
Molecular Interactions of the Long Noncoding RNA NEAT1 in Cancer


Abstract
:Simple Summary
Abstract
1. Introduction
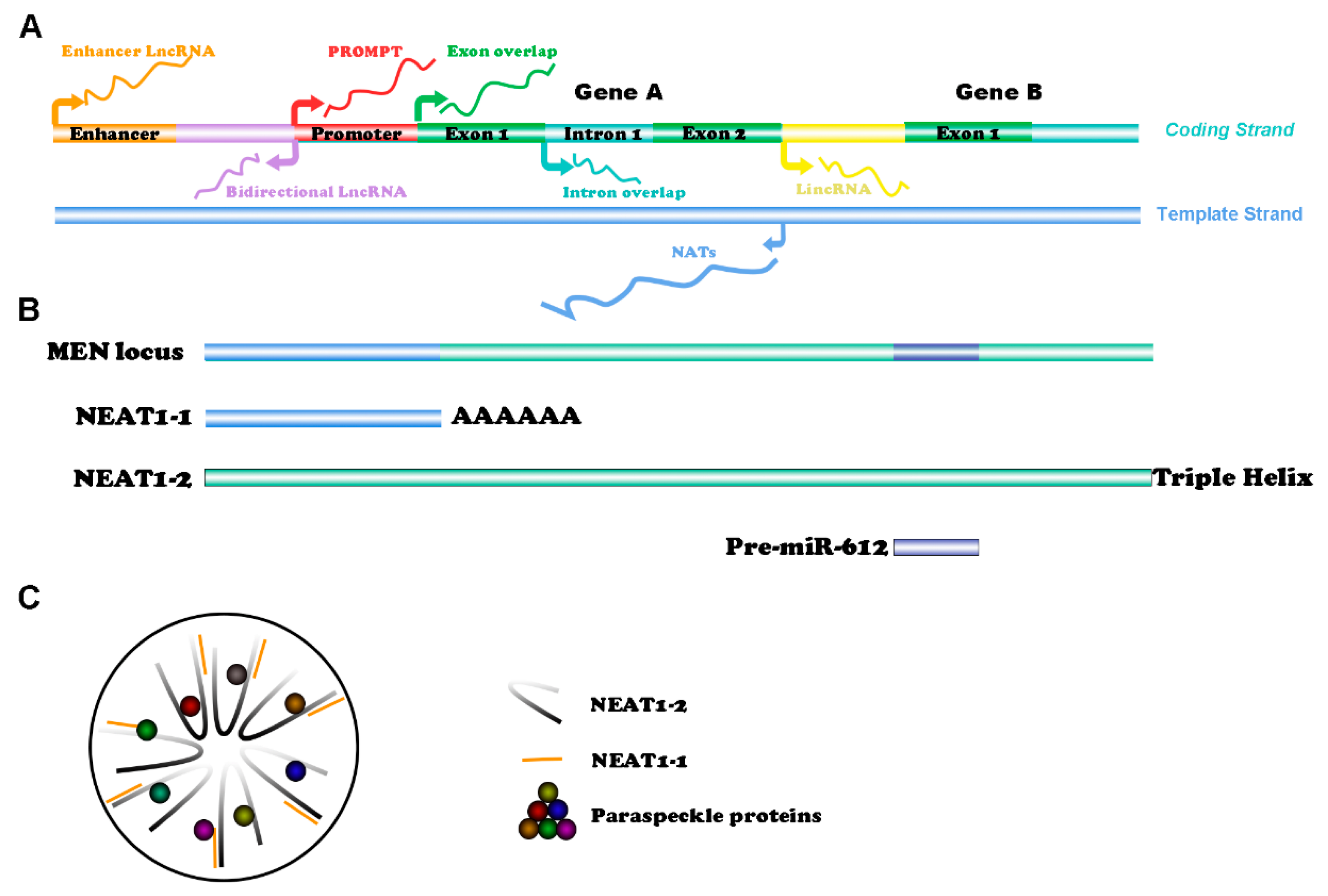
2. Structure of NEAT1 and Paraspeckles
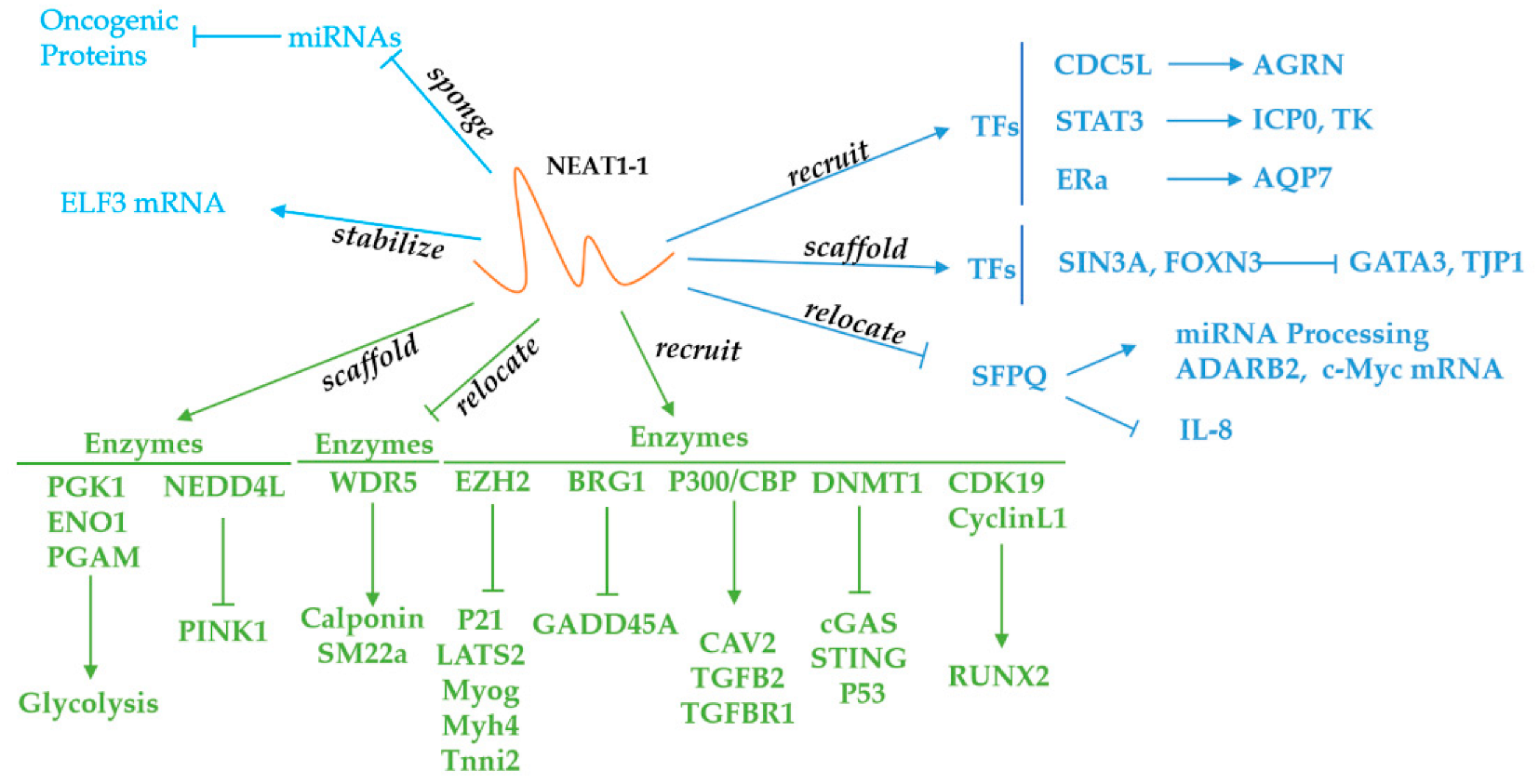
3. NEAT1 Sponges microRNAs and Stabilizes mRNAs
4. NEAT1 Recruits Transcription Factors, Enzymes and Other Proteins
5. NEAT1-Binding Enzymes
6. NEAT1-Binding Transcription Factors
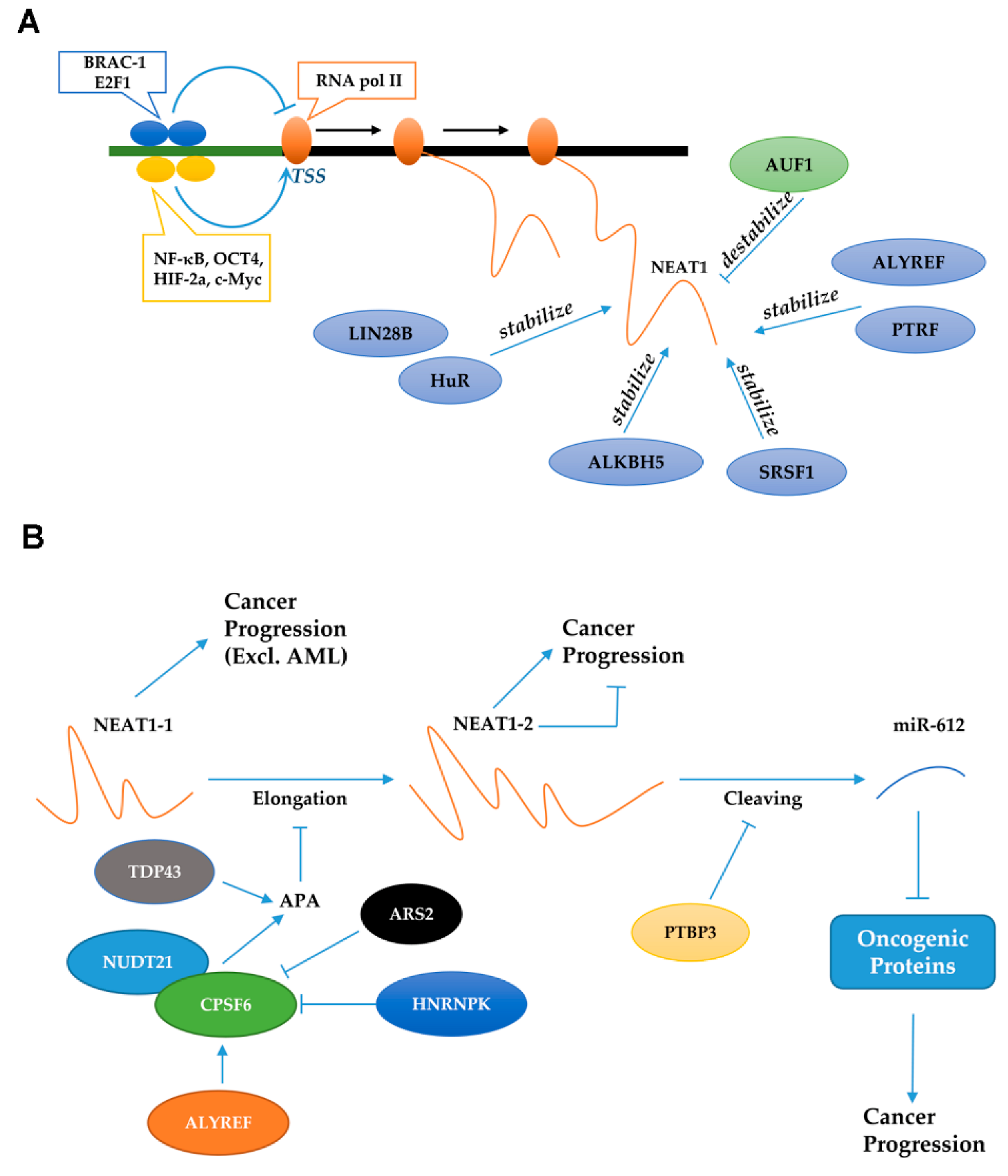
7. NEAT1 Interacts with DNA Sequences to Regulate Transcription
8. Factors That Regulate NEAT1 Expression
9. NEAT1-Containing Exosomes in Cancer
10. Conclusions and Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Djebali, S.; Davis, C.A.; Merkel, A.; Dobin, A.; Lassmann, T.; Mortazavi, A.; Tanzer, A.; Lagarde, J.; Lin, W.; Schlesinger, F.; et al. Landscape of transcription in human cells. Nature 2012, 489, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Kopp, F.; Mendell, J.T. Functional Classification and Experimental Dissection of Long Noncoding RNAs. Cell 2018, 172, 393–407. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Yang, L.; Chen, L.L. The Diversity of Long Noncoding RNAs and Their Generation. Trends Genet. 2017, 33, 540–552. [Google Scholar] [CrossRef] [PubMed]
- Mello, S.S.; Sinow, C.; Raj, N.; Mazur, P.K.; Bieging-Rolett, K.; Broz, D.K.; Imam, J.F.C.; Vogel, H.; Wood, L.D.; Sage, J.; et al. Neat1is a p53-inducible lincRNA essential for transformation suppression. Genes Dev. 2017, 31, 1095–1108. [Google Scholar] [CrossRef]
- Yu, X.; Li, Z.; Zheng, H.; Chan, M.T.; Wu, W.K. NEAT1: A novel cancer-related long non-coding RNA. Cell Prolif. 2017, 50, e12329. [Google Scholar] [CrossRef]
- Liang, J.; Liu, C.; Xu, D.; Xie, K.; Li, A. LncRNA NEAT1 facilitates glioma progression via stabilizing PGK1. J. Transl. Med. 2022, 20, 80. [Google Scholar] [CrossRef]
- Klec, C.; Prinz, F.; Pichler, M. Involvement of the long noncoding RNA NEAT1 in carcinogenesis. Mol. Oncol. 2019, 13, 46–60. [Google Scholar] [CrossRef]
- Fang, L.; Sun, J.; Pan, Z.; Song, Y.; Zhong, L.; Zhang, Y.; Liu, Y.; Zheng, X.; Huang, P. Long non-coding RNA NEAT1 promotes hepatocellular carcinoma cell proliferation through the regulation of miR-129-5p-VCP-IκB. Am. J. Physiol. Gastrointest. Liver Physiol. 2017, 313, G150–G156. [Google Scholar] [CrossRef]
- Xiong, W.; Huang, C.; Deng, H.; Jian, C.; Zen, C.; Ye, K.; Zhong, Z.; Zhao, X.; Zhu, L. Oncogenic non-coding RNA NEAT1 promotes the prostate cancer cell growth through the SRC3/IGF1R/AKT pathway. Int. J. Biochem. Cell Biol. 2018, 94, 125–132. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, Y.; Fu, X.; Lu, Z. Long non-coding RNA NEAT1 promoted ovarian cancer cells’ metastasis through regulation of miR-382-3p/ROCK1 axial. Cancer Sci. 2018, 109, 2188–2198. [Google Scholar] [CrossRef]
- Yu, H.; Peng, S.; Chen, X.; Han, S.; Luo, J. Long non-coding RNA NEAT1 serves as a novel biomarker for treatment response and survival profiles via microRNA-125a in multiple myeloma. J. Clin. Lab. Anal. 2020, 34, e23399. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Yi, Z.J.; Ou, Z.L.; Han, T.; Wan, T.; Tang, Y.C.; Wang, Z.C.; Huang, F.Z. RELA/NEAT1/miR-302a-3p/RELA feedback loop modulates pancreatic ductal adenocarcinoma cell proliferation and migration. J. Cell Physiol. 2019, 234, 3583–3597. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Weng, W.; Zhang, Q.; Wu, Y.; Ni, S.; Tan, C.; Xu, M.; Sun, H.; Liu, C.; Wei, P.; et al. The lncRNA NEAT1 activates Wnt/beta-catenin signaling and promotes colorectal cancer progression via interacting with DDX5. J. Hematol. Oncol. 2018, 11, 113. [Google Scholar] [CrossRef] [PubMed]
- Jiang, P.; Wu, X.; Wang, X.; Huang, W.; Feng, Q. NEAT1 upregulates EGCG-induced CTR1 to enhance cisplatin sensitivity in lung cancer cells. Oncotarget 2016, 7, 43337–43351. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Jiang, G.; Ren, W.; Wang, B.; Yang, C.; Li, M. LncRNA NEAT1 regulates 5-fu sensitivity, apoptosis and invasion in colorectal cancer through the MiR-150-5p/CPSF4 axis. OncoTargets Ther. 2020, 13, 6373–6383. [Google Scholar] [CrossRef]
- Zeng, C.; Xu, Y.; Xu, L.; Yu, X.; Cheng, J.; Yang, L.; Chen, S.; Li, Y. Inhibition of long non-coding RNA NEAT1 impairs myeloid differentiation in acute promyelocytic leukemia cells. BMC Cancer 2014, 14, 693. [Google Scholar] [CrossRef]
- Feng, S.; Liu, N.; Chen, X.; Liu, Y.; An, J. Long non-coding RNA NEAT1/miR-338-3p axis impedes the progression of acute myeloid leukemia via regulating CREBRF. Cancer Cell Int. 2020, 20, 112. [Google Scholar] [CrossRef]
- Dong, P.; Xiong, Y.; Yue, J.; Hanley, S.J.B.; Kobayashi, N.; Todo, Y.; Watari, H. Long Non-coding RNA NEAT1: A Novel Target for Diagnosis and Therapy in Human Tumors. Front. Genet. 2018, 9, 471. [Google Scholar] [CrossRef]
- Naganuma, T.; Hirose, T. Paraspeckle formation during the biogenesis of long non-coding RNAs. RNA Biol. 2013, 10, 456–461. [Google Scholar] [CrossRef]
- Hirose, T.; Virnicchi, G.; Tanigawa, A.; Naganuma, T.; Li, R.; Kimura, H.; Yokoi, T.; Nakagawa, S.; Bénard, M.; Fox, A.H.; et al. NEAT1 long noncoding RNA regulates transcription via protein sequestration within subnuclear bodies. Mol. Biol. Cell 2014, 25, 169–183. [Google Scholar] [CrossRef]
- Wang, Z.; Li, K.; Huang, W. Long non-coding RNA NEAT1-centric gene regulation. Cell Mol. Life Sci. 2020, 77, 3769–3779. [Google Scholar] [CrossRef] [PubMed]
- Machitani, M.; Taniguchi, I.; Ohno, M. ARS2 Regulates Nuclear Paraspeckle Formation through 3′-End Processing and Stability of NEAT1 Long Noncoding RNA. Mol. Cell. Biol. 2020, 40, e00269–19. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Qu, S.; Wang, L.; Zhang, H.; Yang, Z.; Wang, J.; Dai, B.; Tao, K.; Shang, R.; Liu, Z.; et al. PTBP3 splicing factor promotes hepatocellular carcinoma by destroying the splicing balance of NEAT1 and pre-miR-612. Oncogene 2018, 37, 6399–6413. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Schmidt, B.F.; Bruchez, M.P.; McManus, C.J. Structural analyses of NEAT1 lncRNAs suggest long-range RNA interactions that may contribute to paraspeckle architecture. Nucleic Acids Res. 2018, 46, 3742–3752. [Google Scholar] [CrossRef] [PubMed]
