
Microbiota and the Immune System—Actors in the Gastric Cancer Story

 , and
, and 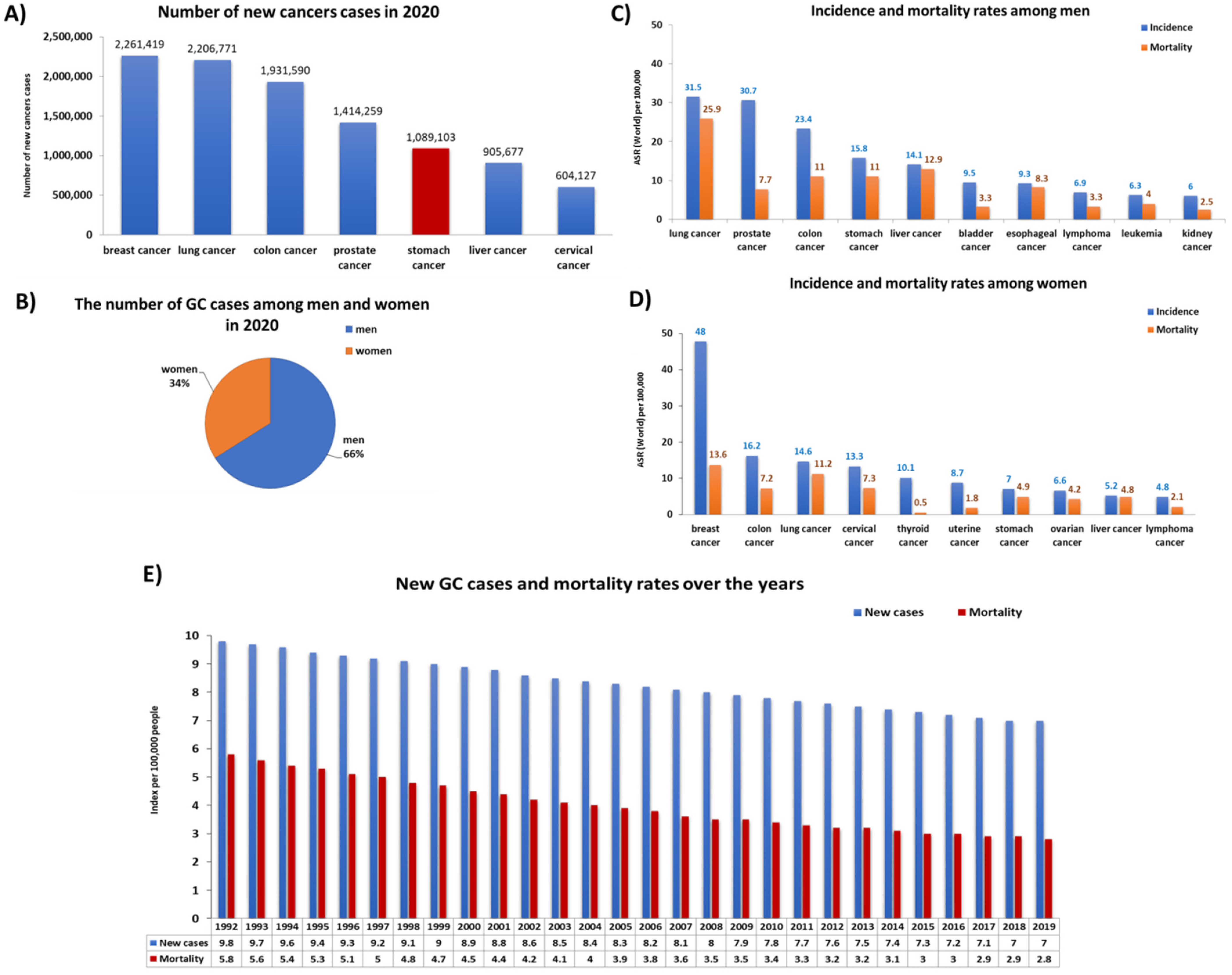{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
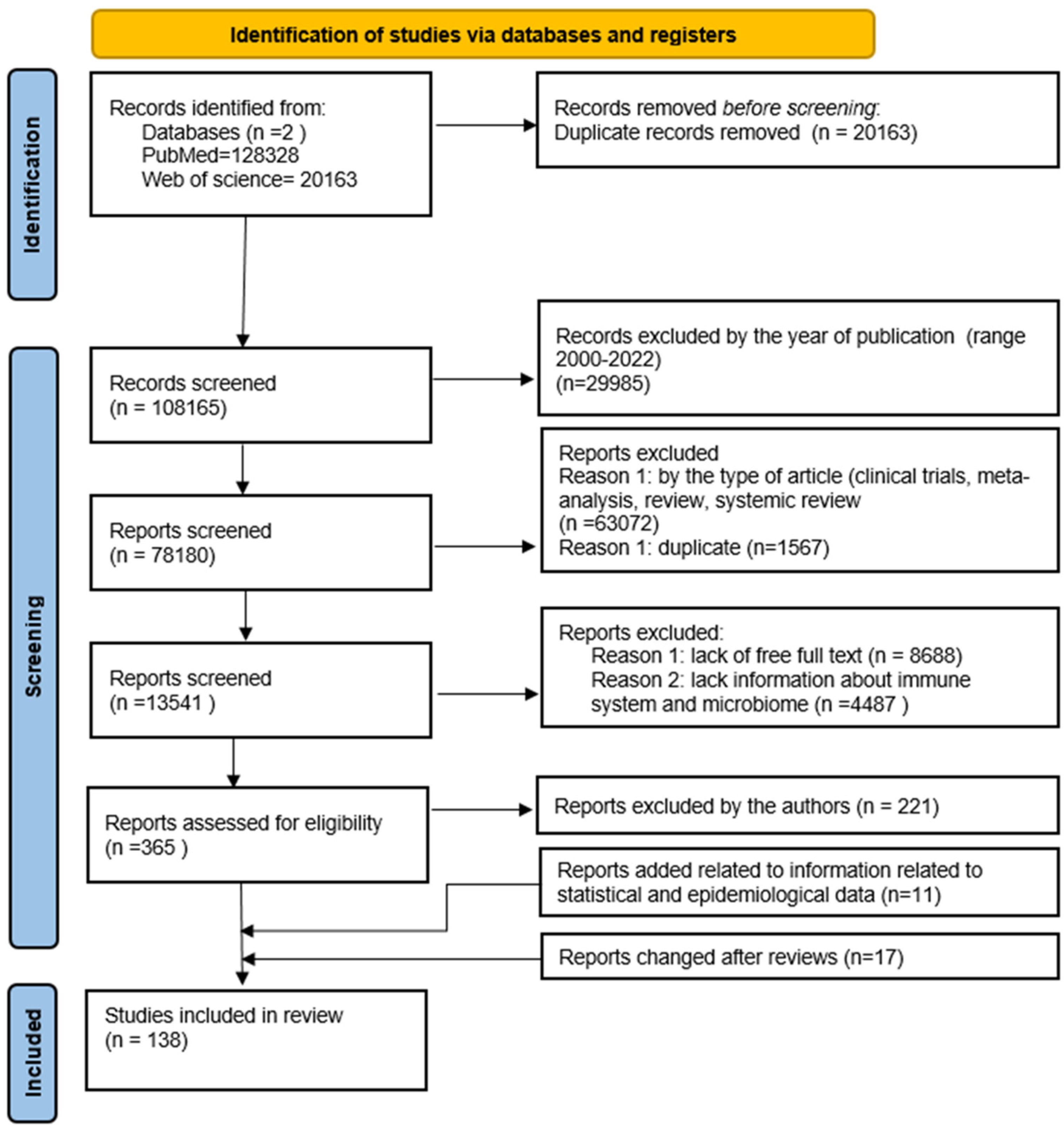
2. Materials and Methods
Search Strategy, Study Selection, and Data Extraction
3. Gastric Cancer—Risk Factors, Symptoms, and Classification
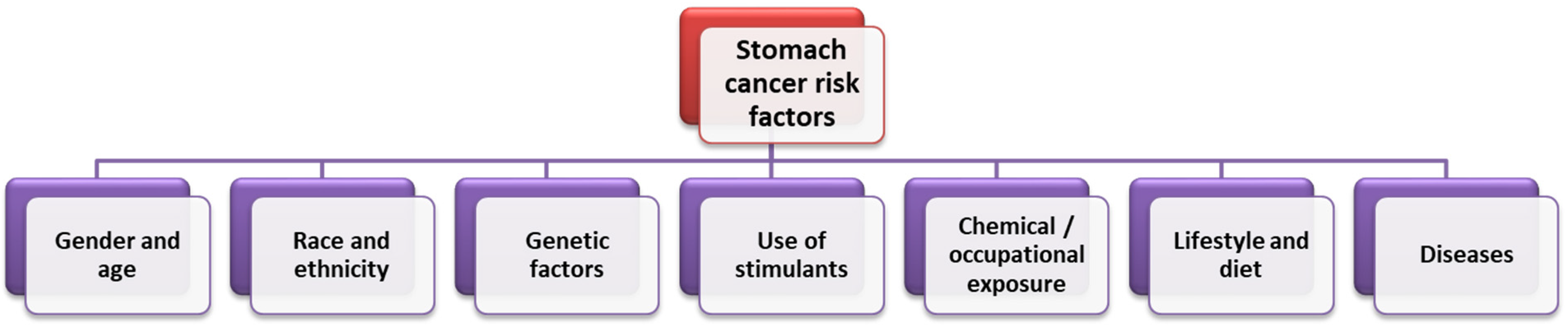
3.1. Selected Risk Factors for the Development of Gastric Cancer
3.1.1. Genetic Factors
3.1.2. Lifestyle and Diet
