
The Role of NOTCH1, GATA3, and c-MYC in T Cell Non-Hodgkin Lymphomas

 ,
,  , , , and
, , , and 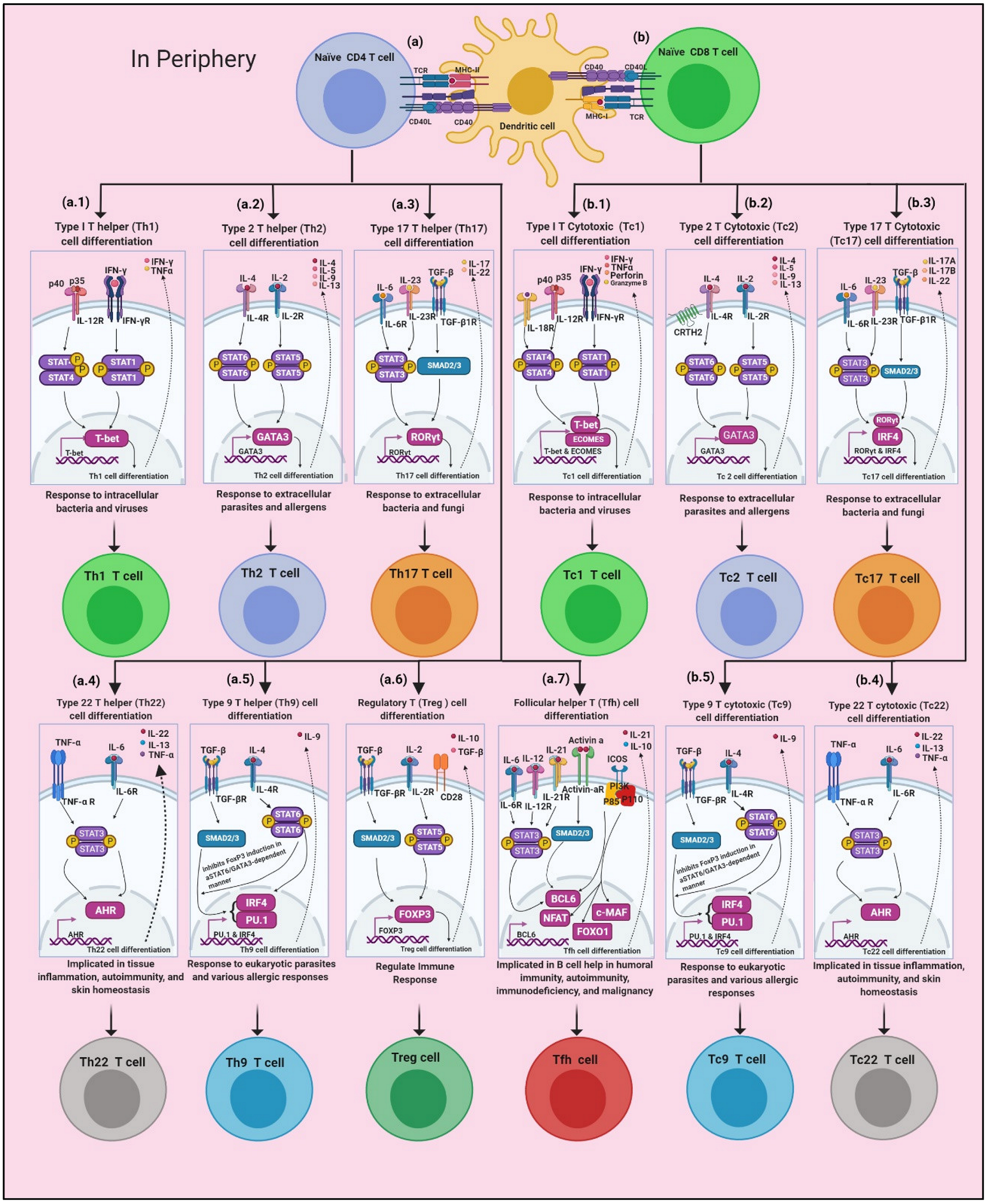{kind=link}
{kind=link}
{kind=link}
{kind=link}
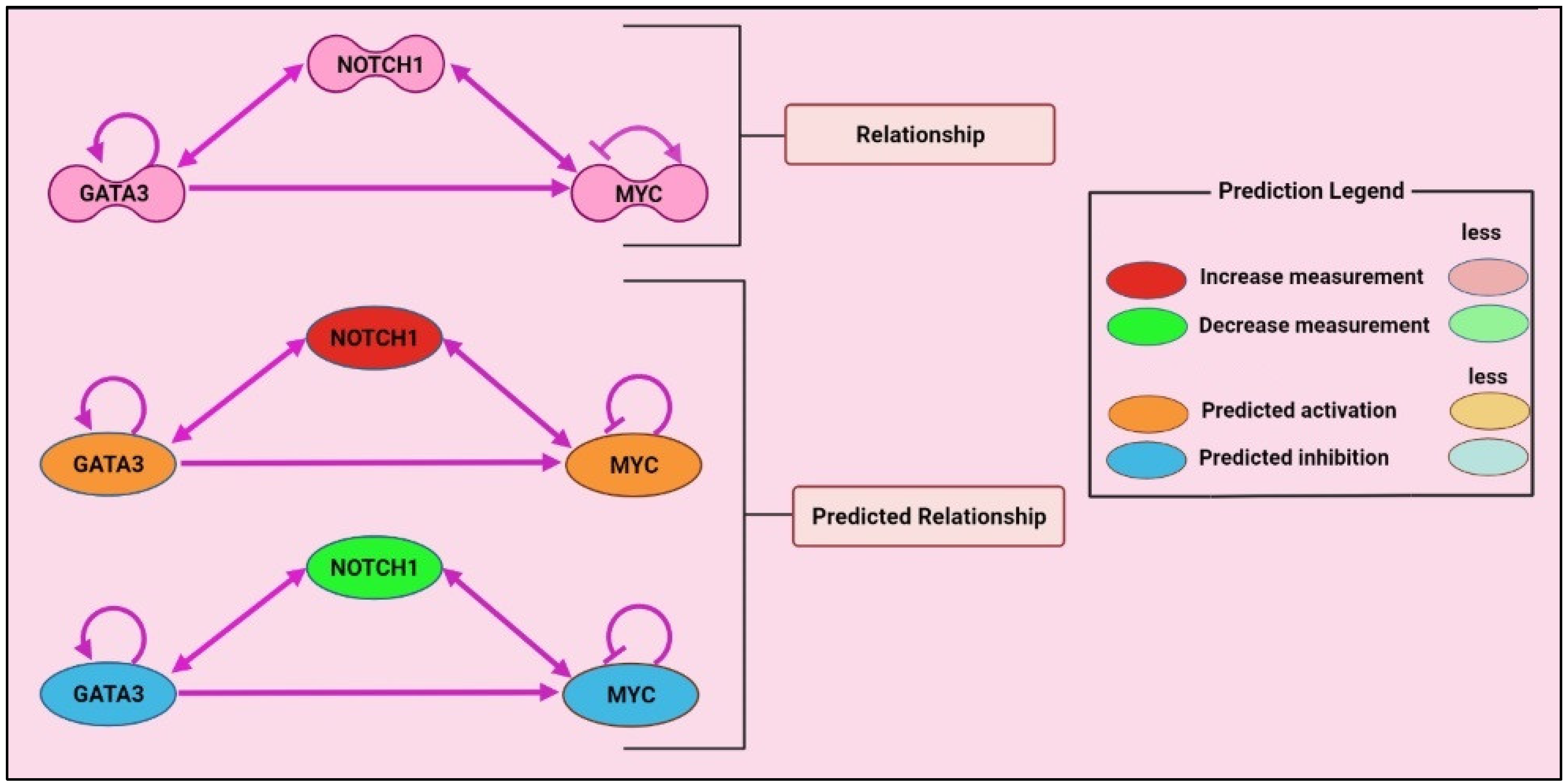{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
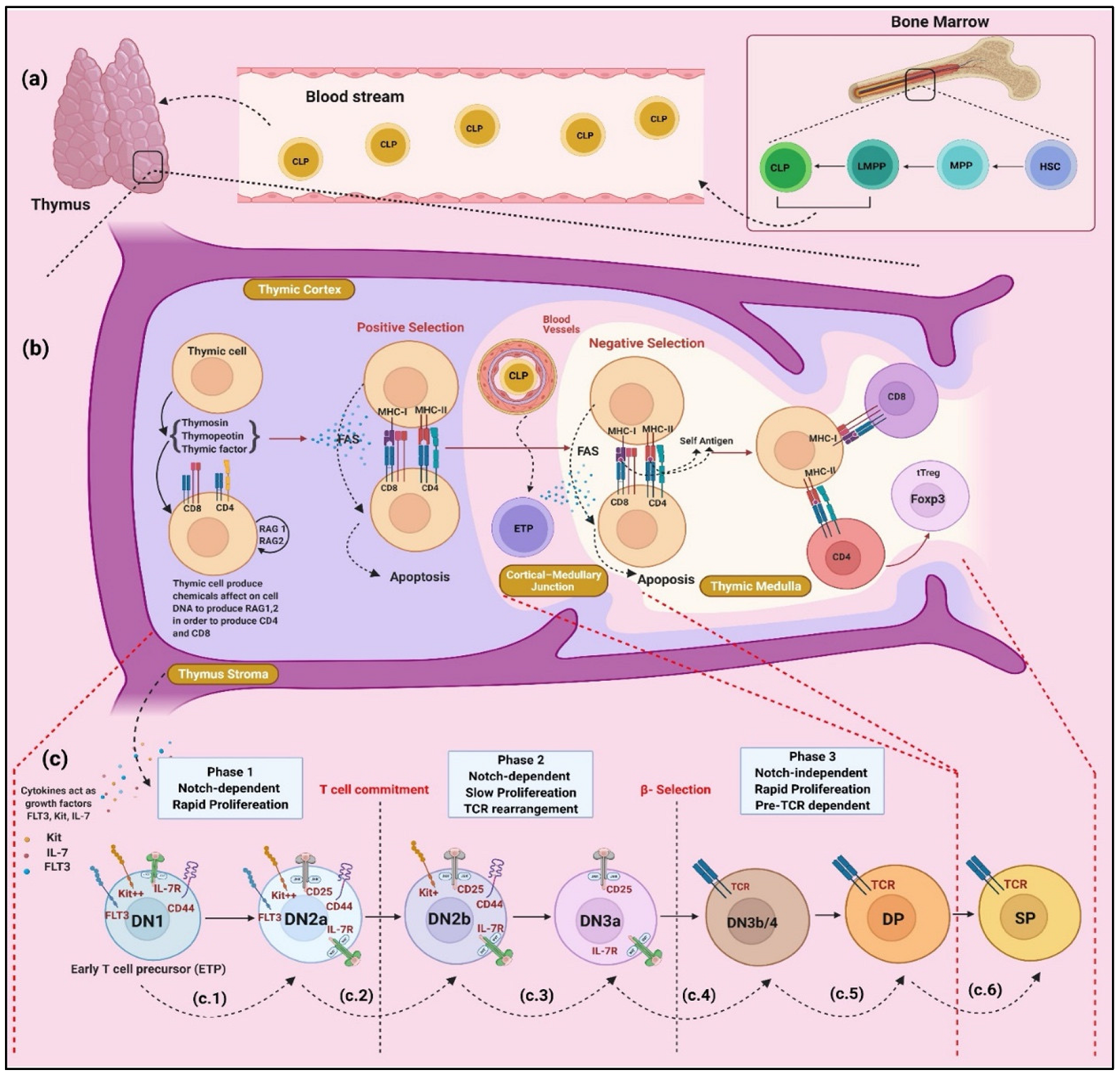
2. Development and Regulation of T Cells
3. NOTCH1 Signalling Pathways in T Cell Development and PTCLs
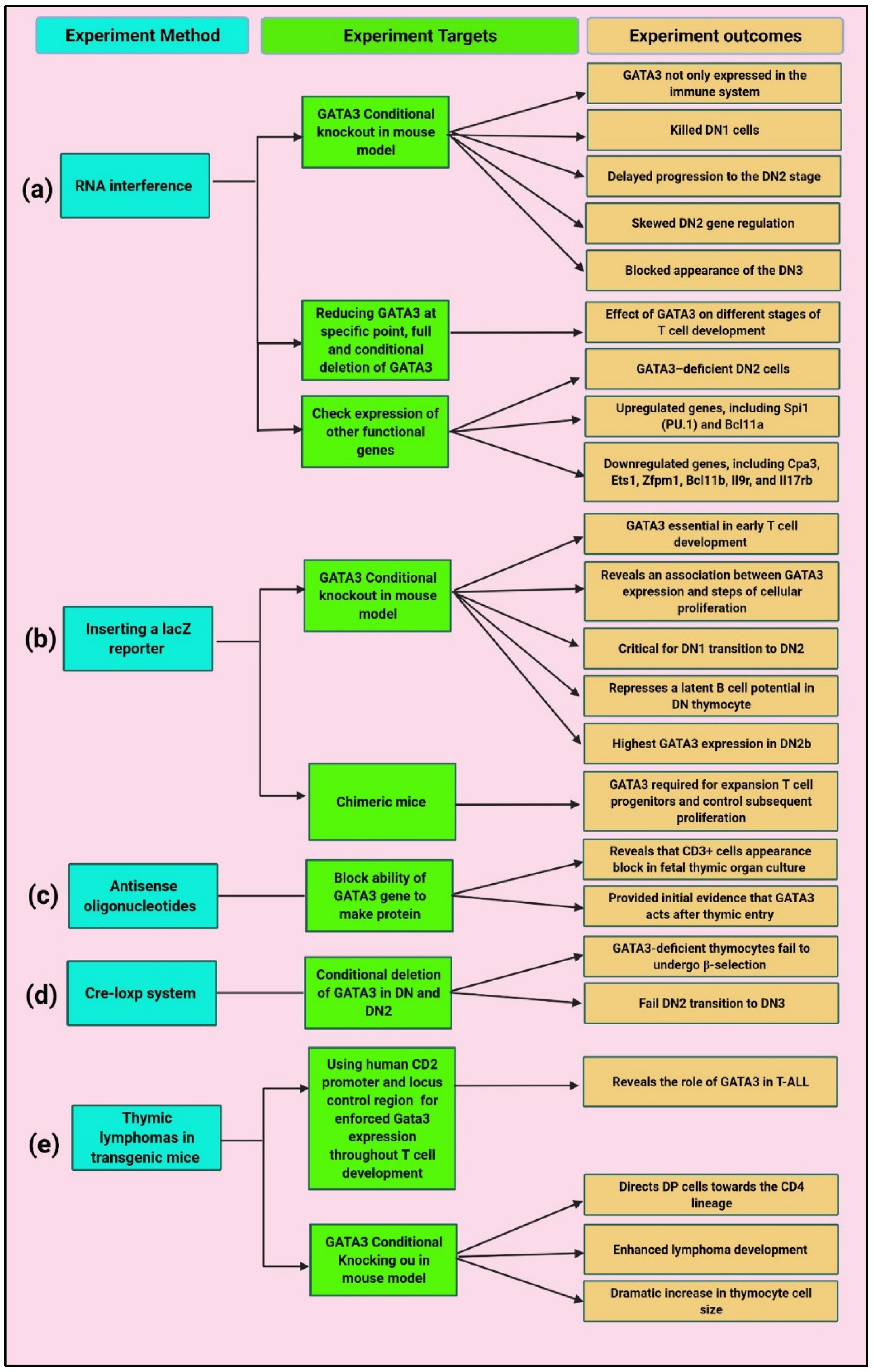
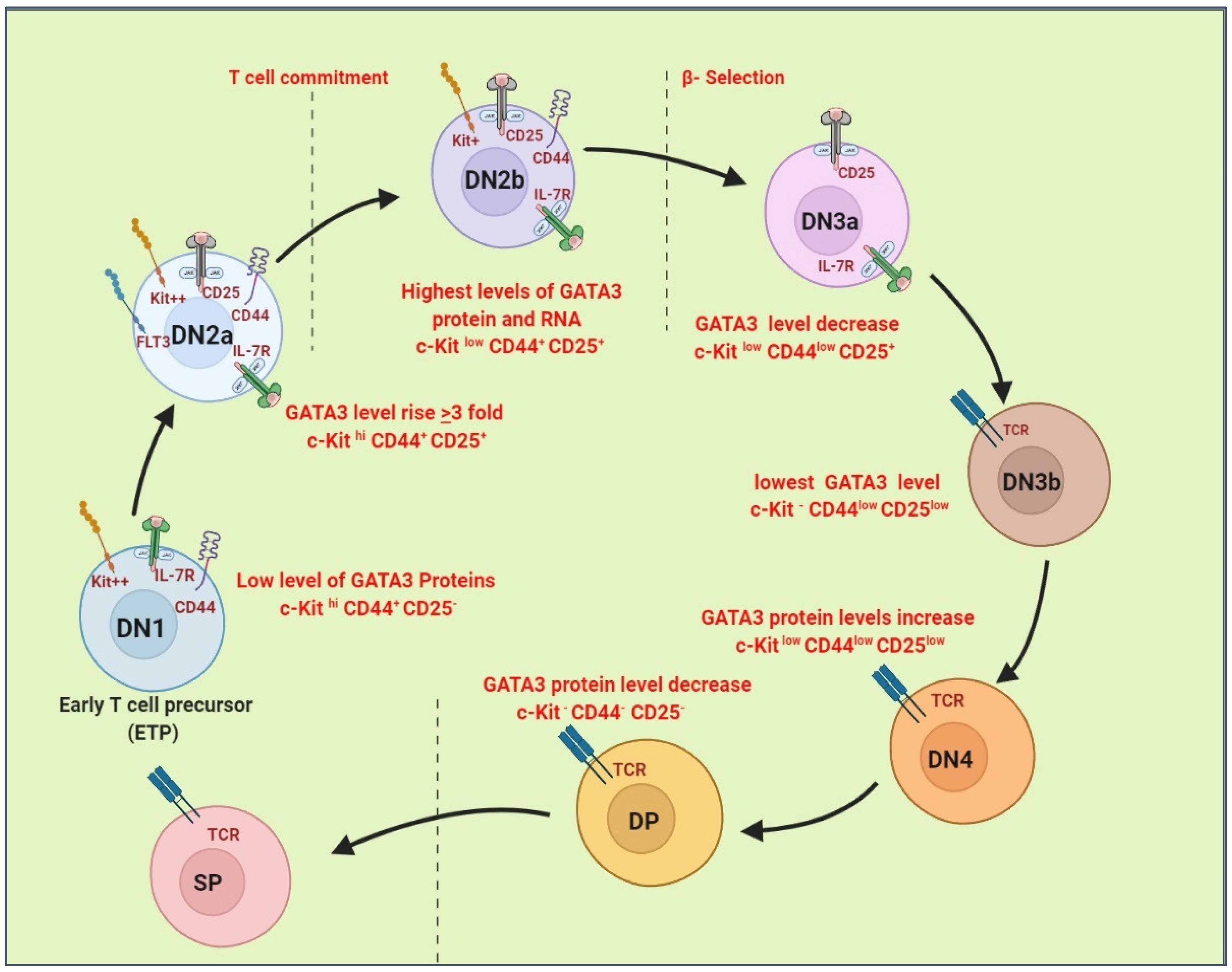
4. Roles of GATA3 in Normal T Cell Development and PTCLs
5. c-MYC Roles in Normal T Cell Development and PTCLs
6. NOTCH1 as a Bridge between GATA3 and c-MYC in T Cell Development
7. Conclusions
8. Future Directions and Therapeutic Targets
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Swerdlow, S.H.; Campo, E.; Pileri, S.A.; Harris, N.L.; Stein, H.; Siebert, R.; Advani, R.; Ghielmini, M.; Salles, G.A.; Zelenetz, A.D.; et al. The 2016 revision of the World Health Organization classification of lymphoid neoplasms. Blood 2016, 127, 2375–2390. [Google Scholar] [CrossRef] [PubMed]
- Arber, D.A.; Orazi, A.; Hasserjian, R.; Thiele, J.; Borowitz, M.J.; Le Beau, M.M.; Bloomfield, C.D.; Cazzola, M.; Vardiman, J.W. The 2016 revision to the World Health Organization classification of myeloid neoplasms and acute leukemia. Blood 2016, 127, 2391–2405. [Google Scholar] [CrossRef] [PubMed]
- Swerdlow, S.H.; Campo, E.; Harris, N.L.; Jaffe, E.S.; Pileri, S.A.; Stein, H.; Thiele, J.; Vardiman, J.W. WHO Classification of Tumours of Haematopoietic and Lymphoid Tissues; International Agency for Research on Cancer: Lyon, France, 2008; Volume 2. [Google Scholar]
- Pileri, S.A.; Ascani, S.; Leoncini, L.; Sabattini, E.; Zinzani, P.L.; Piccaluga, P.P.; Pileri, A., Jr.; Giunti, M.; Falini, B.; Bolis, G.B.; et al. Hodgkin’s lymphoma: The pathologist’s viewpoint. J. Clin. Pathol. 2002, 55, 162–176. [Google Scholar] [CrossRef] [PubMed]
- PDQ Adult Treatment Editorial Board. Adult Non-Hodgkin Lymphoma Treatment (PDQ®). In PDQ Cancer Information Summaries; National Cancer Institute: Bethesda, MD, USA, 2021. [Google Scholar]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Satou, A.; Bennani, N.N.; Feldman, A.L. Update on the classification of T-cell lymphomas, Hodgkin lymphomas, and histiocytic/dendritic cell neoplasms. Expert Rev. Hematol. 2019, 12, 833–843. [Google Scholar] [CrossRef]
- De Leval, L. Approach to nodal-based T-cell lymphomas. Pathology 2020, 52, 78–99. [Google Scholar] [CrossRef]
- Pizzi, M.; Margolskee, E.; Inghirami, G. Pathogenesis of Peripheral T Cell Lymphoma. Annu. Rev. Pathol. 2018, 13, 293–320. [Google Scholar] [CrossRef]
- De Leval, L.; Rickman, D.S.; Thielen, C.; Reynies, A.d.; Huang, Y.-L.; Delsol, G.; Lamant, L.; Leroy, K.; Briere, J.; Molina, T.J.B. The gene expression profile of nodal peripheral T-cell lymphoma demonstrates a molecular link between angioimmunoblastic T-cell lymphoma (AITL) and follicular helper T (TFH) cells. J. Am. Soc. Hematol. 2007, 109, 4952–4963. [Google Scholar] [CrossRef]
- Sakaguchi, S. Naturally arising CD4+ regulatory t cells for immunologic self-tolerance and negative control of immune responses. Annu. Rev. Immunol. 2004, 22, 531–562. [Google Scholar] [CrossRef]
- Kim, W.; Jeon, Y.K.; Kim, T.; Kim, J.E.; Kim, Y.; Lee, S.-H.; Kim, D.-W.; Heo, D.S.; Kim, C.-W. Increased quantity of tumor-infiltrating FOXP3-positive regulatory T cells is an independent predictor for improved clinical outcome in extranodal NK/T-cell lymphoma. Ann. Oncol. 2009, 20, 1688–1696. [Google Scholar] [CrossRef]
- Karwacz, K.; Bricogne, C.; MacDonald, D.; Arce, F.; Bennett, C.L.; Collins, M.; Escors, D. PD-L1 co-stimulation contributes to ligand-induced T cell receptor down-modulation on CD8+ T cells. EMBO Mol. Med. 2011, 3, 581–592. [Google Scholar] [CrossRef] [PubMed]
- Kasprzycka, M.; Marzec, M.; Liu, X.; Zhang, Q.; Wasik, M.A. Nucleophosmin/anaplastic lymphoma kinase (NPM/ALK) oncoprotein induces the T regulatory cell phenotype by activating STAT3. Proc. Natl. Acad. Sci. USA 2006, 103, 9964–9969. [Google Scholar] [CrossRef]
