy. Exp. Mol. Med. 2021, 53, 788–808. [Google Scholar] [CrossRef]
- Wang, S.-C.M.; Dowhan, D.H.; Muscat, G.E.O. Epigenetic Arginine Methylation in Breast Cancer: Emerging Therapeutic Strategies. J. Mol. Endocrinol. 2019, 62, R223–R237. [Google Scholar] [CrossRef]
- Bryant, J.-P.; Heiss, J.; Banasavadi-Siddegowda, Y.K. Arginine Methylation in Brain Tumors: Tumor Biology and Therapeutic Strategies. Cells 2021, 10, 124. [Google Scholar] [CrossRef]
- Xie, V.K.; He, J.; Xie, K. Protein Arginine Methylation Promotes Therapeutic Resistance in Human Pancreatic Cancer. Cytokine Growth Factor Rev. 2020, 55, 58–69. [Google Scholar] [CrossRef]
- Tang, J.; Frankel, A.; Cook, R.J.; Kim, S.; Paik, W.K.; Williams, K.R.; Clarke, S.; Herschman, H.R. PRMT1 Is the Predominant Type I Protein Arginine Methyltransferase in Mammalian Cells. J. Biol. Chem. 2000, 275, 7723–7730. [Google Scholar] [CrossRef]
- Zhao, X.; Jankovic, V.; Gural, A.; Huang, G.; Pardanani, A.; Menendez, S.; Zhang, J.; Dunne, R.; Xiao, A.; Erdjument-Bromage, H.; et al. Methylation of RUNX1 by PRMT1 Abrogates SIN3A Binding and Potentiates Its Transcriptional Activity. Gene Dev. 2008, 22, 640–653. [Google Scholar] [CrossRef] [PubMed]
- Mizutani, S.; Yoshida, T.; Zhao, X.; Nimer, S.D.; Taniwaki, M.; Okuda, T. Loss of RUNX1/AML1 Arginine-methylation Impairs Peripheral T Cell Homeostasis. Br. J. Haematol. 2015, 170, 859–873. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.-I.; Hua, W.-K.; Yao, C.-L.; Hwang, S.-M.; Hung, Y.-C.; Kuan, C.-J.; Leou, J.-S.; Lin, W.-J. Protein-Arginine Methyltransferase 1 Suppresses Megakaryocytic Differentiation via Modulation of the P38 MAPK Pathway in K562 Cells. J. Biol. Chem. 2010, 285, 20595–20606. [Google Scholar] [CrossRef] [PubMed]
- Su, H.; Jiang, M.; Senevirathne, C.; Aluri, S.; Zhang, T.; Guo, H.; Xavier-Ferrucio, J.; Jin, S.; Tran, N.-T.; Liu, S.-M.; et al. Methylation of Dual-Specificity Phosphatase 4 Controls Cell Differentiation. Cell Rep. 2021, 36, 109421. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Tran, N.-T.; Su, H.; Wang, R.; Lu, Y.; Tang, H.; Aoyagi, S.; Guo, A.; Khodadadi-Jamayran, A.; Zhou, D.; et al. Cross-Talk between PRMT1-Mediated Methylation and Ubiquitylation on RBM15 Controls RNA Splicing. Elife 2015, 4, e07938. [Google Scholar] [CrossRef]
- Zhu, L.; He, X.; Dong, H.; Sun, J.; Wang, H.; Zhu, Y.; Huang, F.; Zou, J.; Chen, Z.; Zhao, X.; et al. Protein Arginine Methyltransferase 1 Is Required for Maintenance of Normal Adult Hematopoiesis. Int. J. Biol. Sci. 2019, 15, 2763–2773. [Google Scholar] [CrossRef]
- Su, H.; Sun, C.-W.; Liu, S.-M.; He, X.; Hu, H.; Pawlik, K.M.; Townes, T.M.; Han, X.; Klug, C.A.; Henary, M.; et al. Defining the Epigenetic Status of Blood Cells Using a Cyanine-Based Fluorescent Probe for PRMT1. Blood Adv. 2018, 2, 2829–2836. [Google Scholar] [CrossRef]
- Gupta, P.; Singh, A.; Gowda, P.; Ghosh, S.; Chatterjee, A.; Sen, E. Lactate Induced HIF-1α-PRMT1 Cross Talk Affects MHC I Expression in Monocytes. Exp. Cell Res. 2016, 347, 293–300. [Google Scholar] [CrossRef]
- Fan, Z.; Li, J.; Li, P.; Ye, Q.; Xu, H.; Wu, X.; Xu, Y. Protein Arginine Methyltransferase 1 (PRMT1) Represses MHC II Transcription in Macrophages by Methylating CIITA. Sci. Rep. 2017, 7, 40531. [Google Scholar] [CrossRef]
- Cho, J.-H.; Lee, R.; Kim, E.; Choi, Y.E.; Choi, E.-J. PRMT1 Negatively Regulates Activation-Induced Cell Death in Macrophages by Arginine Methylation of GAPDH. Exp. Cell Res. 2018, 368, 50–58. [Google Scholar] [CrossRef]
- Dolezal, E.; Infantino, S.; Drepper, F.; Börsig, T.; Singh, A.; Wossning, T.; Fiala, G.J.; Minguet, S.; Warscheid, B.; Tarlinton, D.M.; et al. The BTG2-PRMT1 Module Limits Pre-B Cell Expansion by Regulating the CDK4-Cyclin-D3 Complex. Nat. Immunol. 2017, 18, 911–920. [Google Scholar] [CrossRef] [PubMed]
- Infantino, S.; Light, A.; O’Donnell, K.; Bryant, V.; Avery, D.T.; Elliott, M.; Tangye, S.G.; Belz, G.; Mackay, F.; Richard, S.; et al. Arginine Methylation Catalyzed by PRMT1 Is Required for B Cell Activation and Differentiation. Nat. Commun. 2017, 8, 891. [Google Scholar] [CrossRef] [PubMed]
