Simple Summary
Metabolic alterations (i.e., high glucose levels) enhance cancer cell aggressiveness by acting on tumor and on its environment. Mammary adipose tissue-derived mesenchymal stromal/stem cells (MAT-MSCs) stand in close proximity of breast cancer (BC) cells but have a still unclear role. We investigated whether changes in glucose concentration may impact on the interaction between human MAT-MSCs and BC cells. We provided evidence that in presence of cancer cells and high glucose levels, MAT-MSCs display pro-tumorigenic functions, while cancer cells become more aggressive. These results highlight that a metabolic control helps to reduce cancer outgrowth, at least in part preserving the functions of the adipose microenvironment.
Abstract
Adiposity and diabetes affect breast cancer (BC) progression. We addressed whether glucose may affect the interaction between mammary adipose tissue-derived mesenchymal stromal/stem cells (MAT-MSCs) and BC cells. Two-dimensional co-cultures and spheroids were established in 25 mM or 5.5 mM glucose (High Glucose-HG or Low Glucose-LG) by using MAT-MSCs and MCF7 or MDA-MB231 BC cells. Gene expression was measured by qPCR, while protein levels were measured by cytofluorimetry and ELISA. CD44high/CD24low BC stem-like sub-population was quantified by cytofluorimetry. An in vivo zebrafish model was assessed by injecting spheroid-derived labeled cells. MAT-MSCs co-cultured with BC cells showed an inflammatory/senescent phenotype with increased abundance of IL-6, IL-8, VEGF and p16INK4a, accompanied by altered levels of CDKN2A and LMNB1. BC cells reduced multipotency and increased fibrotic features modulating OCT4, SOX2, NANOG, αSMA and FAP in MAT-MSCs. Of note, these co-culture-mediated changes in MAT-MSCs were partially reverted in LG. Only in HG, MAT-MSCs increased CD44high/CD24low MCF7 sub-population and promoted their ability to form mammospheres. Injection in zebrafish embryos of HG spheroid-derived MCF7 and MAT-MSCs was followed by a significant cellular migration and caudal dissemination. Thus, MAT-MSCs enhance the aggressiveness of BC cells in a HG environment.
1. Introduction
Diabetes and cancer are serious diseases, whose prevalence is rapidly increasing worldwide. It has been estimated that the number of diabetic patients will reach 578 million in 2035 and 700 million in 2045 [1] and that the number of global cancer patients will be 22 million in 2032 [2]. Epidemiological and biological studies have associated type 2 diabetes (T2D) and cancer [3]. In particular, the risk of developing breast cancer (BC) is 20–28% higher in diabetic than in healthy patients [4]. Notably, BC prognosis and overall/disease-free survival are worsened by diabetes or impaired glucose tolerance [5,6].
Cancer cells establish bidirectional communication with surrounding stromal cells to reprogram the microenvironment and obtain support for their own growth and dissemination [7]. Importantly, they accumulate metabolic alterations and take advantage of selected metabolites (including glucose), also affecting the fate of normal cells within the tumor microenvironment [8].
The breast microenvironment is composed of numerous stromal cell types, including endothelial and immune cells, fibroblasts and adipocytes. Stromal cells enhance tumor progression through mutual and dynamic communication with cancer cells [9,10,11]. For instance, cancer cells can transform stromal fibroblasts into myofibroblastic phenotypes called cancer-associated fibroblasts (CAFs) and metastasis associated fibroblasts (MAFs). CAFs secrete growth factors and cytokines to sustain cancer progression; MAFs convoy cancer cells for metastasis [12]. Adipocytes contribute to sustaining tumor phenotypes by either acting as energy reservoirs for neighboring embedded cancer cells or through secretion of signaling molecules and vesicles containing proteins, lipids and nucleic acids [9,11]. Notably, adipocytes mediate, at least in part, the impact of metabolic alterations on the BC phenotype. Indeed, we have demonstrated that glucose promotes adipocyte release of IGF1, CCL5 and IL-8 to support BC cell proliferation, invasiveness and drug-resistance [13,14,15].
Nevertheless, adipose tissue (AT) is an abundant source of mesenchymal stromal/stem cells (MSCs) showing self-renewal activity, multi-lineage differentiation potential and secreting a plethora of factors with immunomodulatory, angiogenic, neurotrophic, pro- and anti-inflammatory functions [16,17,18]. MSCs are largely modified by external stimuli. For instance, cytokines secreted by BC cells seem to play a role in the transformation of bone marrow-MSCs in CAF [19]. Moreover, exogenous factors, such as metabolic and stress-related insults may trigger a senescent phenotype in MSCs. Senescent cells display changes in gene expression and a characteristic secretory pattern called the senescence-associated secretory phenotype (SASP), which includes multiple cytokines, growth factors, proteases and matrix metalloproteinases. In most studies, the SASP was reported to sustain cancer cells [20,21]. Thus, by switching on multiple, but still unclear mechanisms, MSCs may favor tumor growth and progression. However, different studies indicate MSCs as vectors for consolidated and innovative therapies [9,22,23,24,25].
Here, we have attempted to clarify whether glucose may interfere with the dialogue between mammary adipose-derived MSCs (MAT-MSCs) and BC cells.
2. Materials and Methods
2.1. Isolation of MSCs from MAT
MAT biopsies were obtained from healthy women (N = 20) undergoing surgical mammary reduction, free of neoplastic, metabolic or endocrine diseases. Informed consents were collected before surgical procedure. Protocol was approved by the ethical committee of the University of Naples “Federico II” (prot. n. 138/16). Biopsies were minced and digested by collagenase solution (cat. C2139-1G, 1 mg/mL-Sigma-Aldrich, St. Louis, MO, USA) to isolate MAT-MSCs [13,16,26].
2.2. Cell Cultures
MAT-MSCs were cultured in DMEM-F12 (1:1). MCF7 (ER+, PR+, HER2−) and MDA-MB231 (ER−, PR−, HER2−) human BC cell lines were cultured in DMEM 25mM glucose. Both media were supplemented with 10% FBS, 2 mM glutamine, 100 unit/mL penicillin, and 100 unit/mL streptomycin. As indicated below, co-culture experiments cells were exposed to 25 mM glucose (standard/high glucose; HG), resembling hyperglycemia in humans, or to 5.5 mM glucose (Low Glucose; LG), resembling normoglycemia. Cultures were maintained in a humidified atmosphere of 95% air and 5% CO2 at 37 °C. Media, sera and antibiotics were obtained from Lonza (Basel, Switzerland). Cells were routinely screened for mycoplasma contamination.
2.3. Adipogenic and Osteogenic Differentiation of MAT-MSCs
Adipocyte differentiation of MAT-MSCs was obtained by the alternation (every three days, two times) of an adipocyte differentiation Induction Mix (850 nM insulin, 10 μM dexamethasone, 0.5 mM 3-IsoButyl-1-MethylXanthine, 33 μM biotin, 17 μM pantothenate and 1 μM rosiglitazone) and an adipocyte differentiation maintaining mix (850 nM insulin and 1 μM rosiglitazone). Then, the cells were stimulated (every two days, two times) with 1 μM rosiglitazone. All the process was carried out in complete culture medium. Adipocyte differentiation was reached within 17 days and lipid accumulation determined by oil red O staining [27]. Osteogenic differentiation of MAT-MSCs was reached within 21 days and mineralization foci detected by alizarin red S [16]. Images of stained monolayers were taken using the Olympus DP20 microscope digital camera system (Olympus Corporation, Tokyo, Japan). All chemicals were obtained from Sigma-Aldrich (St. Louis, MO, USA).
2.4. 2D-Cultures
MAT-MSCs (2.5 × 104 cells) were seeded in the bottom chamber of a transwell-6 culture system (cat. 353090, 0.4 μm pore size, Costar, MA, USA) while MCF7 or MDA-MB231 were plated (3.5 × 104 cells) in the upper chamber. Cells were co-cultured in the presence of HGor LG. In parallel, BC cells and MAT-MSCs were monocultured in HG or LG medium. After 72 h, conditioned media (CM) were collected, and cells were harvested for RNA extraction or cytofluorimetric analysis.
2.5. 3D-Cultures
Mammosphere-forming assay: MCF7 cells were plated in ultra-low attachment 96-wells (cat. 7007, Corning, NY, USA) with or without MAT-MSCs (ratio 4:1) in HG or LG medium supplemented with 5% MammoCultTM proliferation supplement (cat. 05622, STEMCELL Technologies, Vancouver, BC, Canada). After 10 days, mammosphere number was quantified (number of formed spheres/number of wells containing cells × 100) [28]. In parallel, mammosphere diameter was measured by a software associated to the Olympus DP20 microscope digital camera system.
Spheroid formation: MCF7 (4 × 105) were plated in ultra-low attachment 100 mm dish (cat. 3262, Corning, NY, USA) with or without MAT-MSCs (1 × 105) in HG or LG medium supplemented with 5% FBS. After 72 h, spheroids were mechanically disaggregated to obtain cells for cytofluorimetric analysis or zebrafish injection.
2.6. Cytofluorimetric Analysis
Immunophenotypic characterization of MAT-MSCs: MAT-MSCs were incubated with PE-anti-CD73 (Cat. 550257), FITC-anti-CD90 (Cat. 555595) and APC-Cy7-anti-CD45 antibodies (Cat. 641399) as well as dye/isotype-matched antibodies (all from BD Biosciences, San Diego, CA, USA), following manufacturer’s instructions.
