

Anoikis-Associated Lung Cancer Metastasis: Mechanisms and Therapies
 , ,
, , 
Abstract
Simple Summary
Abstract
1. Introduction
2. Methodology
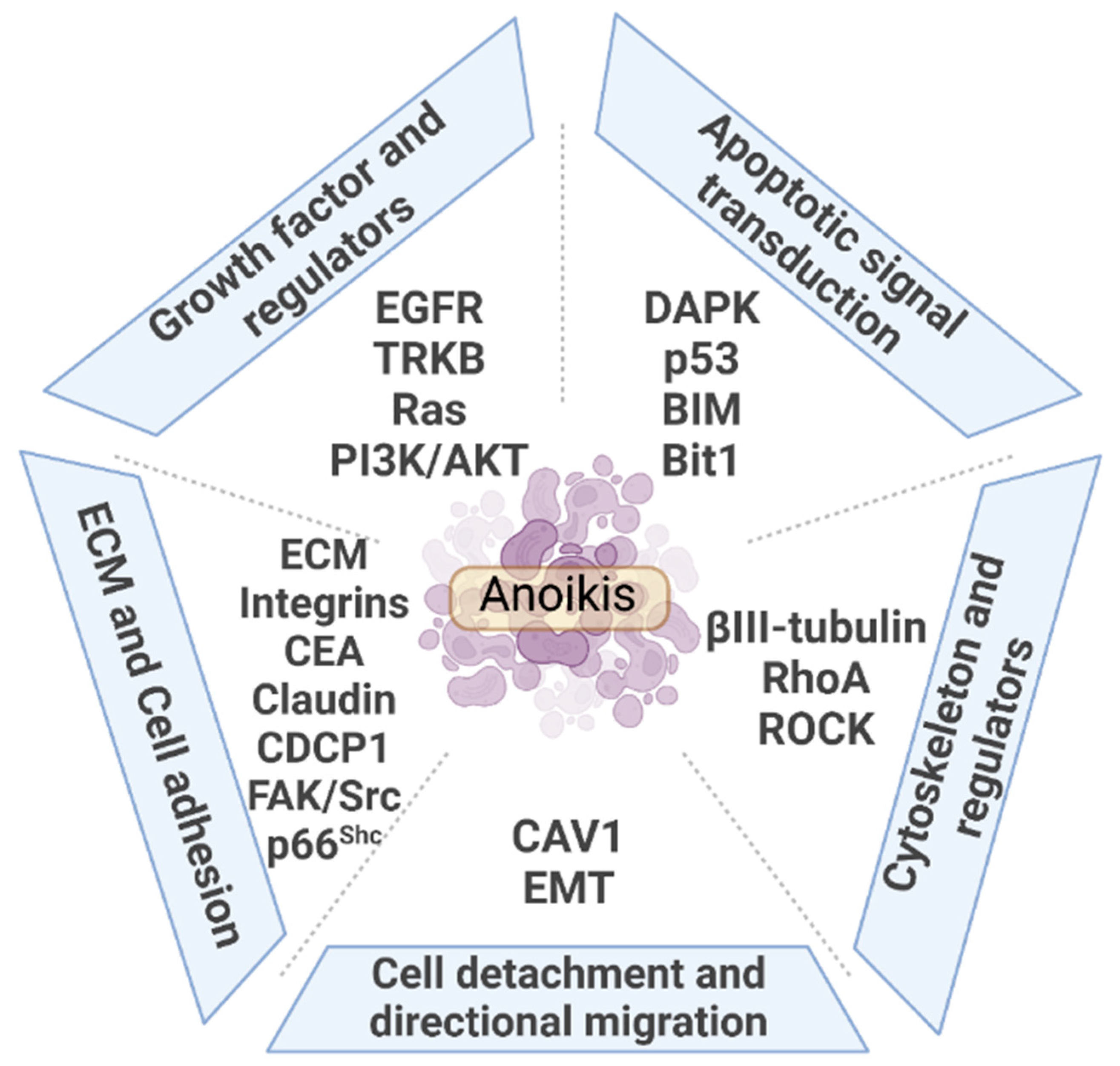
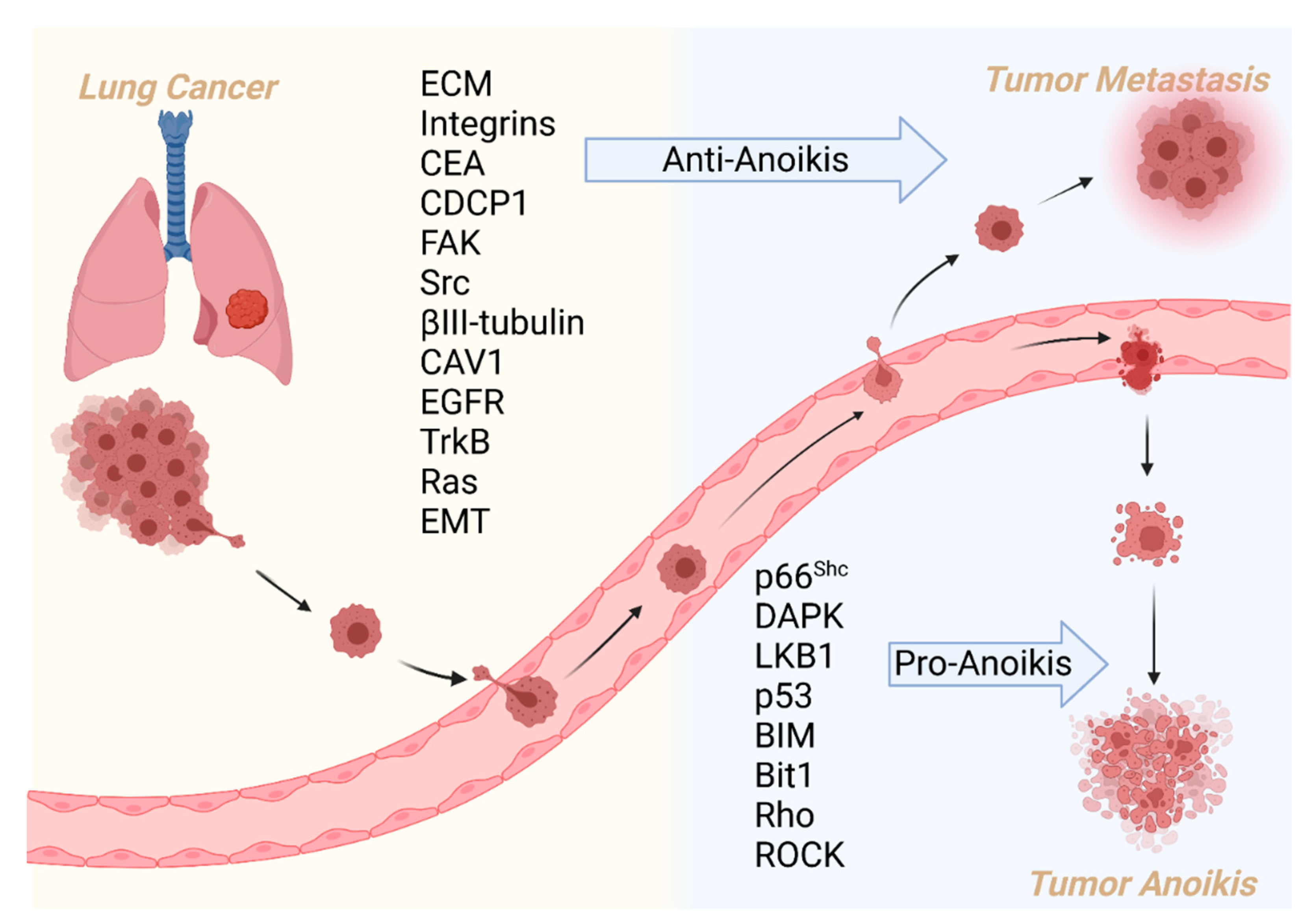
3. Regulators and Mechanism of Anoikis in Lung Cancer
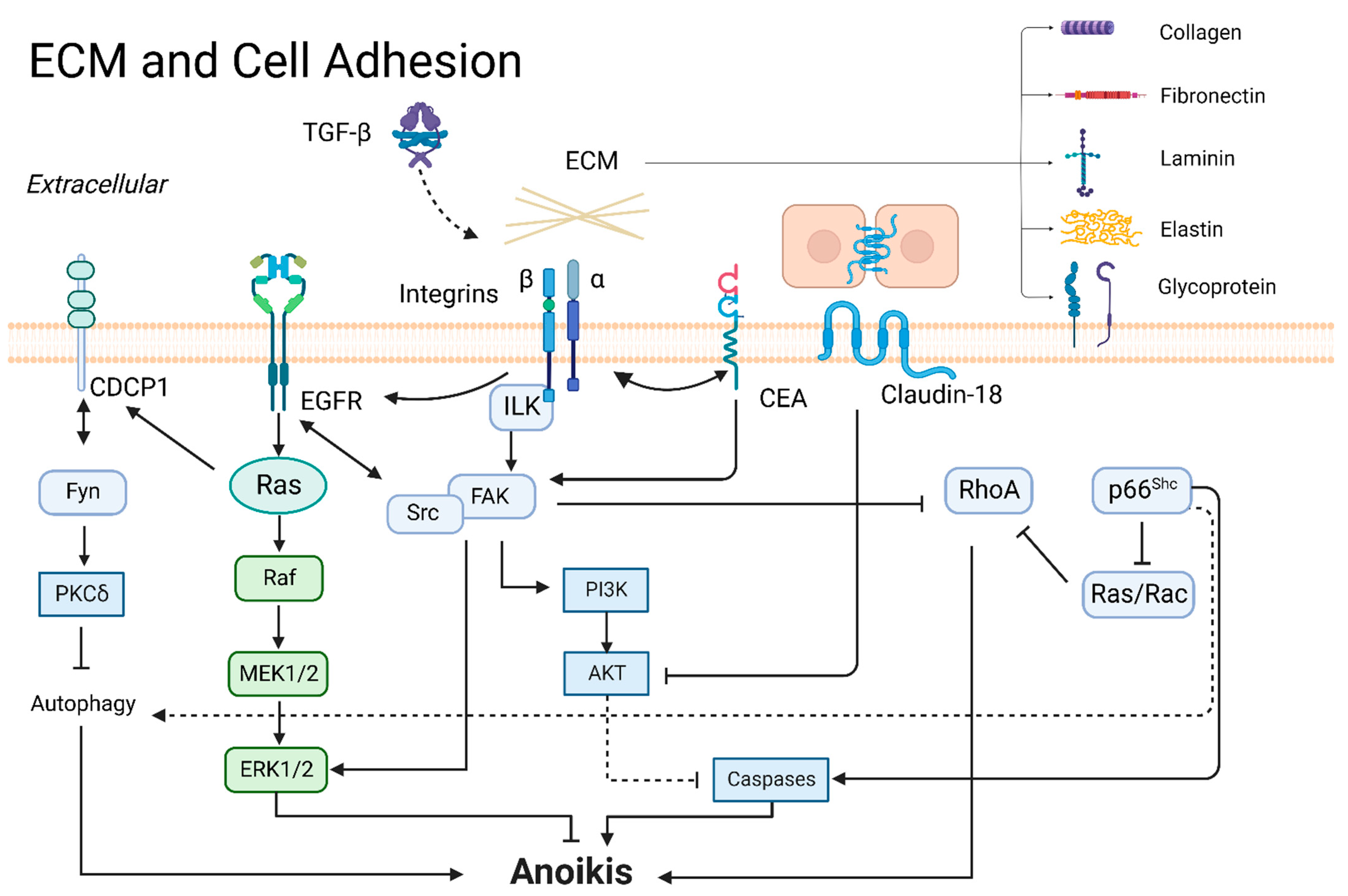
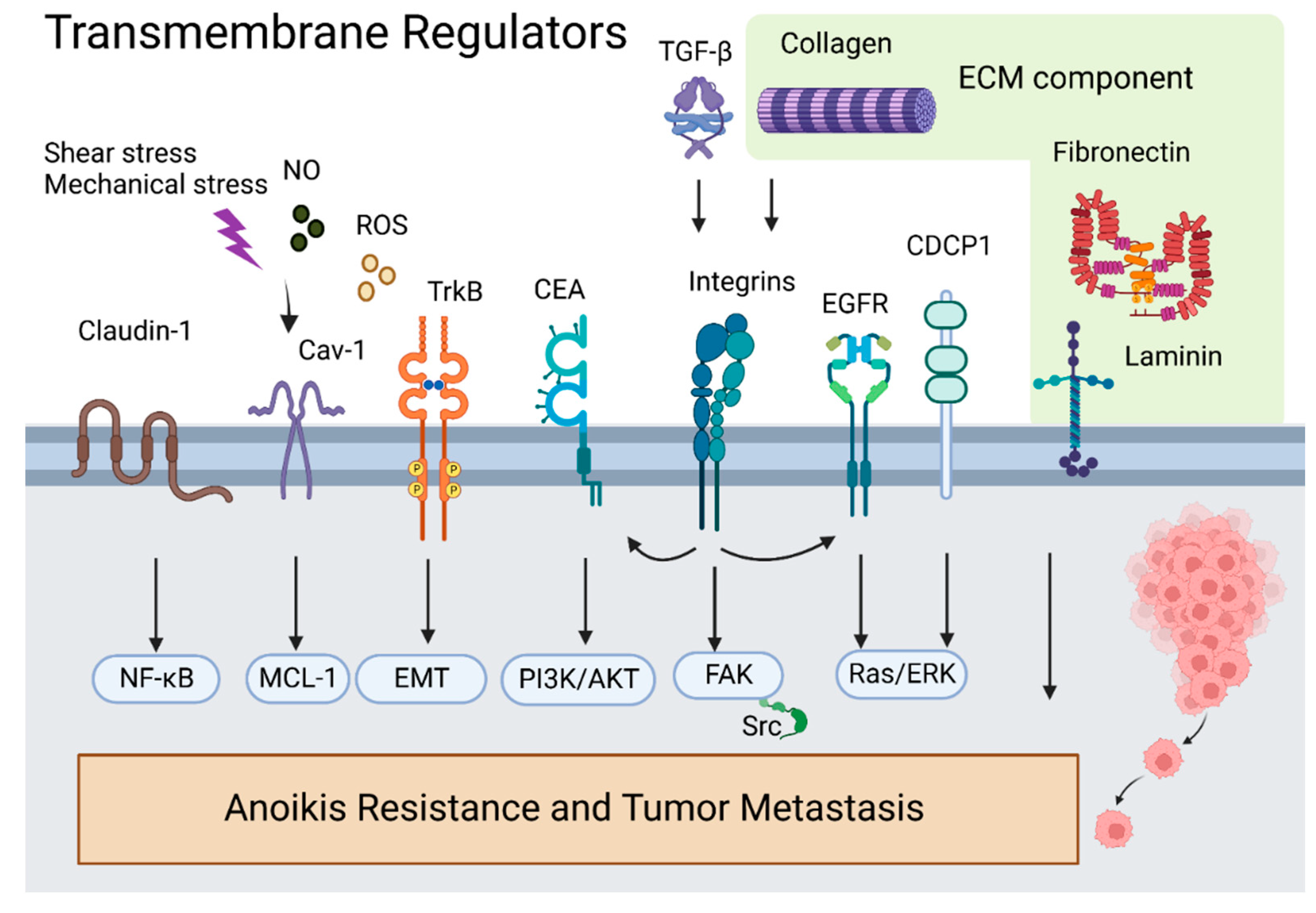
3.1. ECM and Cell Adhesion
3.1.1. ECM
3.1.2. Integrins
3.1.3. CEA
3.1.4. Claudins and Occludin
3.1.5. CUB-Domain-Containing Protein 1 (CDCP1)
3.1.6. FAK
3.1.7. Tyrosine-Protein Kinase Src
3.1.8. p66Shc
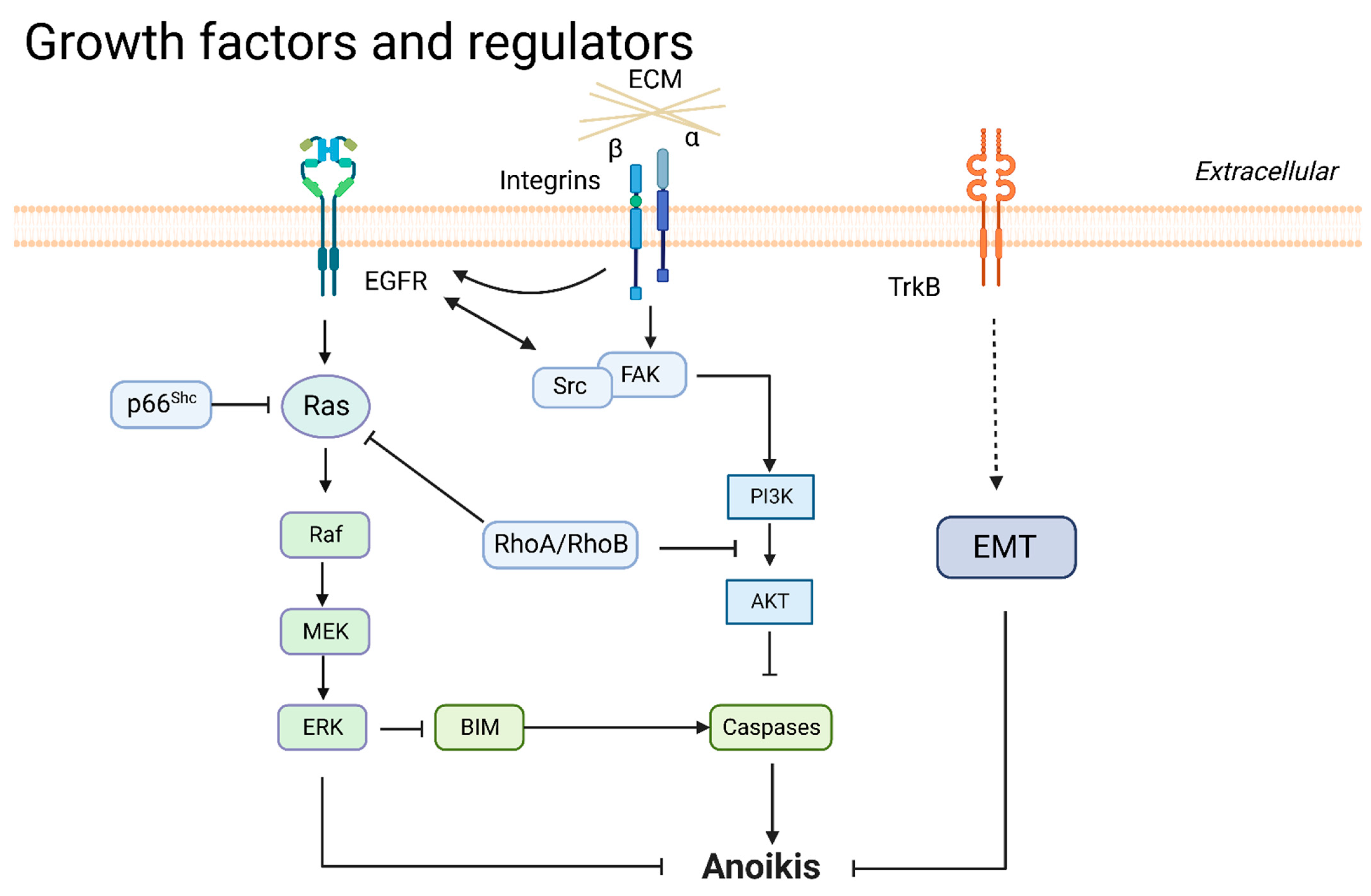
3.2. Growth Factors and Regulators
3.2.1. EGFR
3.2.2. Neurotrophic Tyrosine Kinase Receptor 2 (NTRK2/TrkB)
3.2.3. Ras
3.2.4. PI3K/AKT
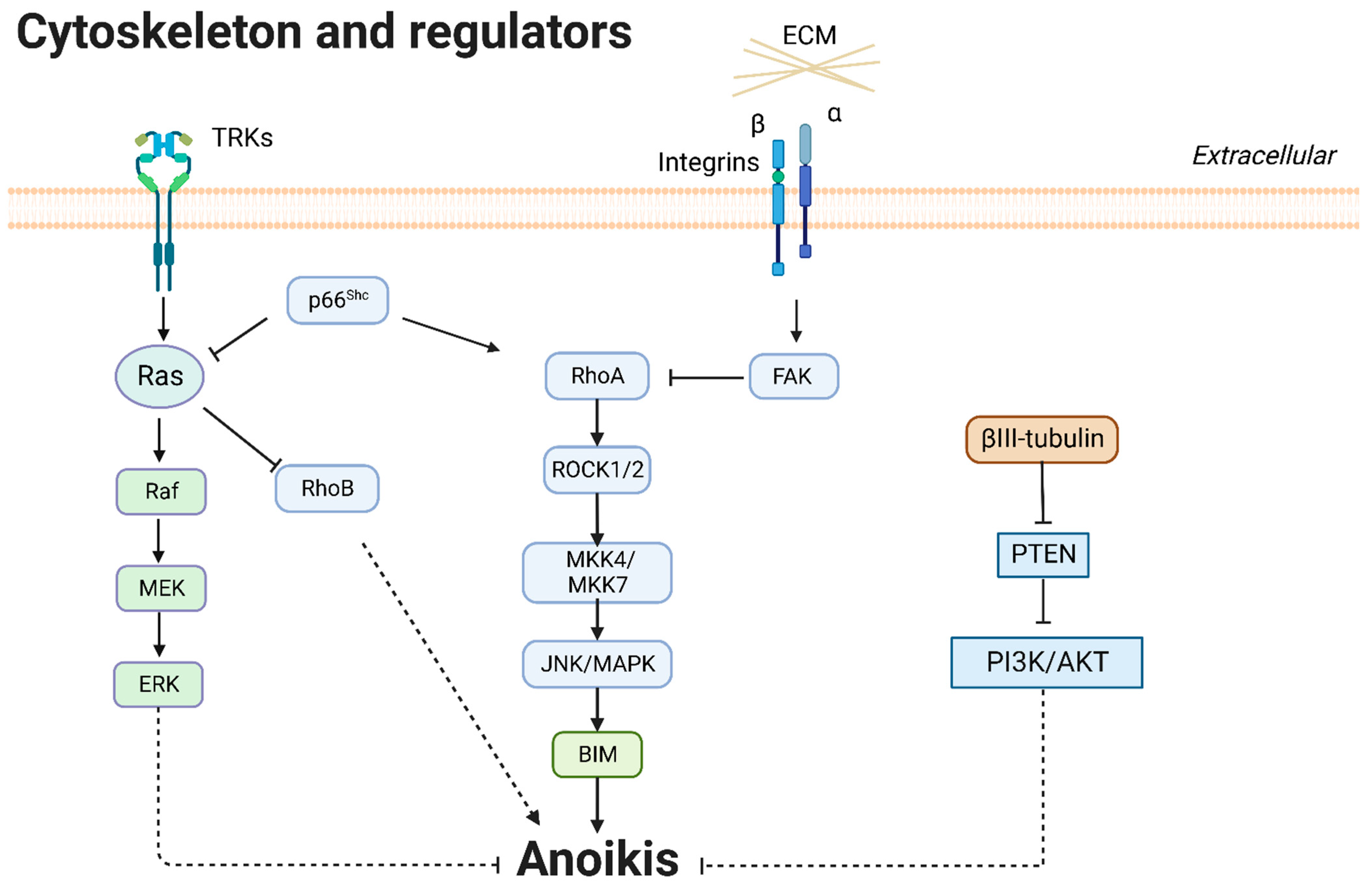
3.3. Cytoskeleton and Regulators
3.3.1. βIII-Tubulin
3.3.2. Rho and Rho-Associated Kinase (ROCK)
3.4. Cell Detachment and Directional Migration
3.4.1. CAV1
3.4.2. EMT
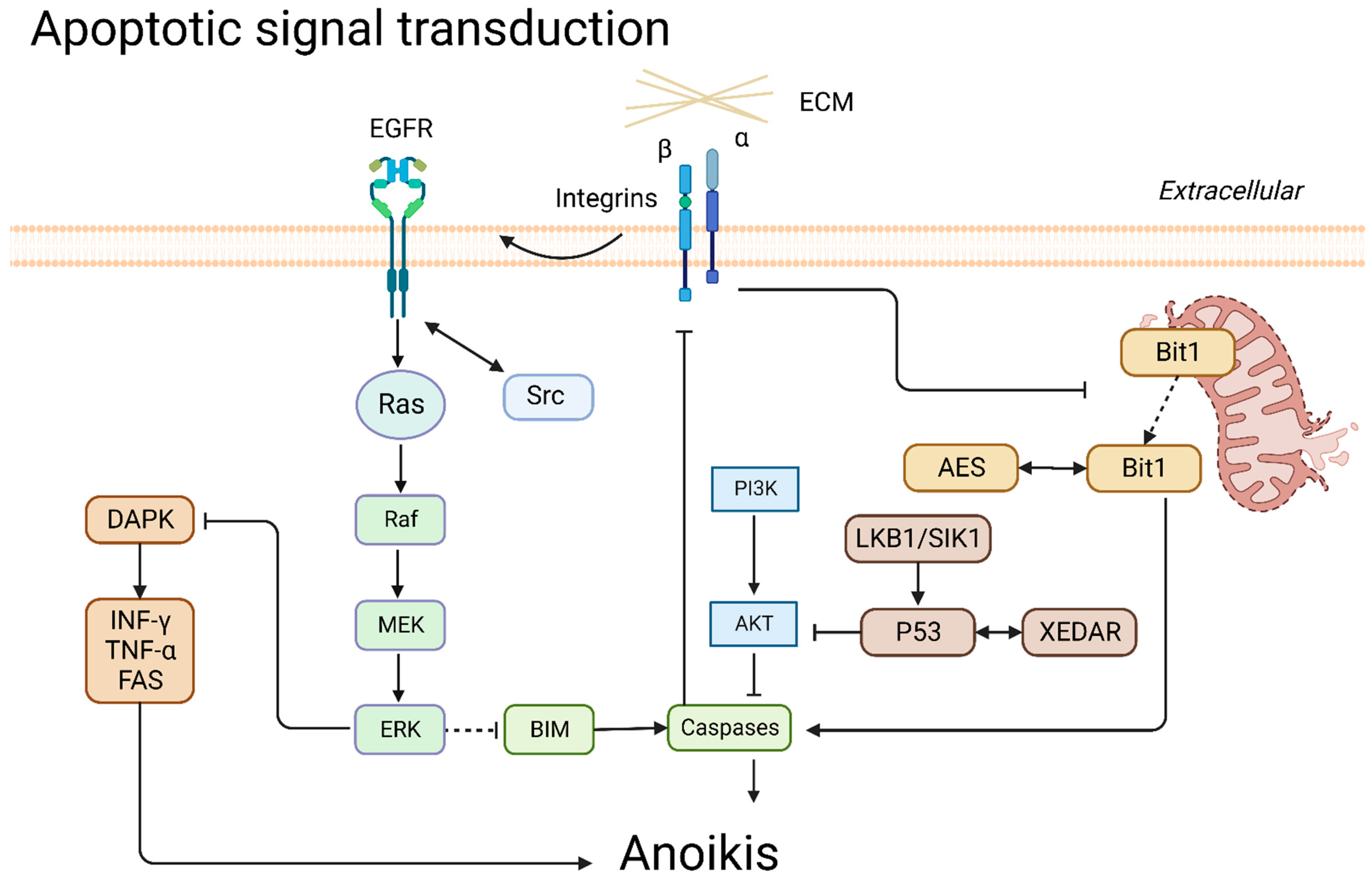
3.5. Apoptotic Signal Transduction
3.5.1. Death-Associated Protein Kinase (DAPK)
3.5.2. p53
3.5.3. BIM
3.5.4. Bit1
3.6. Other Regulators
3.6.1. LKB1
3.6.2. Other Regulators

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factors | Anoikis | Model(s) | Mechanism | Ref |
|---|---|---|---|---|
| Fibronectin | ↓ | A549, H460, and H1975 | Fibronectin upregulated cell desmosomal interactions. | [19] |
| Laminin 5 | ↓ | A549, PC-14, LC-2/ad, RERF-LC-KJ, NCI-H322, and NCI-H358 | Laminin 5/integrin/FAK signaling pathway activated the expression of EGFR. | [20] |
| Collagen XVII | ↓ | A549 | S727 phosphorylation of STAT3 activated collagen XVII to maintain the stability of laminin 5. | [21] |
| Collagen IV | ↓ | Lung-metastasizing M-27 cells | Collagen IV activated integrin α2/FAK and increased reactivity to IGF-I. | [22] |
| CRABP2 | ↓ | L1, C9F6, and H1650 | CRABP2 coupled with HuR promoted integrin β1/FAK/ERK signaling. | [25] |
| Integrin β3 | ↓ | HCC827, H1975, A549, H292, and H1299 | The activation of the TGFβ1/integrin β3 axis overcame acquired resistance to EGFR-TKIs and anoikis. | [27] |
| CEA | ↓ | L6 and LR-73 | CEA inactivated caspase-9 and caspase-8 and enhanced PI3K/AKT pathway. | [28,30,31] |
| Claudin-1, Serglycin, and CD44 | ↓ | H1299, H322, H358, H23, H928, H460, and A549 | Serglycin interacting with CD44 enhanced claudin-1 expression to promote EMT and anoikis resistance. | [33] |
| Claudin-18 | ↑ | A549, RERF-LC-AI, IA-LM, WA-hT, PC-3, and RERF-LC–MS | Claudin-18 inactivated PDK1 and phospho-AKT levels. | [34] |
| Occludin and TTF-1 | ↑ | H441, A549, H1299 | TTF-1 transactivated occludin to promote anoikis. | [35] |
| CDCP1 | ↓ | A549, PC14, H322, H520, and H157 | Tyr734-phosphorylated CDCP1 regulated PKCδ and inhibited autophagy. | [36,38,39] |
| FAK | ↓ | A549 | Phosphorylated tyrosine sites (Tyr397, Tyr861, Tyr925) in FAK bound to intracellular proteins of Src and stimulated PI3K/AKT pathway, MAPK/ERK pathway, and MAPK/p38 pathway. | [42] |
| Olfactomedin III | ↓ | Poorly differentiated human squamous carcinoma, named DLKP cell line | Olfactomedin III upregulated phospho-FAK and phospho-Paxillin and kept procaspase-3 from activation. | [43] |
| Src | ↓ | SK-LU-1, H522, H1437, A549, H460, and H1792 | EGFR, PDGFR, and ZIC2 increased the phosphorylation of Src to induce anoikis resistance. | [48,50,52] |
| p130Cas | ↓ | A549 and H1792 | Src contributed to the phosphorylation of p130Cas in the tumor cells. | [49] |
| ZIC2 | ↓ | HFL1, Calu-3, H1975, H1395, H520, A549, H1299, H226, SK-MES-1, and BEAS-2B | ZIC2 promoted tumorigenesis and anoikis resistance of NSCLC by Src/FAK signaling. | [50] |
| FAM188B | ↓ | A549, H1299, H1975 | FAM188B potentiated the stabilization of EGFR expression. | [52] |
| p66Shc | ↑ | LLC, H69, and H209 | p66Shc potentiated autophagy through cleavage of caspase 7 and PARP. | [57,58] |
| Aiolos | ↓ | A549, H1155 | Aiolos disrupted the enhancer of p66Shc at the transcription level. | [61] |
| PRDM1 | ↑ | HUVEC, A549, Beas-2B, H209, H69, H1155 | PRDM1 was silenced by aiolos. | [62] |
| EGFR | ↓ | A549, H1703, Calu-6, H460, H358, HCC2279, BEAS-2B | EGFR mediated phosphorylation of c-Src and ERK to attenuate anoikis. | [51,52,66] |
| NOX4 | ↓ | A549, H1703, Calu-6, H460, H358, HCC2279, BEAS-2B | NOX4 increased the activation of Src and EGFR, attenuating anoikis | [51] |
| TrkB | ↓ | Primary cell cultures derived from malignant pleural effusions, L1 sarcoma cells from a primary, spontaneous lung tumor in Balb/c | TrkB inhibited EMT. The expression levels of TrkB and E-cadherin were opposite in lung adenocarcinoma samples. | [68,69,70] |
| Ras | ↓ | A549, PC14, H322, H520, and H157 | Ras/ERK signaling activated CDCP1 to induce anoikis resistance. Ras/ERK pathway was regulated through CAV1. | [38,88] |
| βIII-tubulin | ↓ | H460 and A549 | High levels of βIII-tubulin enhanced phospho-AKT activity via PTEN suppression. | [76]. |
| RhoA | ↑ | BEAS-2B, LLC, H69, and H209 | RhoA counteracted FAK and was activated by p66Shc, causing anoikis. | [10,56] |
| RhoB | ↑ | NIH-3T3, A549 | RhoB was suppressed by Ras/PI3K/AKT. Suppression of RhoB resulted in anoikis resistance. | [65,73] |
| ROCK | ↑ | BEAS-2B | RhoA/ROCK activated MKK4/MKK7/JNK/BIM to promote apoptosis. | [10] |
| NO | ↓ | H460, H23, H292 | NO exposure enhanced EMT and upregulated CAV1, allowing escape from anoikis and promoting migration. | [89,90] |
| MCL-1 | ↓ | NIH3T3, H460 | Mcl-1 degradation activated anoikis. CAV1 interacted with MCL-1 to prevent MCL-1 from degradation via ubiquitination. | [92,93] |
| H2O2 | ↓ | H460, G361 | H2O2 inhibited the formation of the CAV1–ubiquitin complex. | [96,100] |
| miR-222-3p | ↓ | A549-GR | miR-222-3p directly targeted the promoter of SOCS3 to enhance gemcitabine resistance and anti-anoikis. | [103] |
| CAV1 | ↓ | H460 | CAV1, as a membrane adaptor to kinase Fyn, activated integrin signaling. CAV1 was negatively regulated by oxidative stress. | [88,90,96] |
| ZEB1 | ↓ | HBECs, HepG2, H358, H1155, H1299, and A549 | ZEB1 was a negative regulator of p66Shc. | [63] |
| Notch-1 | ↓ | PC-9 | Notch-1 enhanced EMT markers, including vimentin and snail. | [110] |
| FAIM2 | ↓ | HARA, HARA-B4, H1395, A549, and NIH3T3 | FAIM2 enhanced Wnt/β-catenin signaling pathway to facilitate EMT and anoikis resistance. | [111] |
| DAPK | ↑ | H460, A427, A549, and CL1-0 | CCN2 promoted DAPK kinase activity and then activated the p53 pathway. | [52] |
| p53 and XEDAR | ↑ | A549, H1299, and HeLa | p53 enhanced cleavage caspase 3 and AKT activity and promoted XEDAR expression to inhibit integrin/FAK signaling. | [44,115] |
| CRR9 | ↓ | A549, H838 | CRR9 interacted with PI3K to activate oncogene Ras. | [74] |
| Contactin 1 | ↓ | H446, H526 | Contactin 1 activated the AKT pathway to promote EMT phenotype. | [77] |
| MMP-7 | ↓ | BALB/c 3T3 | MMP-7 degraded IGFBP-3 and activated the AKT pathway, resulting in anoikis. | [79] |
| IL13Rα2 | ↓ | HBE-135, HBE-154, PC9, H1975, A549, HTB-57, H2170, H1299, H358, H3255, H1838, and HCC827 | IL13Rα2 activated PI3K/AKT and TAZ, resulting in migration, invasion, and anoikis resistance. | [80] |
| lncRNA VAL | ↓ | A549, HCC827, H1650, H596, H1975, H1299, H292, H2009, and H2030 | lncRNA VAL bound to vimentin and inhibited vimentin degradation. | [81] |
| Farnesylated AKT1 | ↓ | MCF10A and A549 | Farnesylated AKT1 suppressed anoikis and caused resistance to DNA-reactive agents, but not altered cell cycle (M-phase) specific chemotherapeutics. | [78] |
| BIM | ↑ | NIH3T3, A549, BEAS-2B | BIM induced endangerment of mitochondria and apoptosis. | [92,119] |