- Wilcox, R.A.; Feldman, A.L.; Wada, D.A.; Yang, Z.-Z.; Comfere, N.I.; Dong, H.; Kwon, E.D.; Novak, A.J.; Markovic, S.N.; Pittelkow, M.R.; et al. B7-H1 (PD-L1, CD274) suppresses host immunity in T-cell lymphoproliferative disorders. J. Am. Soc. Hematol. 2009, 114, 2149–2158. [Google Scholar] [CrossRef] [PubMed]
- Jo, J.-C.; Kim, M.; Choi, Y.; Kim, H.-J.; Kim, J.E.; Chae, S.W.; Kim, H.; Cha, H.J. Expression of programmed cell death 1 and programmed cell death ligand 1 in extranodal NK/T-cell lymphoma, nasal type. Ann. Hematol. 2017, 96, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.-X.; Bai, B.; Cai, Q.-C.; Cai, Q.-Q.; Wang, X.-X.; Wu, X.-Y.; Huang, H.-Q. High numbers of tumor-associated macrophages correlate with poor prognosis in patients with mature T-and natural killer cell lymphomas. Med. Oncol. 2012, 29, 3522–3528. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, J.; Liu, Z.; Deffenbacher, K.; Chan, W.C.J.B.P.; Haematology, R.C. Gene expression profiling in lymphoma diagnosis and management. Best Pract. Res. Clin. Haematol. 2009, 22, 191–210. [Google Scholar] [CrossRef]
- Zain, J.M. Aggressive T-cell lymphomas: 2019 updates on diagnosis, risk stratification, and management. Am. J. Hematol. 2019, 94, 929–946. [Google Scholar] [CrossRef]
- Piccaluga, P.P.; Fuligni, F.; De Leo, A.; Bertuzzi, C.; Rossi, M.; Bacci, F.; Sabattini, E.; Agostinelli, C.; Gazzola, A.; Laginestra, M.A. Molecular profiling improves classification and prognostication of nodal peripheral T-cell lymphomas: Results of a phase III diagnostic accuracy study. J. Clin. Oncol. 2013, 31, 3019–3025. [Google Scholar] [CrossRef]
- Iqbal, J.; Amador, C.; McKeithan, T.W.; Chan, W.C. Molecular and genomic landscape of peripheral T-cell lymphoma. In T-Cell and NK-Cell Lymphomas; Springer: Berlin/Heidelberg, Germany, 2019; pp. 31–68. [Google Scholar]
- Van Hamburg, J.P.; de Bruijn, M.J.; Dingjan, G.M.; Beverloo, H.B.; Diepstraten, H.; Ling, K.W.; Hendriks, R.W. Cooperation of Gata3, c-Myc and Notch in malignant transformation of double positive thymocytes. Mol. Immunol. 2008, 45, 3085–3095. [Google Scholar] [CrossRef]
- Iqbal, J.; Wright, G.; Wang, C.; Rosenwald, A.; Gascoyne, R.D.; Weisenburger, D.D.; Greiner, T.C.; Smith, L.; Guo, S.; Wilcox, R.A.; et al. Gene expression signatures delineate biological and prognostic subgroups in peripheral T-cell lymphoma. J. Am. Soc. Hematol. 2014, 123, 2915–2923. [Google Scholar] [CrossRef]
- Manso, R.; Bellas, C.; Martín-Acosta, P.; Mollejo, M.; Menárguez, J.; Rojo, F.; Llamas, P.; Piris, M.A.; Rodríguez-Pinilla, S.M. C-MYC is related to GATA3 expression and associated with poor prognosis in nodal peripheral T-cell lymphomas. Haematologica 2016, 101, e336. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Belver, L.; Yang, A.Y.; Albero, R.; Herranz, D.; Brundu, F.G.; Quinn, S.A.; Perez-Duran, P.; Alvarez, S.; Gianni, F.; Rashkovan, M.; et al. GATA3-Controlled Nucleosome Eviction Drives MYC Enhancer Activity in T-cell Development and Leukemia. Cancer Discov. 2019, 9, 1774–1791. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Bevan, M.J. CD8(+) T cells: Foot soldiers of the immune system. Immunity 2011, 35, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Perry, S.S.; Wang, H.; Pierce, L.J.; Yang, A.M.; Tsai, S.; Spangrude, G.J. L-selectin defines a bone marrow analog to the thymic early T-lineage progenitor. Blood 2004, 103, 2990–2996. [Google Scholar] [CrossRef] [PubMed]
- Ghaedi, M.; Steer, C.A.; Martinez-Gonzalez, I.; Halim, T.Y.F.; Abraham, N.; Takei, F. Common-Lymphoid-Progenitor-Independent Pathways of Innate and T Lymphocyte Development. Cell Rep. 2016, 15, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Yui, M.A.; Rothenberg, E.V. Developmental gene networks: A triathlon on the course to T cell identity. Nat. Rev. Immunol. 2014, 14, 529–545. [Google Scholar] [CrossRef] [PubMed]