- Hata, K.; Yanase, N.; Sudo, K.; Kiyonari, H.; Mukumoto, Y.; Mizuguchi, J.; Yokosuka, T. Differential Regulation of T-cell Dependent and T-cell Independent Antibody Responses through Arginine Methyltransferase PRMT1 in Vivo. FEBS Lett. 2016, 590, 1200–1210. [Google Scholar] [CrossRef]
- Sung, B.-Y.; Lin, Y.-H.; Kong, Q.; Shah, P.D.; Bieler, J.G.; Palmer, S.; Weinhold, K.J.; Chang, H.-R.; Huang, H.; Avery, R.K.; et al. Wnt Activation Promotes Memory T Cell Polyfunctionality via Epigenetic Regulator PRMT1. J. Clin. Investig. 2022, 132, e140508. [Google Scholar] [CrossRef] [PubMed]
- Sen, S.; He, Z.; Ghosh, S.; Dery, K.J.; Yang, L.; Zhang, J.; Sun, Z. PRMT1 Plays a Critical Role in Th17 Differentiation by Regulating Reciprocal Recruitment of STAT3 and STAT5. J. Immunol. 2018, 201, 440–450. [Google Scholar] [CrossRef] [PubMed]
- Kagoya, Y.; Saijo, H.; Matsunaga, Y.; Guo, T.; Saso, K.; Anczurowski, M.; Wang, C.-H.; Sugata, K.; Murata, K.; Butler, M.O.; et al. Arginine Methylation of FOXP3 Is Crucial for the Suppressive Function of Regulatory T Cells. J. Autoimmun. 2019, 97, 10–21. [Google Scholar] [CrossRef]
- Vadnais, C.; Chen, R.; Fraszczak, J.; Yu, Z.; Boulais, J.; Pinder, J.; Frank, D.; Khandanpour, C.; Hébert, J.; Dellaire, G.; et al. GFI1 Facilitates Efficient DNA Repair by Regulating PRMT1 Dependent Methylation of MRE11 and 53BP1. Nat. Commun. 2018, 9, 1418. [Google Scholar] [CrossRef]
- He, X.; Zhu, Y.; Lin, Y.-C.; Li, M.; Du, J.; Dong, H.; Sun, J.; Zhu, L.; Wang, H.; Ding, Z.; et al. PRMT1-Mediated FLT3 Arginine Methylation Promotes Maintenance of FLT3-ITD+ Acute Myeloid Leukemia. Blood 2019, 134, 548–560. [Google Scholar] [CrossRef]
- Siriboonpiputtana, T.; Zeisig, B.B.; Zarowiecki, M.; Fung, T.K.; Mallardo, M.; Tsai, C.; Lau, P.N.I.; Hoang, Q.C.; Veiga, P.; Barnes, J.; et al. Transcriptional Memory of Cells of Origin Overrides Β-catenin Requirement of MLL Cancer Stem Cells. EMBO J. 2017, 36, 3139–3155. [Google Scholar] [CrossRef]
- Cheung, N.; Chan, L.C.; Thompson, A.; Cleary, M.L.; So, C.W.E. Protein Arginine-Methyltransferase-Dependent Oncogenesis. Nat. Cell Biol. 2007, 9, 1208–1215. [Google Scholar] [CrossRef]
- Cheung, N.; Fung, T.K.; Zeisig, B.B.; Holmes, K.; Rane, J.K.; Mowen, K.A.; Finn, M.G.; Lenhard, B.; Chan, L.C.; So, C.W.E. Targeting Aberrant Epigenetic Networks Mediated by PRMT1 and KDM4C in Acute Myeloid Leukemia. Cancer Cell 2016, 29, 32–48. [Google Scholar] [CrossRef] [PubMed]
- Shia, W.-J.; Okumura, A.J.; Yan, M.; Sarkeshik, A.; Lo, M.-C.; Matsuura, S.; Komeno, Y.; Zhao, X.; Nimer, S.D.; Yates, J.R.; et al. PRMT1 Interacts with AML1-ETO to Promote Its Transcriptional Activation and Progenitor Cell Proliferative Potential. Blood 2012, 119, 4953–4962. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Mi, Y.; Song, J.; Zhang, P.; Liu, Y. PRMT1-RBM15 Axis Regulates Megakaryocytic Differentiation of Human Umbilical Cord Blood CD34+ Cells. Exp. Ther. Med. 2018, 15, 2563–2568. [Google Scholar] [CrossRef] [PubMed]
- Zou, L.; Zhang, H.; Du, C.; Liu, X.; Zhu, S.; Zhang, W.; Li, Z.; Gao, C.; Zhao, X.; Mei, M.; et al. Correlation of SRSF1 and PRMT1 Expression with Clinical Status of Pediatric Acute Lymphoblastic Leukemia. J. Hematol. Oncol. 2012, 5, 42. [Google Scholar] [CrossRef]
- Li, J.; Zhao, Z.; Carter, C.; Ehrlich, L.I.R.; Bedford, M.T.; Richie, E.R. Coactivator-Associated Arginine Methyltransferase 1 Regulates Fetal Hematopoiesis and Thymocyte Development. J. Immunol. 2013, 190, 597–604. [Google Scholar] [CrossRef]
- Kim, J.; Lee, J.; Yadav, N.; Wu, Q.; Carter, C.; Richard, S.; Richie, E.; Bedford, M.T. Loss of CARM1 Results in Hypomethylation of Thymocyte Cyclic AMP-Regulated Phosphoprotein and Deregulated Early T Cell Development. J. Biol. Chem. 2004, 279, 25339–25344. [Google Scholar] [CrossRef]
- Lai, Y.; Li, X.; Li, T.; Li, X.; Nyunoya, T.; Chen, K.; Kitsios, G.; Nouraie, M.; Zhang, Y.; McVerry, B.J.; et al. Protein Arginine N-Methyltransferase 4 (PRMT4) Contributes to Lymphopenia in Experimental Sepsis. Thorax 2022, 1–11. [Google Scholar] [CrossRef]
- Zika, E.; Fauquier, L.; Vandel, L.; Ting, J.P.-Y. Interplay among Coactivator-Associated Arginine Methyltransferase 1, CBP, and CIITA in IFN-γ-Inducible MHC-II Gene Expression. Proc. Natl. Acad. Sci. USA 2005, 102, 16321–16326. [Google Scholar] [CrossRef]