- Prinz, F.; Kapeller, A.; Pichler, M.; Klec, C. The Implications of the Long Non-Coding RNA NEAT1 in Non-Cancerous Diseases. Int. J. Mol. Sci. 2019, 20, 627. [Google Scholar] [CrossRef] [PubMed]
- Nishimoto, Y.; Nakagawa, S.; Hirose, T.; Okano, H.J.; Takao, M.; Shibata, S.; Suyama, S.; Kuwako, K.; Imai, T.; Murayama, S.; et al. The long non-coding RNA nuclear-enriched abundant transcript 1_2 induces paraspeckle formation in the motor neuron during the early phase of amyotrophic lateral sclerosis. Mol. Brain 2013, 6, 31. [Google Scholar] [CrossRef] [PubMed]
- Ghafouri-Fard, S.; Taheri, M. Nuclear Enriched Abundant Transcript 1 (NEAT1): A long non-coding RNA with diverse functions in tumorigenesis. Biomed. Pharmacother. 2019, 111, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Prasanth, K.V.; Prasanth, S.G.; Xuan, Z.; Hearn, S.; Freier, S.M.; Bennett, C.F.; Zhang, M.Q.; Spector, D.L. Regulating gene expression through RNA nuclear retention. Cell 2005, 123, 249–263. [Google Scholar] [CrossRef]
- Zhang, Z.; Carmichael, G.G. The fate of dsRNA in the nucleus: A p54(nrb)-containing complex mediates the nuclear retention of promiscuously A-to-I edited RNAs. Cell 2001, 106, 465–475. [Google Scholar] [CrossRef]
- Xu, L.D.; Öhman, M. ADAR1 Editing and its Role in Cancer. Genes 2018, 10, 12. [Google Scholar] [CrossRef]
- Shen, X.; Song, S.; Li, C.; Zhang, J. Synonymous mutations in representative yeast genes are mostly strongly non-neutral. Nature 2022, 606, 725–731. [Google Scholar] [CrossRef] [PubMed]
- Imamura, K.; Imamachi, N.; Akizuki, G.; Kumakura, M.; Kawaguchi, A.; Nagata, K.; Kato, A.; Kawaguchi, Y.; Sato, H.; Yoneda, M.; et al. Long noncoding RNA NEAT1-dependent SFPQ relocation from promoter region to paraspeckle mediates IL8 expression upon immune stimuli. Mol. Cell 2014, 53, 393–406. [Google Scholar] [CrossRef] [PubMed]
- Zan, J.; Zhao, X.; Deng, X.; Ding, H.; Wang, B.; Lu, M.; Wei, Z.; Huang, Z.; Wang, S. Paraspeckle Promotes Hepatocellular Carcinoma Immune Escape by Sequestering IFNGR1 mRNA. Cell. Mol. Gastroenterol. Hepatol. 2021, 12, 465–487. [Google Scholar] [CrossRef] [PubMed]
- Tay, Y.; Rinn, J.; Pandolfi, P.P. The multilayered complexity of ceRNA crosstalk and competition. Nature 2014, 505, 344–352. [Google Scholar] [CrossRef]
- Sun, W.; Qin, Y.; Wang, Z.; Dong, W.; He, L.; Zhang, T.; Zhang, H. The NEAT1_2/miR-491 Axis Modulates Papillary Thyroid Cancer Invasion and Metastasis Through TGM2/NFκb/FN1 Signaling. Front. Oncol. 2021, 11, 610547. [Google Scholar] [CrossRef]
- Li, W.; Zhang, Z.; Liu, X.; Cheng, X.; Zhang, Y.; Han, X.; Zhang, Y.; Liu, S.; Yang, J.; Xu, B.; et al. The FOXN3-NEAT1-SIN3A repressor complex promotes progression of hormonally responsive breast cancer. J. Clin. Investig. 2017, 127, 3421–3440. [Google Scholar] [CrossRef]
- Jiang, X.; Zhou, Y.; Sun, A.-J.; Xue, J.-L. NEAT1 contributes to breast cancer progression through modulating miR-448 and ZEB1. J. Cell. Physiol. 2018, 233, 8558–8566. [Google Scholar] [CrossRef]
- Liu, X.; Yao, W.; Xiong, H.; Li, Q.; Li, Y. LncRNA NEAT1 accelerates breast cancer progression through regulating miR-410-3p/CCND1 axis. Cancer Biomark. 2020, 29, 277–290. [Google Scholar] [CrossRef]
- Li, X.; Wang, S.; Li, Z.; Long, X.; Guo, Z.; Zhang, G.; Zu, J.; Chen, Y.; Wen, L. The lncRNA NEAT1 facilitates cell growth and invasion via the miR-211/HMGA2 axis in breast cancer. Int. J. Biol. Macromol. 2017, 105, 346–353. [Google Scholar] [CrossRef]
- Yao, L.; Chen, L.; Zhou, H.; Duan, F.; Wang, L.; Zhang, Y. Long Noncoding RNA NEAT1 Promotes the Progression of Breast Cancer by Regulating miR-138-5p/ZFX Axis. Cancer Biother. Radiopharm. 2020. [Google Scholar] [CrossRef]
- Zhou, D.P.; Gu, J.; Wang, Y.P.; Wu, H.G.; Cheng, W.; Wang, Q.P.; Zheng, G.P.; Wang, X.D. Long non-coding RNA NEAT1 transported by extracellular vesicles contributes to breast cancer development by sponging microRNA-141-3p and regulating KLF12. Cell Biosci. 2021, 11, 68. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Liu, Z.; Li, Z.; Wang, S.; Shen, N.; Xin, Y.; Huang, T. Long non-coding RNA nuclear paraspeckle assembly transcript 1 interacts with microRNA-107 to modulate breast cancer growth and metastasis by targeting carnitine palmitoyltransferase-1. Int. J. Oncol. 2019, 55, 1125–1136. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Malhotra, A. Evaluation of Potential of Long Noncoding RNA NEAT1 in Colorectal Cancer. J. Environ. Pathol. Toxicol. Oncol. Off. Organ Int. Soc. Environ. Toxicol. Cancer 2020, 39, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Zhong, F.; Zhang, W.; Cao, Y.; Wen, Q.; Cao, Y.; Lou, B.; Li, J.; Shi, W.; Liu, Y.; Luo, R.; et al. LncRNA NEAT1 promotes colorectal cancer cell proliferation and migration via regulating glial cell-derived neurotrophic factor by sponging miR-196a-5p. Acta Biochim. Biophys. Sin. 2018, 50, 1190–1199. [Google Scholar] [CrossRef]
- Liu, K.; Lei, S.; Kuang, Y.; Jin, Q.; Long, D.; Liu, C.; Jiang, Y.; Zhao, H.; Yao, H. A Novel Mechanism of the c-Myc/NEAT1 Axis Mediating Colorectal Cancer Cell Response to Photodynamic Therapy Treatment. Front. Oncol. 2021, 11, 652831. [Google Scholar] [CrossRef]
- Zhuang, S.T.; Cai, Y.J.; Liu, H.P.; Qin, Y.; Wen, J.F. LncRNA NEAT1/miR-185-5p/IGF2 axis regulates the invasion and migration of colon cancer. Mol. Genet. Genom. Med. 2020, 8, e1125. [Google Scholar] [CrossRef]
- Gao, Z.; Shi, Y.; Wang, J.; Li, W.; Bao, Y.; Wu, D.; Gu, Y. Long non-coding RNA NEAT1 absorbs let-7 g-5p to induce epithelial-mesenchymal transition of colon cancer cells through upregulating BACH1. Dig. Liver Dis. 2021. [Google Scholar] [CrossRef]
- Wu, D.; Li, H.; Wang, J.; Li, H.; Xiao, Q.; Zhao, X.; Huo, Z. LncRNA NEAT1 promotes gastric cancer progression via miR-1294/AKT1 axis. Open Med. 2020, 15, 1028–1038. [Google Scholar] [CrossRef]
- Gao, M.; Liu, L.; Zhang, D.; Yang, Y.; Chang, Z. Long non-coding rna neat1 serves as sponge for mir-365a-3p to promote gastric cancer progression via regulating abcc4. OncoTargets Ther. 2020, 13, 3977–3985. [Google Scholar] [CrossRef]
- Jiang, Y.; Jin, S.; Tan, S.; Xue, Y.; Cao, X. Long noncoding RNA NEAT1 regulates radio-sensitivity via microRNA-27b-3p in gastric cancer. Cancer Cell Int. 2020, 20, 581. [Google Scholar] [CrossRef]