- Artavanis-Tsakonas, S.; Rand, M.D.; Lake, R.J. Notch signaling: Cell fate control and signal integration in development. Science 1999, 284, 770–776. [Google Scholar] [CrossRef]
- Almotiri, A.; Alzahrani, H.; Menendez-Gonzalez, J.B.; Abdelfattah, A.; Alotaibi, B.; Saleh, L.; Greene, A.; Georgiou, M.; Gibbs, A.; Alsayari, A.; et al. Zeb1 modulates hematopoietic stem cell fates required for suppressing acute myeloid leukemia. J. Clin. Investig. 2021, 131, e129115. [Google Scholar] [CrossRef]
- Kueh, H.Y.; Yui, M.A.; Ng, K.K.; Pease, S.S.; Zhang, J.A.; Damle, S.S.; Freedman, G.; Siu, S.; Bernstein, I.D.; Elowitz, M.B.; et al. Asynchronous combinatorial action of four regulatory factors activates Bcl11b for T cell commitment. Nat. Immunol. 2016, 17, 956–965. [Google Scholar] [CrossRef]
- Li, L.; Leid, M.; Rothenberg, E.V. An early T cell lineage commitment checkpoint dependent on the transcription factor Bcl11b. Science 2010, 329, 89–93. [Google Scholar] [CrossRef]
- Agata, Y.; Tamaki, N.; Sakamoto, S.; Ikawa, T.; Masuda, K.; Kawamoto, H.; Murre, C. Regulation of T cell receptor beta gene rearrangements and allelic exclusion by the helix-loop-helix protein, E47. Immunity 2007, 27, 871–884. [Google Scholar] [CrossRef] [PubMed]
- Wojciechowski, J.; Lai, A.; Kondo, M.; Zhuang, Y. E2A and HEB are required to block thymocyte proliferation prior to pre-TCR expression. J. Immunol. 2007, 178, 5717–5726. [Google Scholar] [CrossRef] [PubMed]
- Georgescu, C.; Longabaugh, W.J.; Scripture-Adams, D.D.; David-Fung, E.S.; Yui, M.A.; Zarnegar, M.A.; Bolouri, H.; Rothenberg, E.V. A gene regulatory network armature for T lymphocyte specification. Proc. Natl. Acad. Sci. USA 2008, 105, 20100–20105. [Google Scholar] [CrossRef]
- Schwartz, R.; Engel, I.; Fallahi-Sichani, M.; Petrie, H.T.; Murre, C. Gene expression patterns define novel roles for E47 in cell cycle progression, cytokine-mediated signaling, and T lineage development. Proc. Natl. Acad. Sci. USA 2006, 103, 9976–9981. [Google Scholar] [CrossRef] [PubMed]
- Welinder, E.; Mansson, R.; Mercer, E.M.; Bryder, D.; Sigvardsson, M.; Murre, C. The transcription factors E2A and HEB act in concert to induce the expression of FOXO1 in the common lymphoid progenitor. Proc. Natl. Acad. Sci. USA 2011, 108, 17402–17407. [Google Scholar] [CrossRef] [PubMed]
- Maillard, I.; Tu, L.; Sambandam, A.; Yashiro-Ohtani, Y.; Millholland, J.; Keeshan, K.; Shestova, O.; Xu, L.; Bhandoola, A.; Pear, W.S. The requirement for Notch signaling at the β-selection checkpoint in vivo is absolute and independent of the pre–T cell receptor. J. Exp. Med. 2006, 203, 2239–2245. [Google Scholar] [CrossRef] [PubMed]
- Kreslavsky, T.; Gleimer, M.; Miyazaki, M.; Choi, Y.; Gagnon, E.; Murre, C.; Sicinski, P.; von Boehmer, H.J.I. β-Selection-induced proliferation is required for αβ T cell differentiation. Immunity 2012, 37, 840–853. [Google Scholar] [CrossRef]
- Tussiwand, R.; Engdahl, C.; Gehre, N.; Bosco, N.; Ceredig, R.; Rolink, A. The preTCR-dependent DN3 to DP transition requires Notch signaling, is improved by CXCL12 signaling and is inhibited by IL-7 signaling. Eur. J. Immunol. 2011, 41, 3371–3380. [Google Scholar] [CrossRef]
- Janas, M.L.; Varano, G.; Gudmundsson, K.; Noda, M.; Nagasawa, T.; Turner, M. Thymic development beyond beta-selection requires phosphatidylinositol 3-kinase activation by CXCR4. J. Exp. Med. 2010, 207, 247–261. [Google Scholar] [CrossRef]
- Hosokawa, H.; Rothenberg, E.V. Cytokines, Transcription Factors, and the Initiation of T-Cell Development. Cold Spring Harb Perspect. Biol. 2018, 10, a028621. [Google Scholar] [CrossRef]
- Kelliher, M.A.; Roderick, J.E. NOTCH Signaling in T-Cell-Mediated Anti-Tumor Immunity and T-Cell-Based Immunotherapies. Front. Immunol. 2018, 9, 1718. [Google Scholar] [CrossRef] [PubMed]
- Artavanis-Tsakonas, S.; Matsuno, K.; Fortini, M.E. Notch signaling. Science 1995, 268, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Laky, K.; Fowlkes, B.J. Notch signaling in CD4 and CD8 T cell development. Curr. Opin. Immunol. 2008, 20, 197–202. [Google Scholar] [CrossRef]