| 14-3-3ζ | ↓ | A549 | Deficiency of 14-3-3ζ upregulated BAD and BIM and decreased MCL-1 to increase BAX, cleaved caspase 7, and cleaved caspase 3 toward tumor cell anoikis. | [120] |
| Bit1 | ↑ | A549 | Bit1/AES complex initiated cell death complex and inhibited EMT through upregulation of E-cadherin. | [121,122] |
| TLE1 | ↓ | A549 | TLE1 interfered with AES formation Bit1/AES complex. | [122,124] |
| NQO1 | ↓ | A549, H292 | NQO1 decreased ROS formation and anoikis sensitization. | [137,138] |
| ΔNp63α | ↓ | HCC95 | ΔNp63α protected cells from oxidative stress, DNA damage, anoikis, and ferroptosis through upregulation of glutathione metabolism. | [139] |
| ILF2 | ↓ | HUVEC-C, HBEC-5i, BEAS-2B, A549, H460, H1155, and H1299 | ILF2 directly inhibited PTEN via binding to its upstream regulatory region. | [140] |
| SPIB | ↓ | A549, H1703, H1975, H446, H520, H226, SK-MES-1, H460, H1299 | Activated SPIB directly enhanced SNAP47 transcription in lung cancer cells and increased anoikis resistance. | [141] |
| Keratin 14 | ↓ | KrasG12D/Trp53L/L cell lines from de novo KP tumors | Gastrokine 1 cooperated with keratin 14, inhibited anoikis, and promoted cancer metastasis. | [142] |
| SCD1 | ↓ | H460 | Loss of SCD1 induced cellular damage and anoikis. | [143] |
| MSP | ↓ | SBC-5, H1048 | MSP phosphorylated RON to promote liver metastases in lung cancer. | [144] |
| KLF12 | ↑ | A549, H23, H460 | KLF12 promotes cell cycle transition through the S phase to induce anoikis. | [145] |
| TRIM62 | ↑ | Trim62+/− and Trim62−/− C57BL/6J mice | The loss of TRIM62 synergizes with K-Ras mutation, promoting EMT, tumorigenesis, and metastasis in lung cancer. | [146] |
| SH2B3 | ↑ | A549, NCI-H358, NCI-H1650, NCI-H460, NCI-H1688, Calu-1; A549 and NCI-H1688 tumor-bearing nude mouse | SH2B3 inhibits JAK2/STAT3 and PI3K/AKT signaling pathways to induce anoikis in lung cancer. | [147] |
4. Therapy for Anoikis in Lung Cancer
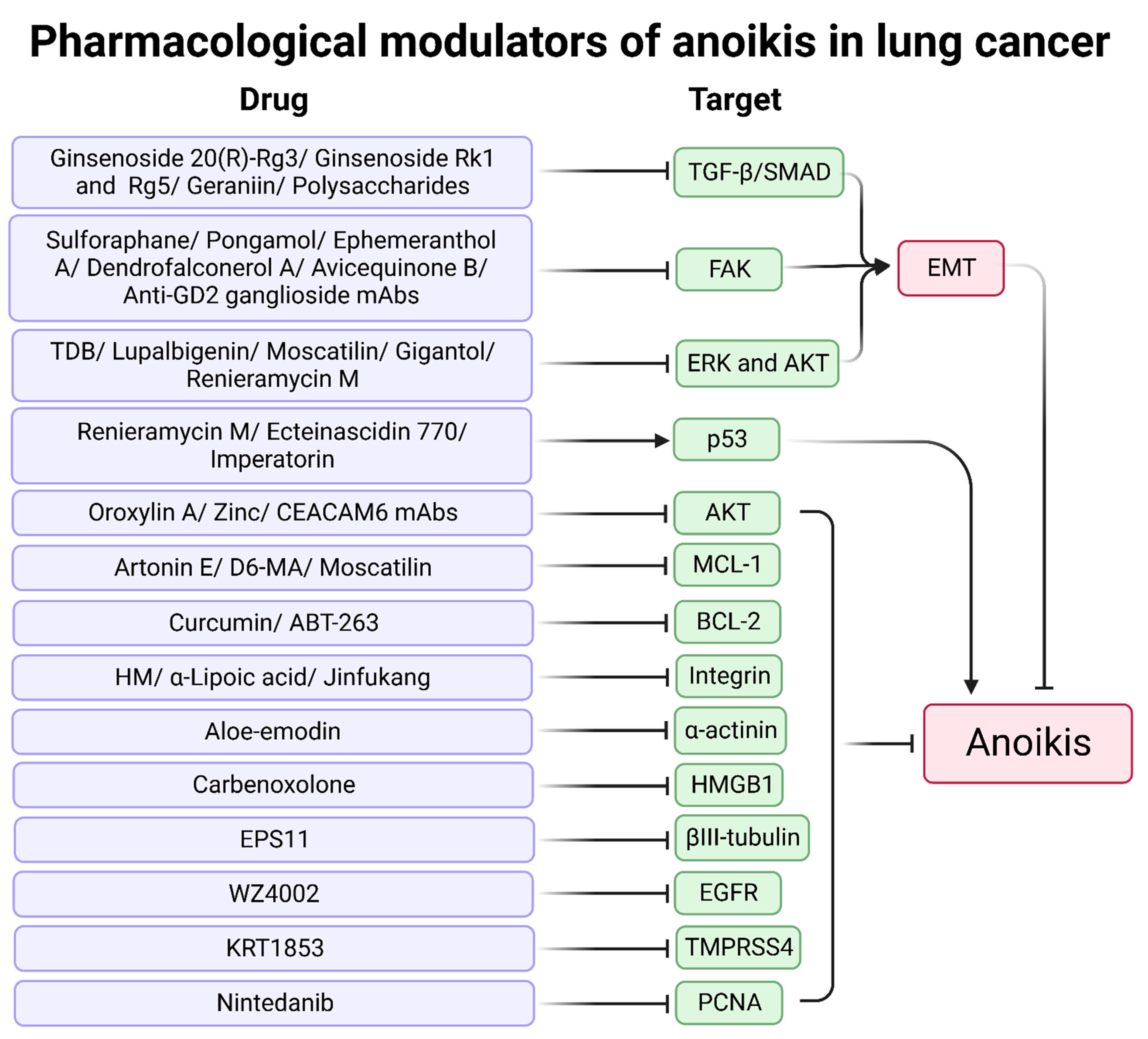
4.1. Natural Products
4.2. Synthetic Products
| Modulators | Tested Model(s) | Mechanism | Ref |
|---|---|---|---|
| Renieramycin M | H460 | p53 activation | [148] |
| Ecteinascidin 770 | H23, H460 | p53 activation | [149] |
| Imperatorin | H23, H292, and A549 | p53 activation and MCL-1 downregulation | [150] |
| Artonin E | H460, A549, and H292 | MCL-1 downregulation | [151] |
| Oroxylin A | A549 | Inhibitory effect of c-Src/AKT pathway | [152] |
| TDB | H292 | EMT inhibition via inactivation of phospho-ERK and phospho-AKT | [153] |
| Lupalbigenin | H460 | Phospho-AKT, phospho-ERK, and BCL-2 inhibition | [154] |
| Sulforaphane | A549 and CL1-5 | Reduction of FAK, AKT, and β-catenin and upregulation of p21 | [155] |
| Pongamol | H460 | Inhibition of EMT through FAK and AKT/mTOR signaling pathways | [156] |
| EPS11 | H460, A549, and H1299 A549 xenograft with BALB/c-nu mice | Inhibitory effect on βIII-tubulin | [157] |
| Ginsenoside 20(R)-Rg3 | A549 | EMT inhibition through the TGF-β/SMAD pathway | [158] |
| Ginsenosides Rk1 and Rg5 | A549 | Inhibition of TGF-β1-induced EMT | [159] |
| Jorunnamycin A | H460, H292, and H23 | Suppression of EMT and apoptosis-related protein | [160] |
| Moscatilin | H460 | EMT inhibition involves repression of ERK, AKT, MCL-1, and CAV1 | [161] |
| Gigantol | H460 | EMT inhibition involves suppression of ERK, AKT, and CAV1 | [162] |
| Ephemeranthol A | H460 | EMT inhibition via FAK/AKT pathway | [163] |
| Dendrofalconerol A | H460 | Suppression of phospho-FAK and Rho-GTP | [164] |
| Chrysotobibenzyl, chrysotoxine, crepidatin, and moscatilin | H23 | Anti-anoikis activities | [165] |
| 4,4′-Dihydroxy-3,5-dimethoxybibenzyl, 4,5,4′-trihydroxy-3,3′-dimethoxybibenzyl, chrysoeriol, and luteolin | H292 | Anti-anoikis activities | [166] |
| Aloe-emodin | H460 | Downregulation of α-actinin and MAPK pathway (JNK, ERK1, p38) and upregulation of PKCδ | [167,168] |
| Curcumin | H460 | Upregulation of the degradation of BCL-2 | [169] |
| Geraniin | A549 | EMT inhibition via TGF-β1 | [170] |
| Polysaccharide | A549 | Inhibition of TGF-β1-induced EMT | [171] |
| Oat avenanthramides | A549 | Suppression of EGFR | [172] |
| Jinfukang | H1975 | Suppression of integrin/Src pathway | [173] |
| Modified Bu-Fei decoction | A549 and LLC-bearing mice | Inhibition of ANGPTL4 expression through suppressing HIF-1α signaling | [174] |
| ABT-263 | LC-KJ, HCC827, H1650, and H1975 | A BCL-2 inhibitor enhanced Src inhibitors | [175] |
| WZ4002 | HCC827 and H1975 | A third-generation EGFR inhibitor | [176] |
| KRT1853 | H322 | TMPRSS4 serine protease inhibitors, repression of JNK/MAPK, PI3K/AKT, and NF-κB pathways | [177] |
| Nintedanib | A549, H1299, and H460 | Downregulation of PCNA | [178] |
| Renieramycin M | H460 | Downregulation of phospho-ERK, phospho-AKT, BCL-2, and MCL-1 | [179] |
| Avicequinone B | H460, H292, and H23 | Diminution of integrin/FAK/Src, MCL-1, and BCL-2 | [180] |
| Carbenoxolone | C57BL/6J mice injected with LLC | Inhibition of HMGB1 | [181] |
| D6-MA | H460 | MCL-1 downregulation | [182] |
| HM | H292 | Inhibition of integrin β3 | [183] |
| α-Lipoic acid | H460 | Integrin β1 and β3 inhibition | [184] |
| Zinc | H460 | Downregulation of AKT and CAV1 | [185] |
| Anti-GD2 ganglioside mAbs | NCI-417, ACC-LC-171, and ACC-LC-96 in vitro | FAK reduction and p38 activation | [186] |
| Anti-LN-332 mAbs | KLN-205 tumors in DBA/2 mice | Blocking cell–matrix interaction | [187] |
| CEACAM6 mAbs | Balb/C xenotransplanted A549 | Inhibitory effects on phospho-AKT and upregulation of paclitaxel chemosensitivity | [188,189] |
5. Discussion
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Fehlmann, T.; Kahraman, M.; Ludwig, N.; Backes, C.; Galata, V.; Keller, V.; Geffers, L.; Mercaldo, N.; Hornung, D.; Weis, T.; et al. Evaluating the Use of Circulating MicroRNA Profiles for Lung Cancer Detection in Symptomatic Patients. JAMA Oncol. 2020, 6, 714–723. [Google Scholar] [CrossRef] [PubMed]
- Sohn, M.; Shin, S.; Yoo, J.-Y.; Goh, Y.; Lee, I.H.; Bae, Y.S. Ahnak promotes tumor metastasis through transforming growth factor-β-mediated epithelial-mesenchymal transition. Sci. Rep. 2018, 8, 14379. [Google Scholar] [CrossRef] [PubMed]
- Talbot, L.J.; Bhattacharya, S.D.; Kuo, P.C. Epithelial-mesenchymal transition, the tumor microenvironment, and metastatic behavior of epithelial malignancies. Int. J. Biochem. Mol. Biol. 2012, 3, 117–136. [Google Scholar]
- Jiang, B.; Zhang, T.; Liu, F.; Sun, Z.; Shi, H.; Hua, D.; Yang, C. The co-stimulatory molecule B7-H3 promotes the epithelial-mesenchymal transition in colorectal cancer. Oncotarget 2016, 7, 31755–31771. [Google Scholar] [CrossRef]
- Kamińska, K.; Szczylik, C.; Bielecka, Z.F.; Bartnik, E.; Porta, C.; Lian, F.; Czarnecka, A.M. The role of the cell-cell interactions in cancer progression. J. Cell. Mol. Med. 2015, 19, 283–296. [Google Scholar] [CrossRef]
- Frisch, S.M.; Francis, H. Disruption of epithelial cell-matrix interactions induces apoptosis. J. Cell Biol. 1994, 124, 619–626. [Google Scholar] [CrossRef]
- Kockx, M.M.; Herman, A.G. Apoptosis in atherosclerosis: Beneficial or detrimental? Cardiovasc. Res. 2000, 45, 736–746. [Google Scholar] [CrossRef]
- Dobler, D.; Ahmed, N.; Song, L.; Eboigbodin, K.E.; Thornalley, P.J. Increased Dicarbonyl Metabolism in Endothelial Cells in Hyperglycemia Induces Anoikis and Impairs Angiogenesis by RGD and GFOGER Motif Modification. Diabetes 2006, 55, 1961–1969. [Google Scholar] [CrossRef]
- Haun, F.; Neumann, S.; Peintner, L.; Wieland, K.; Habicht, J.; Schwan, C.; Østevold, K.; Koczorowska, M.M.; Biniossek, M.; Kist, M.; et al. Identification of a novel anoikis signalling pathway using the fungal virulence factor gliotoxin. Nat. Commun. 2018, 9, 3524. [Google Scholar] [CrossRef]