- Zhou, Y.; Sha, Z.; Yang, Y.; Wu, S.; Chen, H. lncRNA NEAT1 regulates gastric carcinoma cell proliferation, invasion and apoptosis via the miR-500a-3p/XBP-1 axis. Mol. Med. Rep. 2021, 24, 503. [Google Scholar] [CrossRef]
- Tan, H.Y.; Wang, C.; Liu, G.; Zhou, X. Long noncoding RNA NEAT1-modulated miR-506 regulates gastric cancer development through targeting STAT3. J. Cell. Biochem. 2019, 120, 4827–4836. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chang, L.; Wu, Q.; Zuo, F. Long non-coding RNA NEAT1 increases the aggressiveness of gastric cancer by regulating the microRNA-142-5p/JAG1 axis. Exp. Ther. Med. 2021, 22, 862. [Google Scholar] [CrossRef] [PubMed]
- Zhai, S.; Gao, F.; Zhao, C.; Li, H.; Jiao, X. Lncrna neat1 enhances cell proliferation and invasion by regulating the MIR-221-5p/ SOX11 axis in gastric cancer. Basic Clin. Pharmacol. Toxicol. 2021, 128, 33–34. [Google Scholar]
- Rao, X.; Liu, X.; Liu, N.; Zhang, Y.; Zhang, Z.; Zhou, L.; Han, G.; Cen, R.; Shi, N.; Zhu, H.; et al. Long noncoding RNA NEAT1 promotes tumorigenesis in H. pylori gastric cancer by sponging miR-30a to regulate COX-2/BCL9 pathway. Helicobacter 2021, 26, e12847. [Google Scholar] [CrossRef]
- Li, B.; Lu, X.; Ma, C.; Sun, S.; Shu, X.; Wang, Z.; Sun, W. Long non-coding RNA NEAT1 promotes human glioma tumor progression via miR-152-3p/CCT6A pathway. Neurosci. Lett. 2020, 732, 135086. [Google Scholar] [CrossRef]
- Zhou, K.; Zhang, C.; Yao, H.; Zhang, X.; Zhou, Y.; Che, Y.; Huang, Y. Knockdown of long non-coding RNA NEAT1 inhibits glioma cell migration and invasion via modulation of SOX2 targeted by miR-132. Mol. Cancer 2018, 17, 105. [Google Scholar] [CrossRef]
- Yu, H.; Xu, A.; Wu, B.; Wang, M.; Chen, Z. Long noncoding RNA NEAT1 promotes progression of glioma as a ceRNA by sponging miR-185-5p to stimulate DNMT1/mTOR signaling. J. Cell. Physiol. 2021, 236, 121–130. [Google Scholar] [CrossRef]
- Li, Z.; Liu, Y.-H.; Diao, H.-Y.; Ma, J.; Yao, Y.-L. Long noncoding RNA NEAT1 promotes glioma pathogenesis by regulating miR-449b-5p/c-Met axis. Tumor Biol. 2016, 37, 673–683. [Google Scholar] [CrossRef]
- Wu, D.M.; Wang, S.; Wen, X.; Han, X.R.; Wang, Y.J.; Fan, S.H.; Zhang, Z.F.; Shan, Q.; Lu, J.; Zheng, Y.L. Long noncoding RNA nuclear enriched abundant transcript 1 impacts cell proliferation, invasion, and migration of glioma through regulating miR-139-5p/ CDK6. J. Cell. Physiol. 2019, 234, 5972–5987. [Google Scholar] [CrossRef]
- Zhang, X.N.; Zhou, J.; Lu, X.J. The long noncoding RNA NEAT1 contributes to hepatocellular carcinoma development by sponging miR-485 and enhancing the expression of the STAT3. J. Cell. Physiol. 2018, 233, 6733–6741. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Hao, L.P.; Song, H.; Chu, X.Y.; Wang, R. The Potential Roles of Exosomal Non-Coding RNAs in Hepatocellular Carcinoma. Front. Oncol. 2022, 12, 790916. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Xiao, T.; Xiao, Y.; Li, Y. Silencing of long non-coding RNA NEAT1 inhibits hepatocellular carcinoma progression by downregulating SMO by sponging microRNA-503. Mol. Med. Rep. 2020, 23, 168. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Wang, X.; Zhou, Y.; Cheng, L.; Zhang, Y.; Zhang, Y. Long noncoding rna neat1 promotes cell proliferation and invasion and suppresses apoptosis in hepatocellular carcinoma by regulating mirna-22-3p/akt2 in vitro and in vivo. OncoTargets Ther. 2019, 12, 8991–9004. [Google Scholar] [CrossRef] [PubMed]
- Tu, J.; Zhao, Z.; Xu, M.; Lu, X.; Chang, L.; Ji, J. NEAT1 upregulates TGF-β1 to induce hepatocellular carcinoma progression by sponging hsa-mir-139-5p. J. Cell. Physiol. 2018, 233, 8578–8587. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Huang, C.; Zhu, Z.; Hou, Y.; Huang, S.; Sun, C.; Tang, Y.; Zhang, Z.; Wang, L.; Chen, H.; et al. LncRNA NEAT1 regulates the proliferation and migration of hepatocellular carcinoma cells by acting as a miR-320a molecular sponge and targeting L antigen family member 3. Int. J. Oncol. 2020, 57, 1001–1012. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Liang, Y.; Song, R.; Yang, G.; Han, J.; Lan, Y.; Pan, S.; Zhu, M.; Liu, Y.; Wang, Y.; et al. Long non-coding RNA NEAT1-modulated abnormal lipolysis via ATGL drives hepatocellular carcinoma proliferation. Mol. Cancer 2018, 17, 90. [Google Scholar] [CrossRef]
- Sun, C.; Li, S.; Zhang, F.; Xi, Y.; Wang, L.; Bi, Y.; Li, D. Long non-coding RNA NEAT1 promotes non-small cell lung cancer progression through regulation of miR-377-3p-E2F3 pathway. Oncotarget 2016, 7, 51784–51814. [Google Scholar] [CrossRef]
- Kong, X.; Zhao, Y.; Li, X.; Tao, Z.; Hou, M.; Ma, H. Overexpression of HIF-2α-dependent NEAT1 promotes the progression of non-small cell lung cancer through miR-101-3p/SOX9/Wnt/β-catenin signal pathway. Cell. Physiol. Biochem. 2019, 52, 368–381. [Google Scholar] [CrossRef]
- Zhao, M.M.; Ge, L.Y.; Yang, L.F.; Zheng, H.X.; Chen, G.; Wu, L.Z.; Shi, S.M.; Wang, N.; Hang, Y.P. Lncrna neat1/mir-204/nuak1 axis is a potential therapeutic target for non-small cell lung cancer. Cancer Manag. Res. 2020, 12, 13357–13368. [Google Scholar] [CrossRef]
- Chen, L.M.; Niu, Y.D.; Xiao, M.; Li, X.J.; Lin, H. LncRNA NEAT1 regulated cell proliferation, invasion, migration and apoptosis by targeting has-miR-376b-3p/SULF1 axis in non-small cell lung cancer. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 4810–4821. [Google Scholar] [CrossRef]
- Luo, M.; Zhang, L.; Yang, H.; Luo, K.; Qing, C. Long non-coding RNA NEAT1 promotes ovarian cancer cell invasion and migration by interacting with miR-1321 and regulating tight junction protein 3 expression. Mol. Med. Rep. 2020, 22, 3429–3439. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Wang, Y. Long non-coding RNA NEAT1 facilitates the growth, migration, and invasion of ovarian cancer cells via the let-7 g/MEST/ATGL axis. Cancer Cell Int. 2021, 21, 437. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Sun, X.; Huang, Y.; Si, Q.; Li, M. Long non-coding RNA NEAT1 modifies cell proliferation, colony formation, apoptosis, migration and invasion via the miR-4500/BZW1 axis in ovarian cancer. Mol. Med. Rep. 2020, 22, 3347–3357. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; He, C.; Yang, F.; Qin, L.; Lu, X.; Wu, J. Long non-coding RNA-NEAT1, a sponge for MIR-98-5p, promotes expression of oncogene HMGA2 in prostate cancer. Biosci. Rep. 2019, 39, BSR20190635. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Guo, S.; Zhang, Y.; Zhao, Y.; Li, X.; Jia, Y.; Xu, Y.; Ma, B. LncRNA NEAT1 promotes docetaxel resistance in prostate cancer by regulating ACSL4 via sponging miR-34a-5p and miR-204-5p. Cell. Signal. 2020, 65, 109422. [Google Scholar] [CrossRef] [PubMed]