Quantification of CD44high/CD24low BC cell sub-population: MCF7 were incubated with APC-anti-CD44 (cat. 130-113-338) and PE-anti-CD24 (cat. 130-112-656) as well as dye/isotype control matched antibodies (cat. 130-112-656, all from Miltenyi Biotec, Bergisch Gladbach, Germany), following manufacturer’s instructions.
Protein levels in 3D-cultures: Cell membranes were permeabilizated by using the Cytofix/Cytoperm kit (cat. 554714, BD Biosciences) before incubation (4C, 30 min) with specific or isotype control antibodies. Positivity to cytokeratin (FITC-anti-cytokeratin; cat. 130-112-743, Miltenyi Biotec) was used to discriminate MCF7 from MAT-MSCs. PE-anti-OCT4 was from Cell Signaling (cat. 2750S, Danvers, MA, USA), anti-Nanog was from Santa Cruz Biotechnology (cat. sc-374103, Dallas, TX, USA), secondary anti-mouse PE antibody used to conjugate Nanog antibody was from R&D System (cat. F0102B, Minneapolis, MN, USA), anti-αSMA was from Dako (cat. M0851, Carpinteria, CA, USA), secondary anti-mouse PE antibody used to conjugate anti-αSMA antibody was from Miltenyi Biotec (cat. 130-095-908), APC-anti-FAP was from R&D System (cat. FAB3715A), APC-anti-p16 was from Miltenyi Biotec (cat. 130-116-138). Samples were processed using a BD LSR Fortessa or using a FACSCalibur (BD Biosciences) and analyzed by using BD FACS Diva software or Cell Quest software. 104 events for each sample were acquired in all analyses.
2.7. Multiplex ELISA Assay
CM from co-cultured and monocultured MAT-MSCs, MCF7 and MDA-MB231 were screened for concentrations of PDGF, IL-1ra, IL-1b, IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-10, IL-12, IL-13, IL-15, IL-17, eotaxin, FGF, G-CSF, GM-CSF, IFN-γ, MCP1, MIP-1α, MIP-1β, RANTES/CCL5, TNFα and VEGF using the bio-plex multiplex human cytokine and growth factor kits (cat. M500KCAF0Y, Bio-Rad, Hercules, CA, USA) according to the manufacturer’s protocol [26].
2.8. RNA Isolation and Analysis
Total RNA was isolated using TRIzol solution (cat. 15596026, Life Technologies, Carlsbad, CA, USA) according to the manufacturer’s instructions. RNA samples were quantified by measuring the absorbance at 260 nm and 280 nm (NanoDrop spectrophotometer, Life Technologies). RNA integrity was analyzed by using the digital electrophoresis system Experion with the “RNA StdSens Kit” (Biorad), following the manufacturer’s instructions. Run and result analyses were performed using the Experion software. RNA quality indicator (RQI) value ≥ 9 was considered good for the further analysis.
2.9. RT-PCR
RNA samples were reverse-transcribed using SuperScript III Reverse Transcriptase with oligo dT primers (cat. 18080-044, Life Technologies) according to the manufacturer’s instructions. To check the amplifiable template RNA/cDNA, RT-PCR amplification of housekeeping genes was performed. Amplification reactions were set up using AmpliTaq Gold (cat. N8080247, Life Technologies) and specific primer pairs, designed by Oligo 4.0 (Supplementary Table S1).
2.10. Quantitative Real-Time RT-PCR (qPCR)
qPCR was performed using an iTaq Universal SYBR Green Supermix (cat. 1725124, Biorad), according to the manufacturer’s instructions for the CFX Connect Real-Time system (Biorad). Relative quantification of gene expression was measured by using 2−ΔΔCt method. Expression levels were normalized for the reference sample using peptidylprolyl isomerase A (PPIA) as housekeeping gene.
2.11. In Vivo Zebrafish Model
Cell culture and labeling: MCF7 and MAT-MSCs from 3 different donors were labeled with red cell trackers CM-DiI (cat. C7000, Thermo Fisher Scientific, Waltham, MA, USA) and BioTraker 400 Blu (cat. SCT109, Millipore, MA, USA), respectively—according to manufacturer’s instructions—before spheroid formation. After 72 h, spheroids were trypsinized, washed and resuspended in PBS/EDTA for zebrafish xenotransplantation.
Zebrafish husbandry and xenotransplantation: Animal experiments were in accordance with the European Council Directive 2010/63/EU and approved by the Biogem s.c.ar.l. internal ethics committee. Animal care was in accordance with institution guidelines. Tg(fli1:EGFP) zebrafish line, with green fluorescent vessels was raised, maintained and paired under standard conditions. Zebrafish eggs were obtained from natural spawning and maintained at 28 °C for 48 h in E3 medium (5 mM NaCl, 0.17 mM KCl, 0.33 mM CaCl2, 0.33 mM MgSO4). Two days post-fertilization (dpf), embryos were dechorionated and anesthetized with 0.04% of tricaine (Sigma Aldrich, St. Louis, MO, USA) before cell microinjection. Approximately 100–200 cells/embryo were injected in the perivitelline space of each embryo using a pneumatic PicoPump PV830 injector (World Precision Instruments, Sarasota, FL, USA) equipped with an injection borosilicate glass needle (Sutter Instruments, Novato, CA, USA). Following transplantation (0 h post injection), larvae with correct engraftment in the yolk sac were selected under Leica M205 FA fluorescence stereo microscope (Leica, Mycrosystems, Wetzlar, Germany) for further analysis and kept at 34 °C for 72 h. Embryos injected with same volume of PBS/EDTA were defined as control embryos.
Imaging: Zebrafish larvae were anesthetized and evaluated at 0 and 72 h post-injection by fluorescence stereo microscope. Different filters were selected for fluorescence imaging and captured with a Leica DFC450 C camera. Images of embryos at different stages of each experimental group were analyzed with ImageJ software (National Institutes of Health, Rockville, MD, USA).
2.12. Statistical Analysis
Statistical analyses were performed using GraphPad Prism 7.0 software (GraphPad Software Inc., La Jolla, CA, USA). Kruskal Wallis and Friedman tests followed by Dunn’s correction were applied for multiple comparisons; Wilcoxon and Mann–Whitney U tests were used for pairwise comparisons, as indicated in figure/table legends. Percentages were compared by chi-square with Fisher’s exact test. p-value < 0.05 was considered statistically significant. Sample sizes were determined based on the means and variations of previous pilot experiments. Hence, no statistical power analysis was used. Replicates for all the experiments were from different samples isolated from different human specimens, as indicated in figure legends.
3. Results
3.1. Characterization of MAT-MSCs
MSCs were isolated from MAT biopsies (N = 20) of women undergoing surgical mammary reduction (Figure 1a). Women were aged 37.4 ± 13.7, with normal glucose levels and a BMI of 27.7 ± 3.7 (Supplementary Table S2). Based on anthropometric and biochemical data, no evidence of donor sub-groups was noted (Supplementary Figure S1). MAT-MSCs expressed mesenchymal progenitor cell surface antigens CD90 (99.9% ± 0.07) and CD73 (96.5% ± 2.84) while not expressing the hematopoietic marker CD45 (1.6% ± 0.07) (Figure 1b). Moreover, upon specific stimuli (see Section 2), MAT-MSCs differentiated into both adipocytes (Figure 1c) and osteocyte-like cells (Figure 1d).
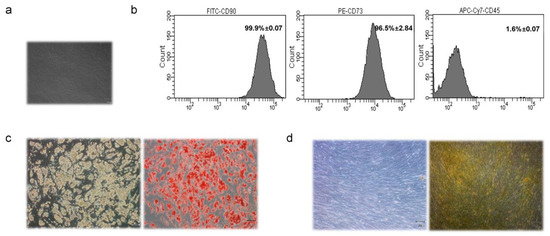
Figure 1.
Characterization of MSCs isolated from Mammary Adipose Tissue (MAT-MSCs). (a) Representative microscopic image (magnification 10×) of MAT-MSCs. (b) Representative dot plots from FACS analysis of MAT-MSCs stained for FITC-anti-CD90, PE-anti-CD73, APC-Cy7-anti-CD45 antigens. (c) Representative microscopic images (magnification 10×) of MAT-MSCs differentiated towards the adipogenic lineage, stained (right) and not (left) with oil red O for lipid accumulation detection. (d) Representative microscopic images (magnification 10×) of MAT-MSCs differentiated towards osteogenic lineage, stained (right) and not (left) with alizarin red S for calcium accumulation detection.
3.2. Profiling of Cytokines, Chemokines, and Growth Factors in MAT-MSCs and BC Cell Co-Cultures
To study the bi-directional communication between MAT-MSCs and BC cells, 2D-cultures were established. Conditioned media (CM) from co-cultured and monocultured MAT-MSCs, MCF7 and MDA-MB231 were collected to define the release of cytokines/chemokines and growth factors. Detectable levels of several molecules were observed in both monocultures and co-cultures (Table 1). The different MAT-MSC isolates displayed a similar secretory profile, without evidence of cell clusters (Supplementary Figure S2a). Moreover, no significant correlations were found between released factors and age, BMI, glucose, cholesterol and triglyceride blood levels of donors (Supplementary Figure S2b). MAT-MSCs secreted a high amount of IL-6, which was the most largely detected cytokine in co-cultures of MAT-MSCs with both MCF7 (421.07 ± 146.91 pg/mL) and MDA-MB231 cells (631.42 ± 286 pg/mL). High levels of IL-6 in co-cultures may derive from MAT-MSCs, since monocultured BC cells released much lower amounts of the cytokine (Table 1). Similar to IL-6, co-cultures of MAT-MSCs with both MCF7 and MDA-MB231 released high amounts of VEGF (173.44 ± 44.43 pg/mL and 115.12 ± 22.21 pg/mL, respectively). Finally, MSCs + MDA-MB231 co-cultures displayed high concentrations of IL-8 (47 ± 17.51 pg/mL), also largely secreted by MDA-MB231 monocultures (20.45 ± 8.71 pg/mL), while barely detected in CM from MAT-MSCs, MCF7 and MSCs + MCF7.