- Huang, E.Y.; Gallegos, A.M.; Richards, S.M.; Lehar, S.M.; Bevan, M.J. Surface expression of Notch1 on thymocytes: Correlation with the double-negative to double-positive transition. J. Immunol. 2003, 171, 2296–2304. [Google Scholar] [CrossRef] [PubMed]
- Garbe, A.I.; von Boehmer, H. TCR and Notch synergize in alphabeta versus gammadelta lineage choice. Trends Immunol. 2007, 28, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Blaumueller, C.M.; Qi, H.; Zagouras, P.; Artavanis-Tsakonas, S. Intracellular cleavage of Notch leads to a heterodimeric receptor on the plasma membrane. Cell 1997, 90, 281–291. [Google Scholar] [CrossRef]
- Rebay, I.; Fleming, R.J.; Fehon, R.G.; Cherbas, L.; Cherbas, P.; Artavanis-Tsakonas, S.J.C. Specific EGF repeats of Notch mediate interactions with Delta and Serrate: Implications for Notch as a multifunctional receptor. Cell 1991, 67, 687–699. [Google Scholar] [CrossRef]
- Ntziachristos, P.; Tsirigos, A.; Van Vlierberghe, P.; Nedjic, J.; Trimarchi, T.; Flaherty, M.S.; Ferres-Marco, D.; da Ros, V.; Tang, Z.; Siegle, J.; et al. Genetic inactivation of the polycomb repressive complex 2 in T cell acute lymphoblastic leukemia. Nat. Med. 2012, 18, 298–301. [Google Scholar] [CrossRef]
- Kopan, R.; Ilagan, M.X. The canonical Notch signaling pathway: Unfolding the activation mechanism. Cell 2009, 137, 216–233. [Google Scholar] [CrossRef]
- Le Borgne, R.; Bardin, A.; Schweisguth, F. The roles of receptor and ligand endocytosis in regulating Notch signaling. Development 2005, 132, 1751–1762. [Google Scholar] [CrossRef]
- Sundaram, M.; Greenwald, I.J.G. Suppressors of a lin-12 hypomorph define genes that interact with both lin-12 and glp-1 in Caenorhabditis elegans. Genet. Soc. Am. 1993, 135, 765–783. [Google Scholar] [CrossRef] [PubMed]
- Kooijman, R.K.; Scholtens, L.E.; Rijkers, G.T.; Zegers, B.J. Differential expression of type I insulin-like growth factor receptors in different stages of human T cells. Eur. J. Immunol. 1995, 25, 931–935. [Google Scholar] [CrossRef] [PubMed]
- Tanigaki, K.; Honjo, T. Regulation of lymphocyte development by Notch signaling. Nat. Immunol. 2007, 8, 451–456. [Google Scholar] [CrossRef] [PubMed]
- Weng, A.P.; Ferrando, A.A.; Lee, W.; Morris, J.P.t.; Silverman, L.B.; Sanchez-Irizarry, C.; Blacklow, S.C.; Look, A.T.; Aster, J.C. Activating mutations of NOTCH1 in human T cell acute lymphoblastic leukemia. Science 2004, 306, 269–271. [Google Scholar] [CrossRef]
- Malyukova, A.; Dohda, T.; von der Lehr, N.; Akhondi, S.; Corcoran, M.; Heyman, M.; Spruck, C.; Grandér, D.; Lendahl, U.; Sangfelt, O. The tumor suppressor gene hCDC4 is frequently mutated in human T-cell acute lymphoblastic leukemia with functional consequences for Notch signaling. Cancer Res. 2007, 67, 5611–5616. [Google Scholar] [CrossRef]
- O’Neil, J.; Grim, J.; Strack, P.; Rao, S.; Tibbitts, D.; Winter, C.; Hardwick, J.; Welcker, M.; Meijerink, J.P.; Pieters, R.; et al. FBW7 mutations in leukemic cells mediate NOTCH pathway activation and resistance to gamma-secretase inhibitors. J. Exp. Med. 2007, 204, 1813–1824. [Google Scholar] [CrossRef]
- Thompson, B.J.; Buonamici, S.; Sulis, M.L.; Palomero, T.; Vilimas, T.; Basso, G.; Ferrando, A.; Aifantis, I. The SCFFBW7 ubiquitin ligase complex as a tumor suppressor in T cell leukemia. J. Exp. Med. 2007, 204, 1825–1835. [Google Scholar] [CrossRef]
- Clurman, B.E.; Sheaff, R.J.; Thress, K.; Groudine, M.; Roberts, J.M. Turnover of cyclin E by the ubiquitin-proteasome pathway is regulated by cdk2 binding and cyclin phosphorylation. Genes Dev. 1996, 10, 1979–1990. [Google Scholar] [CrossRef]
- Inuzuka, H.; Shaik, S.; Onoyama, I.; Gao, D.; Tseng, A.; Maser, R.S.; Zhai, B.; Wan, L.; Gutierrez, A.; Lau, A.W.; et al. SCF(FBW7) regulates cellular apoptosis by targeting MCL1 for ubiquitylation and destruction. Nature 2011, 471, 104–109. [Google Scholar] [CrossRef]
- Sugimoto, K.; Himeno, M.J.B. Biotechnology, Biochemistry. Casein kinase II site of human centromere protein B (CENP-B) is phosphorylated in vitro. Biosci. Biotechnol. 1992, 56, 1174–1175. [Google Scholar] [CrossRef]