- Streubel, G.; Bouchard, C.; Berberich, H.; Zeller, M.S.; Teichmann, S.; Adamkiewicz, J.; Müller, R.; Klempnauer, K.-H.; Bauer, U.-M. PRMT4 Is a Novel Coactivator of C-Myb-Dependent Transcription in Haematopoietic Cell Lines. PLoS Genet. 2013, 9, e1003343. [Google Scholar] [CrossRef]
- Greenblatt, S.M.; Man, N.; Hamard, P.-J.; Asai, T.; Karl, D.; Martinez, C.; Bilbao, D.; Stathias, V.; Jermakowicz, A.M.; Duffort, S.; et al. CARM1 Is Essential for Myeloid Leukemogenesis but Dispensable for Normal Hematopoiesis. Cancer Cell 2018, 33, 1111–1127.e5. [Google Scholar] [CrossRef]
- Vu, L.P.; Perna, F.; Wang, L.; Voza, F.; Figueroa, M.E.; Tempst, P.; Erdjument-Bromage, H.; Gao, R.; Chen, S.; Paietta, E.; et al. PRMT4 Blocks Myeloid Differentiation by Assembling a Methyl-RUNX1-Dependent Repressor Complex. Cell Rep. 2013, 5, 1625–1638. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Zhao, X.; Perna, F.; Wang, L.; Koppikar, P.; Abdel-Wahab, O.; Harr, M.W.; Levine, R.L.; Xu, H.; Tefferi, A.; et al. JAK2V617F-Mediated Phosphorylation of PRMT5 Downregulates Its Methyltransferase Activity and Promotes Myeloproliferation. Cancer Cell 2011, 19, 283–294. [Google Scholar] [CrossRef]
- Liu, F.; Cheng, G.; Hamard, P.-J.; Greenblatt, S.; Wang, L.; Man, N.; Perna, F.; Xu, H.; Tadi, M.; Luciani, L.; et al. Arginine Methyltransferase PRMT5 Is Essential for Sustaining Normal Adult Hematopoiesis. J. Clin. Investig. 2015, 125, 3532–3544. [Google Scholar] [CrossRef]
- Hamard, P.-J.; Santiago, G.E.; Liu, F.; Karl, D.L.; Martinez, C.; Man, N.; Mookhtiar, A.K.; Duffort, S.; Greenblatt, S.; Verdun, R.E.; et al. PRMT5 Regulates DNA Repair by Controlling the Alternative Splicing of Histone-Modifying Enzymes. Cell Rep. 2018, 24, 2643–2657. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.Q.; Li, Y.; Yang, C.; Li, J.; Tan, S.H.; Chin, D.W.L.; Nakamura-Ishizu, A.; Yang, H.; Suda, T. PRMT5 Modulates Splicing for Genome Integrity and Preserves Proteostasis of Hematopoietic Stem Cells. Cell Rep. 2019, 26, 2316–2328.e6. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Ali, S.; Duan, X.; Liu, S.; Du, J.; Liu, C.; Dai, H.; Zhou, M.; Zhou, L.; Yang, L.; et al. JMJD1B Demethylates H4R3me2s and H3K9me2 to Facilitate Gene Expression for Development of Hematopoietic Stem and Progenitor Cells. Cell Rep. 2018, 23, 389–403. [Google Scholar] [CrossRef]
- Quillien, A.; Gilbert, G.; Boulet, M.; Ethuin, S.; Waltzer, L.; Vandel, L. Prmt5 Promotes Vascular Morphogenesis Independently of Its Methyltransferase Activity. PLoS Genet. 2021, 17, e1009641. [Google Scholar] [CrossRef]
- Zheng, Y.; Chen, Z.; Zhou, B.; Chen, S.; Han, L.; Chen, N.; Ma, Y.; Xie, G.; Yang, J.; Nie, H.; et al. PRMT5 Deficiency Enforces the Transcriptional and Epigenetic Programs of Klrg1+CD8+ Terminal Effector T Cells and Promotes Cancer Development. J. Immunol. 2021, 208, ji2100523. [Google Scholar] [CrossRef]
- Gullà, A.; Hideshima, T.; Bianchi, G.; Fulciniti, M.; Samur, M.K.; Qi, J.; Tai, Y.-T.; Harada, T.; Morelli, E.; Amodio, N.; et al. Protein Arginine Methyltransferase 5 Has Prognostic Relevance and Is a Druggable Target in Multiple Myeloma. Leukemia 2018, 32, 996–1002. [Google Scholar] [CrossRef]
- Xia, T.; Liu, M.; Zhao, Q.; Ouyang, J.; Xu, P.; Chen, B. PRMT5 Regulates Cell Pyroptosis by Silencing CASP1 in Multiple Myeloma. Cell Death Dis. 2021, 12, 851. [Google Scholar] [CrossRef]
- Wang, L.; Pal, S.; Sif, S. Protein Arginine Methyltransferase 5 Suppresses the Transcription of the RB Family of Tumor Suppressors in Leukemia and Lymphoma Cells. Mol. Cell Biol. 2008, 28, 6262–6277. [Google Scholar] [CrossRef] [PubMed]
- Pal, S.; Baiocchi, R.A.; Byrd, J.C.; Grever, M.R.; Jacob, S.T.; Sif, S. Low Levels of MiR-92b/96 Induce PRMT5 Translation and H3R8/H4R3 Methylation in Mantle Cell Lymphoma. EMBO J. 2007, 26, 3558–3569. [Google Scholar] [CrossRef]
- Karkhanis, V.; Alinari, L.; Ozer, H.G.; Chung, J.; Zhang, X.; Sif, S.; Baiocchi, R.A. Protein Arginine Methyltransferase 5 Represses Tumor Suppressor MiRNAs That Down-Regulate CYCLIN D1 and c-MYC Expression in Aggressive B-Cell Lymphoma. J. Biol. Chem. 2020, 295, 1165–1180. [Google Scholar] [CrossRef]