3.1.3. H. pylori Infection, Obesity, and other Diseases
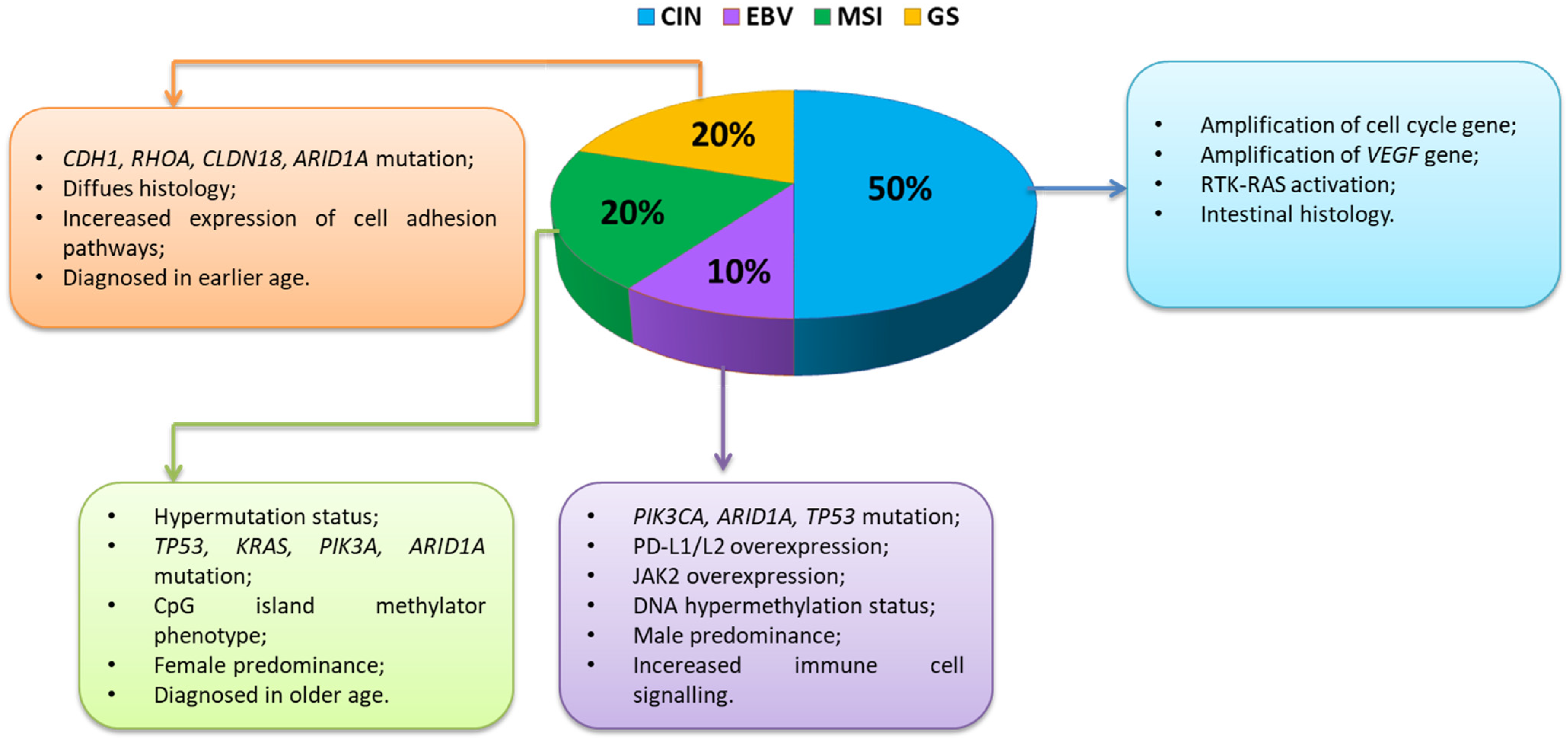
3.2. Classification of Gastric Cancer
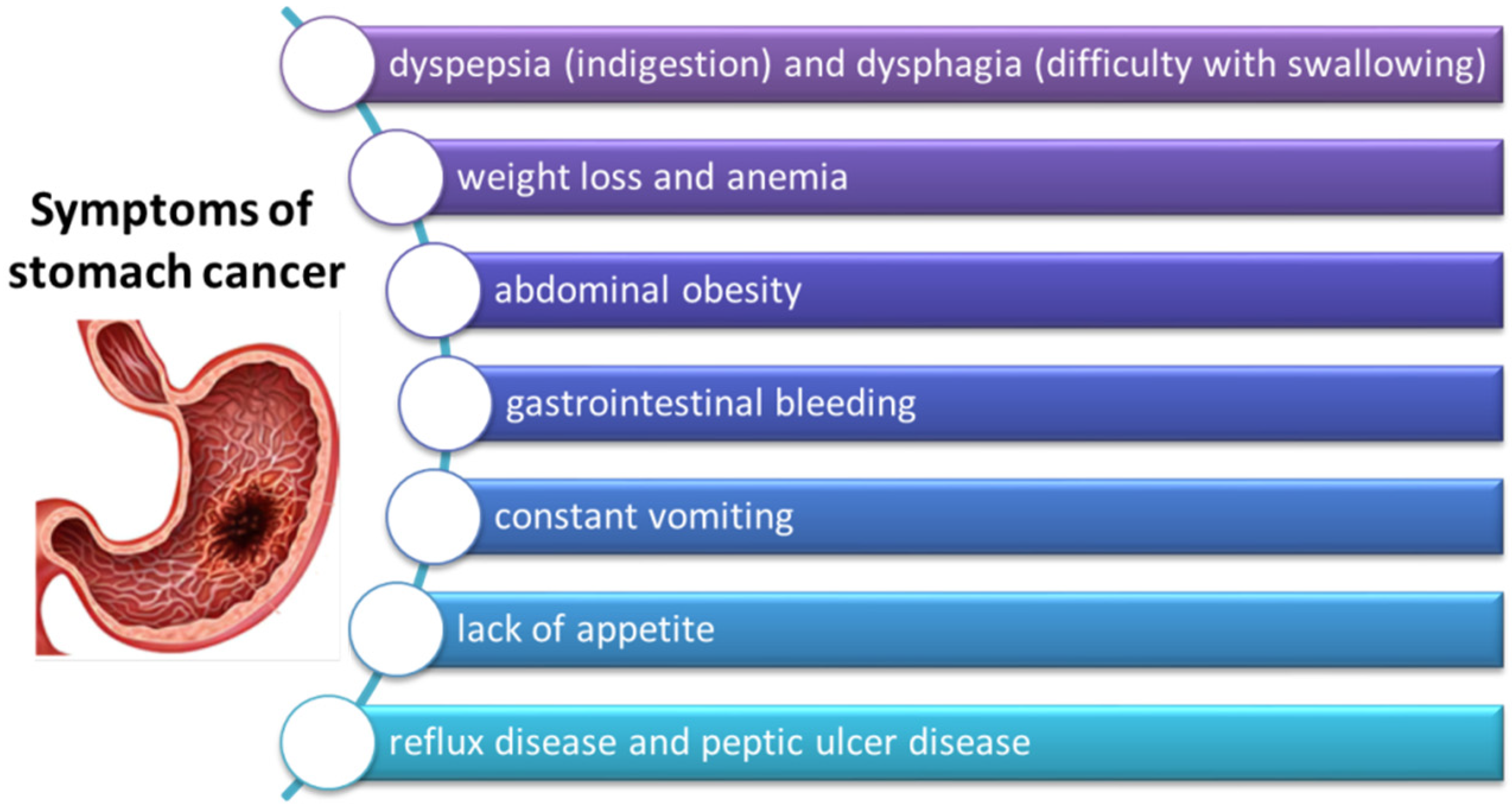
3.3. Symptoms of GC
4. The Role of the Microbiome in the Development and Progression of GC
4.1. Stomach as a Living Environment for Microorganisms
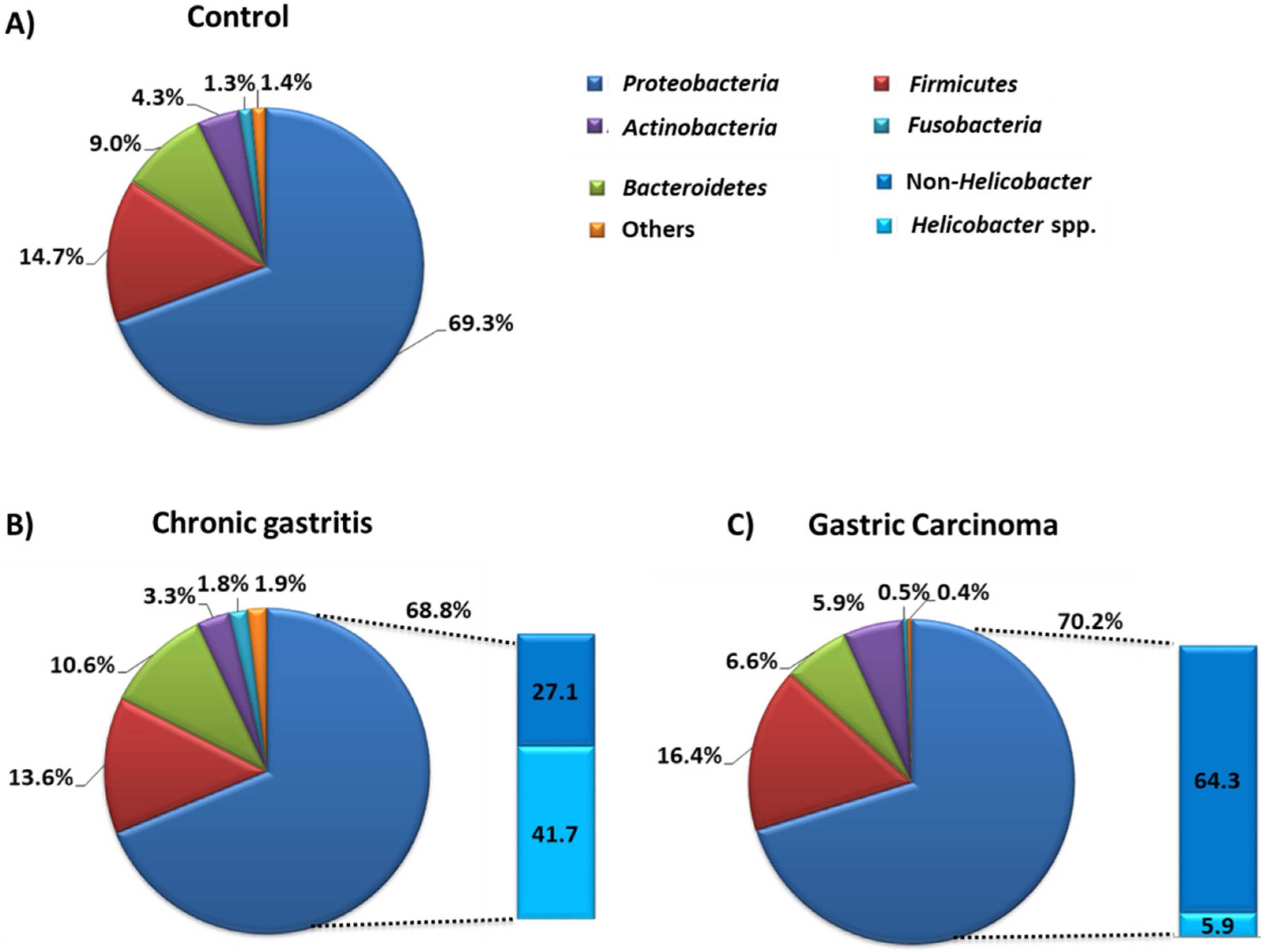
4.2. Symbiosis and Dysbiosis of the Stomach Microbiota
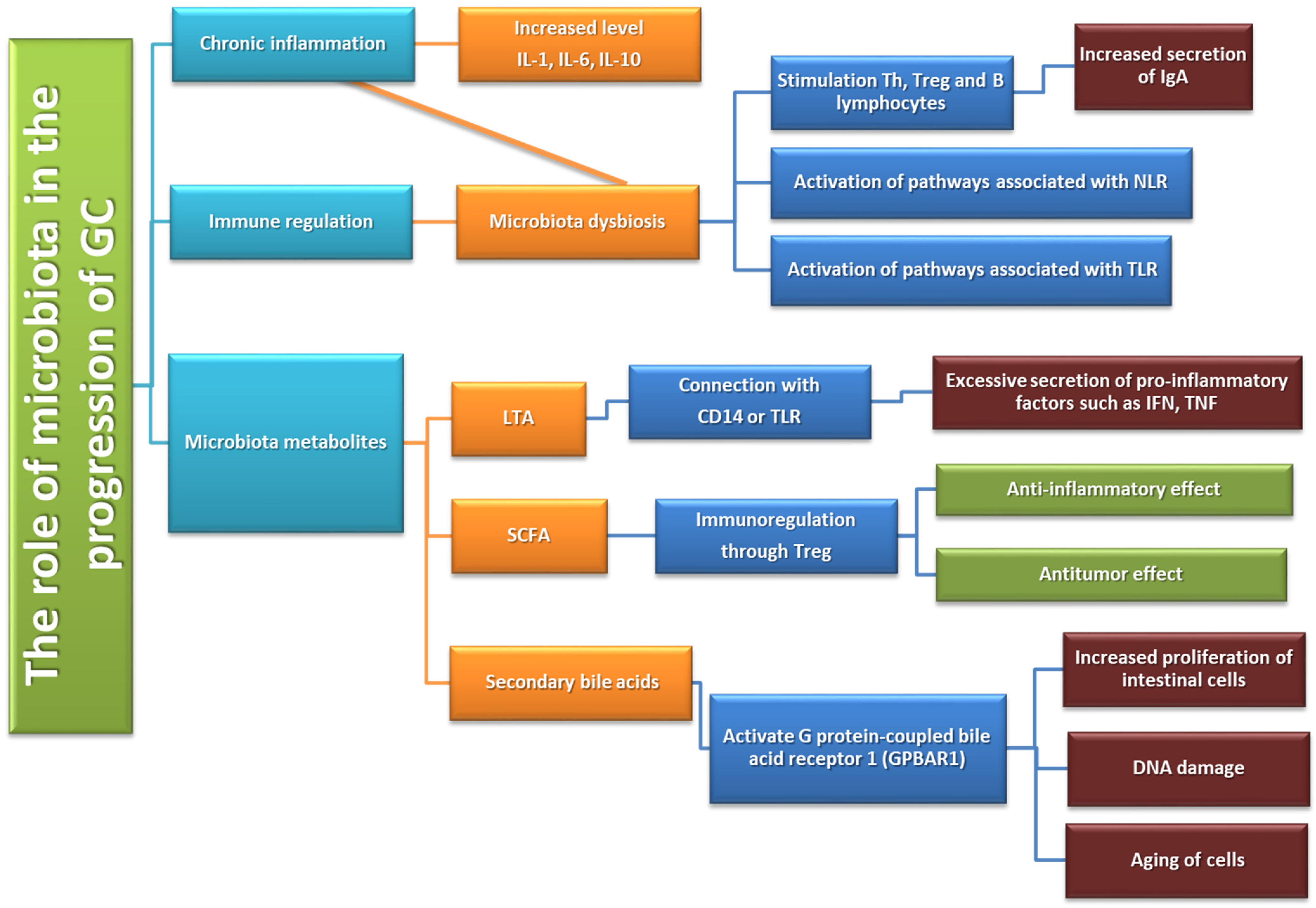
The Role of Gastric Microbiota Metabolites in the Pathogenesis of GC