- Tan, Y.; Sangfelt, O.; Spruck, C. The Fbxw7/hCdc4 tumor suppressor in human cancer. Cancer Lett. 2008, 271, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Jin, J.; Schlisio, S.; Harper, J.W.; Kaelin, W.G., Jr. The v-Jun point mutation allows c-Jun to escape GSK3-dependent recognition and destruction by the Fbw7 ubiquitin ligase. Cancer Cell 2005, 8, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Ellisen, L.W.; Bird, J.; West, D.C.; Soreng, A.L.; Reynolds, T.C.; Smith, S.D.; Sklar, J. TAN-1, the human homolog of the Drosophila notch gene, is broken by chromosomal translocations in T lymphoblastic neoplasms. Cell 1991, 66, 649–661. [Google Scholar] [CrossRef]
- Li, X.; von Boehmer, H. Notch Signaling in T-Cell Development and T-ALL. ISRN Hematol. 2011, 2011, 921706. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Nieva, P.; Santos, J.; Fernandez-Piqueras, J. Defective expression of Notch1 and Notch2 in connection to alterations of c-Myc and Ikaros in gamma-radiation-induced mouse thymic lymphomas. Carcinogenesis 2004, 25, 1299–1304. [Google Scholar] [CrossRef] [PubMed]
- Bellavia, D.; Campese, A.F.; Alesse, E.; Vacca, A.; Felli, M.P.; Balestri, A.; Stoppacciaro, A.; Tiveron, C.; Tatangelo, L.; Giovarelli, M.; et al. Constitutive activation of NF-κB and T-cell leukemia/lymphoma in Notch3 transgenic mice. EMBO J. 2000, 19, 3337–3348. [Google Scholar] [CrossRef]
- Vercauteren, S.M.; Sutherland, H.J. Constitutively active Notch4 promotes early human hematopoietic progenitor cell maintenance while inhibiting differentiation and causes lymphoid abnormalities in vivo. Blood 2004, 104, 2315–2322. [Google Scholar] [CrossRef]
- Jundt, F.; Anagnostopoulos, I.; Forster, R.; Mathas, S.; Stein, H.; Dorken, B. Activated Notch1 signaling promotes tumor cell proliferation and survival in Hodgkin and anaplastic large cell lymphoma. Blood 2002, 99, 3398–3403. [Google Scholar] [CrossRef]
- Kamstrup, M.R.; Ralfkiaer, E.; Skovgaard, G.L.; Gniadecki, R. Potential involvement of Notch1 signalling in the pathogenesis of primary cutaneous CD30-positive lymphoproliferative disorders. Br. J. Dermatol. 2008, 158, 747–753. [Google Scholar] [CrossRef]
- Indraccolo, S.; Minuzzo, S.; Masiero, M.; Pusceddu, I.; Persano, L.; Moserle, L.; Reboldi, A.; Favaro, E.; Mecarozzi, M.; Di Mario, G.; et al. Cross-talk between tumor and endothelial cells involving the Notch3-Dll4 interaction marks escape from tumor dormancy. AACR 2009, 69, 1314–1323. [Google Scholar] [CrossRef]
- Palomero, T.; Sulis, M.L.; Cortina, M.; Real, P.J.; Barnes, K.; Ciofani, M.; Caparros, E.; Buteau, J.; Brown, K.; Perkins, S.L.; et al. Mutational loss of PTEN induces resistance to NOTCH1 inhibition in T-cell leukemia. Nat. Med. 2007, 13, 1203–1210. [Google Scholar] [CrossRef] [PubMed]
- Palomero, T.; Dominguez, M.; Ferrando, A.A. The role of the PTEN/AKT Pathway in NOTCH1-induced leukemia. Cell Cycle 2008, 7, 965–970. [Google Scholar] [CrossRef] [PubMed]
- Joshi, I.; Minter, L.M.; Telfer, J.; Demarest, R.M.; Capobianco, A.J.; Aster, J.C.; Sicinski, P.; Fauq, A.; Golde, T.E.; Osborne, B.A. Notch signaling mediates G1/S cell-cycle progression in T cells via cyclin D3 and its dependent kinases. Blood 2009, 113, 1689–1698. [Google Scholar] [CrossRef] [PubMed]
- Kamstrup, M.R.; Biskup, E.; Gjerdrum, L.M.; Ralfkiaer, E.; Niazi, O.; Gniadecki, R. The importance of Notch signaling in peripheral T-cell lymphomas. Leuk. Lymphoma 2014, 55, 639–644. [Google Scholar] [CrossRef]
- Kamstrup, M.R.; Gjerdrum, L.M.R.; Biskup, E.; Thyssing Lauenborg, B.; Ralfkiaer, E.; Woetmann, A.; Ødum, N.; Gniadecki, R. Notch1 as a potential therapeutic target in cutaneous T-cell lymphoma. Blood 2010, 116, 2504–2512. [Google Scholar] [CrossRef]
- Kamstrup, M.R.; Biskup, E.; Gniadecki, R. Notch signalling in primary cutaneous CD30+ lymphoproliferative disorders: A new therapeutic approach? Br. J. Dermatol. 2010, 163, 781–788. [Google Scholar] [CrossRef]