- Dong, P.; Xu, D.; Konno, Y.; Ihira, K.; Watari, H. Lncrna neat1-mediated mir-361 downregulation contributes to EMT and sphere formation of cervical cancer cells via increasing hsp90 expression. Int. J. Gynecol. Cancer 2020, 30, A128–A129. [Google Scholar] [CrossRef]
- Li, O.; Jiang, B.; Yi, W.M.; Zhang, Y.; Yang, P.Z.; Guo, C.; Sun, Z.P.; Peng, C. LncRNA NEAT1 promotes cell proliferation, migration, and invasion via the miR-186-5p/PTP4A1 axis in cholangiocarcinoma. Kaohsiung J. Med. Sci. 2021, 37, 379–391. [Google Scholar] [CrossRef]
- Fan, J.T.; Zhou, Z.Y.; Luo, Y.L.; Luo, Q.; Chen, S.B.; Zhao, J.C.; Chen, Q.R. Exosomal lncRNA NEAT1 from cancer-associated fibroblasts facilitates endometrial cancer progression via miR-26a/b-5p-mediated STAT3/YKL-40 signaling pathway. Neoplasia 2021, 23, 692–703. [Google Scholar] [CrossRef]
- Li, Y.; Chen, D.; Gao, X.; Li, X.; Shi, G. LncRNA NEAT1 Regulates Cell Viability and Invasion in Esophageal Squamous Cell Carcinoma through the miR-129/CTBP2 Axis. Dis. Markers 2017, 2017, 5314649. [Google Scholar] [CrossRef]
- Yang, F.; Tang, Z.; Duan, A.; Yi, B.; Shen, N.; Bo, Z.; Yin, L.; Zhu, B.; Qiu, Y.; Li, J. Long noncoding RNA NEAT1 upregulates survivin and facilitates gallbladder cancer progression by sponging microRNA-335. OncoTargets Ther. 2020, 13, 2357–2367. [Google Scholar] [CrossRef] [PubMed]
- Lulli, V.; Buccarelli, M.; Ilari, R.; Castellani, G.; De Dominicis, C.; Di Giamberardino, A.; D′Alessandris, Q.G.; Giannetti, S.; Martini, M.; Stumpo, V.; et al. Mir-370-3p impairs glioblastoma stem-like cell malignancy regulating a complex interplay between HMGA2/HIF1A and the oncogenic long non-coding RNA (LncRNA) neat1. Int. J. Mol. Sci. 2020, 21, 3610. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.B.; Yan, X.H.; Tian, M.; Zhang, S.; Liu, J.L.; Sheng, Y.X.; Dong, L.; Zhang, W.L. Long non-coding RNA NEAT1 regulates Hodgkin’s lymphoma cell proliferation and invasion via miR-448 mediated regulation of DCLK1. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 6219–6227. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Zheng, C.; Wu, S. Long non-coding RNA NEAT1 promotes the malignancy of laryngeal squamous cell carcinoma by regulating the microRNA-204-5p/SEMA4B axis. Oncol. Lett. 2021, 22, 802. [Google Scholar] [CrossRef]
- Zhou, W.J.; Wang, H.Y.; Zhang, J.; Dai, H.Y.; Yao, Z.X.; Zheng, Z.; Meng-Yan, S.; Wu, K. NEAT1/miR-200b-3p/SMAD2 axis promotes progression of melanoma. Aging 2020, 12, 22759–22775. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, C.; Chen, C.; Wu, F.; Shen, P.; Zhang, P.; He, G.; Li, X. Long non-coding RNA NEAT1 regulates epithelial membrane protein 2 expression to repress nasopharyngeal carcinoma migration and irradiation-resistance through miR-101-3p as a competing endogenous RNA mechanism. Oncotarget 2017, 8, 70156–70171. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Li, J.; Xiao, J.K.; Xiao, L.; Xu, B.W.; Li, C. The lncRNA NEAT1 promotes the epithelial-mesenchymal transition and metastasis of osteosarcoma cells by sponging miR-483 to upregulate STAT3 expression. Cancer Cell Int. 2021, 21, 90. [Google Scholar] [CrossRef]
- Hu, H.; Chen, W.; Zhang, S.; Xue, Y.; He, Y.; Gu, Y. NEAT1/miR-101-dependent up-regulation of DNA-PKcs enhances malignant behaviors of pancreatic ductal adenocarcinoma cells. J. Cancer 2021, 12, 5622–5632. [Google Scholar] [CrossRef]
- Chen, X.; Zhao, S.; Li, Q.; Xu, C.; Yu, Y.; Ge, H. LncRNA NEAT1 Knockdown Inhibits Retinoblastoma Progression by miR-3619-5p/LASP1 Axis. Front. Genet. 2020, 11, 574145. [Google Scholar] [CrossRef]
- Luo, Y.; Hao, T.; Zhang, J.; Zhang, M.; Sun, P.; Wu, L. MicroRNA-592 suppresses the malignant phenotypes of thyroid cancer by regulating lncRNA NEAT1 and downregulating NOVA1. Int. J. Mol. Med. 2019, 44, 1172–1182. [Google Scholar] [CrossRef]
- Li, Y.; Huang, W.Q.; Chen, L.L. LncRNA NEAT1 regulates proliferation, migration and invasion of tongue squamous cell carcinoma cells by regulating miR-339-5p/ITGA3 axis. Shanghai J. Stomatol. 2020, 29, 267–274. [Google Scholar]
- Shao, X.; Zheng, X.; Ma, D.; Liu, Y.; Liu, G. Inhibition of lncRNA-NEAT1 sensitizes 5-Fu resistant cervical cancer cells through de-repressing the microRNA-34a/LDHA axis. Biosci. Rep. 2021, 41, BSR20200533. [Google Scholar] [CrossRef] [PubMed]
- Rostami, M.; Kharajo, R.S.; Parsa-Kondelaji, M.; Ayatollahi, H.; Sheikhi, M.; Keramati, M.R. Altered expression of NEAT1 variants and P53, PTEN, and BCL-2 genes in patients with acute myeloid leukemia. Leuk. Res. 2022, 115, 106807. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Deng, S.; Pang, X.; Song, Y.; Luo, S.; Jin, L.; Pan, Y. LncRNA NEAT1 silenced miR-133b promotes migration and invasion of breast cancer cells. Int. J. Mol. Sci. 2019, 20, 3616. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Chen, Y.; Kong, W.; Li, Y.; Lu, F. Tumor protein D52 promotes breast cancer proliferation and migration via the long non-coding RNA NEAT1/microRNA-218-5p axis. Ann. Transl. Med. 2021, 9, 1008. [Google Scholar] [CrossRef]
- Lo, P.K.; Zhang, Y.; Wolfson, B.; Gernapudi, R.; Yao, Y.; Duru, N.; Zhou, Q. Dysregulation of the BRCA1/long non-coding RNA NEAT1 signaling axis contributes to breast tumorigenesis. Oncotarget 2016, 7, 65067–65089. [Google Scholar] [CrossRef]
- Shen, X.; Zhao, W.; Zhang, Y.; Liang, B. Long non-coding RNA-neat1 promotes cell migration and invasion via regulating MiR-124/NF-κB pathway in cervical cancer. OncoTargets Ther. 2020, 13, 3265–3276. [Google Scholar] [CrossRef]
- Yuan, L.Y.; Zhou, M.; Lv, H.; Qin, X.; Zhou, J.; Mao, X.; Li, X.; Xu, Y.; Liu, Y.; Xing, H. Involvement of NEAT1/miR-133a axis in promoting cervical cancer progression via targeting SOX4. J. Cell. Physiol. 2019, 234, 18985–18993. [Google Scholar] [CrossRef]
- Zhu, Z.; Du, S.; Yin, K.; Ai, S.; Yu, M.; Liu, Y.; Shen, Y.; Liu, M.; Jiao, R.; Chen, X.; et al. Knockdown long noncoding RNA nuclear paraspeckle assembly transcript 1 suppresses colorectal cancer through modulating miR-193a-3p/KRAS. Cancer Med. 2019, 8, 261–275. [Google Scholar] [CrossRef]