- Simpson, C.D.; Anyiwe, K.; Schimmer, A.D. Anoikis resistance and tumor metastasis. Cancer Lett. 2008, 272, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Chaffer, C.L.; San Juan, B.P.; Lim, E.; Weinberg, R.A. EMT, cell plasticity and metastasis. Cancer Metastasis Rev. 2016, 35, 645–654. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Wang, Z.; Hackert, T.; Pitzer, C.; Zöller, M. Tspan8 and Tspan8/CD151 knockout mice unravel the contribution of tumor and host exosomes to tumor progression. J. Exp. Clin. Cancer Res. 2018, 37, 312. [Google Scholar] [CrossRef] [PubMed]
- Baethge, C.; Goldbeck-Wood, S.; Mertens, S. SANRA—A scale for the quality assessment of narrative review articles. Res. Integr. Peer Rev. 2019, 4, 5. [Google Scholar] [CrossRef]
- Theocharis, A.D.; Skandalis, S.S.; Gialeli, C.; Karamanos, N.K. Extracellular matrix structure. Adv. Drug Deliv. Rev. 2016, 97, 4–27. [Google Scholar] [CrossRef]
- Taddei, M.L.; Giannoni, E.; Fiaschi, T.; Chiarugi, P. Anoikis: An emerging hallmark in health and diseases. J. Pathol. 2012, 226, 380–393. [Google Scholar] [CrossRef]
- Pickup, M.W.; Mouw, J.K.; Weaver, V.M. The extracellular matrix modulates the hallmarks of cancer. EMBO Rep. 2014, 15, 1243–1253. [Google Scholar] [CrossRef]
- Enkhbat, M.; Zhong, B.; Chang, R.; Geng, J.; Lu, L.-S.; Chen, Y.-J.; Wang, P.-Y. Harnessing Focal Adhesions to Accelerate p53 Accumulation and Anoikis of A549 Cells Using Colloidal Self-Assembled Patterns (cSAPs). ACS Appl. Bio Mater. 2022, 5, 322–333. [Google Scholar] [CrossRef]
- Han, H.-J.; Sung, J.Y.; Kim, S.-H.; Yun, U.-J.; Kim, H.; Jang, E.-J.; Yoo, H.-E.; Hong, E.K.; Goh, S.-H.; Moon, A.; et al. Fibronectin regulates anoikis resistance via cell aggregate formation. Cancer Lett. 2021, 508, 59–72. [Google Scholar] [CrossRef]
- Kodama, K.; Ishii, G.; Miyamoto, S.; Goya, M.; Zhang, S.-C.; Sangai, T.; Yoshikawa, T.; Hasebe, T.; Hitomi, Y.; Izumi, K.; et al. Laminin 5 expression protects against anoikis at aerogenous spread and lepidic growth of human lung adenocarcinoma. Int. J. Cancer 2005, 116, 876–884. [Google Scholar] [CrossRef]
- Liu, C.-C.; Lin, S.-P.; Hsu, H.-S.; Yang, S.-H.; Lin, C.-H.; Yang, M.-H.; Hung, M.-C.; Hung, S.-C. Suspension survival mediated by PP2A-STAT3-Col XVII determines tumour initiation and metastasis in cancer stem cells. Nat. Commun. 2016, 7, 11798. [Google Scholar] [CrossRef] [PubMed]
- Burnier, J.V.; Wang, N.; Michel, R.P.; Hassanain, M.; Li, S.; Lu, Y.; Metrakos, P.; Antecka, E.; Burnier, M.N.; Ponton, A.; et al. Type IV collagen-initiated signals provide survival and growth cues required for liver metastasis. Oncogene 2011, 30, 3766–3783. [Google Scholar] [CrossRef] [PubMed]
- Jang, I.; Beningo, K. Integrins, CAFs and Mechanical Forces in the Progression of Cancer. Cancers 2019, 11, 721. [Google Scholar] [CrossRef] [PubMed]
- Hamidi, H.; Ivaska, J. Every step of the way: Integrins in cancer progression and metastasis. Nat. Rev. Cancer 2018, 18, 533–548. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.-I.; Lin, Y.-P.; Tseng, C.-W.; Chen, H.-J.; Wang, L.-H. Crabp2 Promotes Metastasis of Lung Cancer Cells via HuR and Integrin β1/FAK/ERK Signaling. Sci. Rep. 2019, 9, 845. [Google Scholar] [CrossRef] [PubMed]
- Winograd-Katz, S.E.; Fässler, R.; Geiger, B.; Legate, K.R. The integrin adhesome: From genes and proteins to human disease. Nat. Rev. Mol. Cell Biol. 2014, 15, 273–288. [Google Scholar] [CrossRef]
- Wang, C.; Wang, T.; Lv, D.; Li, L.; Yue, J.; Chen, H.-Z.; Xu, L. Acquired Resistance to EGFR TKIs Mediated by TGFβ1/Integrin β3 Signaling in EGFR-Mutant Lung Cancer. Mol. Cancer Ther. 2019, 18, 2357–2367. [Google Scholar] [CrossRef]
- Ordonez, C.; Zhai, A.B.; Camacho-Leal, P.; Demarte, L.; Fan, M.M.; Stanners, C.P. GPI-anchored CEA family glycoproteins CEA and CEACAM6 mediate their biological effects through enhanced integrin α5β1-fibronectin interaction. J. Cell. Physiol. 2007, 210, 757–765. [Google Scholar] [CrossRef]
- Stanners, C.P. Cell Adhesion and Communication Mediated by the CEA Family; CRC Press: London, UK, 1998. [Google Scholar] [CrossRef]
- Camacho-Leal, P.; Zhai, A.B.; Stanners, C.P. A co-clustering model involving α5β1 integrin for the biological effects of GPI-anchored human carcinoembryonic antigen (CEA). J. Cell. Physiol. 2007, 211, 791–802. [Google Scholar] [CrossRef]
- Camacholeal, P.; Stanners, C.P. The human carcinoembryonic antigen (CEA) GPI anchor mediates anoikis inhibition by inactivation of the intrinsic death pathway. Oncogene 2008, 27, 1545–1553. [Google Scholar] [CrossRef]
- Kim, E.Y.; Cha, Y.J.; Jeong, S.; Chang, Y.S. Overexpression of CEACAM6 activates Src-FAK signaling and inhibits anoikis, through homophilic interactions in lung adenocarcinomas. Transl. Oncol. 2022, 20, 101402. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.-Y.; Hsu, H.-S.; Tyan, S.-W.; Li, F.-Y.; Shew, J.-Y.; Lee, W.-H.; Chen, J.-Y. Serglycin in tumor microenvironment promotes non-small cell lung cancer aggressiveness in a CD44-dependent manner. Oncogene 2017, 36, 2457–2471. [Google Scholar] [CrossRef]
- Shimobaba, S.; Taga, S.; Akizuki, R.; Hichino, A.; Endo, S.; Matsunaga, T.; Watanabe, R.; Yamaguchi, M.; Yamazaki, Y.; Sugatani, J.; et al. Claudin-18 inhibits cell proliferation and motility mediated by inhibition of phosphorylation of PDK1 and Akt in human lung adenocarcinoma A549 cells. Biochim. Biophys. Acta 2016, 1863, 1170–1178. [Google Scholar] [CrossRef] [PubMed]
- Runkle, E.A.; Rice, S.J.; Qi, J.; Masser, D.; Antonetti, D.A.; Winslow, M.M.; Mu, D. Occludin Is a Direct Target of Thyroid Transcription Factor-1 (TTF-1/NKX2–1). J. Biol. Chem. 2012, 287, 28790–28801. [Google Scholar] [CrossRef] [PubMed]
- Uekita, T.; Jia, L.; Narisawa-Saito, M.; Yokota, J.; Kiyono, T.; Sakai, R. CUB Domain-Containing Protein 1 Is a Novel Regulator of Anoikis Resistance in Lung Adenocarcinoma. Mol. Cell. Biol. 2007, 27, 7649–7660. [Google Scholar] [CrossRef]
- Miyazawa, Y.; Uekita, T.; Hiraoka, N.; Fujii, S.; Kosuge, T.; Kanai, Y.; Nojima, Y.; Sakai, R. CUB Domain–Containing Protein 1, a Prognostic Factor for Human Pancreatic Cancers, Promotes Cell Migration and Extracellular Matrix Degradation. Cancer Res. 2010, 70, 5136–5146. [Google Scholar] [CrossRef]
- Uekita, T.; Fujii, S.; Miyazawa, Y.; Iwakawa, R.; Narisawa-Saito, M.; Nakashima, K.; Tsuta, K.; Tsuda, H.; Kiyono, T.; Yokota, J.; et al. Oncogenic Ras/ERK Signaling Activates CDCP1 to Promote Tumor Invasion and Metastasis. Mol. Cancer Res. 2014, 12, 1449–1459. [Google Scholar] [CrossRef]
- Uekita, T.; Fujii, S.; Miyazawa, Y.; Hashiguchi, A.; Abe, H.; Sakamoto, M.; Sakai, R. Suppression of autophagy by CUB domain-containing protein 1 signaling is essential for anchorage-independent survival of lung cancer cells. Cancer Sci. 2013, 104, 865–870. [Google Scholar] [CrossRef]
- Schaller, M.D. FAK and paxillin: Regulators of N-cadherin adhesion and inhibitors of cell migration? J. Cell Biol. 2004, 166, 157–159. [Google Scholar] [CrossRef]
- Alanko, J.; Ivaska, J. Endosomes: Emerging Platforms for Integrin-Mediated FAK Signalling. Trends Cell Biol. 2016, 26, 391–398. [Google Scholar] [CrossRef]
- Liu, G.; Meng, X.; Jin, Y.; Bai, J.; Zhao, Y.; Cui, X.; Chen, F.; Fu, S. Inhibitory role of focal adhesion kinase on anoikis in the lung cancer cell A549. Cell Biol. Int. 2008, 32, 663–670. [Google Scholar] [CrossRef] [PubMed]