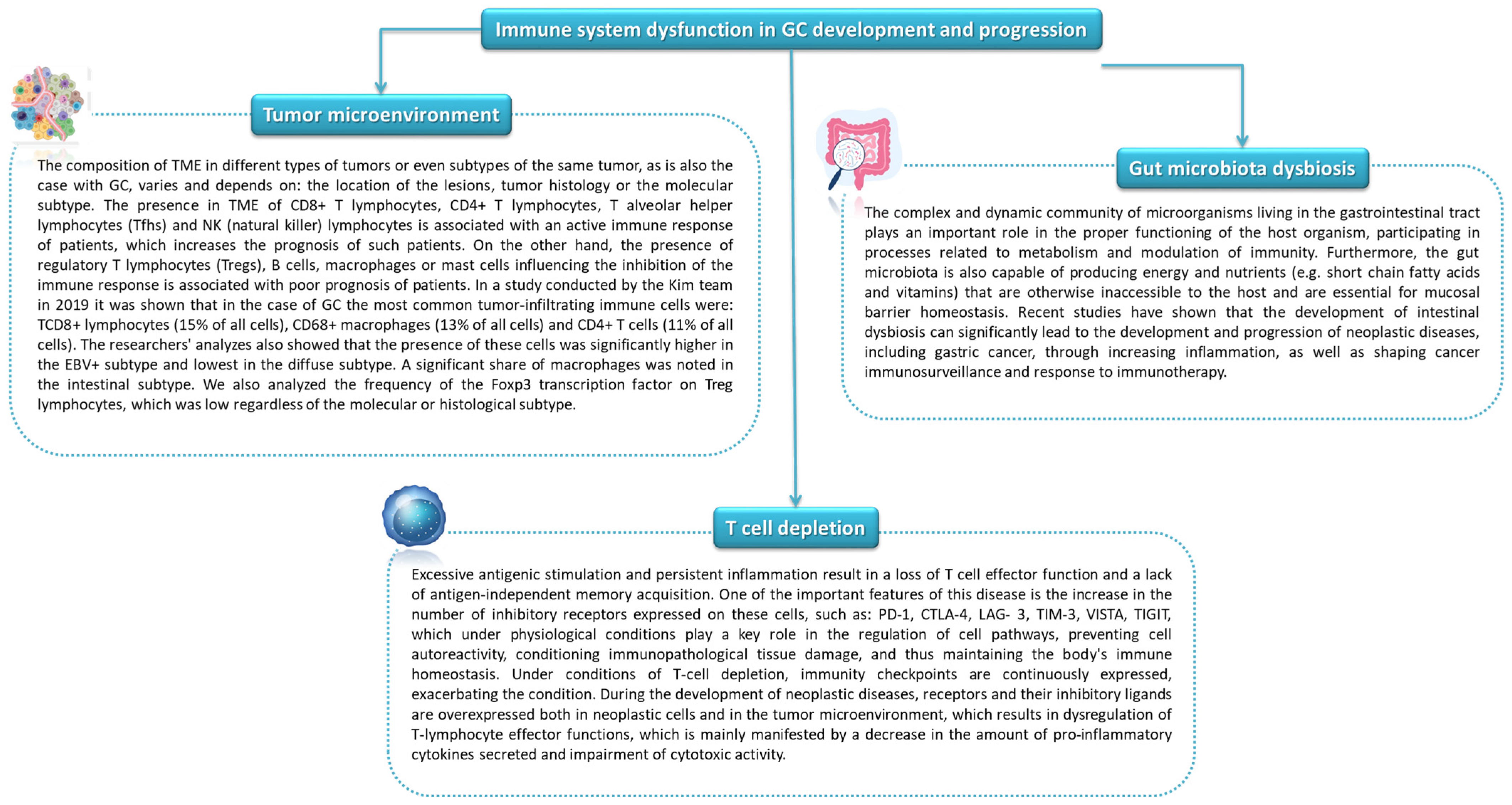
5. The Role of Selected Microorganisms and Immunity in the Development and Progression of Gastric Cancer
5.1. The Role of Selected Microorganisms and Inflamantion
5.2. Influence of the Gut Microbiota on Anti-Tumor Immunity
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Stomach (Gastric) Cancer Key Statistics. Available online: https://www.cancer.org/cancer/stomach-cancer/about/key-statistics.html (accessed on 12 May 2022).