- Li, Y.; Chitnis, N.; Nakagawa, H.; Kita, Y.; Natsugoe, S.; Yang, Y.; Li, Z.; Wasik, M.; Klein-Szanto, A.J.P.; Rustgi, A.K.; et al. PRMT5 Is Required for Lymphomagenesis Triggered by Multiple Oncogenic Drivers. Cancer Discov. 2015, 5, 288–303. [Google Scholar] [CrossRef] [PubMed]
- Chung, J.; Karkhanis, V.; Tae, S.; Yan, F.; Smith, P.; Ayers, L.W.; Agostinelli, C.; Pileri, S.; Denis, G.V.; Baiocchi, R.A.; et al. Protein Arginine Methyltransferase 5 (PRMT5) Inhibition Induces Lymphoma Cell Death through Reactivation of the Retinoblastoma Tumor Suppressor Pathway and Polycomb Repressor Complex 2 (PRC2) Silencing. J. Biol. Chem. 2013, 288, 35534–35547. [Google Scholar] [CrossRef] [PubMed]
- Chung, J.; Karkhanis, V.; Baiocchi, R.A.; Sif, S. Protein Arginine Methyltransferase 5 (PRMT5) Promotes Survival of Lymphoma Cells via Activation of WNT/β-Catenin and AKT/GSK3β Proliferative Signaling. J. Biol. Chem. 2019, 294, 7692–7710. [Google Scholar] [CrossRef] [PubMed]
- Koh, C.M.; Bezzi, M.; Low, D.H.P.; Ang, W.X.; Teo, S.X.; Gay, F.P.H.; Al-Haddawi, M.; Tan, S.Y.; Osato, M.; Sabò, A.; et al. MYC Regulates the Core Pre-MRNA Splicing Machinery as an Essential Step in Lymphomagenesis. Nature 2015, 523, 96–100. [Google Scholar] [CrossRef]
- Zhu, F.; Guo, H.; Bates, P.D.; Zhang, S.; Zhang, H.; Nomie, K.J.; Li, Y.; Lu, L.; Seibold, K.R.; Wang, F.; et al. PRMT5 Is Upregulated by B-Cell Receptor Signaling and Forms a Positive-Feedback Loop with PI3K/AKT in Lymphoma Cells. Leukemia 2019, 33, 2898–2911. [Google Scholar] [CrossRef]
- Lu, X.; Fernando, T.M.; Lossos, C.; Yusufova, N.; Liu, F.; Fontán, L.; Durant, M.; Geng, H.; Melnick, J.; Luo, Y.; et al. PRMT5 Interacts with the BCL6 Oncoprotein and Is Required for Germinal Center Formation and Lymphoma Cell Survival. Blood 2018, 132, 2026–2039. [Google Scholar] [CrossRef]
- Liu, F.; Xu, Y.; Lu, X.; Hamard, P.-J.; Karl, D.L.; Man, N.; Mookhtiar, A.K.; Martinez, C.; Lossos, I.S.; Sun, J.; et al. PRMT5-Mediated Histone Arginine Methylation Antagonizes Transcriptional Repression by Polycomb Complex PRC2. Nucleic Acids Res. 2020, 48, 2956–2968. [Google Scholar] [CrossRef]
- Nie, M.; Wang, Y.; Guo, C.; Li, X.; Wang, Y.; Deng, Y.; Yao, B.; Gui, T.; Ma, C.; Liu, M.; et al. CARM1-Mediated Methylation of Protein Arginine Methyltransferase 5 Represses Human γ-Globin Gene Expression in Erythroleukemia Cells. J. Biol. Chem. 2018, 293, 17454–17463. [Google Scholar] [CrossRef] [PubMed]
- Radzisheuskaya, A.; Shliaha, P.V.; Grinev, V.; Lorenzini, E.; Kovalchuk, S.; Shlyueva, D.; Gorshkov, V.; Hendrickson, R.C.; Jensen, O.N.; Helin, K. PRMT5 Methylome Profiling Uncovers a Direct Link to Splicing Regulation in Acute Myeloid Leukemia. Nat. Struct. Mol. Biol. 2019, 26, 999–1012. [Google Scholar] [CrossRef] [PubMed]
- Serio, J.; Ropa, J.; Chen, W.; Mysliwski, M.; Saha, N.; Chen, L.; Wang, J.; Miao, H.; Cierpicki, T.; Grembecka, J.; et al. The PAF Complex Regulation of Prmt5 Facilitates the Progression and Maintenance of MLL Fusion Leukemia. Oncogene 2018, 37, 450–460. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Cheng, B.; Xiong, J.; Ma, D.; Liu, X.; Wang, L.; Zhang, X.; Wang, J. Protein Arginine Methyltransferase 5 Promotes the Migration of AML Cells by Regulating the Expression of Leukocyte Immunoglobulin-Like Receptor B4. BioMed Res. Int. 2021, 2021, 7329072. [Google Scholar] [CrossRef]
- Huang, X.; Yang, Y.; Zhu, D.; Zhao, Y.; Wei, M.; Li, K.; Zhu, H.; Zheng, X. PRMT5-Mediated RNF4 Methylation Promotes Therapeutic Resistance of APL Cells to As2O3 by Stabilizing Oncoprotein PML-RARα. Cell Mol. Life Sci. 2022, 79, 319. [Google Scholar] [CrossRef]
- Hyllus, D.; Stein, C.; Schnabel, K.; Schiltz, E.; Imhof, A.; Dou, Y.; Hsieh, J.; Bauer, U.-M. PRMT6-Mediated Methylation of R2 in Histone H3 Antagonizes H3 K4 Trimethylation. Gene. Dev. 2007, 21, 3369–3380. [Google Scholar] [CrossRef]
- Schneider, L.; Herkt, S.; Wang, L.; Feld, C.; Wesely, J.; Kuvardina, O.N.; Meyer, A.; Oellerich, T.; Häupl, B.; Seifried, E.; et al. PRMT6 Activates Cyclin D1 Expression in Conjunction with the Transcription Factor LEF1. Oncogenesis 2021, 10, 42. [Google Scholar] [CrossRef]
- Herglotz, J.; Kuvardina, O.N.; Kolodziej, S.; Kumar, A.; Hussong, H.; Grez, M.; Lausen, J. Histone Arginine Methylation Keeps RUNX1 Target Genes in an Intermediate State. Oncogene 2013, 32, 2565–2575. [Google Scholar] [CrossRef]