- Keenan, J.; Joyce, H.; Aherne, S.; O’Dea, S.; Doolan, P.; Lynch, V.; Clynes, M. Olfactomedin III expression contributes to anoikis-resistance in clonal variants of a human lung squamous carcinoma cell line. Exp. Cell Res. 2012, 318, 593–602. [Google Scholar] [CrossRef][Green Version]
- Tanikawa, C.; Furukawa, Y.; Yoshida, N.; Arakawa, H.; Nakamura, Y.; Matsuda, K. XEDAR as a putative colorectal tumor suppressor that mediates p53-regulated anoikis pathway. Oncogene 2009, 28, 3081–3092. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.M.; Brugge, J.S. Cellular functions regulated by Src family kinases. Annu. Rev. Cell Dev. Biol. 1997, 13, 513–609. [Google Scholar] [CrossRef] [PubMed]
- Schlessinger, J. New Roles for Src Kinases in Control of Cell Survival and Angiogenesis. Cell 2000, 100, 293–296. [Google Scholar] [CrossRef]
- Sakuma, Y.; Takeuchi, T.; Nakamura, Y.; Yoshihara, M.; Matsukuma, S.; Nakayama, H.; Ohgane, N.; Yokose, T.; Kameda, Y.; Tsuchiya, E.; et al. Lung adenocarcinoma cells floating in lymphatic vessels resist anoikis by expressing phosphorylated Src. J. Pathol. 2009, 220, 574–585. [Google Scholar] [CrossRef]
- Wei, L.; Yang, Y.; Zhang, X.; Yu, Q. Altered regulation of Src upon cell detachment protects human lung adenocarcinoma cells from anoikis. Oncogene 2004, 23, 9052–9061. [Google Scholar] [CrossRef]
- Wei, L.; Yang, Y.; Zhang, X.; Yu, Q. Anchorage-independent phosphorylation of p130Cas protects lung adenocarcinoma cells from anoikis. J. Cell. Biochem. 2002, 87, 439–449. [Google Scholar] [CrossRef]
- Liu, A.; Xie, H.; Li, R.; Ren, L.; Yang, B.; Dai, L.; Lu, W.; Liu, B.; Ren, D.; Zhang, X.; et al. Silencing ZIC2 abrogates tumorigenesis and anoikis resistance of non-small cell lung cancer cells by inhibiting Src/FAK signaling. Mol. Ther. Oncolytics 2021, 22, 195–208. [Google Scholar] [CrossRef]
- Kim, H.; Sung, J.Y.; Park, E.-K.; Kho, S.; Koo, K.H.; Park, S.-Y.; Goh, S.-H.; Jeon, Y.K.; Oh, S.; Park, B.-K.; et al. Regulation of anoikis resistance by NADPH oxidase 4 and epidermal growth factor receptor. Br. J. Cancer 2017, 116, 370–381. [Google Scholar] [CrossRef]
- Jang, E.-J.; Sung, J.Y.; Yoo, H.-E.; Jang, H.; Shim, J.; Oh, E.-S.; Goh, S.-H.; Kim, Y.-N. FAM188B Downregulation Sensitizes Lung Cancer Cells to Anoikis via EGFR Downregulation and Inhibits Tumor Metastasis In Vivo. Cancers 2021, 13, 247. [Google Scholar] [CrossRef] [PubMed]
- Debnath, J. p66(Shc) and Ras: Controlling anoikis from the inside-out. Oncogene 2010, 29, 5556–5558. [Google Scholar] [CrossRef] [PubMed]
- Frisch, S.M.; Schaller, M.D. The Wind God Promotes Lung Cancer. Cancer Cell 2014, 25, 551–552. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Myers, D.P.; Wu, R.F.; Nwariaku, F.E.; Terada, L.S. p66Shc mediates anoikis through RhoA. J. Cell Biol. 2007, 179, 23–31. [Google Scholar] [CrossRef]
- Ma, Z.; Liu, Z.; Wu, R.-F.; Terada, L.S. p66Shc restrains Ras hyperactivation and suppresses metastatic behavior. Oncogene 2010, 29, 5559–5567. [Google Scholar] [CrossRef]
- Zheng, Z.; Yang, J.; Zhao, D.; Gao, D.; Yan, X.; Yao, Z.; Liu, Z.; Ma, Z. Downregulated adaptor protein p66Shcmitigates autophagy process by low nutrient and enhances apoptotic resistance in human lung adenocarcinoma A549 cells. FEBS J. 2013, 280, 4522–4530. [Google Scholar] [CrossRef]
- Cai, Z.; Zhao, D.; Sun, Y.; Gao, D.; Li, X.; Yang, J.; Ma, Z. Detachment-Based Equilibrium of Anoikic Cell Death and Autophagic Cell Survival Through Adaptor Protein p66Shc. Anat. Rec. 2018, 299, 325–333. [Google Scholar] [CrossRef]
- Du, W.; Jiang, Y.; Zheng, Z.; Zhang, Z.; Chen, N.; Ma, Z.; Yao, Z.; Terada, L.; Liu, Z. Feedback loop between p66Shc and Nrf2 promotes lung cancer progression. Cancer Lett. 2013, 337, 58–65. [Google Scholar] [CrossRef]
- Billot, K.; Soeur, J.; Chereau, F.; Arrouss, I.; Merle-Beral, H.; Huang, M.-E.; Mazier, D.; Baud, V.; Rebollo, A. Deregulation of Aiolos expression in chronic lymphocytic leukemia is associated with epigenetic modifications. Blood 2011, 117, 1917–1927. [Google Scholar] [CrossRef]
- Li, X.; Xu, Z.; Du, W.; Zhang, Z.; Wei, Y.; Wang, H.; Zhu, Z.; Qin, L.; Wang, L.; Niu, Q.; et al. Aiolos Promotes Anchorage Independence by Silencing p66Shc Transcription in Cancer Cells. Cancer Cell 2014, 25, 575–589. [Google Scholar] [CrossRef]
- Zhu, Z.; Wang, H.; Wei, Y.; Meng, F.; Liu, Z.; Zhang, Z. Downregulation of PRDM1 promotes cellular invasion and lung cancer metastasis. Tumor Biol. 2017, 39, 1010428317695929. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Gao, D.; Wang, H.; Yang, J.; Yan, X.; Liu, Z.; Ma, Z. Negative feedback loop between p66Shc and ZEB1 regulates fibrotic EMT response in lung cancer cells. Cell Death Dis. 2015, 6, e1708. [Google Scholar] [CrossRef] [PubMed]
- Reginato, M.J.; Mills, K.R.; Paulus, J.K.; Lynch, D.K.; Sgroi, D.C.; Debnath, J.; Muthuswamy, S.K.; Brugge, J.S. Integrins and EGFR coordinately regulate the pro-apoptotic protein Bim to prevent anoikis. Nat. Cell Biol. 2003, 5, 733–740. [Google Scholar] [CrossRef] [PubMed]
- Jiang, K.; Sun, J.; Cheng, J.; Djeu, J.Y.; Wei, S.; Sebti, S. Akt Mediates Ras Downregulation of RhoB, a Suppressor of Transformation, Invasion, and Metastasis. Mol. Cell. Biol. 2004, 24, 5565–5576. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.-C.; Yang, M.-H.; Lin, B.-R.; Chen, S.-T.; Pan, S.-H.; Hsiao, M.; Lai, T.-C.; Lin, S.-K.; Jeng, Y.-M.; Chu, C.-Y.; et al. CCN2 inhibits lung cancer metastasis through promoting DAPK-dependent anoikis and inducing EGFR degradation. Cell Death Differ. 2013, 20, 443–455. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Miłoszewska, J.; Przybyszewska, M.; Gos, M.; Swoboda, P.; Trembacz, H. TrkB expression level correlates with metastatic properties of L1 mouse sarcoma cells cultured in non-adhesive conditions. Cell Prolif. 2013, 46, 146–152. [Google Scholar] [CrossRef]
- Smit, M.A.; Geiger, T.R.; Song, J.-Y.; Gitelman, I.; Peeper, D.S. A Twist-Snail Axis Critical for TrkB-Induced Epithelial-Mesenchymal Transition-Like Transformation, Anoikis Resistance, and Metastasis. Mol. Cell. Biol. 2009, 29, 3722–3737. [Google Scholar] [CrossRef]
- Ricci, A.; De Vitis, C.; Noto, A.; Fattore, L.; Mariotta, S.; Cherubini, E.; Roscilli, G.; Liguori, G.; Scognamiglio, G.; Rocco, G.; et al. TrkB is responsible for EMT transition in malignant pleural effusions derived cultures from adenocarcinoma of the lung. Cell Cycle 2013, 12, 1696–1703. [Google Scholar] [CrossRef][Green Version]
- Geiger, T.R.; Song, J.-Y.; Rosado, A.; Peeper, D.S. Functional Characterization of Human Cancer-Derived TRKB Mutations. PLoS ONE 2011, 6, e16871. [Google Scholar] [CrossRef]
- Voisin, L.; Julien, C.; Duhamel, S.; Gopalbhai, K.; Claveau, I.; Saba-El-Leil, M.K.; Rodrigue-Gervais, I.G.; Gaboury, L.; Lamarre, D.; Basik, M.; et al. Activation of MEK1 or MEK2 isoform is sufficient to fully transform intestinal epithelial cells and induce the formation of metastatic tumors. BMC Cancer 2008, 8, 337. [Google Scholar] [CrossRef]
- Pearson, G.; Robinson, F.; Beers Gibson, T.; Xu, B.E.; Karandikar, M.; Berman, K.; Cobb, M.H. Mitogen-Activated Protein (MAP) Kinase Pathways: Regulation and Physiological Functions. Endocr. Rev. 2001, 22, 153–183. [Google Scholar] [CrossRef] [PubMed]
- Jiang, K.; Delarue, F.L.; Sebti, S.M. EGFR, ErbB2 and Ras but not Src suppress RhoB expression while ectopic expression of RhoB antagonizes oncogene-mediated transformation. Oncogene 2004, 23, 1136–1145. [Google Scholar] [CrossRef] [PubMed]
- James, M.A.; Vikis, H.G.; Tate, E.; Rymaszewski, A.L.; You, M. CRR9/CLPTM1L Regulates Cell Survival Signaling and Is Required for Ras Transformation and Lung Tumorigenesis. Cancer Res. 2014, 74, 1116–1127. [Google Scholar] [CrossRef]