- Liu, Z.; Gu, Y.; Cheng, X.; Jiang, H.; Huang, Y.; Zhang, Y.; Yu, G.; Cheng, Y.; Zhou, L. Upregulation lnc-NEAT1 contributes to colorectal cancer progression through sponging miR-486-5p and activating NR4A1β-catenin pathway. Cancer Biomark. 2021, 30, 309–319. [Google Scholar] [CrossRef]
- Liu, H.; Li, A.; Sun, Z.; Zhang, J.; Xu, H. Long non-coding RNA NEAT1 promotes colorectal cancer progression by regulating miR-205-5p/VEGFA axis. Hum. Cell 2020, 33, 386–396. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Chen, J.J.; Lv, Q.; Qin, J.; Huang, Y.Z.; Yu, M.H.; Zhong, M. Long non-coding RNA NEAT1 promotes colorectal cancer progression by competitively binding miR-34a with SIRT1 and enhancing the Wnt/β-catenin signaling pathway. Cancer Lett. 2019, 440–441, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Zhai, J.; Fu, Y. Overexpression of Nuclear Enriched Autosomal Transcript 1 Facilitates Cell Proliferation, Migration Invasion, and Suppresses Apoptosis in Endometrial Cancer by Targeting MicroRNA-202-3p/T Cell Immunoglobulin and Mucin Domain 4 Axis. Cancer Biother. Radiopharm. 2020. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Ge, L.; Xu, X.J.; Yang, T.; Yuan, Y.; Ma, X.L.; Zhang, X.H. LncRNA NEAT1 promotes endometrial cancer cell proliferation, migration and invasion by regulating the MIR-144-3p/EZH2 axis. Radiol. Oncol. 2019, 53, 434–442. [Google Scholar] [CrossRef]
- Dong, P.; Xiong, Y.; Yue, J.; Xu, D.; Ihira, K.; Konno, Y.; Kobayashi, N.; Todo, Y.; Watari, H. Long noncoding RNA NEAT1 drives aggressive endometrial cancer progression via miR-361-regulated networks involving STAT3 and tumor microenvironment-related genes. J. Exp. Clin. Cancer Res. 2019, 38, 295. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhao, X.; Guo, Z.; Ma, X.; Song, Y.; Guo, Y. Regulation of NEAT1/miR-214-3p on the growth, migration and invasion of endometrial carcinoma cells. Arch. Gynecol. Obstet. 2017, 295, 1469–1475. [Google Scholar] [CrossRef]
- Luo, J.; Xie, K.; Gao, X.; Yao, Y.; Wang, G.; Shao, C.; Li, X.; Xu, Y.; Ren, B.; Hu, L.; et al. Long Noncoding RNA Nuclear Paraspeckle Assembly Transcript 1 Promotes Progression and Angiogenesis of Esophageal Squamous Cell Carcinoma Through miR-590-3p/MDM2 Axis. Front. Oncol. 2020, 10, 618930. [Google Scholar] [CrossRef]
- Yang, L.; Wang, M.; He, P. Lncrna neat1 promotes the progression of gastric cancer through modifying the mir-1224-5p/rsf1 signaling axis. Cancer Manag. Res. 2020, 12, 11845–11855. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, M.; Sun, G. Long non-coding RNA NEAT1 regulates the proliferation, migration and invasion of gastric cancer cells via targeting miR-335-5p/ROCK1 axis. Pharmazie 2018, 73, 150–155. [Google Scholar] [CrossRef]
- Li, Y.; Ding, X.; Xiu, S.; Du, G.; Liu, Y. LncRNA neat1 promotes proliferation, migration and invasion via regulating miR-296-5p/CNN2 axis in hepatocellular carcinoma cells. OncoTargets Ther. 2019, 12, 9887–9897. [Google Scholar] [CrossRef]
- Xia, Y.; Zhou, Y.; Han, H.; Li, P.; Wei, W.; Lin, N. lncRNA NEAT1 facilitates melanoma cell proliferation, migration, and invasion via regulating miR-495-3p and E2F3. J. Cell. Physiol. 2019, 234, 19592–19601. [Google Scholar] [CrossRef] [PubMed]
- Ding, F.; Lai, J.; Gao, Y.; Wang, G.; Shang, J.; Zhang, D.; Zheng, S. NEAT1/miR-23a-3p/KLF3: A novel regulatory axis in melanoma cancer progression. Cancer Cell Int. 2019, 19, 217. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Fang, P.; Li, W.J.; Zhang, J.; Wang, G.P.; Jiang, D.F.; Chen, F.P. LncRNA NEAT1 sponges miR-214 to regulate M2 macrophage polarization by regulation of B7-H3 in multiple myeloma. Mol. Immunol. 2020, 117, 20–28. [Google Scholar] [CrossRef]
- Wang, H.; Jiang, Y.; Fu, L.; Zhu, X. Mir-222 emerges as a cerna of lncrna neat1, and inhibits cell viability and migration of nasopharyngeal carcinoma cells via aldh1. Basic Clin. Pharmacol. Toxicol. 2021, 128, 94–95. [Google Scholar]
- Qi, L.; Liu, F.; Zhang, F.; Zhang, S.; Lv, L.; Bi, Y.; Yu, Y. lncRNA NEAT1 competes against let-7a to contribute to non-small cell lung cancer proliferation and metastasis. Biomed. Pharmacother. 2018, 103, 1507–1515. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Mo, Q.; Wan, X.; Dan, J.; Hu, H. NEAT1/hsa-mir-98-5p/MAPK6 axis is involved in non–small-cell lung cancer development. J. Cell. Biochem. 2019, 120, 2836–2846. [Google Scholar] [CrossRef]
- Liu, J.; Li, K.; Wang, R.; Chen, S.; Wu, J.; Li, X.; Ning, Q.; Yang, G.; Pang, Y. The interplay between ATF2 and NEAT1 contributes to lung adenocarcinoma progression. Cancer Cell Int. 2020, 20, 594. [Google Scholar] [CrossRef]
- Yu, P.F.; Wang, Y.; Lv, W.; Kou, D.; Hu, H.L.; Guo, S.S.; Zhao, Y.J. LncRNA NEAT1/MIR-1224/KLF3 contributes to cell proliferation, apoptosis and invasion in lung cancer. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 8403–8410. [Google Scholar] [CrossRef]
- Zhang, L.; Lu, X.Q.; Zhou, X.Q.; Liu, Q.B.; Chen, L.; Cai, F. NEAT1 induces osteosarcoma development by modulating the miR-339-5p/TGF-β1 pathway. J. Cell. Physiol. 2019, 234, 5097–5105. [Google Scholar] [CrossRef]
- Tan, H.; Zhao, L. lncRNA nuclear-enriched abundant transcript 1 promotes cell proliferation and invasion by targeting miR-186-5p/HIF-1α in osteosarcoma. J. Cell. Biochem. 2019, 120, 6502–6514. [Google Scholar] [CrossRef]
- Cao, J.; Zhang, Y.; Yang, J.; He, S.; Li, M.; Yan, S.; Chen, Y.; Qu, C.; Xu, L. NEAT1 regulates pancreatic cancer cell growth, invasion and migration though mircroRNA-335-5p/c-met axis. Am. J. Cancer Res. 2016, 6, 2361–2374. [Google Scholar] [PubMed]
- Luan, L.; Hu, Q.; Wang, Y.; Lu, L.; Ling, J. Knockdown of lncRNA NEAT1 expression inhibits cell migration, invasion and EMT by regulating the miR-24-3p/LRG1 axis in retinoblastoma cells. Exp. Ther. Med. 2021, 21, 367. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Zhang, Z.; Lu, Y.; Xiu, W.; Cui, J. Lncrna neat1 acts as an mir-148b-3p sponge to regulate rock1 inhibition of retinoblastoma growth. Cancer Manag. Res. 2021, 13, 5587–5597. [Google Scholar] [CrossRef] [PubMed]