- Kolodziej, S.; Kuvardina, O.N.; Oellerich, T.; Herglotz, J.; Backert, I.; Kohrs, N.; Buscató, E.; Wittmann, S.K.; Salinas-Riester, G.; Bonig, H.; et al. PADI4 Acts as a Coactivator of Tal1 by Counteracting Repressive Histone Arginine Methylation. Nat. Commun. 2014, 5, 3995. [Google Scholar] [CrossRef]
- Herkt, S.C.; Kuvardina, O.N.; Herglotz, J.; Schneider, L.; Meyer, A.; Pommerenke, C.; Salinas-Riester, G.; Seifried, E.; Bonig, H.; Lausen, J. Protein Arginine Methyltransferase 6 Controls Erythroid Gene Expression and Differentiation of Human CD34+ Progenitor Cells. Haematologica 2018, 103, 18–29. [Google Scholar] [CrossRef]
- Kuvardina, O.N.; Herglotz, J.; Kolodziej, S.; Kohrs, N.; Herkt, S.; Wojcik, B.; Oellerich, T.; Corso, J.; Behrens, K.; Kumar, A.; et al. RUNX1 Represses the Erythroid Gene Expression Program during Megakaryocytic Differentiation. Blood 2015, 125, 3570–3579. [Google Scholar] [CrossRef] [PubMed]
- Günsel, G.G.; Conlon, T.M.; Jeridi, A.; Kim, R.; Ertüz, Z.; Lang, N.J.; Ansari, M.; Novikova, M.; Jiang, D.; Strunz, M.; et al. The Arginine Methyltransferase PRMT7 Promotes Extravasation of Monocytes Resulting in Tissue Injury in COPD. Nat. Commun. 2022, 13, 1303. [Google Scholar] [CrossRef]
- Ying, Z.; Mei, M.; Zhang, P.; Liu, C.; He, H.; Gao, F.; Bao, S. Histone Arginine Methylation by PRMT7 Controls Germinal Center Formation via Regulating Bcl6 Transcription. J. Immunol. 2015, 195, 1538–1547. [Google Scholar] [CrossRef] [PubMed]
- Oksa, L.; Mäkinen, A.; Nikkilä, A.; Hyvärinen, N.; Laukkanen, S.; Rokka, A.; Haapaniemi, P.; Seki, M.; Takita, J.; Kauko, O.; et al. Arginine Methyltransferase PRMT7 Deregulates Expression of RUNX1 Target Genes in T-Cell Acute Lymphoblastic Leukemia. Cancers 2022, 14, 2169. [Google Scholar] [CrossRef]
- Liu, C.; Zou, W.; Nie, D.; Li, S.; Duan, C.; Zhou, M.; Lai, P.; Yang, S.; Ji, S.; Li, Y.; et al. Loss of PRMT7 Reprograms Glycine Metabolism to Selectively Eradicate Leukemia Stem Cells in CML. Cell Metab. 2022, 34, 818–835.e7. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Huang, R.; Li, J.; Liao, X.; Huang, Y.; Lai, Y. Identification of Key Genes and Pathways Associated with RUNX1 Mutations in Acute Myeloid Leukemia Using Bioinformatics Analysis. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2018, 24, 7100–7108. [Google Scholar] [CrossRef]
- Xiao, P.-F.; Tao, Y.-F.; Hu, S.-Y.; Cao, L.; Lu, J.; Wang, J.; Feng, X.; Pan, J.; Chai, Y.-H. MRNA Expression Profiling of Histone Modifying Enzymes in Pediatric Acute Monoblastic Leukemia. Die Pharm. 2017, 72, 177–186. [Google Scholar] [CrossRef]
- Tao, Y.-F.; Pang, L.; Du, X.-J.; Sun, L.-C.; Hu, S.-Y.; Lu, J.; Cao, L.; Zhao, W.-L.; Feng, X.; Wang, J.; et al. Differential MRNA Expression Levels of Human Histone-Modifying Enzymes in Normal Karyotype B Cell Pediatric Acute Lymphoblastic Leukemia. Int. J. Mol. Sci. 2013, 14, 3376–3394. [Google Scholar] [CrossRef]
- Eram, M.S.; Shen, Y.; Szewczyk, M.M.; Wu, H.; Senisterra, G.; Li, F.; Butler, K.V.; Kaniskan, H.U.; Speed, B.A.; dela Seña, C.; et al. A Potent, Selective, and Cell-Active Inhibitor of Human Type I Protein Arginine Methyltransferases. ACS Chem. Biol. 2016, 11, 772–781. [Google Scholar] [CrossRef]
- Fong, J.Y.; Pignata, L.; Goy, P.-A.; Kawabata, K.C.; Lee, S.C.-W.; Koh, C.M.; Musiani, D.; Massignani, E.; Kotini, A.G.; Penson, A.; et al. Therapeutic Targeting of RNA Splicing Catalysis through Inhibition of Protein Arginine Methylation. Cancer Cell 2019, 36, 194–209.e9. [Google Scholar] [CrossRef]
- Wang, C.; Jiang, H.; Jin, J.; Xie, Y.; Chen, Z.; Zhang, H.; Lian, F.; Liu, Y.-C.; Zhang, C.; Ding, H.; et al. Development of Potent Type I Protein Arginine Methyltransferase (PRMT) Inhibitors of Leukemia Cell Proliferation. J. Med. Chem. 2017, 60, 8888–8905. [Google Scholar] [CrossRef] [PubMed]
- Bonham, K.; Hemmers, S.; Lim, Y.; Hill, D.M.; Finn, M.G.; Mowen, K.A. Effects of a Novel Arginine Methyltransferase Inhibitor on T-helper Cell Cytokine Production. FEBS J. 2010, 277, 2096–2108. [Google Scholar] [CrossRef]