Table 1.
Cytokine, chemokine and growth factor concentration (pg/mL) in conditioned media (CM) from mono-cultured and co-cultured cells. MCF7 or MDA-MB231 were co-cultured—and not—with MAT-MSCs in standard/high glucose (25 mM glucose; MSC). After 72 h, CM from monocultured and co-cultured MAT-MSCs, MCF7 and MDA-MB231 were collected and analyzed by using the bio-plex multiplex human cytokine and growth factor kits (see Methods). Data were reported as mean ± SD of 5 independent experiments. For multiple comparisons, Kruskall–Wallis test followed by Dunn’s correction was applied. For pairwise comparisons (CM MAT-MSCs + MCF7 vs. CM MAT-MSCs + MDA-MB231) Mann–Whitney test was assessed. * denotes statistically significant values vs. MAT-MSCs (* adjp < 0.05; ** adjp < 0.01); § denotes statistically significant values compared with MCF7 (§ adjp < 0.05; §§ adjp < 0.01); # denotes statistically significant values vs. MDA-MB231 (# adjp < 0.05; ## adjp < 0.01); ç denotes statistically significant values compared with MAT-MSCs + MCF7 (ç adjp < 0.05; çç adjp < 0.01).
Thus, co-culture media, compared to MAT-MSCs and/or BC cell monoculture media had higher levels of IL-6, VEGF and IL-8; proteins involved in SASP; CAF functions and cancer aggressiveness [20,29,30,31].
3.3. Evaluating the Role of Glucose on BC Cell-Induced Phenotype of MAT-MSCs
Next, we analyzed mRNA levels of IL6, IL8, and VEGF together with established markers of cell senescence CDKN2A and LMNB1, in MAT-MSCs. CDKN2A encodes p16INK4a, a protein elevated in cultured senescent cells, while LMNB1 encodes lamin B1, whose reduction is considered a senescence-associated biomarker [32,33]. As shown in Figure 2a, we found a 3.2- and 2.3-fold increase in IL6 and VEGF—while not of IL8—mRNA levels in MAT-MSCs co-cultured with MCF7 (Figure 2a; adjp < 0.05) compared to control cells, represented by MAT-MSCs monocultures. Furthermore, we observed that co-cultures with MDA-MB231 induced IL6, IL8 and VEGF mRNAs in MAT-MSCs by 5.9-, 36.3- and 2.4-fold, respectively (Figure 2a; adjp < 0.001). Of note, co-cultures with MCF7, while not with MDA-MB231, modified the expression levels of CDKN2A (3-fold increase) and LMNB1 (about 50% reduction) in MAT-MSCs (Figure 2a; adjp < 0.05). Then, we measured mRNA levels of ACTA2, encoding α-SMA, a marker of myofibroblast trans-differentiation, and observed that both MCF7 and MDA-MB231 promoted its expression in MAT-MSCs by 2- and 3-fold, respectively (Figure 2a; adjp < 0.01). No correlation between gene expression profiles and antropometric and biochemical data of the donors was found in MAT-MSCs.
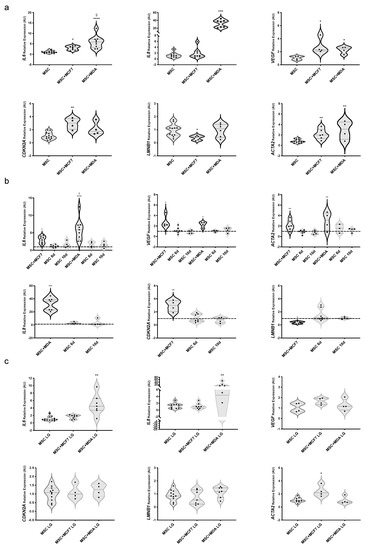
Figure 2.
Effect of glucose on MAT-MSCs phenotype in 2D-cultures with BC cells. MAT-MSCs were co-cultured—and not—with MCF7 or MDA-MB231 for 72 h in (a,b) standard/high glucose (25 mM glucose; MSC) or (c) low glucose concentration (5.5 mM; MSC LG). After 72 h, BC cells were removed while MAT-MSCs were grown for another 3 (MSC 6d) or 7 days (MSC 10d). mRNA expression levels of senescence/fibrosis markers (IL6, IL8, VEGF, CDKN2A, LMNB1, ACTA2) were determined by qPCR (see Section 2 and Supplementary Table S1. Data were normalized on the peptidyl-prolyl cis-trans isomerase A (PPIA) gene as per internal standard. Results were represented as: violin plot of 7–9 (a,c) or 3–5 (b) independent triplicate experiments showing mRNA levels of IL6, IL8, VEGF, CDKNA2, LMNB1 and ACTA2 in MAT-MSCs co-cultured with BC cells as relative expression (2−ΔΔCt) compared to that in monocultured MAT-MSCs (control cells). In (b) monocultured cells (control cells) are represented as a dotted line. Data were analyzed using non-parametric Kruskall–Wallis test followed by Dunn’s correction for multiple comparisons. For pairwise comparisons (MSC + MCF7 vs. MSC + MDA-MB231), Mann–Whitney test was assessed. * denotes statistically significant values compared with monocultured MAT-MSCs (* adjp < 0.05; ** adjp < 0.01; *** adjp < 0.001, **** adjp < 0.0001). ç denotes statistically significant values compared with MSC + MCF7 (ç adjp < 0.05).
Interestingly, we found that BC modulation of senescence and trans-differentiation markers in MAT-MSCs was not maintained for 10 days upon the end of co-culture (i.e., the removal of BC cells) (Figure 2b).
Next, we investigated the impact of glucose lowering (25 mM to 5.5 mM; HG → LG) onto the MAT-MSCs phenotype acquired upon co-culture with BC cells. We previously demonstrated that glucose did not modify the MAT-MSC proliferation rate, while increasing BC cell proliferation [13,16]. Here, we found that glucose lowering per sè did not modulate IL8, VEGF, CDKN2A and LMNB1 mRNA levels while it promoted IL6 and ACTA2 expression (Supplementary Figure S3; adjp < 0.01). Co-culturing MAT-MSCs with MCF7 in LG still induced ACTA2 mRNA expression (Figure 2c; adjp < 0.05) while—at variance with HG (Figure 2a)—it did not affect IL6, VEGF, CDKN2A and LMNB1 mRNA levels (Figure 2c).
In parallel, in LG, MDA-MB231 still upregulated IL6 and IL8 but lost the ability to increase mRNA levels of VEGF and ACTA2 in MAT-MSCs (Figure 2c). Thus, glucose lowering reverted, at least in part, the acquisition of senescent/fibrotic phenotype induced by BC cells in MAT-MSCs.
In addition, in these co-culture systems, neither glucose nor MAT-MSCs modulated expression levels of inflammatory and senescence-associated markers in MCF7 (Supplementary Figure S4) and in MDA-MB231 (Supplementary Figure S5).
Notably, we also observed that co-cultures with BC cells significantly reduced mRNA levels of multipotency markers OCT4 (about 30% reduction upon co-culture with MCF7), SOX2 (about 80% reduction upon co-culture with both MCF7 and MDA-MB231) and NANOG (about 80% reduction upon co-culture with MDA-MB231) in MAT-MSCs (Figure 3a; adjp < 0.05). Glucose lowering did not modify the expression of multipotency-related genes in MAT-MSCs (Supplementary Figure S3b) while totally restoring cancer cell action on their expression levels (Figure 3b).
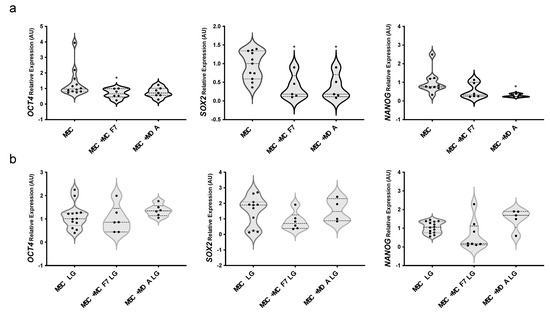
Figure 3.
Effect of glucose on MAT-MSCs multipotency in 2D cultures with BC cells. MAT-MSCs were co-cultured—and not—with MCF7 or MDA-MB231 BC cells in (a) standard/high glucose (25 mM glucose; MSC) or (b) low glucose concentration (5.5 mM; MSC LG). After for 72 h, mRNA expression levels of multipotency markers (OCT4, SOX2, NANOG) were determined by qPCR (see Methods and Supplementary Table S1). Data were normalized on PPIA gene as internal standard. Results were represented as violin plot of 7–9 independent triplicate experiments showing mRNA levels of OCT4, SOX2 and NANOG in MAT-MSCs co-cultured with BC cells as relative expression (2−ΔΔCt) compared to that in mono-cultured MAT-MSCs. Data were analyzed using non-parametric Kruskall–Wallis test followed by Dunn’s correction for multiple comparisons. * denotes statistically significant values compared with MSC (* adjp < 0.05).