- Van Der Fits, L.; Qin, Y.; Out-Luiting, J.J.; Vermeer, K.G.; Whittaker, S.; Van Es, J.H.; Tensen, C.P.; Vermeer, M.H. NOTCH1 signaling as a therapeutic target in Sezary syndrome. J. Investig. Dermatol. 2012, 132, 2810–2817. [Google Scholar] [CrossRef]
- Katsumura, K.R.; Bresnick, E.H.; Group, G.F.M. The GATA factor revolution in hematology. Blood 2017, 129, 2092–2102. [Google Scholar] [CrossRef]
- Dovat, S.; Ronni, T.; Russell, D.; Ferrini, R.; Cobb, B.S.; Smale, S.T. A common mechanism for mitotic inactivation of C2H2 zinc finger DNA-binding domains. Genes Dev. 2002, 16, 2985–2990. [Google Scholar] [CrossRef]
- Ting, C.-N.; Olson, M.C.; Barton, K.P.; Leiden, J.M. Transcription factor GATA-3 is required for development of the T-cell lineage. Nature 1996, 384, 474–478. [Google Scholar] [CrossRef]
- Pai, S.Y.; Truitt, M.L.; Ting, C.N.; Leiden, J.M.; Glimcher, L.H.; Ho, I.C. Critical roles for transcription factor GATA-3 in thymocyte development. Immunity 2003, 19, 863–875. [Google Scholar] [CrossRef]
- Vosshenrich, C.A.; Garcia-Ojeda, M.E.; Samson-Villeger, S.I.; Pasqualetto, V.; Enault, L.; Richard-Le Goff, O.; Corcuff, E.; Guy-Grand, D.; Rocha, B.; Cumano, A.; et al. A thymic pathway of mouse natural killer cell development characterized by expression of GATA-3 and CD127. Nat. Immunol. 2006, 7, 1217–1224. [Google Scholar] [CrossRef] [PubMed]
- Samson, S.I.; Richard, O.; Tavian, M.; Ranson, T.; Vosshenrich, C.A.; Colucci, F.; Buer, J.; Grosveld, F.; Godin, I.; Di Santo, J.P. GATA-3 promotes maturation, IFN-gamma production, and liver-specific homing of NK cells. Immunity 2003, 19, 701–711. [Google Scholar] [CrossRef]
- Wang, Y.; Su, M.A.; Wan, Y.Y. An essential role of the transcription factor GATA-3 for the function of regulatory T cells. Immunity 2011, 35, 337–348. [Google Scholar] [CrossRef] [PubMed]
- Hoyler, T.; Klose, C.S.; Souabni, A.; Turqueti-Neves, A.; Pfeifer, D.; Rawlins, E.L.; Voehringer, D.; Busslinger, M.; Diefenbach, A. The transcription factor GATA-3 controls cell fate and maintenance of type 2 innate lymphoid cells. Immunity 2012, 37, 634–648. [Google Scholar] [CrossRef] [PubMed]
- Ho, I.-C.; Pai, S.-Y. GATA-3-not just for Th2 cells anymore. Cell Mol. Immunol. 2007, 4, 15–29. [Google Scholar]
- Ho, I.-C.; Tai, T.-S.; Pai, S.-Y. GATA3 and the T-cell lineage: Essential functions before and after T-helper-2-cell differentiation. Nat. Rev. Immunol. 2009, 9, 125–135. [Google Scholar] [CrossRef]
- Hattori, N.; Kawamoto, H.; Fujimoto, S.; Kuno, K.; Katsura, Y. Involvement of transcription factors TCF-1 and GATA-3 in the initiation of the earliest step of T cell development in the thymus. J. Exp. Med. 1996, 184, 1137–1147. [Google Scholar] [CrossRef]
- García-Ojeda, M.E.; Klein Wolterink, R.G.; Lemaître, F.; Richard-Le Goff, O.; Hasan, M.; Hendriks, R.W.; Cumano, A.; Di Santo, J.P. GATA-3 promotes T-cell specification by repressing B-cell potential in pro–T cells in mice. Blood 2013, 121, 1749–1759. [Google Scholar] [CrossRef]
- Wei, G.; Abraham, B.J.; Yagi, R.; Jothi, R.; Cui, K.; Sharma, S.; Narlikar, L.; Northrup, D.L.; Tang, Q.; Paul, W.E.; et al. Genome-wide analyses of transcription factor GATA3-mediated gene regulation in distinct T cell types. Immunity 2011, 35, 299–311. [Google Scholar] [CrossRef]
- Zhang, J.A.; Mortazavi, A.; Williams, B.A.; Wold, B.J.; Rothenberg, E.V. Dynamic transformations of genome-wide epigenetic marking and transcriptional control establish T cell identity. Cell 2012, 149, 467–482. [Google Scholar] [CrossRef] [PubMed]
- Hendriks, R.W.; Nawijn, M.C.; Engel, J.D.; van Doorninck, H.; Grosveld, F.; Karis, A. Expression of the transcription factor GATA-3 is required for the development of the earliest T cell progenitors and correlates with stages of cellular proliferation in the thymus. Eur. J. Immunol. 1999, 29, 1912–1918. [Google Scholar] [CrossRef]
- Derbinski, J.; Schulte, A.; Kyewski, B.; Klein, L. Promiscuous gene expression in medullary thymic epithelial cells mirrors the peripheral self. Nat. Immunol. 2001, 2, 1032–1039. [Google Scholar] [CrossRef] [PubMed]