- Bissinger, E.-M.; Heinke, R.; Spannhoff, A.; Eberlin, A.; Metzger, E.; Cura, V.; Hassenboehler, P.; Cavarelli, J.; Schüle, R.; Bedford, M.T.; et al. Acyl Derivatives of P-Aminosulfonamides and Dapsone as New Inhibitors of the Arginine Methyltransferase HPRMT1. Bioorg. Med. Chem. 2011, 19, 3717–3731. [Google Scholar] [CrossRef] [PubMed]
- Spannhoff, A.; Machmur, R.; Heinke, R.; Trojer, P.; Bauer, I.; Brosch, G.; Schüle, R.; Hanefeld, W.; Sippl, W.; Jung, M. A Novel Arginine Methyltransferase Inhibitor with Cellular Activity. Bioorg. Med. Chem. Lett. 2007, 17, 4150–4153. [Google Scholar] [CrossRef]
- Yan, L.; Yan, C.; Qian, K.; Su, H.; Kofsky-Wofford, S.A.; Lee, W.-C.; Zhao, X.; Ho, M.-C.; Ivanov, I.; Zheng, Y.G. Diamidine Compounds for Selective Inhibition of Protein Arginine Methyltransferase 1. J. Med. Chem. 2014, 57, 2611–2622. [Google Scholar] [CrossRef] [PubMed]
- Drew, A.E.; Moradei, O.; Jacques, S.L.; Rioux, N.; Boriack-Sjodin, A.P.; Allain, C.; Scott, M.P.; Jin, L.; Raimondi, A.; Handler, J.L.; et al. Identification of a CARM1 Inhibitor with Potent In Vitro and In Vivo Activity in Preclinical Models of Multiple Myeloma. Sci. Rep. 2017, 7, 17993. [Google Scholar] [CrossRef]
- Nakayama, K.; Szewczyk, M.M.; dela Sena, C.; Wu, H.; Dong, A.; Zeng, H.; Li, F.; de Freitas, R.F.; Eram, M.S.; Schapira, M.; et al. TP-064, a Potent and Selective Small Molecule Inhibitor of PRMT4 for Multiple Myeloma. Oncotarget 2018, 9, 18480–18493. [Google Scholar] [CrossRef]
- Veazey, K.J.; Cheng, D.; Lin, K.; Villarreal, O.D.; Gao, G.; Perez-Oquendo, M.; Van, H.T.; Stratton, S.A.; Green, M.; Xu, H.; et al. CARM1 Inhibition Reduces Histone Acetyltransferase Activity Causing Synthetic Lethality in CREBBP/EP300-Mutated Lymphomas. Leukemia 2020, 34, 3269–3285. [Google Scholar] [CrossRef]
- Chan-Penebre, E.; Kuplast, K.G.; Majer, C.R.; Boriack-Sjodin, P.A.; Wigle, T.J.; Johnston, L.D.; Rioux, N.; Munchhof, M.J.; Jin, L.; Jacques, S.L.; et al. A Selective Inhibitor of PRMT5 with in Vivo and in Vitro Potency in MCL Models. Nat. Chem. Biol. 2015, 11, 432–437. [Google Scholar] [CrossRef]
- Secker, K.-A.; Keppeler, H.; Duerr-Stoerzer, S.; Schmid, H.; Schneidawind, D.; Hentrich, T.; Schulze-Hentrich, J.M.; Mankel, B.; Fend, F.; Schneidawind, C. Inhibition of DOT1L and PRMT5 Promote Synergistic Anti-Tumor Activity in a Human MLL Leukemia Model Induced by CRISPR/Cas9. Oncogene 2019, 38, 7181–7195. [Google Scholar] [CrossRef]
- Schnormeier, A.-K.; Pommerenke, C.; Kaufmann, M.; Drexler, H.G.; Koeppel, M. Genomic Deregulation of PRMT5 Supports Growth and Stress Tolerance in Chronic Lymphocytic Leukemia. Sci. Rep. 2020, 10, 9775. [Google Scholar] [CrossRef] [PubMed]
- Duncan, K.W.; Rioux, N.; Boriack-Sjodin, P.A.; Munchhof, M.J.; Reiter, L.A.; Majer, C.R.; Jin, L.; Johnston, L.D.; Chan-Penebre, E.; Kuplast, K.G.; et al. Structure and Property Guided Design in the Identification of PRMT5 Tool Compound EPZ015666. ACS Med. Chem. Lett. 2016, 7, 162–166. [Google Scholar] [CrossRef] [PubMed]
- Gerhart, S.V.; Kellner, W.A.; Thompson, C.; Pappalardi, M.B.; Zhang, X.-P.; de Oca, R.M.; Penebre, E.; Duncan, K.; Boriack-Sjodin, A.; Le, B.; et al. Activation of the P53-MDM4 Regulatory Axis Defines the Anti-Tumour Response to PRMT5 Inhibition through Its Role in Regulating Cellular Splicing. Sci. Rep. 2018, 8, 9711. [Google Scholar] [CrossRef] [PubMed]
- Szewczyk, M.M.; Luciani, G.M.; Vu, V.; Murison, A.; Dilworth, D.; Barghout, S.H.; Lupien, M.; Arrowsmith, C.H.; Minden, M.D.; Barsyte-Lovejoy, D. PRMT5 Regulates ATF4 Transcript Splicing and Oxidative Stress Response. Redox. Biol. 2022, 51, 102282. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Zhou, J.; Xu, F.; Jin, B.; Cui, L.; Wang, Y.; Du, X.; Li, J.; Li, P.; Ren, R.; et al. Targeting Methyltransferase PRMT5 Eliminates Leukemia Stem Cells in Chronic Myelogenous Leukemia. J. Clin. Investig. 2016, 126, 3961–3980. [Google Scholar] [CrossRef]
- Bonday, Z.Q.; Cortez, G.S.; Grogan, M.J.; Antonysamy, S.; Weichert, K.; Bocchinfuso, W.P.; Li, F.; Kennedy, S.; Li, B.; Mader, M.M.; et al. LLY-283, a Potent and Selective Inhibitor of Arginine Methyltransferase 5, PRMT5, with Antitumor Activity. ACS Med. Chem. Lett. 2018, 9, 612–617. [Google Scholar] [CrossRef]