- Stomach Cancer-Statistics. Available online: https://www.cancer.net/cancer-types/stomach-cancer/statistics (accessed on 12 May 2022).
- Rawla, P.; Barsouk, A. Epidemiology of Gastric Cancer: Global Trends, Risk Factors and Prevention. Prz. Gastroenterol. 2019, 14, 26–38. [Google Scholar] [CrossRef]
- Cancer Today. Available online: http://gco.iarc.fr/today/home (accessed on 12 May 2022).
- Cancer of the Stomach-Cancer Stat Facts. Available online: https://seer.cancer.gov/statfacts/html/stomach.html (accessed on 12 May 2022).
- Shah, S.C. Gastric Cancer: A Neglected Threat to Racial and Ethnic Minorities in the USA. Lancet Gastroenterol. Hepatol. 2021, 6, 266–267. [Google Scholar] [CrossRef]
- Pereira, C.; Park, J.-H.; Campelos, S.; Gullo, I.; Lemos, C.; Solorzano, L.; Martins, D.; Gonçalves, G.; Leitão, D.; Lee, H.-J.; et al. Comparison of East-Asia and West-Europe Cohorts Explains Disparities in Survival Outcomes and Highlights Predictive Biomarkers of Early Gastric Cancer Aggressiveness. Int. J. Cancer 2022, 150, 868–880. [Google Scholar] [CrossRef]
- Katoh, H.; Ishikawa, S. Lifestyles, Genetics, and Future Perspectives on Gastric Cancer in East Asian Populations. J. Hum. Genet. 2021, 66, 887–899. [Google Scholar] [CrossRef]
- Stomach Cancer Incidence Statistics. Available online: https://www.cancerresearchuk.org/health-professional/cancer-statistics/statistics-by-cancer-type/stomach-cancer/incidence (accessed on 12 May 2022).
- Stomach (Gastric) Cancer Survival Rates. Available online: https://www.cancer.org/cancer/stomach-cancer/detection-diagnosis-staging/survival-rates.html (accessed on 12 May 2022).
- Khatoon, J.; Rai, R.P.; Prasad, K.N. Role of Helicobacter pylori in gastric cancer: Updates. World J. Gastrointest. Oncol. 2016, 8, 147–158. [Google Scholar] [CrossRef]
- Carcas, L.P. Gastric cancer review. J. Carcinog. 2014, 13, 14. [Google Scholar] [CrossRef]
- Lee, Y.Y.; Derakhshan, M.H. Environmental and Lifestyle Risk Factors of Gastric Cancer. Arch. Iran. Med. 2013, 16, 358–365. [Google Scholar]
- Yang, L.; Ying, X.; Liu, S.; Lyu, G.; Xu, Z.; Zhang, X.; Li, H.; Li, Q.; Wang, N.; Ji, J. Gastric Cancer: Epidemiology, Risk Factors and Prevention Strategies. Chin. J. Cancer Res. 2020, 32, 695–704. [Google Scholar] [CrossRef]
- Sitarz, R.; Skierucha, M.; Mielko, J.; Offerhaus, G.J.A.; Maciejewski, R.; Polkowski, W.P. Gastric Cancer: Epidemiology, Prevention, Classification, and Treatment. Cancer Manag. Res. 2018, 10, 239–248. [Google Scholar] [CrossRef]
- Wang, M.; Busuttil, R.A.; Pattison, S.; Neeson, P.J.; Boussioutas, A. Immunological Battlefield in Gastric Cancer and Role of Immunotherapies. World J. Gastroenterol. 2016, 22, 6373–6384. [Google Scholar] [CrossRef]
- Kumar, S.; Patel, G.K.; Ghoshal, U.C. Helicobacter Pylori-Induced Inflammation: Possible Factors Modulating the Risk of Gastric Cancer. Pathogens 2021, 10, 1099. [Google Scholar] [CrossRef]
- Zhang, R.-G.; Duan, G.-C.; Fan, Q.-T.; Chen, S.-Y. Role of Helicobacter Pylori Infection in Pathogenesis of Gastric Carcinoma. World J. Gastrointest. Pathophysiol. 2016, 7, 97–107. [Google Scholar] [CrossRef]
- Kwak, Y.; Seo, A.N.; Lee, H.E.; Lee, H.S. Tumor Immune Response and Immunotherapy in Gastric Cancer. J. Pathol. Transl. Med. 2020, 54, 20–33. [Google Scholar] [CrossRef]
- Chen, Y.; Sun, Z.; Chen, W.; Liu, C.; Chai, R.; Ding, J.; Liu, W.; Feng, X.; Zhou, J.; Shen, X.; et al. The Immune Subtypes and Landscape of Gastric Cancer and to Predict Based on the Whole-Slide Images Using Deep Learning. Front. Immunol. 2021, 12, 685992. [Google Scholar] [CrossRef]
- Becerril-Rico, J.; Alvarado-Ortiz, E.; Toledo-Guzmán, M.E.; Pelayo, R.; Ortiz-Sánchez, E. The Cross Talk between Gastric Cancer Stem Cells and the Immune Microenvironment: A Tumor-Promoting Factor. Stem. Cell Res. Ther. 2021, 12, 498. [Google Scholar] [CrossRef]
- Hu, B.; El Hajj, N.; Sittler, S.; Lammert, N.; Barnes, R.; Meloni-Ehrig, A. Gastric Cancer: Classification, Histology and Application of Molecular Pathology. J. Gastrointest. Oncol. 2012, 3, 251–261. [Google Scholar] [CrossRef]
- Gullo, I.; Grillo, F.; Mastracci, L.; Vanoli, A.; Carneiro, F.; Saragoni, L.; Limarzi, F.; Ferro, J.; Parente, P.; Fassan, M. Precancerous Lesions of the Stomach, Gastric Cancer and Hereditary Gastric Cancer Syndromes. Pathologica 2020, 112, 166–185. [Google Scholar] [CrossRef]
- Poorolajal, J.; Moradi, L.; Mohammadi, Y.; Cheraghi, Z.; Gohari-Ensaf, F. Risk Factors for Stomach Cancer: A Systematic Review and Meta-Analysis. Epidemiol. Health 2020, 42, e2020004. [Google Scholar] [CrossRef]
- Zhao, Y.; Deng, X.; Song, G.; Qin, S.; Liu, Z. The GSTM1 Null Genotype Increased Risk of Gastric Cancer: A Meta-Analysis Based on 46 Studies. PLoS ONE 2013, 8, e81403. [Google Scholar] [CrossRef]
- Zhang, X.-L.; Cui, Y.-H. GSTM1 Null Genotype and Gastric Cancer Risk in the Chinese Population: An Updated Meta-Analysis and Review. OncoTargets Ther. 2015, 8, 969–975. [Google Scholar] [CrossRef]
- Hereditary Diffuse Gastric Cancer (HDGC). Available online: https://www.hopkinsmedicine.org/health/conditions-and-diseases/hereditary-diffuse-gastric-cancer-hdgc (accessed on 12 May 2022).