3.4. Evaluating the Role of Glucose on MAT-MSC-Induced Phenotype of BC Cells
To investigate the effect of MAT-MSCs and the possible role of glucose to the BC cell stem-like phenotype, mRNA levels of OCT4, SOX2, and NANOG were measured in BC cells upon co-culture with MAT-MSCs in HG or LG. We found a significant increase of OCT4 mRNA levels in MCF7 co-cultured with MAT-MSCs in HG while not in LG (Figure 4a; pval < 0.05) as compared to the relative monocultures. SOX2 and NANOG mRNA levels remained unaltered in co-cultured MCF7 (Figure 4a). Notably, glucose did not directly affect the expression of these genes in MCF7 cells (Supplementary Figure S6). Moreover, we observed that stemness genes did not change in MDA-MB231 upon co-culture with MAT-MSCs neither in LG nor in HG (Figure 4b).
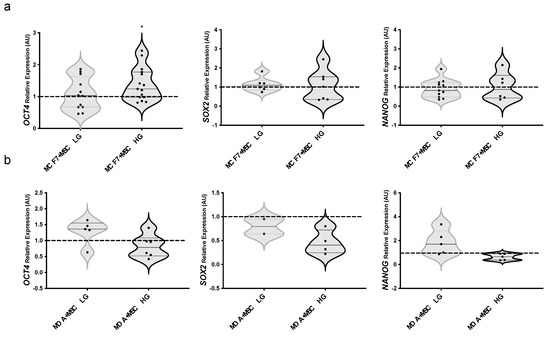
Figure 4.
Effect of glucose on BC cells stemness in 2D cultures with MAT-MSCs. (a) MCF7 or (b) MDA-MB231 were co-cultured-and not—with MAT-MSCs in low glucose (5.5 mM; LG) or standard/high glucose (25 mM; HG) medium. After 72 h, mRNA expression levels of stemness genes (OCT4, SOX2, NANOG) were determined by qPCR (see Methods and Supplementary Table S1). Data were normalized on PPIA gene as internal standard. Results were represented as violin plot of 7–15 independent triplicate experiments showing mRNA levels of OCT4, SOX2 and NANOG in (a) MCF7 + MSC LG/HG and (b) MDA-MB231 + MSC LG/HG as relative expression (2−ΔΔCt) compared to that in monocultured cells (MCF7/MDA-MB231 LG/HG; dotted line). Data were analyzed using the non-parametric Wilcoxon test for pairwise comparisons. * denotes statistically significant values (* pval < 0.05).
Next, we quantified CD44high/CD24low stem-like BC sub-population upon co-culture with MAT-MSCs. As shown in Figure 5, co-culturing MCF7 with MAT-MSCs in HG, while not in LG, determined a 15-fold increase of CD44high/CD24low sub-population (Figure 5; adjp < 0.01). Thus, MAT-MSCs induced a stem-like phenotype in MCF7.
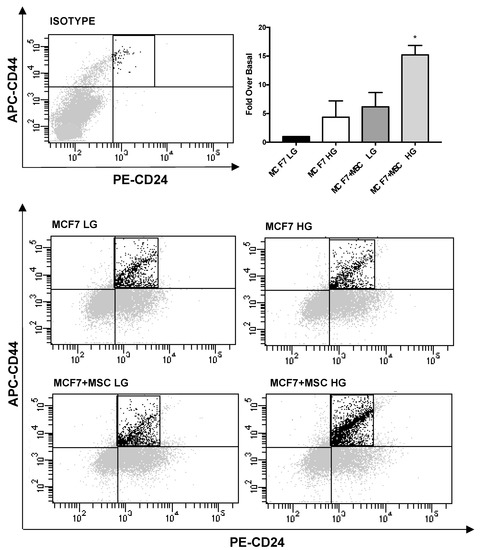
Figure 5.
Quantification of CD44high/CD24low subpopulation in BC cells co-cultured with MAT-MSCs. MCF7 were co-cultured—and not—with MAT-MSCs in LG or HG medium. After 72 h, CD44high/CD24low stem cell subpopulation was identified in monocultured (MCF7 LG/HG) and co-cultured (MCF7 + MSC LG/HG) MCF7 by FACS analysis (See Section 2). Representative dot plots show MCF7 stained with APC-anti-CD44 and PE-anti-CD24 as well as dye/isotype matched antibodies. In the graph, the percentage of population was reported as fold over basal (MCF7 LG) and showed CD44high/CD24low cell subpopulation as mean ± SD of 4 independent experiments. Data were analyzed using the non-parametric Friedman test followed by Dunn’s correction for multiple comparisons. * denotes statistically significant values compared with MCF7 LG (* adjp < 0.01).
3.5. Stem Phenotype of MAT-MSCs and BC Cells in 3D Cultures
Three-dimensional spheroids were established with MCF7 in presence and in absence of MAT-MSCs, in HG or LG. MCF7 were able to form a 2.8-fold higher number of mammospheres, also characterized by 50% increased diameter, when co-cultured with MAT-MSCs in HG, while not in LG (Figure 6; adjp < 0.05).
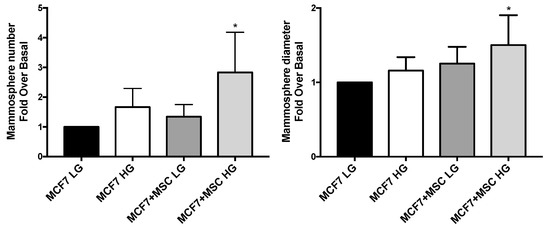
Figure 6.
Effect of glucose and MAT-MSCs on mammosphere formation. Three-dimensional cultures were set up by using MCF7 and MAT-MSCs (ratio 4:1) in LG or HG concentrations. After 10 days, mammosphere number and diameter were obtained. Results were reported as fold over basal (MCF7 LG). Bars represent mean ± SD of 4–5 independent experiments showing mammosphere number and diameter in BC cells, co-cultured and not with MAT-MSCs. Data were analyzed using the non-parametric Friedman test followed by Dunn’s correction for multiple comparisons. * denotes statistically significant values compared with MCF7 LG (* adjp < 0.05).
Notably, when disaggregated, MCF7 included in spheroids with MAT-MSCs in HG displayed a 4.3-fold increase in OCT44 protein (adjp < 0.05) with no change of NANOG levels (Figure 7a, Supplementary Figure S7a). In parallel, in the same spheroids, a significant increase in α-SMA (adjp < 0.05), FAP (adjp < 0.01) and p16INK4a (adjp < 0.05) proteins was observed in MAT-MSCs (Figure 7b, Supplementary Figure S7b). In addition, in these cells NANOG protein levels did not change. However, a 50% significant reduction of OCT4 (adjp < 0.05) was detected (Figure 7b, Supplementary Figure S7b).
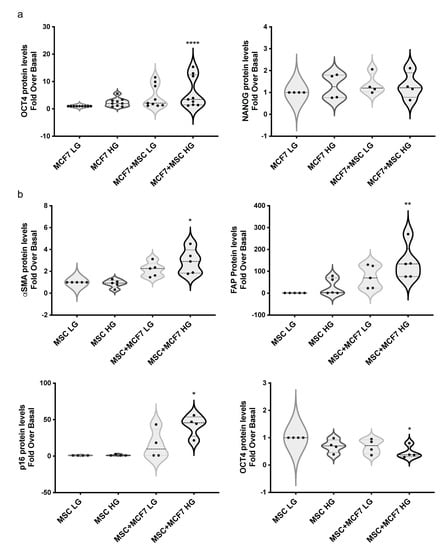
Figure 7.
Effect of glucose on MAT-MSCs and MCF7 crosstalk in 3D organoids. Three-dimensional cultures were set up by using MCF7 and MAT-MSCs (ratio 4:1) in LG or HG concentrations. After 72 h, spheroids were mechanically disaggregated and cells were stained with PE-anti-OCT4, PE-anti-NANOG, PE-anti-α-SMA, APC-anti-FAP and APC-anti-p16, as well as dye/isotype matched antibodies for FACS analysis. Results were reported as violin plot of 4–9 independent experiments showing protein levels of (a) Oct4 and Nanog in MCF7 or (b) α-SMA, FAP, p16 and OCT4 in MAT-MSCs as fold over basal (MCF7 LG/MSC LG). Data were analyzed using the non-parametric Friedman test followed by Dunn’s correction for multiple comparisons. * denotes statistically significant values compared with MCF7 LG or MSC LG (* adjp < 0.05; ** adjp < 0.01, **** adjp < 0.0001).
3.6. Glucose Modulates Invasiveness of MAT-MSCs and MCF7 Spheroids in Zebrafish Xenograft Model
Finally, spheroid-derived cells were engrafted in 2-day-old zebrafish embryos (Figure 8a). Immediately after injection, cells were localized in the yolk sac (Figure 8a, red spot). At 72 h post-injection, cells were able to migrate in head and tail regions (Figure 8b, white arrows). Monocultured MCF7 in LG and HG showed a similar dissemination rate. Indeed, 63% of xenografts with invasive MCF7 in LG, and 76% with invasive cells in HG were detected (Figure 8b). Interestingly, when injected MCF7+MSCs in LG, only 30% of xenografts displayed invasive BC cells, a percentage significantly reduced as compared to xenografts with MCF7 monocultures. In this condition, the majority of zebrafish displayed a clear localized tumor mass (Figure 8b, bigger white arrow). At variance, when injected, MCF7 + MSCs in HG 90% of xenografts showed invasive cells, a percentage significantly higher compared to xenografts with MCF7 monocultures or MCF7 + MSCs in LG (Figure 8b).