- Wei, L.; Yang, Y.; Yu, Q. Tyrosine kinase-dependent, phosphatidylinositol 3′-kinase, and mitogen-activated protein ki-nase-independent signaling pathways prevent lung adenocarcinoma cells from anoikis. Cancer Res. 2001, 61, 2439–2444. [Google Scholar] [PubMed]
- McCarroll, J.A.; Gan, P.P.; Erlich, R.B.; Liu, M.; Dwarte, T.; Sagnella, S.S.; Akerfeldt, M.C.; Yang, L.; Parker, A.L.; Chang, M.H.; et al. TUBB3/βIII-Tubulin Acts through the PTEN/AKT Signaling Axis to Promote Tumorigenesis and Anoikis Resistance in Non–Small Cell Lung Cancer. Cancer Res. 2015, 75, 415–425. [Google Scholar] [CrossRef]
- Xu, S.; Lam, S.-K.; Cheng, P.N.-M.; Ho, J.C.-M. Contactin 1 modulates pegylated arginase resistance in small cell lung cancer through induction of epithelial-mesenchymal transition. Sci. Rep. 2019, 9, 12030. [Google Scholar] [CrossRef]
- Schmidt, M.; Hövelmann, S.; Beckers, T.L. A novel form of constitutively active farnesylated Akt1 prevents mammary epithelial cells from anoikis and suppresses chemotherapy-induced apoptosis. Br. J. Cancer 2002, 87, 924–932. [Google Scholar] [CrossRef]
- Miyamoto, S.; Yano, K.; Sugimoto, S.; Ishii, G.; Hasebe, T.; Endoh, Y.; Kodama, K.; Goya, M.; Chiba, T.; Ochiai, A. Matrix Metalloproteinase-7 Facilitates Insulin-Like Growth Factor Bioavailability through Its Proteinase Activity on Insulin-Like Growth Factor Binding Protein 3. Cancer Res. 2004, 64, 665–671. [Google Scholar] [CrossRef]
- Xie, M.; Wu, X.-J.; Zhang, J.-J.; He, C.-S. IL-13 receptor α2 is a negative prognostic factor in human lung cancer and stimulates lung cancer growth in mice. Oncotarget 2015, 6, 32902–32913. [Google Scholar] [CrossRef]
- Tian, H.; Lian, R.; Li, Y.; Liu, C.; Liang, S.; Li, W.; Tao, T.; Wu, X.; Ye, Y.; Yang, X.; et al. AKT-induced lncRNA VAL promotes EMT-independent metastasis through diminishing Trim16-dependent Vimentin degradation. Nat. Commun. 2020, 11, 5127. [Google Scholar] [CrossRef]
- Jordan, M.A.; Wilson, L. Microtubules as a target for anticancer drugs. Nat. Rev. Cancer 2004, 4, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Heasman, S.J.; Ridley, A.J. Mammalian Rho GTPases: New insights into their functions from in vivo studies. Nat. Rev. Mol. Cell Biol. 2008, 9, 690–701. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.D.; Kiosses, W.B.; Schwartz, M.A. Regulation of the small GTP-binding protein Rho by cell adhesion and the cytoskeleton. EMBO J. 1999, 18, 578–585. [Google Scholar] [CrossRef]
- Riento, K.; Ridley, A.J. ROCKs: Multifunctional kinases in cell behaviour. Nat. Rev. Mol. Cell Biol. 2003, 4, 446–456. [Google Scholar] [CrossRef] [PubMed]
- Sargiacomo, M.; Scherer, P.E.; Tang, Z.; Kübler, E.; Song, K.S.; Sanders, M.C.; Lisanti, M.P. Oligomeric structure of caveolin: Implications for caveolae membrane organization. Proc. Natl. Acad. Sci. USA 1995, 92, 9407–9411. [Google Scholar] [CrossRef]
- Grande-García, A.; Echarri, A.; de Rooij, J.; Alderson, N.B.; Waterman-Storer, C.M.; Valdivielso, J.M.; del Pozo, M.A. Caveolin-1 regulates cell polarization and directional migration through Src kinase and Rho GTPases. J. Cell Biol. 2007, 177, 683–694. [Google Scholar] [CrossRef]
- Wary, K.K.; Mariotti, A.; Zurzolo, C.; Giancotti, F.G. A Requirement for Caveolin-1 and Associated Kinase Fyn in Integrin Signaling and Anchorage-Dependent Cell Growth. Cell 1998, 94, 625–634. [Google Scholar] [CrossRef]
- Chanvorachote, P.; Nimmannit, U.; Lu, Y.; Talbott, S.; Jiang, B.-H.; Rojanasakul, Y. Nitric Oxide Regulates Lung Carcinoma Cell Anoikis through Inhibition of Ubiquitin-Proteasomal Degradation of Caveolin-1. J. Biol. Chem. 2009, 284, 28476–28484. [Google Scholar] [CrossRef]
- Chanvorachote, P.; Pongrakhananon, V.; Chunhacha, P. Prolonged Nitric Oxide Exposure Enhances Anoikis Resistance and Migration through Epithelial-Mesenchymal Transition and Caveolin-1 Upregulation. BioMed Res. Int. 2014, 2014, 941359. [Google Scholar] [CrossRef]
- Boisvert-Adamo, K.; Longmate, W.; Abel, E.V.; Aplin, A.E. Mcl-1 Is Required for Melanoma Cell Resistance to Anoikis. Mol. Cancer Res. 2009, 7, 549–556. [Google Scholar] [CrossRef]
- Woods, N.T.; Yamaguchi, H.; Lee, F.Y.; Bhalla, K.N.; Wang, H.-G. Anoikis, Initiated by Mcl-1 Degradation and Bim Induction, Is Deregulated during Oncogenesis. Cancer Res. 2007, 67, 10744–10752. [Google Scholar] [CrossRef] [PubMed]
- Chunhacha, P.; Pongrakhananon, V.; Rojanasakul, Y.; Chanvorachote, P. Caveolin-1 regulates Mcl-1 stability and anoikis in lung carcinoma cells. Am. J. Physiol. Physiol. 2012, 302, C1284–C1292. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Guan, L.; Li, S.; Jiang, Y.; Xiong, N.; Li, L.; Wu, C.; Zeng, H.; Liu, Y. Mechanosensitive caveolin-1 activation-induced PI3K/Akt/mTOR signaling pathway promotes breast cancer motility, invadopodia formation and metastasis in vivo. Oncotarget 2016, 7, 16227–16247. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Chen, Y.; Zhang, Y.; Jiang, X.; Jiang, Y.; Qin, X.; Yang, H.; Wu, C.; Liu, Y. Shear stress promotes anoikis resistance of cancer cells via caveolin-1-dependent extrinsic and intrinsic apoptotic pathways. J. Cell. Physiol. 2019, 234, 3730–3743. [Google Scholar] [CrossRef] [PubMed]
- Rungtabnapa, P.; Nimmannit, U.; Halim, H.; Rojanasakul, Y.; Chanvorachote, P. Hydrogen peroxide inhibits non-small cell lung cancer cell anoikis through the inhibition of caveolin-1 degradation. Am. J. Physiol. Physiol. 2011, 300, C235–C245. [Google Scholar] [CrossRef] [PubMed]
- Halim, H.; Luanpitpong, S.; Chanvorachote, P. Acquisition of anoikis resistance up-regulates caveolin-1 expression in human non-small cell lung cancer cells. Anticancer Res. 2012, 32, 1649–1658. [Google Scholar] [PubMed]
- Chanvorachote, P.; Pongrakhananon, V.; Halim, H. Caveolin-1 regulates metastatic behaviors of anoikis resistant lung cancer cells. Mol. Cell. Biochem. 2015, 399, 291–302. [Google Scholar] [CrossRef] [PubMed]
- Songserm, T.; Pongrakhananon, V.; Chanvorachote, P. Sub-toxic cisplatin mediates anoikis resistance through hydrogen peroxide-induced caveolin-1 up-regulation in non-small cell lung cancer cells. Anticancer Res. 2012, 32, 1659–1669. [Google Scholar]
- Halim, H.; Chanvorachote, P. Long-term hydrogen peroxide exposure potentiates anoikis resistance and anchorage-independent growth in lung carcinoma cells. Cell Biol. Int. 2012, 36, 1055–1066. [Google Scholar] [CrossRef]
- Lloyd, P.G.; Hardin, C.D. Caveolae in cancer: Two sides of the same coin? Focus on “Hydrogen peroxide inhibits non-small cell lung cancer cell anoikis through the inhibition of caveolin-1 degradation”. Am. J. Physiol. Physiol. 2011, 300, C232–C234. [Google Scholar] [CrossRef]
- Guo, X.; Wang, Z.; Sun, Q.; Sun, C.; Hua, H.; Huang, Q. The inhibitory effect of microRNA-1827 on anoikis resistance in lung adenocarcinoma A549 cells via targeting caveolin-1. Acta Biochim. Biophys. Sin. 2020, 52, 1148–1155. [Google Scholar] [CrossRef] [PubMed]
- Wei, F.; Ma, C.; Zhou, T.; Dong, X.; Luo, Q.; Geng, L.; Ding, L.; Zhang, Y.; Zhang, L.; Li, N.; et al. Exosomes derived from gemcitabine-resistant cells transfer malignant phenotypic traits via delivery of miRNA-222-3p. Mol. Cancer 2017, 16, 132. [Google Scholar] [CrossRef] [PubMed]
- Thiery, J.P.; Acloque, H.; Huang, R.Y.J.; Nieto, M.A. Epithelial-Mesenchymal Transitions in Development and Disease. Cell 2009, 139, 871–890. [Google Scholar] [CrossRef] [PubMed]
- Powan, P.; Luanpitpong, S.; He, X.; Rojanasakul, Y.; Chanvorachote, P. Detachment-induced E-cadherin expression promotes 3D tumor spheroid formation but inhibits tumor formation and metastasis of lung cancer cells. Am. J. Physiol. Physiol. 2017, 313, C556–C566. [Google Scholar] [CrossRef] [PubMed]