- Luo, W.; Fedda, F.; Lynch, P.; Tan, D. CDH1 Gene and Hereditary Diffuse Gastric Cancer Syndrome: Molecular and Histological Alterations and Implications for Diagnosis and Treatment. Front. Pharmacol. 2018, 9, 1421. [Google Scholar] [CrossRef]
- Long, Z.-W.; Yu, H.-M.; Wang, Y.-N.; Liu, D.; Chen, Y.-Z.; Zhao, Y.-X.; Bai, L. Association of IL-17 Polymorphisms with Gastric Cancer Risk in Asian Populations. World J. Gastroenterol. WJG 2015, 21, 5707–5718. [Google Scholar] [CrossRef]
- Chen, L.; Shi, Y.; Zhu, X.; Guo, W.; Zhang, M.; Che, Y.; Tang, L.; Yang, X.; You, Q.; Liu, Z. IL10 Secreted by Cancer associated Macrophages Regulates Proliferation and Invasion in Gastric Cancer Cells via CMet/STAT3 Signaling. Oncol. Rep. 2019, 42, 595–604. [Google Scholar] [CrossRef]
- Tang, J.; Pan, R.; Xu, L.; Ma, Q.; Ying, X.; Zhao, J.; Zhao, H.; Miao, L.; Xu, Y.; Duan, S.; et al. IL10 Hypomethylation Is Associated with the Risk of Gastric Cancer. Oncol. Lett. 2021, 21, 241. [Google Scholar] [CrossRef]
- Alizadeh, A.M.; Afrouzan, H.; Dinparast-Djadid, N.; Sawaya, A.C.H.F.; Azizian, S.; Hemmati, H.R.; Mohagheghi, M.A.; Erfani, S. Chemoprotection of MNNG-Initiated Gastric Cancer in Rats Using Iranian Propolis. Arch. Iran. Med. 2015, 18, 18–23. [Google Scholar]
- Kim, T.-J.; Kim, M.-K.; Jung, D. MNNG-Regulated Differentially Expressed Genes that Contribute to Cancer Development in Stomach Cells. Korean J. Clin. Lab. Sci. 2021, 53, 353–362. [Google Scholar] [CrossRef]
- Kamineni, A.; Williams, M.A.; Schwartz, S.M.; Cook, L.S.; Weiss, N.S. The Incidence of Gastric Carcinoma in Asian Migrants to the United States and Their Descendants. Cancer Causes Control 1999, 10, 77–83. [Google Scholar] [CrossRef]
- Taylor, V.M.; Ko, L.K.; Hwang, J.H.; Sin, M.-K.; Inadomi, J.M. Gastric Cancer in Asian American Populations: A Neglected Health Disparity. Asian Pac. J. Cancer Prev. 2015, 15, 10565–10571. [Google Scholar] [CrossRef]
- Red Meat, Processed Meat and Cancer. Available online: https://www.cancercouncil.com.au/1in3cancers/lifestyle-choices-and-cancer/red-meat-processed-meat-and-cancer/ (accessed on 12 May 2022).
- Zhu, H.; Yang, X.; Zhang, C.; Zhu, C.; Tao, G.; Zhao, L.; Tang, S.; Shu, Z.; Cai, J.; Dai, S.; et al. Red and Processed Meat Intake Is Associated with Higher Gastric Cancer Risk: A Meta-Analysis of Epidemiological Observational Studies. PLoS ONE 2013, 8, e70955. [Google Scholar] [CrossRef]
- Borucka, A.; Ostaszewski, K. Koncepcja Resilience. Kluczowe Pojęcia i Wybrane Zagadnienia. Med. Wieku. Rozw. 2008, 12, 587–597. [Google Scholar]
- Kim, J.H.; Lee, J.; Choi, I.J.; Kim, Y.-I.; Kwon, O.; Kim, H.; Kim, J. Dietary Carotenoids Intake and the Risk of Gastric Cancer: A Case—Control Study in Korea. Nutrients 2018, 10, 1031. [Google Scholar] [CrossRef]
- Chen, Q.-H.; Wu, B.-K.; Pan, D.; Sang, L.-X.; Chang, B. Beta-Carotene and Its Protective Effect on Gastric Cancer. World J. Clin. Cases 2021, 9, 6591–6607. [Google Scholar] [CrossRef]
- Liu, C.; Russell, R.M. Nutrition and Gastric Cancer Risk: An Update. Nutr. Rev. 2008, 66, 237–249. [Google Scholar] [CrossRef]
- Tsugane, S.; Sasazuki, S. Diet and the Risk of Gastric Cancer: Review of Epidemiological Evidence. Gastric. Cancer 2007, 10, 75–83. [Google Scholar] [CrossRef]
- Agudo, A.; Cayssials, V.; Bonet, C.; Tjønneland, A.; Overvad, K.; Boutron-Ruault, M.-C.; Affret, A.; Fagherazzi, G.; Katzke, V.; Schübel, R.; et al. Inflammatory Potential of the Diet and Risk of Gastric Cancer in the European Prospective Investigation into Cancer and Nutrition (EPIC) Study. Am. J. Clin. Nutr. 2018, 107, 607–616. [Google Scholar] [CrossRef]
- Wroblewski, L.E.; Peek, R.M.; Wilson, K.T. Helicobacter Pylori and Gastric Cancer: Factors That Modulate Disease Risk. Clin. Microbiol. Rev. 2010, 23, 713–739. [Google Scholar] [CrossRef]
- Helicobacter Pylori and Cancer—NCI. Available online: https://www.cancer.gov/about-cancer/causes-prevention/risk/infectious-agents/h-pylori-fact-sheet (accessed on 12 May 2022).
- Ishaq, S.; Nunn, L. Helicobacter Pylori and Gastric Cancer: A State of the Art Review. Gastroenterol. Hepatol. Bed Bench 2015, 8, S6–S14. [Google Scholar]
- Díaz, P.; Valenzuela Valderrama, M.; Bravo, J.; Quest, A.F.G. Helicobacter pylori and Gastric Cancer: Adaptive Cellular Mechanisms Involved in Disease Progression. Front. Microbiol. 2018, 9, 5. [Google Scholar] [CrossRef]
- Choi, I.J.; Kim, C.G.; Lee, J.Y.; Kim, Y.-I.; Kook, M.-C.; Park, B.; Joo, J. Family History of Gastric Cancer and Helicobacter Pylori Treatment. New Engl. J. Med. 2020, 382, 427–436. [Google Scholar] [CrossRef]
- Yang, J.; Zhou, X.; Liu, X.; Ling, Z.; Ji, F. Role of the Gastric Microbiome in Gastric Cancer: From Carcinogenesis to Treatment. Front. Microbiol. 2021, 12, 641322. [Google Scholar] [CrossRef]
- Yang, P.; Zhou, Y.; Chen, B.; Wan, H.-W.; Jia, G.-Q.; Bai, H.-L.; Wu, X.-T. Overweight, Obesity and Gastric Cancer Risk: Results from a Meta-Analysis of Cohort Studies. Eur. J. Cancer 2009, 45, 2867–2873. [Google Scholar] [CrossRef]
- Hirabayashi, M.; Inoue, M.; Sawada, N.; Saito, E.; Abe, S.K.; Hidaka, A.; Iwasaki, M.; Yamaji, T.; Shimazu, T.; Shibuya, K.; et al. Effect of Body-Mass Index on the Risk of Gastric Cancer: A Population-Based Cohort Study in A Japanese Population. Cancer Epidemiol. 2019, 63, 101622. [Google Scholar] [CrossRef] [PubMed]
- Perre, A. Battling Obesity and Cancer? An Integrated Team? S Approach—Onco’Zine 2018. Available online: https://www.oncozine.com/battling-obesity-and-cancer-an-integrated-teams-approach/ (accessed on 19 June 2022).