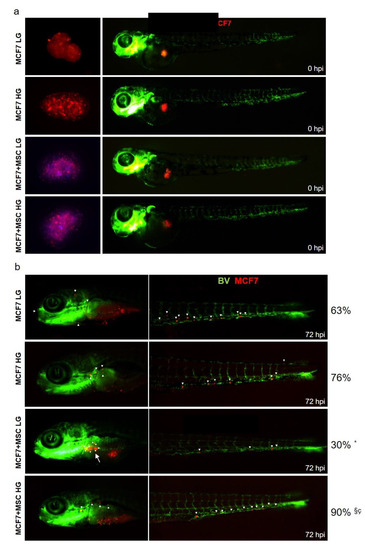
Figure 8.
Effect of glucose on MCF7-MAT-MSCs organoids in xenograft zebrafish models. MCF7 (labeled with red dye), cultured in spheroids alone or with MAT-MSCs (labeled with blue dye) in LG or HG, were injected into the PVS of 48 hpf zebrafish embryos. MCF7 and MCF7 + MSCs spheroids in LG and HG ((a), left) were disaggregated, injected and visualized under fluorescence stereo microscope at time point 0 hrs post-injection ((a), right). BV = blood vessel. MCF7 dissemination in LG and HG was analyzed 72 hrs post-injection (b). White arrows indicate migrated MCF7 cells in zebrafish head and tail. Results have been presented as percentage of xenograft with invasive BC cells. Data were analyzed using the chi-square with Fisher’s exact test. * denotes statistically significant values compared with MCF7 LG (* pval < 0.0001). § denotes statistically significant values compared with MCF7 HG (§ pval < 0.01). ç denotes statistically significant values compared with MSC + MCF7 LG (ç pval < 0.0001).
Thus, glucose also modulates BC-invasive features in in vivo models.
4. Discussion
Obesity and type 2 diabetes are epidemic diseases strictly associated with BC risk and progression. Many systemic factors, dysregulated in obesity and T2D, may contribute to BC, including insulin, insulin-like growth factor 1 (IGF1), glucose, lipids, inflammatory cytokines, immune cells, steroids, the autonomic nervous system, adipokines and the microbiome [9,34]. These factors may have direct pro-tumorigenic effects on BC cells but may also disrupt the local tumor microenvironment (TME). For instance, high glucose levels promote the proliferation, invasion, and migration of BC cells and enhance the chemoresistance of tumors via abnormal glucose metabolism. In parallel, glucose may modify the TME, increasing the PH, the lactic acid, production of inflammatory factors, imbalance of ROS and the cellular composition/phenotype [13,35].
Within the breast, cancer cells communicate with adipocytes and their mesenchymal precursors which release signaling molecules and provide mechanical support and energy supply [9,11]. Metabolic imbalances, such as high glucose levels, may affect such communication, contributing to the association between metabolic diseases and BC progression. Previously, we demonstrated that glucose promotes adipocyte release of IGF1, CCL5 and IL-8, enhancing BC cell proliferation, invasiveness and tamoxifen resistance [13,14,15].
Here, we have shown for the first time that glucose participates in the dialogue between mammary MSCs and BC cells, which leads to a MSC CAF-like phenotype and BC cell stem-like features.
To date, the contribution of MSCs in BC progression has been investigated mainly by using murine MSCs (mMSCs), human bone marrow-derived MSCs (BM-MSCs) or human abdominal adipose tissue (AT)-derived MSCs. However, biological differences between BM-MSCs and AT-derived MSCs have been described and, in the context of AT, the functional diversity of depots is well established [16,17,36]. Indeed, although different studies reported that MSCs support BC cell growth and migration [9,23], a piece of literature assigns tumor-suppressive functions to MSCs, considering them as suitable carriers for anti-cancer drug delivery [22,37]. Recently, it has been shown that MSCs isolated from mammary AT (MAT-MSCs) of patients with BC displayed a dysregulated secretory profile and enhanced tumorigenicity compared to MAT-MSCs isolated from healthy donors, suggesting permanent changes induced by cancer cells in local MSCs [25]. Moreover, it has been reported that MAT-MSCs are largely modified by doxorubicin and paclitaxel, which influence cytokine release and potentiate the ability of MSCs to stimulate BC invasive potential in vivo [24]. Thus, local MSCs exhibit a pivotal role in BC progression and are extensively affected by external stimuli.
Here, we defined the role of MSCs in BC progression by taking advantage of MAT-MSCs obtained from a cohort of healthy women. In this population no evidence of cell clusters due to interindividual differences were found. This is consistent with other studies showing no differences in gene expression profiles in MAT-MSCs obtained from healthy donors with different ages [25]. However, a future research direction will be the deeper study of the pro/anti tumorigenic effect of MAT-MSCs obtained from well-defined groups of donors with different anthropometric/clinical data.
In this study, we closely examined the role of glucose in the communication that MAT-MSCs establish with BC cells. Two- and three-dimensional in vitro co-cultures and in vivo zebrafish embryo models were established in 25 mM or 5.5 mM glucose (HG or LG, respectively), resembling hyperglycemia or close-to-normal fasting glucose levels in humans. Through this model, we provided evidence that MSCs are largely modified by ER+ and TNBC cells and promote BC cell stemness in an HG environment.
Interestingly, within a screening of cytokines, chemokines and growth factors, we observed increased release of IL-6, IL-8 and VEGF in co-cultures of MAT-MSCs with BC cells. Consistently, significantly higher IL6 and VEGF mRNA levels were detected when MAT-MSCs were co-cultured with MCF7 and MDA-MB231; in contrast, IL-8 expression was increased in MAT-MSCs only upon co-culture with MDA-MB231. IL-6, IL-8 and VEGF play a crucial role in tumor microenvironment and represent key signals in the communication between adipose, cancer, endothelial and immune cells [13,28,30,31,38]. High serum levels of IL-6, IL-8 and VEGF have been associated with poor BC prognosis/overall survival [31], but also with insulin resistance and diabetic complications [13,39,40]. Hence, targeted inhibitors are actually in use or in clinical trial either for BC [38] or diabetes [39,40,41]. Notably, IL-6, IL-8 and VEGF are also considered senescence-associated secretory phenotype (SASP) factors [29]. Here, we showed their over-expression, accompanied by a de-regulation of senescence markers LMNB1 and CDKN2A (the latter also at protein levels) in MAT-MSCs particularly upon co-culture with MCF7 cells, suggesting the acquisition of a senescent phenotype. In this model, although we did not find β-galactosidase (β-gal) activation, CDKN2A (and p16INK4A) increase and LMNB1 decline were detected [20,32,33]. In senescent cells their modifications appear earlier compared to β-gal, whose activity takes 7–10 days to develop upon specific senescent stimuli [32].
IL-6, IL-8 and VEGF are largely secreted also by cancer-associated fibroblasts (CAFs), the most heterogeneous population of stromal cells in the BC microenvironment. CAFs modulate cancer metastasis and influence angiogenesis, tumor mechanics, drug access and therapy responses [42,43]. Different studies reported the presence of senescent CAFs in solid tumors [20], with a more evident pro-tumorigenic role for senescent, compared to non-senescent CAFs [44,45]. Here, we have found that MAT-MSCs, cultured with BC cells, show an up-regulation of α-SMA and FAP, largely recognized markers of CAFs in BC. Notably, α-SMA expression in BC environment has been correlated with metastasis and poorer overall survival rate [46] and its expression is common to senescent and non-senescent CAFs [20]. The contemporary presence of α-SMA and FAP identifies also a new specific CAF subpopulation (referred to as CAF-S1), which promotes immunosuppression and immunotherapy resistance in BC [42,43]. FAP is still considered the most promising CAF therapeutic target. Indeed, FAP inhibition to eradicate CAFs has been associated with decreased collagen content and tumor burden, and improved immunotherapy effectiveness [47,48,49]. Our data suggest that, in presence of BC cells, MAT-MSCs acquire a CAF-like phenotype, but only in presence of ER+ cells they become senescent CAFs.
Interestingly, almost all markers of senescence and fibrosis were not induced by BC cells in MAT-MSCs in an LG environment. These results highlight that MAT-MSCs may transdifferentiate in CAFs and that their heterogeneity in BC could be driven by cancer subtypes. Notably, we have evidenced that fibro/senescent features acquired by MAT-MSCs upon co-culture with BC cells in HG are not stably retained. Hence, once removed from cancer cells, IL6, IL8, VEGF, CDKN2A and LMNB1 mRNA levels in MSCs return to levels observed in MSCs without cancer cells. However, it should be reported that CAFs are characterized by high plasticity and their properties may change upon longer-term cultures [50].
The loosening of an undifferentiated state of MAT-MSCs cultured with BC cells is pointed up also by the finding that in HG co-cultures they showed a reduction of OCT4, SOX2 and NANOG, which play a critical role for the maintenance of the multipotent/pluripotent state [51]. Few studies have investigated their functions in adult MSCs. It has been shown that these factors inhibit lineage genes by binding to the DMT1 promoter and downregulating p16 expression [52]. Accordingly, we have detected a reduction of multipotency-related genes and an increase in p16INK4A. Such effects might be, at least in part, related to the increase of α-SMA. Indeed, it has been documented that α-SMA-positive BM-MSCs exhibited a fibrotic phenotype, with reduced self-renewal, adipogenic differentiation, OCT4 and SOX2 levels [53]. However, in LG co-cultures, we detected no difference in multipotency genes, despite increased levels of α-SMA. Another possible mechanism may be represented by the glutamine withdrawal in the culture medium. BC cells display an upregulation of glutamine metabolism [54], while on the other hand, glutamine depletion has been shown to induce OCT4 degradation in embryonic stem cells [55].