- Frisch, S.M.; Schaller, M.; Cieply, B. Mechanisms that link the oncogenic epithelial–mesenchymal transition to suppression of anoikis. J. Cell Sci. 2013, 126, 21–29. [Google Scholar] [CrossRef]
- Denlinger, C.E.; Ikonomidis, J.S.; Reed, C.E.; Spinale, F.G. Epithelial to mesenchymal transition: The doorway to metastasis in human lung cancers. J. Thorac. Cardiovasc. Surg. 2010, 140, 505–513. [Google Scholar] [CrossRef]
- Adeshakin, F.O.; Adeshakin, A.O.; Afolabi, L.O.; Yan, D.; Zhang, G.; Wan, X. Mechanisms for Modulating Anoikis Resistance in Cancer and the Relevance of Metabolic Reprogramming. Front. Oncol. 2021, 11, 626577. [Google Scholar] [CrossRef]
- Salvi, A.; Thanabalu, T. WIP promotes in-vitro invasion ability, anchorage independent growth and EMT progression of A549 lung adenocarcinoma cells by regulating RhoA levels. Biochem. Biophys. Res. Commun. 2017, 482, 1353–1359. [Google Scholar] [CrossRef]
- Xie, M.; Zhang, L.; He, C.-S.; Xu, F.; Liu, J.-L.; Hu, Z.-H.; Zhao, L.-P.; Tian, Y. Activation of Notch-1 enhances epithelial-mesenchymal transition in gefitinib-acquired resistant lung cancer cells. J. Cell. Biochem. 2012, 113, 1501–1513. [Google Scholar] [CrossRef]
- She, K.; Yang, W.; Li, M.; Xiong, W.; Zhou, M. FAIM2 Promotes Non-Small Cell Lung Cancer Cell Growth and Bone Metastasis by Activating the Wnt/β-Catenin Pathway. Front. Oncol. 2021, 11, 690142. [Google Scholar] [CrossRef]
- Bialik, S.; Kimchi, A. The Death-Associated Protein Kinases: Structure, Function, and Beyond. Annu. Rev. Biochem. 2006, 75, 189–210. [Google Scholar] [CrossRef] [PubMed]
- Kwon, T.; Youn, H.; Son, B.; Kim, D.; Seong, K.M.; Park, S.; Kim, W.; Youn, B. DANGER is involved in high glucose-induced radioresistance through inhibiting DAPK-mediated anoikis in non-small cell lung cancer. Oncotarget 2016, 7, 7193–7206. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Liu, P.; Wang, Z.C.; Zou, L.; Santiago, S.; Garbitt, V.; Gjoerup, O.V.; Iglehart, J.D.; Miron, A.; Richardson, A.L.; et al. SIK1 Couples LKB1 to p53-Dependent Anoikis and Suppresses Metastasis. Sci. Signal. 2009, 2, ra35. [Google Scholar] [CrossRef]
- Bachelder, R.E.; Ribick, M.J.; Marchetti, A.; Falcioni, R.; Soddu, S.; Davis, K.R.; Mercurio, A.M. P53 Inhibits α6β4 Integrin Survival Signaling by Promoting the Caspase 3–Dependent Cleavage of Akt/PKB. J. Cell Biol. 1999, 147, 1063–1072. [Google Scholar] [CrossRef]
- Marcus, A.I.; Zhou, W. LKB1 Regulated Pathways in Lung Cancer Invasion and Metastasis. J. Thorac. Oncol. 2010, 5, 1883–1886. [Google Scholar] [CrossRef]
- Ji, H.; Ramsey, M.R.; Hayes, D.N.; Fan, C.; McNamara, K.; Kozlowski, P.; Torrice, C.; Wu, M.C.; Shimamura, T.; Perera, S.A.; et al. LKB1 modulates lung cancer differentiation and metastasis. Nature 2007, 448, 807–810. [Google Scholar] [CrossRef]
- Platt, R.J.; Chen, S.; Zhou, Y.; Yim, M.J.; Swiech, L.; Kempton, H.R.; Dahlman, J.E.; Parnas, O.; Eisenhaure, T.M.; Jovanovic, M.; et al. CRISPR-Cas9 Knockin Mice for Genome Editing and Cancer Modeling. Cell 2014, 159, 440–455. [Google Scholar] [CrossRef]
- Akiyama, T.; Dass, C.R.; Choong, P.F. Bim-targeted cancer therapy: A link between drug action and underlying molecular changes. Mol. Cancer Ther. 2009, 8, 3173–3180. [Google Scholar] [CrossRef]
- Li, Z.; Zhao, J.; Du, Y.; Park, H.R.; Sun, S.-Y.; Bernal-Mizrachi, L.; Aitken, A.; Khuri, F.R.; Fu, H. Down-regulation of 14-3-3ζ suppresses anchorage-independent growth of lung cancer cells through anoikis activation. Proc. Natl. Acad. Sci. USA 2008, 105, 162–167. [Google Scholar] [CrossRef]
- Yao, X.; Pham, T.; Temple, B.; Gray, S.; Cannon, C.; Chen, R.; Abdel-Mageed, A.B.; Biliran, H. The Anoikis Effector Bit1 Inhibits EMT through Attenuation of TLE1-Mediated Repression of E-Cadherin in Lung Cancer Cells. PLoS ONE 2016, 11, e0163228. [Google Scholar] [CrossRef]
- Yao, X.; Gray, S.; Pham, T.; Delgardo, M.; Nguyen, A.; Do, S.; Ireland, S.K.; Chen, R.; Abdel-Mageed, A.B.; Biliran, H. Downregulation of Bit1 expression promotes growth, anoikis resistance, and transformation of immortalized human bronchial epithelial cells via Erk activation-dependent suppression of E-cadherin. Biochem. Biophys. Res. Commun. 2018, 495, 1240–1248. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Jennings, S.; Ireland, S.K.; Pham, T.; Temple, B.; Davis, M.; Chen, R.; Davenport, I.; Biliran, H. The Anoikis Effector Bit1 Displays Tumor Suppressive Function in Lung Cancer Cells. PLoS ONE 2014, 9, e101564. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Pham, T.; Temple, B.; Gray, S.; Cannon, C.; Hardy, C.; Fletcher, K.; Ireland, S.K.; Hossain, A.; Chen, R.; et al. TLE1 inhibits anoikis and promotes tumorigenicity in human lung cancer cells through ZEB1-mediated E-cadherin repression. Oncotarget 2017, 8, 72235–72249. [Google Scholar] [CrossRef] [PubMed]
- Brunquell, C.; Biliran, H.; Jennings, S.; Ireland, S.K.; Chen, R.; Ruoslahti, E. TLE1 Is an Anoikis Regulator and Is Downregulated by Bit1 in Breast Cancer Cells. Mol. Cancer Res. 2012, 10, 1482–1495. [Google Scholar] [CrossRef] [PubMed]
- Shaw, R.J. Tumor Suppression by LKB1: SIK-ness Prevents Metastasis. Sci. Signal. 2009, 2, pe55. [Google Scholar] [CrossRef] [PubMed]
- Mack, H.I.; Munger, K. The LKB1 tumor suppressor differentially affects anchorage independent growth of HPV positive cervical cancer cell lines. Virology 2013, 446, 9–16. [Google Scholar] [CrossRef][Green Version]
- Jin, L.; Chun, J.; Pan, C.; Kumar, A.; Zhang, G.; Ha, Y.; Li, D.; Alesi, G.N.; Kang, Y.; Zhou, L.; et al. The PLAG1-GDH1 Axis Promotes Anoikis Resistance and Tumor Metastasis through CamKK2-AMPK Signaling in LKB1-Deficient Lung Cancer. Mol. Cell 2018, 69, 87–99.e7. [Google Scholar] [CrossRef]
- Glutaminolysis Drives Lung Cancer Metastasis via the PLAG1–GDH1 Axis. Cancer Discov. 2018, 8, 135. [CrossRef]
- Coloff, J.L. Glutamate Dehydrogenase to the Rescue. Mol. Cell 2018, 69, 1–2. [Google Scholar] [CrossRef]
- Kuang, B.-H.; Zhang, M.-Q.; Xu, L.-H.; Hu, L.-J.; Wang, H.-B.; Zhao, W.-F.; Du, Y.; Zhang, X. Proline-rich tyrosine kinase 2 and its phosphorylated form pY881 are novel prognostic markers for non-small-cell lung cancer progression and patients’ overall survival. Br. J. Cancer 2013, 109, 1252–1263. [Google Scholar] [CrossRef]
- Baldacci, S.; Kherrouche, Z.; Cockenpot, V.; Stoven, L.; Copin, M.C.; Werkmeister, E.; Marchand, N.; Kyheng, M.; Tulasne, D.; Cortot, A.B. MET amplification increases the metastatic spread of EGFR-mutated NSCLC. Lung Cancer 2018, 125, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Tykhomyrov, A.A.; Nedzvetsky, V.S.; Aĝca, C.A.; Guzyk, M.M.; Korsa, V.V.; Grinenko, T.V. Plasminogen/plasmin affects expression of glycolysis regulator TIGAR and induces autophagy in lung adenocarcinoma A549 cells. Exp. Oncol. 2020, 42, 270–276. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Lee, J.; Kljavin, N.M.; Haley, B.; Daemen, A.; Johnson, L.; Liang, Y. Requirement for BUB1B/BUBR1 in tumor progression of lung adenocarcinoma. Genes Cancer 2015, 6, 106–118. [Google Scholar] [CrossRef]
- Choi, E.-B.; Yang, A.-Y.; Kim, S.C.; Lee, J.; Choi, J.K.; Choi, C.; Kim, M.-Y. PARP1 enhances lung adenocarcinoma metastasis by novel mechanisms independent of DNA repair. Oncogene 2016, 35, 4569–4579. [Google Scholar] [CrossRef]
- Dey, S.; Sayers, C.M.; Verginadis, I.I.; Lehman, S.L.; Cheng, Y.; Cerniglia, G.J.; Tuttle, S.W.; Feldman, M.D.; Zhang, P.J.; Fuchs, S.Y.; et al. ATF4-dependent induction of heme oxygenase 1 prevents anoikis and promotes metastasis. J. Clin. Investig. 2015, 125, 2592–2608. [Google Scholar] [CrossRef]