- Cui, X.; Zhang, H.; Cao, A.; Cao, L.; Hu, X. Cytokine TNF-α Promotes Invasion and Metastasis of Gastric Cancer by down-Regulating Pentraxin3. J. Cancer 2020, 11, 1800–1807. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, H.; Hirata, Y.; Nakagawa, H.; Sakamoto, K.; Hayakawa, Y.; Takahashi, R.; Nakata, W.; Sakitani, K.; Serizawa, T.; Hikiba, Y.; et al. Interleukin-6 Mediates Epithelial–Stromal Interactions and Promotes Gastric Tumorigenesis. PLoS ONE 2013, 8, e60914. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.-F.; Chen, P.-C.; Chang, T.-M.; Hou, C.-H. Monocyte Chemoattractant Protein-1 Promotes Cancer Cell Migration via c-Raf/MAPK/AP-1 Pathway and MMP-9 Production in Osteosarcoma. J. Exp. Clin. Cancer Res. 2020, 39, 254. [Google Scholar] [CrossRef]
- Menéndez, P.; Padilla, D.; Villarejo, P.; Menéndez, J.M.; Lora, D. Does Bariatric Surgery Decrease Gastric Cancer Risk? HepatoGastroenterol. Hepatol. Bed Bench. 2012, 59, 409–412. [Google Scholar] [CrossRef]
- Aurello, P.; Petrucciani, N.; Antolino, L.; Giulitti, D.; D’Angelo, F.; Ramacciato, G. Follow-up after Curative Resection for Gastric Cancer: Is it Time to Tailor it? World J. Gastroenterol. 2017, 23, 3379–3387. [Google Scholar] [CrossRef]
- Lahner, E.; Annibale, B. Pernicious Anemia: New Insights from a Gastroenterol Point of View. World J. Gastroenterol. 2009, 15, 5121–5128. [Google Scholar] [CrossRef]
- Cisło, M.; Filip, A.A.; Arnold Offerhaus, G.J.; Ciseł, B.; Rawicz-Pruszyński, K.; Skierucha, M.; Polkowski, W.P. Distinct Molecular Subtypes of Gastric Cancer: From Laurén to Molecular Pathology. Oncotarget 2018, 9, 19427–19442. [Google Scholar] [CrossRef]
- Park, H.S.; Lee, S.-Y.; Hong, S.N.; Kim, J.H.; Sung, I.-K.; Park, H.S.; Shim, C.S.; Jin, C.J. Early Gastric Cancer-Like Advanced Gastric Cancer Versus Advanced Gastric Cancer-Like Early Gastric Cancer. Clin. Endosc. 2013, 46, 155–160. [Google Scholar] [CrossRef]
- De Sol, A.; Trastulli, S.; Grassi, V.; Corsi, A.; Barillaro, I.; Boccolini, A.; Di Patrizi, M.S.; DI Rocco, G.; Santoro, A.; Cirocchi, R.; et al. Requirement for a Standardised Definition of Advanced Gastric Cancer. Oncol. Lett. 2014, 7, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Machlowska, J.; Baj, J.; Sitarz, M.; Maciejewski, R.; Sitarz, R. Gastric Cancer: Epidemiology, Risk Factors, Classification, Genomic Characteristics and Treatment Strategies. Int. J. Mol. Sci. 2020, 21, 4012. [Google Scholar] [CrossRef]
- Carneiro, F. Pathology of Gastric Cancer. 38. Available online: https://oncologypro.esmo.org/content/download/240507/4025970/file/2019-ESMO-Preceptorship-Gastric-Pathology-Fatima-Carneiro.pdf (accessed on 12 May 2022).
- Friis-Hansen, L. Achlorhydria is Associated with Gastric Microbial Overgrowth and Development of Cancer: Lessons Learned from the Gastrin Knockout Mouse. Scand. J. Clin. Lab. Investig. 2006, 66, 607–622. [Google Scholar] [CrossRef] [PubMed]
- Berlth, F.; Bollschweiler, E.; Drebber, U.; Hoelscher, A.H.; Moenig, S. Pathohistological Classification Systems in Gastric Cancer: Diagnostic Relevance and Prognostic Value. World J. Gastroenterol. 2014, 20, 5679–5684. [Google Scholar] [CrossRef] [PubMed]
- Al-Shamsi, H.O.; Alzaabi, A.A.; Afrit, M.; Abu-Gheida, I.; Musallam, K.M. Clinicopathological Features of Gastric Cancer in a Cohort of Gulf Council Countries’ Patients: A Cross-Sectional Study of 96 Cases. J. Oncol Res. Rev. Rep. 2021, 2, 1–6. [Google Scholar] [CrossRef]
- The Cancer Genome Atlas—Gastric Adenocarcinoma Study—NCI. Available online: https://www.cancer.gov/about-nci/organization/ccg/research/structural-genomics/tcga/studied-cancers/stomach (accessed on 12 May 2022).
- Sohn, B.H.; Hwang, J.-E.; Jang, H.-J.; Lee, H.-S.; Oh, S.C.; Shim, J.-J.; Lee, K.-W.; Kim, E.H.; Yim, S.Y.; Lee, S.H.; et al. Clinical Significance of Four Molecular Subtypes of Gastric Cancer Identified by The Cancer Genome Atlas Project. Clin. Cancer Res. 2017, 23, 4441–4449. [Google Scholar] [CrossRef]
- Serra, O.; Galán, M.; Ginesta, M.M.; Calvo, M.; Sala, N.; Salazar, R. Comparison and Applicability of Molecular Classifications for Gastric Cancer. Cancer Treat. Rev. 2019, 77, 29–34. [Google Scholar] [CrossRef]
- Batalha, S.; Ferreira, S.; Brito, C. The Peripheral Immune Landscape of Breast Cancer: Clinical Findings and In Vitro Models for Biomarker Discovery. Cancers 2021, 13, 1305. [Google Scholar] [CrossRef]
- Shibata, D.; Weiss, L.M. Epstein-Barr Virus-Associated Gastric Adenocarcinoma. Am. J. Pathol. 1992, 140, 769–774. [Google Scholar]
- Alarcón, A.; Figueroa, U.; Espinoza, B.; Sandoval, A.; Carrasco-Aviño, G.; Aguayo, F.R.; Corvalan, A.H. Epstein-Barr Virus–Associated Gastric Carcinoma: The Americas’ Perspective; IntechOpen: London, UK, 2017; ISBN 978-953-51-3528-9. [Google Scholar]
- Tang, C.-T.; Zeng, L.; Yang, J.; Zeng, C.; Chen, Y. Analysis of the Incidence and Survival of Gastric Cancer Based on the Lauren Classification: A Large Population-Based Study Using SEER. Front Oncol. 2020, 10, 1212. [Google Scholar] [CrossRef]
- Macke, L.; Schulz, C.; Malfertheiner, P. The Fear of Gastric Cancer in Patients with Dyspepsia: Challenge in Specialist Care Gastroenterology. Dig. Dis. 2021, 40, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Choi, M. Gastric Cancer Screening and Alarm Symptoms in Early Gastric Cancer; ClinicalTrials.gov; 2012. Available online: https://clinicaltrials.gov/ct2/show/NCT01659632 (accessed on 19 June 2022).
- Maconi, G.; Kurihara, H.; Panizzo, V.; Russo, A.; Cristaldi, M.; Marrelli, D.; Roviello, F.; de Manzoni, G.; Di Leo, A.; Morgagni, P.; et al. Gastric Cancer in Young Patients with no Alarm Symptoms: Focus on Delay in Diagnosis, Stage of Neoplasm and Survival. Scand. J. Gastroenterol. 2003, 38, 1249–1255. [Google Scholar] [CrossRef] [PubMed]
- Stephens, M.R.; Lewis, W.G.; White, S.; Blackshaw, G.R.J.C.; Edwards, P.; Barry, J.D.; Allison, M.C. Prognostic Significance of Alarm Symptoms in Patients with Gastric Cancer. J. Br. Surg. 2005, 92, 840–846. [Google Scholar] [CrossRef] [PubMed]
- Bowrey, D.J.; Griffin, S.M.; Wayman, J.; Karat, D.; Hayes, N.; Raimes, S.A. Use of Alarm Symptoms to Select Dyspeptics for Endoscopy Causes Patients with Curable Esophagogastric Cancer to Be Overlooked. Surg. Endosc. 2006, 20, 1725–1728. [Google Scholar] [CrossRef]
- Rasmussen, S.; Haastrup, P.F.; Balasubramaniam, K.; Christensen, R.D.; Søndergaard, J.; Jarbøl, D.E. Predictive Values of Upper Gastrointestinal Cancer Alarm Symptoms in the General Population: A Nationwide Cohort Study. BMC Cancer 2018, 18, 440. [Google Scholar] [CrossRef]
- Dos Santos Guedes, M.T.; de Jesus, J.P.; de Souza Filho, O.; Fontenele, R.M.; Sousa, A.I. Clinical and Epidemiological Profile of Cases of Deaths from Stomach Cancer in the National Cancer Institute, Brazil. Ecancermedicalscience 2014, 8, 445. [Google Scholar] [CrossRef]
- Mao, L.; Franke, J. Symbiosis, Dysbiosis, and Rebiosis—The Value of Metaproteomics in Human Microbiome Monitoring. Proteomics 2015, 15, 1142–1151. [Google Scholar] [CrossRef]
- Sheflin, A.M.; Whitney, A.K.; Weir, T.L. Cancer-Promoting Effects of Microbial Dysbiosis. Curr. Oncol. Rep. 2014, 16, 406. [Google Scholar] [CrossRef]