- Scripture-Adams, D.D.; Damle, S.S.; Li, L.; Elihu, K.J.; Qin, S.; Arias, A.M.; Butler, R.R.; Champhekar, A.; Zhang, J.A.; Rothenberg, E.V. GATA-3 dose-dependent checkpoints in early T cell commitment. J. Immunol. 2014, 193, 3470–3491. [Google Scholar] [CrossRef]
- Yui, M.A.; Feng, N.; Rothenberg, E.V. Fine-scale staging of T cell lineage commitment in adult mouse thymus. J. Immunol. 2010, 185, 284–293. [Google Scholar] [CrossRef]
- Hernández-Hoyos, G.; Anderson, M.K.; Wang, C.; Rothenberg, E.V.; Alberola-Ila, J.J.I. GATA-3 expression is controlled by TCR signals and regulates CD4/CD8 differentiation. Immunity 2003, 19, 83–94. [Google Scholar] [CrossRef]
- Dang, C.V.; O’Donnell, K.A.; Zeller, K.I.; Nguyen, T.; Osthus, R.C.; Li, F. The c-Myc target gene network. Proc. Semin. Cancer Biol. 2006, 16, 253–264. [Google Scholar] [CrossRef]
- Spencer, C.A.; Groudine, M. Control of c-myc regulation in normal and neoplastic cells. Adv. Cancer Res. 1991, 56, 1–48. [Google Scholar] [CrossRef]
- Knoepfler, P.S. Myc goes global: New tricks for an old oncogene. Cancer Res. 2007, 67, 5061–5063. [Google Scholar] [CrossRef]
- Herrick, D.; Ross, J.J.M. The half-life of c-myc mRNA in growing and serum-stimulated cells: Influence of the coding and 3′untranslated regions and role of ribosome translocation. Mol. Cell Biol. 1994, 14, 2119–2128. [Google Scholar] [CrossRef]
- Hann, S.R.; Eisenman, R.N. Proteins encoded by the human c-myc oncogene: Differential expression in neoplastic cells. Mol. Cell Biol. 1984, 4, 2486–2497. [Google Scholar] [CrossRef] [PubMed]
- Ciechanover, A.; DiGiuseppe, J.A.; Schwartz, A.L.; Brodeur, G.M. Degradation of MYCN oncoprotein by the ubiquitin system. Prog. Clin. Biol. Res. 1991, 366, 37–43. [Google Scholar] [PubMed]
- Miller, D.M.; Thomas, S.D.; Islam, A.; Muench, D.; Sedoris, K. c-Myc and cancer metabolism. Clin. Cancer Res. 2012, 18, 5546–5553. [Google Scholar] [CrossRef]
- Weng, A.P.; Millholland, J.M.; Yashiro-Ohtani, Y.; Arcangeli, M.L.; Lau, A.; Wai, C.; Del Bianco, C.; Rodriguez, C.G.; Sai, H.; Tobias, J.J.G.; et al. c-Myc is an important direct target of Notch1 in T-cell acute lymphoblastic leukemia/lymphoma. Genes Dev. 2006, 20, 2096–2109. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.M.; Calvo, J.A.; Draheim, K.M.; Cunningham, L.A.; Hermance, N.; Beverly, L.; Krishnamoorthy, V.; Bhasin, M.; Capobianco, A.J.; Kelliher, M.A. Notch1 contributes to mouse T-cell leukemia by directly inducing the expression of c-myc. Mol. Cell Biol. 2006, 26, 8022–8031. [Google Scholar] [CrossRef]
- Nie, L.; Xu, M.; Vladimirova, A.; Sun, X.H. Notch-induced E2A ubiquitination and degradation are controlled by MAP kinase activities. EMBO J. 2003, 22, 5780–5792. [Google Scholar] [CrossRef]
- Li, X.; Gounari, F.; Protopopov, A.; Khazaie, K.; von Boehmer, H. Oncogenesis of T-ALL and nonmalignant consequences of overexpressing intracellular NOTCH1. J. Exp. Med. 2008, 205, 2851–2861. [Google Scholar] [CrossRef]
- Satoh, Y.; Matsumura, I.; Tanaka, H.; Ezoe, S.; Sugahara, H.; Mizuki, M.; Shibayama, H.; Ishiko, E.; Ishiko, J.; Nakajima, K.; et al. Roles for c-Myc in self-renewal of hematopoietic stem cells. J. Biol. Chem. 2004, 279, 24986–24993. [Google Scholar] [CrossRef]
- Girard, L.; Hanna, Z.; Beaulieu, N.; Hoemann, C.D.; Simard, C.; Kozak, C.A.; Jolicoeur, P. Frequent provirus insertional mutagenesis of Notch1 in thymomas of MMTVD/myc transgenic mice suggests a collaboration of c-myc and Notch1 for oncogenesis. Genes Dev. 1996, 10, 1930–1944. [Google Scholar] [CrossRef]
- Fang, T.C.; Yashiro-Ohtani, Y.; Del Bianco, C.; Knoblock, D.M.; Blacklow, S.C.; Pear, W.S. Notch directly regulates Gata3 expression during T helper 2 cell differentiation. Immunity 2007, 27, 100–110. [Google Scholar] [CrossRef]