Beside controlling stem cell fate, multipotency genes display a strong association with cancer progression. Indeed, OCT4 is an essential factor for the early stages of mammalian embryogenesis, but also the major regulator of cancer stem cells (CSCs) in BC. CSCs are tumor-initiating cells with a clear role in cancer recurrence, metastasis, and drug resistance. Although different factors may contribute to CSCs generation and maintenance, the mechanisms involved are largely unknown. Moreover, patients with high OCT4 expression in BC showed poor post-progression survival [51,56,57]. Diverse studies provided evidence that MSCs, mainly of murine origin, promote the acquisition of stem-like features of BC cells, via different pathways involving cytokines (i.e., IL-6, IL-8, CCL5), growth factors and miRNAs [58,59]. Here, we have shown, for the first time, that mammary MSCs, in an HG environment, increased OCT4 levels in ER+ BC cells. Instead, MDA-MB231 cells did not modify stem-related genes in presence of stromal stimuli. Similar results were obtained in BT-549 triple negative breast cancer cells. At variance, in BT-474, an ER+ and HER2+ BC cell model, even though not significant, increased levels of OCT4, SOX2 and NANOG in presence of MAT-MSCs, only in HG, were observed. These results need to be confirmed in human samples and in other BC cells, however they are in line with literature data reporting a different crosstalk between BC subtypes and the tumor microenvironment [60,61]. For instance, a recent study based on high-dimensional imaging describes how the specialized cells of the breast tumor microenvironment organize in space, how this organization varies across tumor subtypes and how this may impact clinical outcomes [60]. However, the signaling molecules that regulate the interplay between tumor environment and different BC subtypes have not yet been defined. TNBCs harbor the highest proportion of CSCs compared with other subtypes, contributing to the poor prognosis associated with this subtype [62,63]. Thus, it is possible to hypothesize that TNBC cells, which express high levels of OCT4 and hold numerous CD44high/CD24low cells, compared to other BC subtypes, do not respond to external stimuli (i.e., MAT-MSCs, glucose) to further increase stemness features. In contrast, MCF7, a less aggressive BC model, may be more prone to capturing external stimuli, making them ancillary signals. Accordingly, in these cells, the proportion of the CD44high/CD24low BC cell sub-population was increased in presence of MAT-MSCs in HG. Interestingly, we provided evidence also that in HG, a low number of MSCs was sufficient to expand BC mammosphere number and diameter. We found that spheroid-derived MCF7 and MAT-MSCs exhibit high OCT4 and α-SMA/FAP/p16INK4A protein levels, respectively. Both spheroid-derived cells have also been injected in zebrafish embryos, which are optically transparent and permissive to the xenograft of human tumor cells. This method allows in vivo delivery of a very limited number of cancer cells, mimicking the initial stages of tumor angiogenesis and metastasis; thus, it is now considered a useful approach to study the metastatic homing and colonization [64,65,66]. Zebrafish are becoming increasingly prominent in the study of the tumor microenvironment, since they allow for the reproduction of in vivo conditions in which cancer cells normally grow, consisting of blood vessels, lymphatic vessels, stromal cells, extracellular matrices, proteins and RNAs. However, there are still many needs to address with regard to mimicking the human tumor environment [65,67]. Here, we showed that the injection of spheroid-derived MCF7 + MSCs led to higher tumor invasiveness compared to that of spheroid-derived MCF7, only in HG. Indeed, in LG the percentage of xenografts with invasive cells was significantly lower. Our model is innovative, since it is based on a xenograft generated with BC cells + MAT-MSCs spheroids. It allows for the study of tumor/MSCs crosstalk during cancer cell invasion and metastasis and may be envisioned as novel approach for drug screening toward breast cancer.
5. Conclusions
In high glucose environments BC cells induce profound changes in mammary MSCs, increasing the expression of inflammation, senescence and fibrosis markers, and downregulating multipotency genes. Tumor-educated MAT-MSCs, in turn, promote a CSC phenotype in ER+ BC cells and tumor invasiveness in an in vivo model. Glucose lowering interferes with the dangerous communication between cancer cells and MAT-MSCs, counteracting, at least in part, these pro-tumorigenic effects.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/cancers14215421/s1, Table S1: Primer pairs for qPCR; Table S2: Clinical phenotyping of enrolled population; Figure S1: Heatmap of anthropometric and biochemical data of mammary adipose tissue (MAT) donors; Figure S2: Secretory profile of MAT-MSC isolates and correlation with donor anthropometric/biochemical data; Figure S3: Effect of glucose lowering on MAT-MSC phenotype; Figure S4: Effect of glucose and MAT-MSCs on MCF7 phenotype; Figure S5: Effect of glucose and MAT-MSCs on MDA-MB231 phenotype; Figure S6: Effect of glucose lowering on MCF7 stemness genes.
Author Contributions
M.R.A.: conceptualization; formal analysis; writing—original draft. G.M.: investigation; methodology, data curation. T.M.: investigation; validation; data curation. D.L.: investigation; methodology. G.N.: conceptualization; data curation. F.S.: conceptualization, supervision. F.D.: resources, visualization. F.L.: investigation; methodology. N.P.: data curation, investigation. R.M.M.: conceptualization, supervision. C.R.: data curation, investigation. C.A.: funding acquisition, supervision. C.M.: funding acquisition, project administration. F.B.: funding acquisition, writing—review and editing. V.D.: supervision; writing—original draft; writing—review and editing, conceptualization. P.F.: funding acquisition, supervision; conceptualization, project administration, writing—review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the Regione Campania POR FESR 2014–2020–Objective 1.2.—Realization of Technology Platform to fight oncologic diseases (RARE PLAT NET, SATIN, and COEPICA Projects) and by the Italian Association for the Cancer Research—AIRC (grant IG19001).
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki and approved by the ethical committee of the University of Naples (prot. n 138/16).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
The authors wish to thank Laura Cerchia and Augusta Moccia for scientific support and Antonio D’Andrea for technical support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Saeedi, P.; Petersohn, I.; Salpea, P.; Malanda, B.; Karuranga, S.; Unwin, N.; Colagiuri, S.; Guariguata, L.; Motala, A.A.; Ogurtsova, K.; et al. Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: Results from the International Diabetes Federation Diabetes Atlas, 9(th) edition. Diabetes Res. Clin. Pract. 2019, 157, 107843. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Hu, F.B. The global implications of diabetes and cancer. Lancet 2014, 383, 1947–1948. [Google Scholar] [CrossRef]
- Vigneri, P.; Frasca, F.; Sciacca, L.; Pandini, G.; Vigneri, R. Diabetes and cancer. Endocr.-Relat. Cancer 2009, 16, 1103–1123. [Google Scholar] [CrossRef] [PubMed]
- Larsson, S.C.; Mantzoros, C.S.; Wolk, A. Diabetes mellitus and risk of breast cancer: A meta-analysis. Int. J. Cancer 2007, 121, 856–862. [Google Scholar] [CrossRef]
- Peairs, K.S.; Barone, B.B.; Snyder, C.F.; Yeh, H.C.; Stein, K.B.; Derr, R.L.; Brancati, F.L.; Wolff, A.C. Diabetes mellitus and breast cancer outcomes: A systematic review and meta-analysis. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2011, 29, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Schrauder, M.G.; Fasching, P.A.; Haberle, L.; Lux, M.P.; Rauh, C.; Hein, A.; Bayer, C.M.; Heusinger, K.; Hartmann, A.; Strehl, J.D.; et al. Diabetes and prognosis in a breast cancer cohort. J. Cancer Res. Clin. Oncol. 2011, 137, 975–983. [Google Scholar] [CrossRef]