- Alinari, L.; Mahasenan, K.V.; Yan, F.; Karkhanis, V.; Chung, J.-H.; Smith, E.M.; Quinion, C.; Smith, P.L.; Kim, L.; Patton, J.T.; et al. Selective Inhibition of Protein Arginine Methyltransferase 5 Blocks Initiation and Maintenance of B-Cell Transformation. Blood 2015, 125, 2530–2543. [Google Scholar] [CrossRef]
- Pastore, F.; Bhagwat, N.; Pastore, A.; Radzisheuskaya, A.; Karzai, A.; Krishnan, A.; Li, B.; Bowman, R.L.; Xiao, W.; Viny, A.D.; et al. PRMT5 Inhibition Modulates E2F1 Methylation and Gene-Regulatory Networks Leading to Therapeutic Efficacy in JAK2V617F-Mutant MPN. Cancer Discov. 2020, 10, 1742–1757. [Google Scholar] [CrossRef]
- Zhu, K.; Shao, J.; Tao, H.; Yan, X.; Luo, C.; Zhang, H.; Duan, W. Rational Design, Synthesis and Biological Evaluation of Novel Triazole Derivatives as Potent and Selective PRMT5 Inhibitors with Antitumor Activity. J. Comput. Aid Mol. Des. 2019, 33, 775–785. [Google Scholar] [CrossRef]
- Hu, H.; Owens, E.A.; Su, H.; Yan, L.; Levitz, A.; Zhao, X.; Henary, M.; Zheng, Y.G. Exploration of Cyanine Compounds as Selective Inhibitors of Protein Arginine Methyltransferases: Synthesis and Biological Evaluation. J. Med. Chem. 2015, 58, 1228–1243. [Google Scholar] [CrossRef]
- Qian, K.; Yan, C.; Su, H.; Dang, T.; Zhou, B.; Wang, Z.; Zhao, X.; Ivanov, I.; Ho, M.-C.; Zheng, Y.G. Pharmacophore-Based Screening of Diamidine Small Molecule Inhibitors for Protein Arginine Methyltransferases. RSC Med. Chem. 2020, 12, 95–102. [Google Scholar] [CrossRef]
- Guo, Z.; Zhang, Z.; Yang, H.; Cao, D.; Xu, X.; Zheng, X.; Chen, D.; Wang, Q.; Li, Y.; Li, J.; et al. Design and Synthesis of Potent, Selective Inhibitors of Protein Arginine Methyltransferase 4 against Acute Myeloid Leukemia. J. Med. Chem. 2019, 62, 5414–5433. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Cao, C.; Jin, J.; Fan, Y.; Wang, X.; Zhang, Q.; Hu, H.; Ye, X.; Wang, L.; Ye, F. Identification of PRMT5 Inhibitors with Novel Scaffold Structures through Virtual Screening and Biological Evaluations. J. Mol. Model 2021, 28, 184. [Google Scholar] [CrossRef] [PubMed]
- Brehmer, D.; Beke, L.; Wu, T.; Millar, H.J.; Moy, C.; Sun, W.; Mannens, G.; Pande, V.; Boeckx, A.; van Heerde, E.; et al. Discovery and Pharmacological Characterization of JNJ-64619178, a Novel Small Molecule Inhibitor of PRMT5 with Potent Anti-Tumor Activity. Mol. Cancer Ther. 2021, 20, 2317–2328. [Google Scholar] [CrossRef] [PubMed]
- Zhu, K.; Tao, H.; Song, J.-L.; Jin, L.; Zhang, Y.; Liu, J.; Chen, Z.; Jiang, C.-S.; Luo, C.; Zhang, H. Identification of 5-Benzylidene-2-Phenylthiazolones as Potent PRMT5 Inhibitors by Virtual Screening, Structural Optimization and Biological Evaluations. Bioorg. Chem. 2018, 81, 289–298. [Google Scholar] [CrossRef] [PubMed]
- van Haren, M.J.; Marechal, N.; Troffer-Charlier, N.; Cianciulli, A.; Sbardella, G.; Cavarelli, J.; Martin, N.I. Transition State Mimics Are Valuable Mechanistic Probes for Structural Studies with the Arginine Methyltransferase CARM1. Proc. Natl. Acad. Sci. USA 2017, 114, 3625–3630. [Google Scholar] [CrossRef] [PubMed]
- Siu, L.L.; Rasco, D.W.; Vinay, S.P.; Romano, P.M.; Menis, J.; Opdam, F.L.; Heinhuis, K.M.; Egger, J.L.; Gorman, S.A.; Parasrampuria, R.; et al. 438OMETEOR-1: A Phase I Study of GSK3326595, a First-in-Class Protein Arginine Methyltransferase 5 (PRMT5) Inhibitor, in Advanced Solid Tumours. Ann. Oncol. 2019, 30, v159. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| PRMT(s) Inhibited | Drug (Alias) | IC50 | Disease | Experimental Models | References |
|---|---|---|---|---|---|
| Type I PRMTs | MS023 [99] | 30 nM (PRMT1) 119 nM (PRMT3) 83 nM (CARM1) 4 nM (PRMT6) 5 nM (PRMT8) | ALL | Cell lines; primary specimens; PDX | [36] |
| AML | Cell lines; primary specimens; PDX | [48,100] | |||
| Compound 28d [101] | 1.1 nM (PRMT1) 22 nM (PRMT3) 63 nM (CARM1) 1.2 nM (PRMT6) 1.1 nM (PRMT8) | MLL | Cell lines | [101] | |
| PRMT1 | AMI-408 (compound 4) [102] | ND | AML | Cell lines; mice xenografts | [51] |
| TC-E 5003(compound 2e) [103] | 1 µM | ALL | Cell lines | [36] | |
| AML | Cell lines | [48] | |||
| RM-65 [104] | 55 µM | AML | Cell lines | [52] | |
| DB75 (Furamidine) [105] | 9.4 µM | AMKL | Cell lines | [35] | |
| CARM1 | EPZ025654 (GSK35336023) [106] | 3 nM | AML | Cell lines; primary specimens; PDX | [60] |