- Madajewski, B.; Boatman, M.A.; Chakrabarti, G.; Boothman, D.A.; Bey, E.A. Depleting Tumor-NQO1 Potentiates Anoikis and Inhibits Growth of NSCLC. Mol. Cancer Res. 2016, 14, 14–25. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Liu, F.; Liu, H.; Wang, G.; Hao, H. Enhanced glycometabolism as a mechanism of NQO1 potentiated growth of NSCLC revealed by metabolomic profiling. Biochem. Biophys. Res. Commun. 2018, 496, 31–36. [Google Scholar] [CrossRef]
- Wang, G.X.; Tu, H.-C.; Dong, Y.; Skanderup, A.J.; Wang, Y.; Takeda, S.; Ganesan, Y.T.; Han, S.; Liu, H.; Hsieh, J.J.; et al. ΔNp63 Inhibits Oxidative Stress-Induced Cell Death, Including Ferroptosis, and Cooperates with the BCL-2 Family to Promote Clonogenic Survival. Cell Rep. 2017, 21, 2926–2939. [Google Scholar] [CrossRef]
- Li, N.; Liu, T.; Li, H.; Zhang, L.; Chu, L.; Meng, Q.; Qiao, Q.; Han, W.; Zhang, J.; Guo, M.; et al. ILF2 promotes anchorage independence through direct regulation of PTEN. Oncol. Lett. 2019, 18, 1689–1696. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, G.; Zhou, R.; Li, X.; Sun, Y.; Li, Y.; Du, W.; Yan, X.; Yang, J.; Chang, X.; et al. SPIB promotes anoikis resistance via elevated autolysosomal process in lung cancer cells. FEBS J. 2020, 287, 4696–4709. [Google Scholar] [CrossRef]
- Yao, S.; Huang, H.-Y.; Han, X.; Ye, Y.; Qin, Z.; Zhao, G.; Li, F.; Hu, G.; Hu, L.; Ji, H. Keratin 14-high subpopulation mediates lung cancer metastasis potentially through Gkn1 upregulation. Oncogene 2019, 38, 6354–6369. [Google Scholar] [CrossRef]
- Noto, A.; Raffa, S.; De Vitis, C.; Roscilli, G.; Malpicci, D.; Coluccia, P.; Di Napoli, A.; Ricci, A.; Giovagnoli, M.R.; Aurisicchio, L.; et al. Stearoyl-CoA desaturase-1 is a key factor for lung cancer-initiating cells. Cell Death Dis. 2013, 4, e947. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Hanibuchi, M.; Kuramoto, T.; Yamamori, N.; Goto, H.; Ogawa, H.; Mitsuhashi, A.; Van, T.T.; Kakiuchi, S.; Akiyama, S.-I.; et al. Macrophage stimulating protein promotes liver metastases of small cell lung cancer cells by affecting the organ microenvironment. Clin. Exp. Metastasis 2013, 30, 333–344. [Google Scholar] [CrossRef] [PubMed]
- Godin-Heymann, N.; Brabetz, S.; Murillo, M.M.; Saponaro, M.; Santos, C.R.; Lobley, A.; East, P.; Chakravarty, P.; Matthews, N.; Kelly, G.; et al. Tumour-suppression function of KLF12 through regulation of anoikis. Oncogene 2016, 35, 3324–3334. [Google Scholar] [CrossRef] [PubMed]
- Quintás-Cardama, A.; Post, S.M.; Solis, L.M.; Xiong, S.; Yang, P.; Chen, N.; Wistuba, I.I.; Killary, A.M.; Lozano, G. Loss of the novel tumour suppressor and polarity gene Trim62 (Dear1) synergizes with oncogenic Ras in invasive lung cancer. J. Pathol. 2014, 234, 108–119. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.N.; Zhang, Z.T.; Wang, L.; Wei, H.X.; Zhang, T.; Zhang, L.M.; Lin, H.; Zhang, H.; Wang, S.Q. TGF-β1/SH2B3 axis regulates anoikis resistance and EMT of lung cancer cells by modulating JAK2/STAT3 and SHP2/Grb2 signaling pathways. Cell Death Dis. 2022, 13, 472. [Google Scholar] [CrossRef]
- Halim, H.; Chunhacha, P.; Suwanborirux, K.; Chanvorachote, P. Anticancer and antimetastatic activities of Renieramycin M, a marine tetrahydroisoquinoline alkaloid, in human non-small cell lung cancer cells. Anticancer Res. 2011, 31, 193–201. [Google Scholar]
- Powan, P.; Saito, N.; Suwanborirux, K.; Chanvorachote, P. Ecteinascidin 770, a tetrahydroisoquinoline alkaloid, sensitizes human lung cancer cells to anoikis. Anticancer Res. 2013, 33, 505–512. [Google Scholar]
- Choochuay, K.; Chunhacha, P.; Pongrakhananon, V.; Luechapudiporn, R.; Chanvorachote, P. Imperatorin sensitizes anoikis and inhibits anchorage-independent growth of lung cancer cells. J. Nat. Med. 2013, 67, 599–606. [Google Scholar] [CrossRef]
- Wongpankam, E.; Chunhacha, P.; Pongrakhananon, V.; Sritularak, B.; Chanvorachote, P. Artonin E mediates MCL1 down-regulation and sensitizes lung cancer cells to anoikis. Anticancer Res. 2012, 32, 5343–5351. [Google Scholar]
- Wei, L.; Dai, Q.; Zhou, Y.; Zou, M.; Li, Z.; Lu, N.; Guo, Q. Oroxylin A sensitizes non-small cell lung cancer cells to anoikis via glucose-deprivation-like mechanisms: C-Src and hexokinase II. Biochim. Biophys. Acta 2013, 1830, 3835–3845. [Google Scholar] [CrossRef] [PubMed]
- Chaotham, C.; Pongrakhananon, V.; Sritularak, B.; Chanvorachote, P. A Bibenzyl from Dendrobium ellipsophyllum inhibits epithelial-to-mesenchymal transition and sensitizes lung cancer cells to anoikis. Anticancer Res. 2014, 34, 1931–1938. [Google Scholar] [PubMed]
- Ausawasamrit, A.; Itthiwarapornkul, N.; Chaotham, C.; Sukrong, S.; Chanvorachote, P. Lupalbigenin from Derris scandens Sensitizes Detachment-induced Cell Death in Human Lung Cancer Cells. Anticancer Res. 2015, 35, 2827–2834. [Google Scholar]
- Tsai, J.-Y.; Tsai, S.-H.; Wu, C.-C. The chemopreventive isothiocyanate sulforaphane reduces anoikis resistance and anchorage-independent growth in non-small cell human lung cancer cells. Toxicol. Appl. Pharmacol. 2019, 362, 116–124. [Google Scholar] [CrossRef]
- Putri, H.E.; Sritularak, B.; Chanvorachote, P. Pongamol Inhibits Epithelial to Mesenchymal Transition Through Suppression of FAK/Akt-mTOR Signaling. Anticancer Res. 2021, 41, 6147–6154. [Google Scholar] [CrossRef] [PubMed]
- Cao, R.; Jin, W.; Shan, Y.; Wang, J.; Liu, G.; Kuang, S.; Sun, C. Marine Bacterial Polysaccharide EPS11 Inhibits Cancer Cell Growth via Blocking Cell Adhesion and Stimulating Anoikis. Mar. Drugs 2018, 16, 85. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-J.; Choi, W.-I.; Jeon, B.-N.; Choi, K.-C.; Kim, K.; Kim, T.-J.; Ham, J.; Jang, H.J.; Kang, K.S.; Ko, H. Stereospecific effects of ginsenoside 20-Rg3 inhibits TGF-β1-induced epithelial–mesenchymal transition and suppresses lung cancer migration, invasion and anoikis resistance. Toxicology 2014, 322, 23–33. [Google Scholar] [CrossRef]
- Kim, H.; Choi, P.; Kim, T.; Kim, Y.; Song, B.G.; Park, Y.-T.; Choi, S.-J.; Yoon, C.H.; Lim, W.-C.; Ko, H.; et al. Ginsenosides Rk1 and Rg5 inhibit transforming growth factor-β1-induced epithelial-mesenchymal transition and suppress migration, invasion, anoikis resistance, and development of stem-like features in lung cancer. J. Ginseng Res. 2021, 45, 134–148. [Google Scholar] [CrossRef]
- Ecoy, G.A.U.; Chamni, S.; Suwanborirux, K.; Chanvorachote, P.; Chaotham, C. Jorunnamycin A from Xestospongia sp. Suppresses Epithelial to Mesenchymal Transition and Sensitizes Anoikis in Human Lung Cancer Cells. J. Nat. Prod. 2019, 82, 1861–1873. [Google Scholar] [CrossRef]
- Busaranon, K.; Plaimee, P.; Sritularak, B.; Chanvorachote, P. Moscatilin inhibits epithelial-to-mesenchymal transition and sensitizes anoikis in human lung cancer H460 cells. J. Nat. Med. 2016, 70, 18–27. [Google Scholar] [CrossRef]
- Unahabhokha, T.; Chanvorachote, P.; Pongrakhananon, V. The attenuation of epithelial to mesenchymal transition and induction of anoikis by gigantol in human lung cancer H460 cells. Tumor Biol. 2016, 37, 8633–8641. [Google Scholar] [CrossRef] [PubMed]
- Nonpanya, N.; Prakhongcheep, O.; Petsri, K.; Jitjaicham, C.; Tungsukruthai, S.; Sritularak, B.; Chanvorachote, P. Ephemeranthol A Suppresses Epithelial to Mesenchymal Transition and FAK-Akt Signaling in Lung Cancer Cells. Anticancer Res. 2020, 40, 4989–4999. [Google Scholar] [CrossRef] [PubMed]
- Pengpaeng, P.; Sritularak, B.; Chanvorachote, P. Dendrofalconerol A sensitizes anoikis and inhibits migration in lung cancer cells. J. Nat. Med. 2015, 69, 178–190. [Google Scholar] [CrossRef] [PubMed]