- Zhong, W.; Yang, J.; Li, M.; Li, L.; Li, A. Long noncoding RNA NEAT1 promotes the growth of human retinoblastoma cells via regulation of miR-204/CXCR4 axis. J. Cell. Physiol. 2019, 234, 11567–11576. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Cai, Y.; Zheng, L.; Zhang, Z.; Lin, X.; Jiang, N. Long noncoding RNA NEAT1 regulate papillary thyroid cancer progression by modulating miR-129-5p/KLK7 expression. J. Cell. Physiol. 2018, 233, 6638–6648. [Google Scholar] [CrossRef]
- Sun, W.; Lan, X.; Zhang, H.; Wang, Z.; Dong, W.; He, L.; Zhang, T.; Zhang, P.; Liu, J.; Qin, Y. NEAT1-2 functions as a competing endogenous RNA to regulate ATAD2 expression by sponging microRNA-106b-5p in papillary thyroid cancer. Cell Death Dis. 2018, 9, 380. [Google Scholar] [CrossRef]
- Pan, S.; Liu, R.; Wu, X.; Ma, K.; Luo, W.; Nie, K.; Zhang, C.; Meng, X.; Tong, T.; Chen, X.; et al. LncRNA NEAT1 mediates intestinal inflammation by regulating TNFRSF1B. Ann. Transl. Med. 2021, 9, 773. [Google Scholar] [CrossRef]
- Feng, Y.; Gao, L.; Cui, G.; Cao, Y. LncRNA NEAT1 facilitates pancreatic cancer growth and metastasis through stabilizing ELF3 mRNA. Am. J. Cancer Res. 2020, 10, 237–248. [Google Scholar]
- Ma, F.; Lei, Y.Y.; Ding, M.G.; Luo, L.H.; Xie, Y.C.; Liu, X.L. LncRNA NEAT1 Interacted With DNMT1 to Regulate Malignant Phenotype of Cancer Cell and Cytotoxic T Cell Infiltration via Epigenetic Inhibition of p53, cGAS, and STING in Lung Cancer. Front. Genet. 2020, 11, 250. [Google Scholar] [CrossRef]
- Wang, S.; Zuo, H.; Jin, J.; Lv, W.; Xu, Z.; Fan, Y.; Zhang, J.; Zuo, B. Long noncoding RNA Neat1 modulates myogenesis by recruiting Ezh2. Cell Death Dis. 2019, 10, 505. [Google Scholar] [CrossRef]
- Chen, Q.; Cai, J.; Wang, Q.; Wang, Y.; Liu, M.; Yang, J.; Zhou, J.; Kang, C.; Li, M.; Jiang, C. Long Noncoding RNA NEAT1, Regulated by the EGFR Pathway, Contributes to Glioblastoma Progression Through the WNT/beta-Catenin Pathway by Scaffolding EZH2. Clin. Cancer Res. 2018, 24, 684–695. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Liu, L.; Zhang, S.; Ming, Y.; Liu, S.; Cheng, K.; Zhao, Y. Long noncoding RNA NEAT1 suppresses hepatocyte proliferation in fulminant hepatic failure through increased recruitment of EZH2 to the LATS2 promoter region and promotion of H3K27me3 methylation. Exp. Mol. Med. 2020, 52, 461–472. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.S.I.; Dong, K.; Liu, J.; Wen, T.; Yu, L.; Xu, F.; Kang, X.; Osman, I.; Hu, G.; Bunting, K.M.; et al. Long noncoding RNA NEAT1 (nuclear paraspeckle assembly transcript 1) is critical for phenotypic switching of vascular smooth muscle cells. Proc. Natl. Acad. Sci. USA 2018, 115, E8660–E8667. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhao, Y.; Xu, N.; Zhang, S.; Wang, S.; Mao, Y.; Zhu, Y.; Li, B.; Jiang, Y.; Tan, Y.; et al. NEAT1 regulates neuroglial cell mediating Abeta clearance via the epigenetic regulation of endocytosis-related genes expression. Cell Mol. Life Sci. 2019, 76, 3005–3018. [Google Scholar] [CrossRef] [PubMed]
- Ma, P.; Pan, Y.; Yang, F.; Fang, Y.; Liu, W.; Zhao, C.; Yu, T.; Xie, M.; Jing, X.; Wu, X.; et al. KLF5-Modulated lncRNA NEAT1 Contributes to Tumorigenesis by Acting as a Scaffold for BRG1 to Silence GADD45A in Gastric Cancer. Mol. Nucleic Acids 2020, 22, 382–395. [Google Scholar] [CrossRef]
- Park, M.K.; Zhang, L.; Min, K.W.; Cho, J.H.; Yeh, C.C.; Moon, H.; Hormaechea-Agulla, D.; Mun, H.; Ko, S.; Lee, J.W.; et al. NEAT1 is essential for metabolic changes that promote breast cancer growth and metastasis. Cell Metab. 2021, 33, 2380–2397. [Google Scholar] [CrossRef]
- Wen, S.; Wei, Y.; Zen, C.; Xiong, W.; Niu, Y.; Zhao, Y. Long non-coding RNA NEAT1 promotes bone metastasis of prostate cancer through N6-methyladenosine. Mol. Cancer 2020, 19, 171. [Google Scholar] [CrossRef]
- Huang, Z.; Zhao, J.; Wang, W.; Zhou, J.; Zhang, J. Depletion of LncRNA NEAT1 Rescues Mitochondrial Dysfunction Through NEDD4L-Dependent PINK1 Degradation in Animal Models of Alzheimer’s Disease. Front. Cell. Neurosci. 2020, 14, 28. [Google Scholar] [CrossRef]
- Xu, Y.; Cao, Z.; Ding, Y.; Li, Z.; Xiang, X.; Lai, R.; Sheng, Z.; Liu, Y.; Cai, W.; Hu, R.; et al. Long Non-coding RNA NEAT1 Alleviates Acute-on-Chronic Liver Failure Through Blocking TRAF6 Mediated Inflammatory Response. Front. Physiol. 2019, 10, 1503. [Google Scholar] [CrossRef]
- Shui, X.; Chen, S.; Lin, J.; Kong, J.; Zhou, C.; Wu, J. Knockdown of lncRNA NEAT1 inhibits Th17/CD4(+) T cell differentiation through reducing the STAT3 protein level. J. Cell Physiol. 2019, 234, 22477–22484. [Google Scholar] [CrossRef]
- Li, X.; Wang, X.; Song, W.; Xu, H.; Huang, R.; Wang, Y.; Zhao, W.; Xiao, Z.; Yang, X. Oncogenic Properties of NEAT1 in Prostate Cancer Cells Depend on the CDC5L-AGRN Transcriptional Regulation Circuit. Cancer Res. 2018, 78, 4138–4149. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Fan, P.; Zhao, Y.; Zhang, S.; Lu, J.; Xie, W.; Jiang, Y.; Lei, F.; Xu, N.; Zhang, Y. NEAT1 modulates herpes simplex virus-1 replication by regulating viral gene transcription. Cell Mol. Life Sci. 2017, 74, 1117–1131. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Zhu, J.; Zhang, L.; Shu, J. Long non-coding RNA NEAT1 promotes steatosis via enhancement of estrogen receptor alpha-mediated AQP7 expression in HepG2 cells. Artif. Cells Nanomed. Biotechnol. 2019, 47, 1782–1787. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Shao, C.; Wu, Q.J.; Chen, G.; Zhou, J.; Yang, B.; Li, H.; Gou, L.T.; Zhang, Y.; Wang, Y.; et al. NEAT1 scaffolds RNA-binding proteins and the Microprocessor to globally enhance pri-miRNA processing. Nat. Struct. Mol. Biol. 2017, 24, 816–824. [Google Scholar] [CrossRef]
- Shen, W.; Liang, X.H.; Sun, H.; De Hoyos, C.L.; Crooke, S.T. Depletion of NEAT1 lncRNA attenuates nucleolar stress by releasing sequestered P54nrb and PSF to facilitate c-Myc translation. PLoS ONE 2017, 12, e0173494. [Google Scholar] [CrossRef]
- Jen, H.W.; Gu, D.L.; Lang, Y.D.; Jou, Y.S. PSPC1 Potentiates IGF1R Expression to Augment Cell Adhesion and Motility. Cells 2020, 9, 1490. [Google Scholar] [CrossRef]
- Cooper, D.R.; Carter, G.; Li, P.; Patel, R.; Watson, J.E.; Patel, N.A. Long Non-Coding RNA NEAT1 Associates with SRp40 to Temporally Regulate PPARγ2 Splicing during Adipogenesis in 3T3-L1 Cells. Genes 2014, 5, 1050–1063. [Google Scholar] [CrossRef]