- Quail, D.F.; Joyce, J.A. Microenvironmental regulation of tumor progression and metastasis. Nat. Med. 2013, 19, 1423–1437. [Google Scholar] [CrossRef]
- Hanahan, D.; Coussens, L.M. Accessories to the crime: Functions of cells recruited to the tumor microenvironment. Cancer Cell 2012, 21, 309–322. [Google Scholar] [CrossRef]
- D’Esposito, V.; Ambrosio, M.R.; Giuliano, M.; Cabaro, S.; Miele, C.; Beguinot, F.; Formisano, P. Mammary Adipose Tissue Control of Breast Cancer Progression: Impact of Obesity and Diabetes. Front. Oncol. 2020, 10, 1554. [Google Scholar] [CrossRef]
- Lapeire, L.; Hendrix, A.; Lambein, K.; Van Bockstal, M.; Braems, G.; Van Den Broecke, R.; Limame, R.; Mestdagh, P.; Vandesompele, J.; Vanhove, C.; et al. Cancer-associated adipose tissue promotes breast cancer progression by paracrine oncostatin M and Jak/STAT3 signaling. Cancer Res. 2014, 74, 6806–6819. [Google Scholar] [CrossRef]
- La Camera, G.; Gelsomino, L.; Malivindi, R.; Barone, I.; Panza, S.; De Rose, D.; Giordano, F.; D’Esposito, V.; Formisano, P.; Bonofiglio, D.; et al. Adipocyte-derived extracellular vesicles promote breast cancer cell malignancy through HIF-1alpha activity. Cancer Lett. 2021, 521, 155–168. [Google Scholar] [CrossRef]
- Hemalatha, S.K.; Sengodan, S.K.; Nadhan, R.; Dev, J.; Sushama, R.R.; Somasundaram, V.; Thankappan, R.; Rajan, A.; Latha, N.R.; Varghese, G.R.; et al. Brcal Defective Breast Cancer Cells Induce in vitro Transformation of Cancer Associated Fibroblasts (CAFs) to Metastasis Associated Fibroblasts (MAF). Sci. Rep. 2018, 8, 13903. [Google Scholar] [CrossRef]
- Ambrosio, M.R.; D’Esposito, V.; Costa, V.; Liguoro, D.; Collina, F.; Cantile, M.; Prevete, N.; Passaro, C.; Mosca, G.; De Laurentiis, M.; et al. Glucose impairs tamoxifen responsiveness modulating connective tissue growth factor in breast cancer cells. Oncotarget 2017, 8, 109000–109017. [Google Scholar] [CrossRef] [PubMed]
- D’Esposito, V.; Liguoro, D.; Ambrosio, M.R.; Collina, F.; Cantile, M.; Spinelli, R.; Raciti, G.A.; Miele, C.; Valentino, R.; Campiglia, P.; et al. Adipose microenvironment promotes triple negative breast cancer cell invasiveness and dissemination by producing CCL5. Oncotarget 2016, 7, 24495–24509. [Google Scholar] [CrossRef] [PubMed]
- D’Esposito, V.; Passaretti, F.; Hammarstedt, A.; Liguoro, D.; Terracciano, D.; Molea, G.; Canta, L.; Miele, C.; Smith, U.; Beguinot, F.; et al. Adipocyte-released insulin-like growth factor-1 is regulated by glucose and fatty acids and controls breast cancer cell growth in vitro. Diabetologia 2012, 55, 2811–2822. [Google Scholar] [CrossRef] [PubMed]
- D’Esposito, V.; Lecce, M.; Marenzi, G.; Cabaro, S.; Ambrosio, M.R.; Sammartino, G.; Misso, S.; Migliaccio, T.; Liguoro, P.; Oriente, F.; et al. Platelet-rich plasma counteracts detrimental effect of high-glucose concentrations on mesenchymal stem cells from Bichat fat pad. J. Tissue Eng. Regen. Med. 2020, 14, 701–713. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Chen, F.; Liu, D.; Gu, F.; Wang, Y. Adipose tissue-derived stem cells in breast reconstruction: A brief review on biology and translation. Stem Cell Res. 2021, 12, 8. [Google Scholar] [CrossRef]
- Path, G.; Perakakis, N.; Mantzoros, C.S.; Seufert, J. Stem cells in the treatment of diabetes mellitus—Focus on mesenchymal stem cells. Metab. Clin. Exp. 2019, 90, 1–15. [Google Scholar] [CrossRef]
- Borzone, F.R.; Giorello, M.B.; Sanmartin, M.C.; Yannarelli, G.; Martinez, L.M.; Chasseing, N.A. Mesenchymal stem cells and cancer-associated fibroblasts as a therapeutic strategy for breast cancer. Br. J. Pharmacol. 2022. [Google Scholar] [CrossRef]
- Schosserer, M.; Grillari, J.; Breitenbach, M. The Dual Role of Cellular Senescence in Developing Tumors and Their Response to Cancer Therapy. Front. Oncol. 2017, 7, 278. [Google Scholar] [CrossRef]
- Turinetto, V.; Vitale, E.; Giachino, C. Senescence in Human Mesenchymal Stem Cells: Functional Changes and Implications in Stem Cell-Based Therapy. Int. J. Mol. Sci. 2016, 17, 1164. [Google Scholar] [CrossRef] [PubMed]
- Scioli, M.G.; Storti, G.; D’Amico, F.; Gentile, P.; Kim, B.S.; Cervelli, V.; Orlandi, A. Adipose-Derived Stem Cells in Cancer Progression: New Perspectives and Opportunities. Int. J. Mol. Sci. 2019, 20, 3296. [Google Scholar] [CrossRef] [PubMed]
- Kamat, P.; Schweizer, R.; Kaenel, P.; Salemi, S.; Calcagni, M.; Giovanoli, P.; Gorantla, V.S.; Eberli, D.; Andres, A.C.; Plock, J.A. Human Adipose-Derived Mesenchymal Stromal Cells May Promote Breast Cancer Progression and Metastatic Spread. Plast. Reconstr. Surg. 2015, 136, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Plava, J.; Burikova, M.; Cihova, M.; Trnkova, L.; Smolkova, B.; Babal, P.; Krivosikova, L.; Janega, P.; Rojikova, L.; Drahosova, S.; et al. Chemotherapy-triggered changes in stromal compartment drive tumor invasiveness and progression of breast cancer. J. Exp. Clin. Cancer Res. CR 2021, 40, 302. [Google Scholar] [CrossRef]
- Plava, J.; Cihova, M.; Burikova, M.; Bohac, M.; Adamkov, M.; Drahosova, S.; Rusnakova, D.; Pindak, D.; Karaba, M.; Simo, J.; et al. Permanent Pro-Tumorigenic Shift in Adipose Tissue-Derived Mesenchymal Stromal Cells Induced by Breast Malignancy. Cells 2020, 9, 480. [Google Scholar] [CrossRef]
- D’Esposito, V.; Ambrosio, M.R.; Liguoro, D.; Perruolo, G.; Lecce, M.; Cabaro, S.; Aprile, M.; Marino, A.; Pilone, V.; Forestieri, P.; et al. In severe obesity, subcutaneous adipose tissue cell-derived cytokines are early markers of impaired glucose tolerance and are modulated by quercetin. Int. J. Obes. 2021, 45, 1811–1820. [Google Scholar] [CrossRef]
- Aprile, M.; Cataldi, S.; Ambrosio, M.R.; D’Esposito, V.; Lim, K.; Dietrich, A.; Bluher, M.; Savage, D.B.; Formisano, P.; Ciccodicola, A.; et al. PPARgammaDelta5, a Naturally Occurring Dominant-Negative Splice Isoform, Impairs PPARgamma Function and Adipocyte Differentiation. Cell Rep. 2018, 25, 1577–1592.e6. [Google Scholar] [CrossRef]
- Visciano, C.; Liotti, F.; Prevete, N.; Cali, G.; Franco, R.; Collina, F.; de Paulis, A.; Marone, G.; Santoro, M.; Melillo, R.M. Mast cells induce epithelial-to-mesenchymal transition and stem cell features in human thyroid cancer cells through an IL-8-Akt-Slug pathway. Oncogene 2015, 34, 5175–5186. [Google Scholar] [CrossRef]
- Laberge, R.M.; Sun, Y.; Orjalo, A.V.; Patil, C.K.; Freund, A.; Zhou, L.; Curran, S.C.; Davalos, A.R.; Wilson-Edell, K.A.; Liu, S.; et al. MTOR regulates the pro-tumorigenic senescence-associated secretory phenotype by promoting IL1A translation. Nat. Cell Biol. 2015, 17, 1049–1061. [Google Scholar] [CrossRef]
- Pallegar, N.K.; Christian, S.L. Adipocytes in the Tumour Microenvironment. Adv. Exp. Med. Biol. 2020, 1234, 1–13. [Google Scholar] [CrossRef]
- Todorovic-Rakovic, N.; Milovanovic, J. Interleukin-8 in breast cancer progression. J. Interferon Cytokine Res. Off. J. Int. Soc. Interferon Cytokine Res. 2013, 33, 563–570. [Google Scholar] [CrossRef] [PubMed]
- Freund, A.; Laberge, R.M.; Demaria, M.; Campisi, J. Lamin B1 loss is a senescence-associated biomarker. Mol. Biol. Cell 2012, 23, 2066–2075. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.Y.; Souroullas, G.P.; Diekman, B.O.; Krishnamurthy, J.; Hall, B.M.; Sorrentino, J.A.; Parker, J.S.; Sessions, G.A.; Gudkov, A.V.; Sharpless, N.E. Cells exhibiting strong p16 (INK4a) promoter activation in vivo display features of senescence. Proc. Natl. Acad. Sci. USA 2019, 116, 2603–2611. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, E.J.; LeRoith, D. Hyperinsulinaemia in cancer. Nat. Rev. Cancer 2020, 20, 629–644. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Zheng, Q.; Meng, X. Hyperglycemia and Chemoresistance in Breast Cancer: From Cellular Mechanisms to Treatment Response. Front. Oncol. 2021, 11, 628359. [Google Scholar] [CrossRef]