- Nasr, R.; Shamseddine, A.; Mukherji, D.; Nassar, F.; Temraz, S. The Crosstalk between Microbiome and Immune Response in Gastric Cancer. Int. J. Mol. Sci. 2020, 21, 6586. [Google Scholar] [CrossRef]
- Hao, W.-L.; Lee, Y.-K. Microflora of the Gastrointestinal Tract. In Public Health Microbiol.ogy: Methods and Protocols; Methods in Molecular Biology; Spencer, J.F.T., de Spencer, A.L.R., Eds.; Humana Press: Totowa, NJ, USA, 2004; pp. 491–502. ISBN 978-1-59259-766-6. [Google Scholar]
- O’May, G.A.; Reynolds, N.; Macfarlane, G.T. Effect of PH on an In Vitro Model of Gastric Microbiota in Enteral Nutrition Patients. Appl. Environ. Microbiol. 2005, 71, 4777–4783. [Google Scholar] [CrossRef]
- Martinez, J.E.; Kahana, D.D.; Ghuman, S.; Wilson, H.P.; Wilson, J.; Kim, S.C.J.; Lagishetty, V.; Jacobs, J.P.; Sinha-Hikim, A.P.; Friedman, T.C. Unhealthy Lifestyle and Gut Dysbiosis: A Better Understanding of the Effects of Poor Diet and Nicotine on the Intestinal Microbiome. Front. Endocrinol. 2021, 12, 667066. [Google Scholar] [CrossRef] [PubMed]
- Caguazango, J.C. Ecological Models of Gastric Microbiota Dysbiosis: Helicobacter Pylori and Gastric Carcinogenesis. Med. Microecol. 2020, 3, 100010. [Google Scholar] [CrossRef]
- Ferreira, R.M.; Pereira-Marques, J.; Pinto-Ribeiro, I.; Costa, J.L.; Carneiro, F.; Machado, J.C.; Figueiredo, C. Gastric Microbial Community Profiling Reveals a Dysbiotic Cancer-Associated Microbiota. Gut 2018, 67, 226–236. [Google Scholar] [CrossRef]
- Wang, L.; Zhou, J.; Xin, Y.; Geng, C.; Tian, Z.; Yu, X.; Dong, Q. Bacterial Overgrowth and Diversification of Microbiota in Gastric Cancer. Eur. J. Gastroenterol. Hepatol. 2016, 28, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Griffin, M.; Shakespeare, S.; Shepherd, H.J.; Harding, C.J.; Létard, J.-F.; Desplanches, C.; Goeta, A.E.; Howard, J.A.K.; Powell, A.K.; Mereacre, V.; et al. A Symmetry-Breaking Spin-State Transition in Iron(III). Angew. Chem. Int. Ed. 2011, 50, 896–900. [Google Scholar] [CrossRef] [PubMed]
- Hatakeyama, M. Helicobacter Pylori CagA and Gastric Cancer: A Paradigm for Hit-and-Run Carcinogenesis. Cell Host Microbe 2014, 15, 306–316. [Google Scholar] [CrossRef]
- Vinasco, K.; Mitchell, H.M.; Kaakoush, N.O.; Castaño-Rodríguez, N. Microbial Carcinogenesis: Lactic Acid Bacteria in Gastric Cancer. Biochim. Biophys. Acta Rev. Cancer 2019, 1872, 188309. [Google Scholar] [CrossRef]
- Li, Z.-P.; Liu, J.-X.; Lu, L.-L.; Wang, L.-L.; Xu, L.; Guo, Z.-H.; Dong, Q.-J. Overgrowth of Lactobacillus in Gastric Cancer. World J. Gastrointest. Oncol. 2021, 13, 1099–1108. [Google Scholar] [CrossRef]
- Whiteside, S.A.; Mohiuddin, M.M.; Shlimon, S.; Chahal, J.; MacPherson, C.W.; Jass, J.; Tompkins, T.A.; Creuzenet, C. In Vitro Framework to Assess the Anti-Helicobacter Pylori Potential of Lactic Acid Bacteria Secretions as Alternatives to Antibiotics. Int. J. Mol. Sci. 2021, 22, 5650. [Google Scholar] [CrossRef]
- Ping, W.; Senyan, H.; Li, G.; Yan, C.; Long, L. Increased Lactate in Gastric Cancer Tumor-Infiltrating Lymphocytes is Related to Impaired T Cell Function Due to MiR-34a Deregulated Lactate Dehydrogenase, A. Cell. Physiol. Biochem. 2018, 49, 828–836. [Google Scholar] [CrossRef]
- Aday, U.; Tatlı, F.; Akpulat, F.V.; İnan, M.; Kafadar, M.T.; Bilge, H.; Başol, Ö.; Oğuz, A. Prognostic Significance of Pretreatment Serum Lactate Dehydrogenase-to-Albumin Ratio in Gastric Cancer. Contemp. Oncol. 2020, 24, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Chattopadhyay, I.; Gundamaraju, R.; Jha, N.K.; Gupta, P.K.; Dey, A.; Mandal, C.C.; Ford, B.M. Interplay between Dysbiosis of Gut Microbiome, Lipid Metabolism, and Tumorigenesis: Can Gut Dysbiosis Stand as a Prognostic Marker in Cancer? Dis. Markers 2022, 2022, 2941248. [Google Scholar] [CrossRef] [PubMed]
- Pappas-Gogos, G.; Tepelenis, K.; Fousekis, F.; Katsanos, K.; Pitiakoudis, M.; Vlachos, K. The Implication of Gastric Microbiome in the Treatment of Gastric Cancer. Cancers 2022, 14, 2039. [Google Scholar] [CrossRef] [PubMed]
- Jaye, K.; Li, C.G.; Chang, D.; Bhuyan, D.J. The Role of Key Gut Microbial Metabolites in the Development and Treatment of Cancer. Gut Microbes 2022, 14, 2038865. [Google Scholar] [CrossRef] [PubMed]
- Mertowska, P.; Mertowski, S.; Wojnicka, J.; Korona-Głowniak, I.; Grywalska, E.; Błażewicz, A.; Załuska, W. A Link between Chronic Kidney Disease and Gut Microbiota in Immunological and Nutritional Aspects. Nutrients 2021, 13, 3637. [Google Scholar] [CrossRef]
- Mirzaei, R.; Afaghi, A.; Babakhani, S.; Sohrabi, M.R.; Hosseini-Fard, S.R.; Babolhavaeji, K.; Khani Ali Akbari, S.; Yousefimashouf, R.; Karampoor, S. Role of Microbiota-Derived Short-Chain Fatty Acids in Cancer Development and Prevention. Biomed. Pharmacother. 2021, 139, 111619. [Google Scholar] [CrossRef]
- Coussens, L.M.; Werb, Z. Inflammation and Cancer. Nature 2002, 420, 860–867. [Google Scholar] [CrossRef]
- Singh, N.; Baby, D.; Rajguru, J.P.; Patil, P.B.; Thakkannavar, S.S.; Pujari, V.B. Inflammation and Cancer. Ann. Afr. Med. 2019, 18, 121–126. [Google Scholar] [CrossRef]
- Zhao, H.; Wu, L.; Yan, G.; Chen, Y.; Zhou, M.; Wu, Y.; Li, Y. Inflammation and Tumor Progression: Signaling Pathways and Targeted Intervention. Sig. Transduct. Target Ther. 2021, 6, 1–46. [Google Scholar] [CrossRef]
- Oya, Y.; Hayakawa, Y.; Koike, K. Tumor Microenvironment in Gastric Cancers. Cancer Sci. 2020, 111, 2696–2707. [Google Scholar] [CrossRef]
- Kim, T.S.; da Silva, E.; Coit, D.G.; Tang, L.H. Intratumoral Immune Response to Gastric Cancer Varies by Molecular and Histologic Subtype. Am. J. Surg. Pathol. 2019, 43, 851–860. [Google Scholar] [CrossRef]
- Colombo, M.P.; Piconese, S. Regulatory T-Cell Inhibition versus Depletion: The Right Choice in Cancer Immunotherapy. Nat. Rev. Cancer 2007, 7, 880–887. [Google Scholar] [CrossRef]
- Zhang, Y.; Lazarus, J.; Steele, N.G.; Yan, W.; Lee, H.-J.; Nwosu, Z.C.; Halbrook, C.J.; Menjivar, R.E.; Kemp, S.B.; Sirihorachai, V.R.; et al. Regulatory T-Cell Depletion Alters the Tumor Microenvironment and Accelerates Pancreatic Carcinogenesis. Cancer Discov. 2020, 10, 422–439. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-Q.; Zhang, Y.; Jiang, W.; Chen, Y.-P.; Xu, S.-Y.; Liu, N.; Zhao, Y.; Li, L.; Lei, Y.; Hong, X.-H.; et al. Development and Validation of an Immune Checkpoint-Based Signature to Predict Prognosis in Nasopharyngeal Carcinoma Using Computational Pathology Analysis. J. Immunother. Cancer 2019, 7, 298. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Pan, Z. Influence of Microbiota on Immunity and Immunotherapy for Gastric and Esophageal Cancers. Gastroenterol. Rep. 2020, 8, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Meng, C.; Bai, C.; Brown, T.D.; Hood, L.E.; Tian, Q. Human Gut Microbiota and Gastrointestinal Cancer. Genom. Proteom. Bioinform. 2018, 16, 33–49. [Google Scholar] [CrossRef] [PubMed]
- Cadamuro, A.C.T.; Rossi, A.F.T.; Maniezzo, N.M.; Silva, A.E. Helicobacter Pylori Infection: Host Immune Response, Implications on Gene Expression and MicroRNAs. World J. Gastroenterol. WJG 2014, 20, 1424–1437. [Google Scholar] [CrossRef]