- Mang, Y.; Li, L.; Ran, J.; Zhang, S.; Liu, J.; Li, L.; Chen, Y.; Liu, J.; Gao, Y.; Ren, G. Long noncoding RNA NEAT1 promotes cell proliferation and invasion by regulating hnRNP A2 expression in hepatocellular carcinoma cells. Oncol. Targets 2017, 10, 1003–1016. [Google Scholar] [CrossRef]
- Lee, M.; Sadowska, A.; Bekere, I.; Ho, D.; Gully, B.S.; Lu, Y.; Iyer, K.S.; Trewhella, J.; Fox, A.H.; Bond, C.S. The structure of human SFPQ reveals a coiled-coil mediated polymer essential for functional aggregation in gene regulation. Nucleic Acids Res. 2015, 43, 3826–3840. [Google Scholar] [CrossRef]
- Naganuma, T.; Nakagawa, S.; Tanigawa, A.; Sasaki, Y.F.; Goshima, N.; Hirose, T. Alternative 3’-end processing of long noncoding RNA initiates construction of nuclear paraspeckles. EMBO J. 2012, 31, 4020–4034. [Google Scholar] [CrossRef]
- West, J.A.; Davis, C.P.; Sunwoo, H.; Simon, M.D.; Sadreyev, R.I.; Wang, P.I.; Tolstorukov, M.Y.; Kingston, R.E. The long noncoding RNAs NEAT1 and MALAT1 bind active chromatin sites. Mol. Cell 2014, 55, 791–802. [Google Scholar] [CrossRef] [PubMed]
- Chu, C.; Qu, K.; Zhong, F.L.; Artandi, S.E.; Chang, H.Y. Genomic maps of long noncoding RNA occupancy reveal principles of RNA-chromatin interactions. Mol. Cell 2011, 44, 667–678. [Google Scholar] [CrossRef] [PubMed]
- Jen, J.; Tang, Y.A.; Lu, Y.H.; Lin, C.C.; Lai, W.W.; Wang, Y.C. Oct4 transcriptionally regulates the expression of long non-coding RNAs NEAT1 and MALAT1 to promote lung cancer progression. Mol. Cancer 2017, 16, 104. [Google Scholar] [CrossRef] [PubMed]
- Choudhry, H.; Albukhari, A.; Morotti, M.; Haider, S.; Moralli, D.; Smythies, J.; Schödel, J.; Green, C.M.; Camps, C.; Buffa, F.; et al. Tumor hypoxia induces nuclear paraspeckle formation through HIF-2α dependent transcriptional activation of NEAT1 leading to cancer cell survival. Oncogene 2015, 34, 4482–4490. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zheng, Y.; Sun, Y.; Li, S.; Chen, L.; Jin, X.; Hou, X.; Liu, X.; Chen, Q.; Li, J.; et al. Knockdown of NEAT1 induces tolerogenic phenotype in dendritic cells by inhibiting activation of NLRP3 inflammasome. Theranostics 2019, 9, 3425–3442. [Google Scholar] [CrossRef]
- Modic, M.; Grosch, M.; Rot, G.; Schirge, S.; Lepko, T.; Yamazaki, T.; Lee, F.C.Y.; Rusha, E.; Shaposhnikov, D.; Palo, M.; et al. Cross-Regulation between TDP-43 and Paraspeckles Promotes Pluripotency-Differentiation Transition. Mol. Cell 2019, 74, 951–965. [Google Scholar] [CrossRef]
- Sunwoo, H.; Dinger, M.E.; Wilusz, J.E.; Amaral, P.P.; Mattick, J.S.; Spector, D.L. MEN epsilon/beta nuclear-retained non-coding RNAs are up-regulated upon muscle differentiation and are essential components of paraspeckles. Genome Res. 2009, 19, 347–359. [Google Scholar] [CrossRef]
- Klec, C.; Knutsen, E.; Schwarzenbacher, D.; Jonas, K.; Pasculli, B.; Heitzer, E.; Rinner, B.; Krajina, K.; Prinz, F.; Gottschalk, B.; et al. ALYREF, a novel factor involved in breast carcinogenesis, acts through transcriptional and post-transcriptional mechanisms selectively regulating the short NEAT1 isoform. Cell Mol. Life Sci. 2022, 79, 391. [Google Scholar] [CrossRef]
- Wu, Y.; Yang, L.; Zhao, J.; Li, C.; Nie, J.; Liu, F.; Zhuo, C.; Zheng, Y.; Li, B.; Wang, Z.; et al. Nuclear-enriched abundant transcript 1 as a diagnostic and prognostic biomarker in colorectal cancer. Mol. Cancer 2015, 14, 191. [Google Scholar] [CrossRef]
- Zhang, H.; Su, X.; Burley, S.K.; Zheng, X.F.S. mTOR regulates aerobic glycolysis through NEAT1 and nuclear paraspeckle-mediated mechanism in hepatocellular carcinoma. Theranostics 2022, 12, 3518–3533. [Google Scholar] [CrossRef]
- Chai, Y.; Liu, J.; Zhang, Z.; Liu, L. HuR-regulated lncRNA NEAT1 stability in tumorigenesis and progression of ovarian cancer. Cancer Med. 2016, 5, 1588–1598. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Li, X.; Yu, L.; Wang, R.; Hua, D.; Shi, C.; Sun, C.; Luo, W.; Rao, C.; Jiang, Z.; et al. The RNA-binding protein SRSF1 is a key cell cycle regulator via stabilizing NEAT1 in glioma. Int. J. Biochem. Cell Biol. 2019, 113, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Dong, F.; Qin, X.; Wang, B.; Li, Q.; Hu, J.; Cheng, X.; Guo, D.; Cheng, F.; Fang, C.; Tan, Y.; et al. ALKBH5 Facilitates Hypoxia-Induced Paraspeckle Assembly and IL8 Secretion to Generate an Immunosuppressive Tumor Microenvironment. Cancer Res. 2021, 81, 5876–5888. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Deng, Y.; Zhu, J.; Duan, Y.; Weng, W.; Xu, M.; Ju, X.; Wu, X. Long noncoding RNA NEAT1, regulated by LIN28B, promotes cell proliferation and migration through sponging miR-506 in high-grade serous ovarian cancer. Cell Death Dis. 2018, 9, 861. [Google Scholar] [CrossRef]
- Yi, K.; Cui, X.; Liu, X.; Wang, Y.; Zhao, J.; Yang, S.; Xu, C.; Yang, E.; Xiao, M.; Hong, B.; et al. PTRF/Cavin-1 as a Novel RNA-Binding Protein Expedites the NF-κB/PD-L1 Axis by Stabilizing lncRNA NEAT1, Contributing to Tumorigenesis and Immune Evasion in Glioblastoma. Front. Immunol. 2021, 12, 802795. [Google Scholar] [CrossRef]
- Yoon, J.H.; De, S.; Srikantan, S.; Abdelmohsen, K.; Grammatikakis, I.; Kim, J.; Kim, K.M.; Noh, J.H.; White, E.J.; Martindale, J.L.; et al. PAR-CLIP analysis uncovers AUF1 impact on target RNA fate and genome integrity. Nat. Commun. 2014, 5, 5248. [Google Scholar] [CrossRef]
- Rao, Y.; Fang, Y.; Tan, W.; Liu, D.; Pang, Y.; Wu, X.; Zhang, C.; Li, G. Delivery of Long Non-coding RNA NEAT1 by Peripheral Blood Monouclear Cells-Derived Exosomes Promotes the Occurrence of Rheumatoid Arthritis via the MicroRNA-23a/MDM2/SIRT6 Axis. Front. Cell Dev. Biol. 2020, 8, 551681. [Google Scholar] [CrossRef]
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef]
- Jia, X.; Wei, L.; Zhang, Z. NEAT1 Overexpression Indicates a Poor Prognosis and Induces Chemotherapy Resistance via the miR-491-5p/SOX3 Signaling Pathway in Ovarian Cancer. Front. Genet. 2021, 12, 616220. [Google Scholar] [CrossRef]
- Mo, C.; Huang, B.; Zhuang, J.; Jiang, S.; Guo, S.; Mao, X. LncRNA nuclear-enriched abundant transcript 1 shuttled by prostate cancer cells-secreted exosomes initiates osteoblastic phenotypes in the bone metastatic microenvironment via miR-205-5p/runt-related transcription factor 2/splicing factor proline- and glutamine-rich/polypyrimidine tract-binding protein 2 axis. Clin. Transl. Med. 2021, 11, e493. [Google Scholar] [CrossRef]