- Chanvorachote, P.; Kowitdamrong, A.; Ruanghirun, T.; Sritularak, B.; Mungmee, C.; Likhitwitayawuid, K. Anti-metastatic Activities of Bibenzyls from Dendrobium pulchellum. Nat. Prod. Commun. 2013, 8, 115–118. [Google Scholar] [CrossRef]
- Tanagornmeatar, K.; Chaotham, C.; Sritularak, B.; Likhitwitayawuid, K.; Chanvorachote, P. Cytotoxic and anti-metastatic activities of phenolic compounds from Dendrobium ellipsophyllum. Anticancer Res. 2014, 34, 6573–6579. [Google Scholar]
- Lee, H.-Z.; Yang, W.-H.; Hour, M.-J.; Wu, C.-Y.; Peng, W.-H.; Bao, B.-Y.; Han, P.-H.; Bau, D.-T. Photodynamic activity of aloe-emodin induces resensitization of lung cancer cells to anoikis. Eur. J. Pharmacol. 2010, 648, 50–58. [Google Scholar] [CrossRef]
- Chang, W.-T.; You, B.-J.; Yang, W.-H.; Wu, C.-Y.; Bau, D.-T.; Lee, H.-Z. Protein kinase C delta-mediated cytoskeleton remodeling is involved in aloe-emodin-induced photokilling of human lung cancer cells. Anticancer Res. 2012, 32, 3707–3713. [Google Scholar]
- Pongrakhananon, V.; Nimmannit, U.; Luanpitpong, S.; Rojanasakul, Y.; Chanvorachote, P. Curcumin sensitizes non-small cell lung cancer cell anoikis through reactive oxygen species-mediated Bcl-2 downregulation. Apoptosis 2010, 15, 574–585. [Google Scholar] [CrossRef]
- Ko, H. Geraniin inhibits TGF-β1-induced epithelial–mesenchymal transition and suppresses A549 lung cancer migration, invasion and anoikis resistance. Bioorganic Med. Chem. Lett. 2015, 25, 3529–3534. [Google Scholar] [CrossRef]
- Lim, W.-C.; Choi, J.W.; Song, N.-E.; Cho, C.-W.; Rhee, Y.K.; Hong, H.-D. Polysaccharide isolated from persimmon leaves (Diospyros kaki Thunb.) suppresses TGF-β1-induced epithelial-to-mesenchymal transition in A549 cells. Int. J. Biol. Macromol. 2020, 164, 3835–3845. [Google Scholar] [CrossRef]
- Trabalzini, L.; Ercoli, J.; Trezza, A.; Schiavo, I.; Macrì, G.; Moglia, A.; Spiga, O.; Finetti, F. Pharmacological and In Silico Analysis of Oat Avenanthramides as EGFR Inhibitors: Effects on EGF-Induced Lung Cancer Cell Growth and Migration. Int. J. Mol. Sci. 2022, 23, 8534. [Google Scholar] [CrossRef] [PubMed]
- Que, Z.-J.; Yang, Y.; Liu, H.-T.; Shang-Guan, W.-J.; Yu, P.; Zhu, L.-H.; Li, H.-G.; Liu, H.-M.; Tian, J.-H. Jinfukang regulates integrin/Src pathway and anoikis mediating circulating lung cancer cells migration. J. Ethnopharmacol. 2021, 267, 113473. [Google Scholar] [CrossRef] [PubMed]
- Hao, H.; Guo, Z.; Li, Z.; Li, J.; Jiang, S.; Fu, J.; Jiao, Y.; Deng, X.; Han, S.; Li, P. Modified Bu-Fei decoction inhibits lung metastasis via suppressing angiopoietin-like 4. Phytomedicine 2022, 106, 154409. [Google Scholar] [CrossRef] [PubMed]
- Sakuma, Y.; Tsunezumi, J.; Nakamura, Y.; Yoshihara, M.; Matsukuma, S.; Koizume, S.; Miyagi, Y. ABT-263, a Bcl-2 inhibitor, enhances the susceptibility of lung adenocarcinoma cells treated with Src inhibitors to anoikis. Oncol. Rep. 2011, 25, 661–667. [Google Scholar] [CrossRef]
- Sakuma, Y.; Yamazaki, Y.; Nakamura, Y.; Yoshihara, M.; Matsukuma, S.; Nakayama, H.; Yokose, T.; Kameda, Y.; Koizume, S.; Miyagi, Y. WZ4002, a third-generation EGFR inhibitor, can overcome anoikis resistance in EGFR-mutant lung adenocarcinomas more efficiently than Src inhibitors. Lab. Investig. 2012, 92, 371–383. [Google Scholar] [CrossRef]
- Kim, S.; Ko, D.; Lee, Y.; Jang, S.; Lee, Y.; Lee, I.Y.; Kim, S. Anti-cancer activity of the novel 2-hydroxydiarylamide derivatives IMD-0354 and KRT1853 through suppression of cancer cell invasion, proliferation, and survival mediated by TMPRSS4. Sci. Rep. 2019, 9, 10003. [Google Scholar] [CrossRef]
- Shochet, G.E.; Israeli-Shani, L.; Koslow, M.; Shitrit, D. Nintedanib (BIBF 1120) blocks the tumor promoting signals of lung fibroblast soluble microenvironment. Lung Cancer 2016, 96, 7–14. [Google Scholar] [CrossRef]
- Sirimangkalakitti, N.; Chamni, S.; Suwanborirux, K.; Chanvorachote, P. Renieramycin M Sensitizes Anoikis-resistant H460 Lung Cancer Cells to Anoikis. Anticancer Res. 2016, 36, 1665–1671. [Google Scholar]
- Prateep, A.; Sumkhemthong, S.; Karnsomwan, W.; De-Eknamkul, W.; Chamni, S.; Chanvorachote, P.; Chaotham, C. Avicequinone B sensitizes anoikis in human lung cancer cells. J. Biomed. Sci. 2018, 25, 32. [Google Scholar] [CrossRef]
- Karsch-Bluman, A.; Avraham, S.; Assayag, M.; Schwob, O.; Benny, O. Encapsulated Carbenoxolone Reduces Lung Metastases. Cancers 2019, 11, 1383. [Google Scholar] [CrossRef]
- Pongrakhananon, V.; Stueckle, T.A.; Wang, H.-Y.L.; O’Doherty, G.A.; Dinu, C.Z.; Chanvorachote, P.; Rojanasakul, Y. Monosaccharide digitoxin derivative sensitize human non-small cell lung cancer cells to anoikis through Mcl-1 proteasomal degradation. Biochem. Pharmacol. 2014, 88, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Sriratanasak, N.; Nonpanya, N.; Wattanathana, W.; Chanvorachote, P. Benzoxazine Dimer Analogue Targets Integrin β3 in Lung Cancer Cells and Suppresses Anoikis Resistance and Migration. Anticancer Res. 2020, 40, 2583–2589. [Google Scholar] [CrossRef]
- Puchsaka, P.; Chaotham, C.; Chanvorachote, P. α-Lipoic acid sensitizes lung cancer cells to chemotherapeutic agents and anoikis via integrin β1/β3 downregulation. Int. J. Oncol. 2016, 49, 1445–1456. [Google Scholar] [CrossRef] [PubMed]
- Pramchu-Em, C.; Meksawan, K.; Chanvorachote, P. Zinc Sensitizes Lung Cancer Cells to Anoikis through Down-Regulation of Akt and Caveolin-1. Nutr. Cancer 2016, 68, 312–319. [Google Scholar] [CrossRef] [PubMed]
- Aixinjueluo, W.; Furukawa, K.; Zhang, Q.; Hamamura, K.; Tokuda, N.; Yoshida, S.; Ueda, R.; Furukawa, K. Mechanisms for the Apoptosis of Small Cell Lung Cancer Cells Induced by Anti-GD2 Monoclonal Antibodies. J. Biol. Chem. 2005, 280, 29828–29836. [Google Scholar] [CrossRef]
- Salo, S.; Boutaud, A.; Hansen, A.J.; He, L.; Sun, Y.; Morales, S.; Venturini, A.; Martin, P.; Nokelainen, P.; Betsholtz, C.; et al. Antibodies blocking adhesion and matrix binding domains of laminin-332 inhibit tumor growth and metastasis in vivo. Int. J. Cancer 2009, 125, 1814–1825. [Google Scholar] [CrossRef]
- Hong, K.P.; Shin, M.H.; Yoon, S.; Ji, G.Y.; Moon, Y.R.; Lee, O.-J.; Choi, S.-Y.; Lee, Y.-M.; Koo, J.H.; Lee, H.-C.; et al. Therapeutic effect of anti CEACAM6 monoclonal antibody against lung adenocarcinoma by enhancing anoikis sensitivity. Biomaterials 2015, 67, 32–41. [Google Scholar] [CrossRef]
- Lee, H.; Jang, Y.; Park, S.; Jang, H.; Park, E.-J.; Kim, H.J.; Kim, H. Development and evaluation of a CEACAM6-targeting theranostic nanomedicine for photoacoustic-based diagnosis and chemotherapy of metastatic cancer. Theranostics 2018, 8, 4247–4261. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Luo, Z.; Lin, L.; Sui, X.; Yu, L.; Xu, C.; Zhang, R.; Zhao, Z.; Zhu, Q.; An, B.; et al. Anoikis-Associated Lung Cancer Metastasis: Mechanisms and Therapies. Cancers 2022, 14, 4791. https://doi.org/10.3390/cancers14194791
Wang J, Luo Z, Lin L, Sui X, Yu L, Xu C, Zhang R, Zhao Z, Zhu Q, An B, et al. Anoikis-Associated Lung Cancer Metastasis: Mechanisms and Therapies. Cancers. 2022; 14(19):4791. https://doi.org/10.3390/cancers14194791
Chicago/Turabian StyleWang, Jing, Zhijie Luo, Lizhu Lin, Xinbing Sui, Lili Yu, Cong Xu, Ruonan Zhang, Ziming Zhao, Qianru Zhu, Bo An, and et al. 2022. "Anoikis-Associated Lung Cancer Metastasis: Mechanisms and Therapies" Cancers 14, no. 19: 4791. https://doi.org/10.3390/cancers14194791
APA StyleWang, J., Luo, Z., Lin, L., Sui, X., Yu, L., Xu, C., Zhang, R., Zhao, Z., Zhu, Q., An, B., Wang, Q., Chen, B., Leung, E. L.-H., & Wu, Q. (2022). Anoikis-Associated Lung Cancer Metastasis: Mechanisms and Therapies. Cancers, 14(19), 4791. https://doi.org/10.3390/cancers14194791