- Chang, W.-L.; Yeh, Y.-C.; Sheu, B.-S. The Impacts of H. Pylori Virulence Factors on the Development of Gastroduodenal Diseases. J. Biomed. Sci. 2018, 25, 68. [Google Scholar] [CrossRef]
- Wen, S.; Moss, S.F. Helicobacter Pylori Virulence Factors in Gastric Carcinogenesis. Cancer Lett. 2009, 282, 1–8. [Google Scholar] [CrossRef]
- Foegeding, N.J.; Caston, R.R.; McClain, M.S.; Ohi, M.D.; Cover, T.L. An Overview of Helicobacter Pylori VacA Toxin Biology. Toxins 2016, 8, 173. [Google Scholar] [CrossRef]
- Caston, R.R.; Sierra, J.C.; Foegeding, N.J.; Truelock, M.D.; Campbell, A.M.; Frick-Cheng, A.E.; Bimczok, D.; Wilson, K.T.; McClain, M.S.; Cover, T.L. Functional Properties of Helicobacter Pylori VacA Toxin M1 and M2 Variants. Infect. Immunity 2020, 88, e00032-20. [Google Scholar] [CrossRef] [PubMed]
- Djekic, A.; Müller, A. The Immunomodulator VacA Promotes Immune Tolerance and Persistent Helicobacter Pylori Infection through its Activities on T-Cells and Antigen-Presenting Cells. Toxins 2016, 8, 187. [Google Scholar] [CrossRef]
- Stein, M.; Ruggiero, P.; Rappuoli, R.; Bagnoli, F. Helicobacter Pylori CagA: From Pathogenic Mechanisms to Its Use as an Anti-Cancer Vaccine. Front. Immunol. 2013, 4, 328. [Google Scholar] [CrossRef] [PubMed]
- Takahashi-Kanemitsu, A.; Knight, C.T.; Hatakeyama, M. Molecular Anatomy and Pathogenic Actions of Helicobacter Pylori CagA That Underpin Gastric Carcinogenesis. Cell Mol. Immunol. 2020, 17, 50–63. [Google Scholar] [CrossRef]
- Li, W.; Zhou, Y.; Shang, C.; Sang, H.; Zhu, H. Effects of Environmental PH on the Growth of Gastric Cancer Cells. Gastroenterol. Res. Pract. 2020, 2020, e3245359. [Google Scholar] [CrossRef] [PubMed]
- Hardbower, D.M.; Peek, R.M.; Wilson, K.T. At the Bench: Helicobacter Pylori, Dysregulated Host Responses, DNA Damage, and Gastric Cancer. J. Leukoc. Biol. 2014, 96, 201–212. [Google Scholar] [CrossRef]
- Sayed, I.M.; Sahan, A.Z.; Venkova, T.; Chakraborty, A.; Mukhopadhyay, D.; Bimczok, D.; Beswick, E.J.; Reyes, V.E.; Pinchuk, I.; Sahoo, D.; et al. Helicobacter Pylori Infection Downregulates the DNA Glycosylase NEIL2, Resulting in Increased Genome Damage and Inflammation in Gastric Epithelial Cells. J. Biol. Chem. 2020, 295, 11082–11098. [Google Scholar] [CrossRef] [PubMed]
- Mahdi, B.M. Immune Response to Helicobacter Pylori; IntechOpen: London, UK, 2014; ISBN 978-953-51-1239-6. [Google Scholar]
- Könönen, E.; Gursoy, U.K. Oral Prevotella Species and Their Connection to Events of Clinical Relevance in Gastrointestinal and Respiratory Tracts. Front. Microbiol. 2022, 12, 798763. [Google Scholar] [CrossRef]
- Zhang, X.; Li, C.; Cao, W.; Zhang, Z. Alterations of Gastric Microbiota in Gastric Cancer and Precancerous Stages. Front. Cell. Infect. Microbiol. 2021, 11, 559148. [Google Scholar] [CrossRef]
- Barra, W.F.; Sarquis, D.P.; Khayat, A.S.; Khayat, B.C.M.; Demachki, S.; Anaissi, A.K.M.; Ishak, G.; Santos, N.P.C.; dos Santos, S.E.B.; Burbano, R.R.; et al. Gastric Cancer Microbiome. Pathobiology 2021, 88, 156–169. [Google Scholar] [CrossRef]
- Gunathilake, M.N.; Lee, J.; Choi, I.J.; Kim, Y.-I.; Ahn, Y.; Park, C.; Kim, J. Association between the Relative Abundance of Gastric Microbiota and the Risk of Gastric Cancer: A Case-Control Study. Sci. Rep. 2019, 9, 13589. [Google Scholar] [CrossRef] [PubMed]
- Montalban-Arques, A.; Wurm, P.; Trajanoski, S.; Schauer, S.; Kienesberger, S.; Halwachs, B.; Högenauer, C.; Langner, C.; Gorkiewicz, G. Propionibacterium Acnes Overabundance and Natural Killer Group 2 Member D System Activation in Corpus-Dominant Lymphocytic Gastritis. J. Pathol. 2016, 240, 425–436. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Chen, B.; Yang, T.; Xiao, W.; Qian, L.; Ding, Y.; Ji, M.; Ge, X.; Gong, W. Human Fused NKG2D–IL-15 Protein Controls Xenografted Human Gastric Cancer through the Recruitment and Activation of NK Cells. Cell. Mol. Immunol. 2017, 14, 293–307. [Google Scholar] [CrossRef]
- Liu, X.; Shao, L.; Liu, X.; Ji, F.; Mei, Y.; Cheng, Y.; Liu, F.; Yan, C.; Li, L.; Ling, Z. Alterations of Gastric Mucosal Microbiota across Different Stomach Microhabitats in a Cohort of 276 Patients with Gastric Cancer. EBioMedicine 2019, 40, 336–348. [Google Scholar] [CrossRef]
- Signat, B.; Roques, C.; Poulet, P.; Duffaut, D. Fusobacterium Nucleatum in Periodontal Health and Disease. Curr. Issues Mol. Biol. 2011, 13, 25–36. [Google Scholar] [PubMed]
- Boehm, E.T.; Thon, C.; Kupcinskas, J.; Steponaitiene, R.; Skieceviciene, J.; Canbay, A.; Malfertheiner, P.; Link, A. Fusobacterium Nucleatum Is Associated with Worse Prognosis in Lauren’s Diffuse Type Gastric Cancer Patients. Sci. Rep. 2020, 10, 16240. [Google Scholar] [CrossRef]
- Hsieh, Y.-Y.; Tung, S.-Y.; Pan, H.-Y.; Chang, T.-S.; Wei, K.-L.; Chen, W.-M.; Deng, Y.-F.; Lu, C.-K.; Lai, Y.-H.; Wu, C.-S.; et al. Fusobacterium Nucleatum Colonization is Associated with Decreased Survival of Helicobacter Pylori-Positive Gastric Cancer Patients. World J. Gastroenterol. 2021, 27, 7311–7323. [Google Scholar] [CrossRef]
- Safdar, A.; Rolston, K.V. Stenotrophomonas Maltophilia: Changing Spectrum of a Serious Bacterial Pathogen in Patients with Cancer. Clin. Infect. Dis. 2007, 45, 1602–1609. [Google Scholar] [CrossRef]
- Ling, Z.; Shao, L.; Liu, X.; Cheng, Y.; Yan, C.; Mei, Y.; Ji, F.; Liu, X. Regulatory T Cells and Plasmacytoid Dendritic Cells within the Tumor Microenvironment in Gastric Cancer Are Correlated with Gastric Microbiota Dysbiosis: A Preliminary Study. Front. Immunol. 2019, 10, 533. [Google Scholar] [CrossRef]
- Guo, Q.; Qin, H.; Liu, X.; Zhang, X.; Chen, Z.; Qin, T.; Chang, L.; Zhang, W. The Emerging Roles of Human Gut Microbiota in Gastrointestinal Cancer. Front. Immunol. 2022, 13, 2894. [Google Scholar] [CrossRef]
- Matson, V.; Chervin, C.S.; Gajewski, T.F. Cancer and the Microbiome—Influence of the Commensal Microbiota on Cancer, Immune Responses, and Immunotherapy. Gastroenterology 2021, 160, 600–613. [Google Scholar] [CrossRef] [PubMed]
- AL-Ishaq, R.K.; Koklesova, L.; Kubatka, P.; Büsselberg, D. Immunomodulation by Gut Microbiome on Gastrointestinal Cancers: Focusing on Colorectal Cancer. Cancers 2022, 14, 2140. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Majewski, M.; Mertowska, P.; Mertowski, S.; Smolak, K.; Grywalska, E.; Torres, K. Microbiota and the Immune System—Actors in the Gastric Cancer Story. Cancers 2022, 14, 3832. https://doi.org/10.3390/cancers14153832
Majewski M, Mertowska P, Mertowski S, Smolak K, Grywalska E, Torres K. Microbiota and the Immune System—Actors in the Gastric Cancer Story. Cancers. 2022; 14(15):3832. https://doi.org/10.3390/cancers14153832
Chicago/Turabian StyleMajewski, Marek, Paulina Mertowska, Sebastian Mertowski, Konrad Smolak, Ewelina Grywalska, and Kamil Torres. 2022. "Microbiota and the Immune System—Actors in the Gastric Cancer Story" Cancers 14, no. 15: 3832. https://doi.org/10.3390/cancers14153832
APA StyleMajewski, M., Mertowska, P., Mertowski, S., Smolak, K., Grywalska, E., & Torres, K. (2022). Microbiota and the Immune System—Actors in the Gastric Cancer Story. Cancers, 14(15), 3832. https://doi.org/10.3390/cancers14153832