- Zhuang, L.; Xia, W.; Chen, D.; Ye, Y.; Hu, T.; Li, S.; Hou, M. Exosomal LncRNA-NEAT1 derived from MIF-treated mesenchymal stem cells protected against doxorubicin-induced cardiac senescence through sponging miR-221-3p. J. Nanobiotechnol. 2020, 18, 157. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.L.; Carmichael, G.G. Altered nuclear retention of mRNAs containing inverted repeats in human embryonic stem cells: Functional role of a nuclear noncoding RNA. Mol. Cell 2009, 35, 467–478. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Cancer Type | Sponged miRNA | Regulated mRNA | Ref. |
|---|---|---|---|
| Acute Myeloid Leukemia | 338-3p | CREBRF | [17] |
| Breast Cancer | 133b | TIMM17A | [94] |
| 141-3p | KLF12 | [41] | |
| 218-5p | TPD52 | [95] | |
| 138-5p | ZFX | [40] | |
| 410-3p | Cyclin D1 | [38] | |
| 107 | CPT1A | [42] | |
| 448 | ZEB1 | [37] | |
| 211 | HMGA2 | [39] | |
| 129-5p | WNT4 | [96] | |
| Cervical cancer | 361 | Hsp90 | [77] |
| 124 | NF-κB | [97] | |
| 133a | SOX4 | [98] | |
| 34a | LDHA | [92] | |
| Cholangiocarcinoma | 186-5p | PTP4A1 | [78] |
| Colorectal Cancer | let-7 g-5p | BACH1 | [47] |
| 185-5p | IGF2 | [46] | |
| 150-5p | CPSF4 | [15] | |
| 196a-5p | GDNF | [44] | |
| 193a-3p | KRAS | [99] | |
| 486-5p | NR4A1β | [100] | |
| 205-5p | VEGFA | [101] | |
| 195-5p | CEP55 | [43] | |
| 34a | SIRT1 | [102] | |
| Endometrial Cancer | 202-3p | TIMD4 | [103] |
| 144-3p | EZH2 | [104] | |
| 361 | MEF2D, ROCK1, WNT7A, STAT3, VEGFA, PDE4B and KPNA4 | [105] | |
| 214-3p | HMGA1 | [106] | |
| Esophageal Carcinoma | 590-3p | MDM2 | [107] |
| 129 | CTBP2 | [80] | |
| Gallbladder Cancer | 335 | Survivin | [81] |
| Gastric Cancer | 365a-3p | ABCC4 | [49] |
| 142-5p | JAG1 | [53] | |
| 221-5p | SOX11 | [54] | |
| 30a-3p | COX-2, BCL9 | [55] | |
| 1224-5p | RSF1 | [108] | |
| 1294 | AKT1 | [48] | |
| 506 | STAT3 | [52] | |
| 335-5p | ROCK1 | [109] | |
| 500a-3p | XBP-1 | [51] | |
| Glioblastoma | 370-3p | HIF1A | [82] |
| Glioma | 185-5p | DNMT1 | [58] |
| 152-3p | CCT6A | [56] | |
| 139-5p | CDK6 | [60] | |
| 132 | SOX2 | [57] | |
| 449b-5p | c-Met | [59] | |
| Hepatocellular Carcinoma | 503 | SMO | [63] |
| 320a | LAGE3 | [66] | |
| 22-3p | AKT2 | [64] | |
| 296-5p | Calponin 2 | [110] | |
| 139-5p | TGF-β1 | [65] | |
| 485 | STAT3 | [61] | |
| 124-3p | ATGL1 | [67] | |
| Hodgkin’s Lymphoma | 448 | DCLK1 | [83] |
| Laryngeal Carcinoma | 204-5p | SEMA4B | [84] |
| Melanoma | 495-3p | E2F3 | [111] |
| 200b-3p | SMAD2 | [85] | |
| 23a-3p | KLF3 | [112] | |
| Multiple Myeloma | 214 | B7-H3 | [113] |
| Nasopharyngeal Carcinoma | 222 | ALDH1 | [114] |
| 101-3p | EMP2 | [86] | |
| Lung Cancer | 204 | NUAK1 | [70] |
| 376b-3p | SULF1 | [71] | |
| 101-3p | SOX9 | [69] | |
| let-7a | IGF2 | [115] | |
| 377-3p | E2F3 | [68] | |
| 98-5p | MAPK6 | [116] | |
| 26a-5p | ATF2 | [117] | |
| 1224 | KLF3 | [118] | |
| Osteosarcoma | 483 | STAT3 | [87] |
| 339-5p | TGF-β1 | [119] | |
| 186-5p | HIF-1α | [120] | |
| Ovarian Cancer | 1321 | TJP3 | [72] |
| let-7 g | MEST | [73] | |
| 4500 | BZW1 | [74] | |
| 382-3p | ROCK1 | [10] | |
| Pancreatic Carcinoma | 101 | DNA-PKcs | [88] |
| 302a-3p | RELA | [12] | |
| 335-5p | c-Met | [121] | |
| Prostate Cancer | 34a-5p | ACSL4 | [76] |
| 204-5p | ACSL4 | [76] | |
| 98-5p | HMGA2 | [75] | |
| Retinoblastoma | 24-3p | LRG1 | [122] |
| 148b-3p | ROCK1 | [123] | |
| 3619-5p | LASP1 | [89] | |
| Retinoblastoma | 204 | CXCR4 | [124] |
| Thyroid Cancer | 491 | TGM2 | [35] |
| 592 | NOVA1 | [90] | |
| 129-5p | KLK7 | [125] | |
| 106b-5p | ATAD2 | [126] | |
| Tongue Carcinoma | 339-5p | ITGA3 | [91] |
| RBPs | Downstream Target | Function | Ref. |
|---|---|---|---|
| EZH2 | H3K27 | Trimethylation of H3K27, transcription repression | [131] |
| WDR5 | H3k4 | Trimethylation of H3K4, transcription activation | [133] |
| P300/CBP | H3K27 | Acetylation and decrotonylation, transcription activation | [134] |
| BRG1 | H3K27, H3K4 | Demethylation of H3K4 and trimethylation of H3K27, transcription inactivation | [135] |
| DNMT1 | CGAS, STING and P53 promoter | CpG island methylation, transcription repression | [129] |
| PGK, PGAM, ENO | 1,3-Biphosphoglyceric acid | Glycolysis upregulation | [136] |
| CDK19, CyclinL1 | RNA Pol II | RNA pol II phosphorylation, transcription activation | [137] |
| NEDD4L | PINK1 | Promoting ubiquitination degradation of PINK1 | [138] |
| TRAF6 | - | Inhibiting auto-ubiquitination of TRAF6 | [139] |
| STAT3 | - | Inhibiting ubiquitination degradation of STAT3 | [140] |
| SIN3A, FOXN3 | GATA3, TJP1 promoter | Transcription downregulation | [36] |
| CDC5L | AGRN promoter | Transcription upregulation | [141] |
| STAT3 | ICP0, TK promoter | Transcription upregulation | [142] |
| ERα | AQP7 promoter | Transcription upregulation | [143] |
| SFPQ | IL-8 promoter | Transcription downregulation | [32] |
| ADARB2 promoter | Transcription upregulation | [20] | |
| Drosha/DGCR8 | miRNA processing | [144] | |
| c-Myc mRNA | Transcription upregulation | [145] | |
| DDX5 | β-catenin | Promoting Wnt-β-catenin pathway | [13] |
| PSPC1 | - | IGF1R induction | [146] |
| SRSF 5 | PPAR-γ mRNA | mRNA splicing | [147] |
| U2AF65 | hnRNP A2 mRNA | Increasing hnRNP A2 expression | [148] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gu, J.; Zhang, B.; An, R.; Qian, W.; Han, L.; Duan, W.; Wang, Z.; Ma, Q. Molecular Interactions of the Long Noncoding RNA NEAT1 in Cancer. Cancers 2022, 14, 4009. https://doi.org/10.3390/cancers14164009
Gu J, Zhang B, An R, Qian W, Han L, Duan W, Wang Z, Ma Q. Molecular Interactions of the Long Noncoding RNA NEAT1 in Cancer. Cancers. 2022; 14(16):4009. https://doi.org/10.3390/cancers14164009
Chicago/Turabian StyleGu, Jingtao, Bo Zhang, Rui An, Weikun Qian, Liang Han, Wanxing Duan, Zheng Wang, and Qingyong Ma. 2022. "Molecular Interactions of the Long Noncoding RNA NEAT1 in Cancer" Cancers 14, no. 16: 4009. https://doi.org/10.3390/cancers14164009
APA StyleGu, J., Zhang, B., An, R., Qian, W., Han, L., Duan, W., Wang, Z., & Ma, Q. (2022). Molecular Interactions of the Long Noncoding RNA NEAT1 in Cancer. Cancers, 14(16), 4009. https://doi.org/10.3390/cancers14164009