- Zwick, R.K.; Guerrero-Juarez, C.F.; Horsley, V.; Plikus, M.V. Anatomical, Physiological, and Functional Diversity of Adipose Tissue. Cell Metab. 2018, 27, 68–83. [Google Scholar] [CrossRef]
- Atiya, H.; Frisbie, L.; Pressimone, C.; Coffman, L. Mesenchymal Stem Cells in the Tumor Microenvironment. Adv. Exp. Med. Biol. 2020, 1234, 31–42. [Google Scholar] [CrossRef]
- Wu, Q.; Li, B.; Li, Z.; Li, J.; Sun, S.; Sun, S. Cancer-associated adipocytes: Key players in breast cancer progression. J. Hematol. Oncol. 2019, 12, 95. [Google Scholar] [CrossRef]
- Kusminski, C.M.; Bickel, P.E.; Scherer, P.E. Targeting adipose tissue in the treatment of obesity-associated diabetes. Nat. Rev. Drug Discov. 2016, 15, 639–660. [Google Scholar] [CrossRef]
- Long, L.; Li, Y.; Yu, S.; Li, X.; Hu, Y.; Long, T.; Wang, L.; Li, W.; Ye, X.; Ke, Z.; et al. Scutellarin Prevents Angiogenesis in Diabetic Retinopathy by Downregulating VEGF/ERK/FAK/Src Pathway Signaling. J. Diabetes Res. 2019, 2019, 4875421. [Google Scholar] [CrossRef]
- Citro, A.; Cantarelli, E.; Piemonti, L. The CXCR1/2 Pathway: Involvement in Diabetes Pathophysiology and Potential Target for T1D Interventions. Curr. Diabetes Rep. 2015, 15, 68. [Google Scholar] [CrossRef] [PubMed]
- Magagna, I.; Gourdin, N.; Kieffer, Y.; Licaj, M.; Mhaidly, R.; Andre, P.; Morel, A.; Vincent-Salomon, A.; Paturel, C.; Mechta-Grigoriou, F. CD73-Mediated Immunosuppression Is Linked to a Specific Fibroblast Population That Paves the Way for New Therapy in Breast Cancer. Cancers 2021, 13, 5878. [Google Scholar] [CrossRef] [PubMed]
- Rivas, E.I.; Linares, J.; Zwick, M.; Gomez-Llonin, A.; Guiu, M.; Labernadie, A.; Badia-Ramentol, J.; Llado, A.; Bardia, L.; Perez-Nunez, I.; et al. Targeted immunotherapy against distinct cancer-associated fibroblasts overcomes treatment resistance in refractory HER2+ breast tumors. Nat. Commun. 2022, 13, 5310. [Google Scholar] [CrossRef] [PubMed]
- Hassona, Y.; Cirillo, N.; Heesom, K.; Parkinson, E.K.; Prime, S.S. Senescent cancer-associated fibroblasts secrete active MMP-2 that promotes keratinocyte dis-cohesion and invasion. Br. J. Cancer 2014, 111, 1230–1237. [Google Scholar] [CrossRef]
- Wang, T.; Notta, F.; Navab, R.; Joseph, J.; Ibrahimov, E.; Xu, J.; Zhu, C.Q.; Borgida, A.; Gallinger, S.; Tsao, M.S. Senescent Carcinoma-Associated Fibroblasts Upregulate IL8 to Enhance Prometastatic Phenotypes. Mol. Cancer Res. MCR 2017, 15, 3–14. [Google Scholar] [CrossRef]
- Yamashita, M.; Ogawa, T.; Zhang, X.; Hanamura, N.; Kashikura, Y.; Takamura, M.; Yoneda, M.; Shiraishi, T. Role of stromal myofibroblasts in invasive breast cancer: Stromal expression of alpha-smooth muscle actin correlates with worse clinical outcome. Breast Cancer 2012, 19, 170–176. [Google Scholar] [CrossRef]
- Cremasco, V.; Astarita, J.L.; Grauel, A.L.; Keerthivasan, S.; MacIsaac, K.; Woodruff, M.C.; Wu, M.; Spel, L.; Santoro, S.; Amoozgar, Z.; et al. FAP Delineates Heterogeneous and Functionally Divergent Stromal Cells in Immune-Excluded Breast Tumors. Cancer Immunol. Res. 2018, 6, 1472–1485. [Google Scholar] [CrossRef]
- Han, C.; Liu, T.; Yin, R. Biomarkers for cancer-associated fibroblasts. Biomark. Res. 2020, 8, 64. [Google Scholar] [CrossRef]
- Xin, L.; Gao, J.; Zheng, Z.; Chen, Y.; Lv, S.; Zhao, Z.; Yu, C.; Yang, X.; Zhang, R. Fibroblast Activation Protein-alpha as a Target in the Bench-to-Bedside Diagnosis and Treatment of Tumors: A Narrative Review. Front. Oncol. 2021, 11, 648187. [Google Scholar] [CrossRef]
- Sahai, E.; Astsaturov, I.; Cukierman, E.; DeNardo, D.G.; Egeblad, M.; Evans, R.M.; Fearon, D.; Greten, F.R.; Hingorani, S.R.; Hunter, T.; et al. A framework for advancing our understanding of cancer-associated fibroblasts. Nat. Rev. Cancer 2020, 20, 174–186. [Google Scholar] [CrossRef]
- Seymour, T.; Twigger, A.J.; Kakulas, F. Pluripotency Genes and Their Functions in the Normal and Aberrant Breast and Brain. Int. J. Mol. Sci. 2015, 16, 27288–27301. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.C.; Su, P.F.; Huang, Y.F.; Yew, T.L.; Hung, S.C. Oct4 and Nanog directly regulate Dnmt1 to maintain self-renewal and undifferentiated state in mesenchymal stem cells. Mol. Cell 2012, 47, 169–182. [Google Scholar] [CrossRef]
- Talele, N.P.; Fradette, J.; Davies, J.E.; Kapus, A.; Hinz, B. Expression of alpha-Smooth Muscle Actin Determines the Fate of Mesenchymal Stromal Cells. Stem Cell Rep. 2015, 4, 1016–1030. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Wang, M.; Wang, M.; Yu, X.; Guo, J.; Sun, T.; Li, X.; Yao, L.; Dong, H.; Xu, Y. Metabolic Reprogramming in Triple-Negative Breast Cancer. Front. Oncol. 2020, 10, 428. [Google Scholar] [CrossRef] [PubMed]
- Marsboom, G.; Zhang, G.F.; Pohl-Avila, N.; Zhang, Y.; Yuan, Y.; Kang, H.; Hao, B.; Brunengraber, H.; Malik, A.B.; Rehman, J. Glutamine Metabolism Regulates the Pluripotency Transcription Factor OCT4. Cell Rep. 2016, 16, 323–332. [Google Scholar] [CrossRef]
- Cho, Y.; Kang, H.G.; Kim, S.J.; Lee, S.; Jee, S.; Ahn, S.G.; Kang, M.J.; Song, J.S.; Chung, J.Y.; Yi, E.C.; et al. Post-translational modification of OCT4 in breast cancer tumorigenesis. Cell Death Differ. 2018, 25, 1781–1795. [Google Scholar] [CrossRef]
- Zhang, Q.; Han, Z.; Zhu, Y.; Chen, J.; Li, W. The Role and Specific Mechanism of OCT4 in Cancer Stem Cells: A Review. Int. J. Stem Cells 2020, 13, 312–325. [Google Scholar] [CrossRef]
- Goto, H.; Shimono, Y.; Funakoshi, Y.; Imamura, Y.; Toyoda, M.; Kiyota, N.; Kono, S.; Takao, S.; Mukohara, T.; Minami, H. Adipose-derived stem cells enhance human breast cancer growth and cancer stem cell-like properties through adipsin. Oncogene 2019, 38, 767–779. [Google Scholar] [CrossRef]
- Liu, S.; Ginestier, C.; Ou, S.J.; Clouthier, S.G.; Patel, S.H.; Monville, F.; Korkaya, H.; Heath, A.; Dutcher, J.; Kleer, C.G.; et al. Breast cancer stem cells are regulated by mesenchymal stem cells through cytokine networks. Cancer Res. 2011, 71, 614–624. [Google Scholar] [CrossRef]
- Danenberg, E.; Bardwell, H.; Zanotelli, V.R.T.; Provenzano, E.; Chin, S.F.; Rueda, O.M.; Green, A.; Rakha, E.; Aparicio, S.; Ellis, I.O.; et al. Breast tumor microenvironment structures are associated with genomic features and clinical outcome. Nat. Genet. 2022, 54, 660–669. [Google Scholar] [CrossRef]
- Turner, K.M.; Yeo, S.K.; Holm, T.M.; Shaughnessy, E.; Guan, J.L. Heterogeneity within molecular subtypes of breast cancer. Am. J. Physiol. Cell Physiol. 2021, 321, C343–C354. [Google Scholar] [CrossRef] [PubMed]
- Fultang, N.; Chakraborty, M.; Peethambaran, B. Regulation of cancer stem cells in triple negative breast cancer. Cancer Drug Resist. 2021, 4, 321–342. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Chen, Q.; Zou, Y.; Chen, H.; Qi, L.; Chen, Y. Stem Cells and Cellular Origins of Breast Cancer: Updates in the Rationale, Controversies, and Therapeutic Implications. Front. Oncol. 2019, 9, 820. [Google Scholar] [CrossRef]
- Tobia, C.; Gariano, G.; De Sena, G.; Presta, M. Zebrafish embryo as a tool to study tumor/endothelial cell cross-talk. Biochim. Biophys. Acta 2013, 1832, 1371–1377. [Google Scholar] [CrossRef]
- Chen, X.; Li, Y.; Yao, T.; Jia, R. Benefits of Zebrafish Xenograft Models in Cancer Research. Front. Cell Dev. Biol. 2021, 9, 616551. [Google Scholar] [CrossRef]
- Mondal, C.; Gacha-Garay, M.J.; Larkin, K.A.; Adikes, R.C.; Di Martino, J.S.; Chien, C.C.; Fraser, M.; Eni-Aganga, I.; Agullo-Pascual, E.; Cialowicz, K.; et al. A proliferative to invasive switch is mediated by srGAP1 downregulation through the activation of TGF-beta2 signaling. Cell Rep. 2022, 40, 111358. [Google Scholar] [CrossRef] [PubMed]
- Gamble, J.T.; Elson, D.J.; Greenwood, J.A.; Tanguay, R.L.; Kolluri, S.K. The Zebrafish Xenograft Models for Investigating Cancer and Cancer Therapeutics. Biology 2021, 10, 252. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).