| TP-064 [107] | <10 nM | NHL | Cell lines | [108] | |
| MM | Cell lines | [107] | |||
| EZM2302 (GSK3359088) [106] | 6 nM | MM | Cell lines; xenografted mice gavage | [106] | |
| AML | Transplanted mice gavage | [60] | |||
| NHL | Transplanted mice gavage | [108] | |||
| PRMT5 | EPZ015666 (GSK3235025) [109] | 22 nM | AML | Cell lines; primary specimens; transplanted mice gavage | [82,83,100,110] |
| CLL | Cell lines | [111] | |||
| NHL | Cell lines; PDX | [78] | |||
| MM | Cell lines; primary specimens; xenografted mice gavage | [69] | |||
| EPZ015866 (GSK3203591) [112] | 4 nM | NHL | Cell lines; primary specimens | [79,113] | |
| MM | Cell lines | [70] | |||
| AML | Cell lines | [100,114] | |||
| EPZ015938 (GSK3326595) [113] | 6.2 nM | NHL | Cell lines; PDX; transplanted or xenografted mice gavage | [78,113] | |
| GSK3186000A (compound 14) [112] | 11 nM | AML | Cell lines | [64,80] | |
| PJ-68 [115] | 517 nM | CML | Cell lines; primary specimens; intraperitoneally injected transplanted mice | [115] | |
| HLCL-61 [18] | 7.21–21.46 µM | AML | Cell lines; primary specimens | [18] | |
| LLY-283 [116] | 22 nM | AML | Cell lines | [114] | |
| CMP-5 [117] | ND | NHL | Cell lines; mouse primary specimens | [76] | |
| C220 [118] | ND | MPN | Cell lines; transplanted mice gavage | [118] | |
| PRMT7 | JS1310 [95] | 5 µM | CML | Leukemic mice; primary specimens; PDX | [95] |
| Trial Number | Official Title of the Study | Number of Patients | Drug (Alias) | PRMT Inhibited | HM Included |
|---|---|---|---|---|---|
| NCT03666988 | A Phase I, open-label, dose-escalation study to investigate the safety, pharmacokinetics, pharmacodynamics, and clinical activity of GSK3368715 in patients with solid tumors and DLBCL. | 31 | GSK3368715 (EPZ019997) | Type I PRMTs | DLBCL |
| NCT03614728 | A Phase I/II Study to investigate the safety and clinical activity of GSK3326595 and other agents in patients with MDS and AML. | 30 | GSK3326595 (Pemrametostat) | PRMT5 | MDS; CMML; AML |
| NCT02783300 | A Phase I, open-label, dose-escalation study to investigate the safety, pharmacokinetics, pharmacodynamics, and clinical activity of GSK3326595 in patients with solid tumors and NHL. | 322 | GSK3326595 (Pemrametostat) | PRMT5 | NHL |
| NCT03573310 | A Phase I, first in-human, open-label study of the safety, pharmacokinetics, and pharmacodynamics of JNJ-64619178, an inhibitor of PRMT5 in patients with advanced cancers. | 114 | JNJ-64619178 (Onametostat) | PRMT5 | B NHL; lower risk MDS |
| NCT03886831 | A Phase I, open-label, multicenter, dose-escalation, dose-expansion study of PRT543 in patients with advanced solid tumors and hematologic malignancies. | 227 | PRT543 | PRMT5 | DLBCL; myelodysplasia; myelofibrosis; MCL; AML; CML |
| NCT04089449 | A Phase I, open-label, multicenter, dose-escalation and -expansion study of PRT811 in subjects with advanced solid tumors, CNS lymphoma, and recurrent high-grade gliomas. | 145 | PRT811 | PRMT5 | CNS lymphoma |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sauter, C.; Simonet, J.; Guidez, F.; Dumétier, B.; Pernon, B.; Callanan, M.; Bastie, J.-N.; Aucagne, R.; Delva, L. Protein Arginine Methyltransferases as Therapeutic Targets in Hematological Malignancies. Cancers 2022, 14, 5443. https://doi.org/10.3390/cancers14215443
Sauter C, Simonet J, Guidez F, Dumétier B, Pernon B, Callanan M, Bastie J-N, Aucagne R, Delva L. Protein Arginine Methyltransferases as Therapeutic Targets in Hematological Malignancies. Cancers. 2022; 14(21):5443. https://doi.org/10.3390/cancers14215443
Chicago/Turabian StyleSauter, Camille, John Simonet, Fabien Guidez, Baptiste Dumétier, Baptiste Pernon, Mary Callanan, Jean-Noël Bastie, Romain Aucagne, and Laurent Delva. 2022. "Protein Arginine Methyltransferases as Therapeutic Targets in Hematological Malignancies" Cancers 14, no. 21: 5443. https://doi.org/10.3390/cancers14215443
APA StyleSauter, C., Simonet, J., Guidez, F., Dumétier, B., Pernon, B., Callanan, M., Bastie, J.-N., Aucagne, R., & Delva, L. (2022). Protein Arginine Methyltransferases as Therapeutic Targets in Hematological Malignancies. Cancers, 14(21), 5443. https://doi.org/10.3390/cancers14215443