- Hozumi, K.; Negishi, N.; Tsuchiya, I.; Abe, N.; Hirano, K.i.; Suzuki, D.; Yamamoto, M.; Engel, J.D.; Habu, S. Notch signaling is necessary for GATA3 function in the initiation of T cell development. Eur. J. Immunol. 2008, 38, 977–985. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Martin, M.; Ferrando, A. The NOTCH1-MYC highway toward T-cell acute lymphoblastic leukemia. Blood 2017, 129, 1124–1133. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.M.; Draheim, K.M.; Kelliher, M.A. The Notch1/c-Myc pathway in T cell leukemia. Cell Cycle 2007, 6, 927–930. [Google Scholar] [CrossRef] [PubMed]
- Guruharsha, K.; Kankel, M.W.; Artavanis-Tsakonas, S. The Notch signalling system: Recent insights into the complexity of a conserved pathway. Nat. Rev. Genet. 2012, 13, 654–666. [Google Scholar] [CrossRef]
- Andersson, E.R.; Lendahl, U. Therapeutic modulation of Notch signalling—are we there yet? Nat. Rev. Drug Discov. 2014, 13, 357–378. [Google Scholar] [CrossRef]
- Choi, E.-J.; Hong, J.Y.; Yoon, D.H.; Kang, J.; Park, C.-S.; Huh, J.; Chae, E.J.; Lee, Y.; Ryu, J.-S.; Suh, C. Treatment outcomes of dose-attenuated CHOP chemotherapy in elderly patients with peripheral T cell lymphoma. Blood Res. 2017, 52, 270–275. [Google Scholar] [CrossRef][Green Version]
- Chiarle, R.; Podda, A.; Prolla, G.; Gong, J.; Thorbecke, G.J.; Inghirami, G.J.C.i. CD30 in normal and neoplastic cells. Clin. Immunol. 1999, 90, 157–164. [Google Scholar] [CrossRef]
- Werner, B.; Massone, C.; Kerl, H.; Cerroni, L. Large CD30-positive cells in benign, atypical lymphoid infiltrates of the skin. J. Cutan. Pathol. 2008, 35, 1100–1107. [Google Scholar] [CrossRef]
- Falini, B.; Flenghi, L.; Aversa, F.; Barbabietola, G.; Martelli, M.; Comeli, P.; Tazzari, P.; Broe, M.; Stein, H.; Dürkop, H. Response of refractory Hodgkin’s disease to monoclonal anti-CD30 immunotoxin. Lancet 1992, 339, 1195–1196. [Google Scholar] [CrossRef]
- Tazzari, P.L.; De Totero, D.; Bolognesi, A.; Testoni, N.; Pileri, S.; Roncella, S.; Reato, G.; Stein, H.; Gobbi, M.; Stirpe, F.J.H. An Epstein-Barr virus-infected lymphoblastoid cell line (D430B) that grows in SCID-mice with the morphologic features of a CD30+ anaplastic large cell lymphoma, and is sensitive to anti-CD30 immunotoxins. Haematologica 1999, 84, 988–995. [Google Scholar]
- Doronina, S.O.; Toki, B.E.; Torgov, M.Y.; Mendelsohn, B.A.; Cerveny, C.G.; Chace, D.F.; DeBlanc, R.L.; Gearing, R.P.; Bovee, T.D.; Siegall, C.B.J.N.b. Development of potent monoclonal antibody auristatin conjugates for cancer therapy. Nat. Biotechnol. 2003, 21, 778–784. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Chen, D.; Lu, Y.; Dong, S.-C.; Ma, R.; Tang, W.-y.; Wu, J.-q.; Feng, J.-F.; Wu, J.-Z. A new immunotherapy strategy targeted CD30 in peripheral T-cell lymphomas: CAR-modified T-cell therapy based on CD30 mAb. Cancer Gene Ther. 2022, 29, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Mack, M.; Riethmüller, G.; Kufer, P. A small bispecific antibody construct expressed as a functional single-chain molecule with high tumor cell cytotoxicity. Proc. Natl. Acad. Sci. USA 1995, 92, 7021–7025. [Google Scholar] [CrossRef] [PubMed]
- Solimando, A.G.; Ribatti, D.; Vacca, A.; Einsele, H. Targeting B-cell non Hodgkin lymphoma: New and old tricks. Leuk. Res. 2016, 42, 93–104. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Khreisat, M.J.; Hussain, F.A.; Abdelfattah, A.M.; Almotiri, A.; Al-Sanabra, O.M.; Johan, M.F. The Role of NOTCH1, GATA3, and c-MYC in T Cell Non-Hodgkin Lymphomas. Cancers 2022, 14, 2799. https://doi.org/10.3390/cancers14112799
Al-Khreisat MJ, Hussain FA, Abdelfattah AM, Almotiri A, Al-Sanabra OM, Johan MF. The Role of NOTCH1, GATA3, and c-MYC in T Cell Non-Hodgkin Lymphomas. Cancers. 2022; 14(11):2799. https://doi.org/10.3390/cancers14112799
Chicago/Turabian StyleAl-Khreisat, Mutaz Jamal, Faezahtul Arbaeyah Hussain, Ali Mahmoud Abdelfattah, Alhomidi Almotiri, Ola Mohammed Al-Sanabra, and Muhammad Farid Johan. 2022. "The Role of NOTCH1, GATA3, and c-MYC in T Cell Non-Hodgkin Lymphomas" Cancers 14, no. 11: 2799. https://doi.org/10.3390/cancers14112799
APA StyleAl-Khreisat, M. J., Hussain, F. A., Abdelfattah, A. M., Almotiri, A., Al-Sanabra, O. M., & Johan, M. F. (2022). The Role of NOTCH1, GATA3, and c-MYC in T Cell Non-Hodgkin Lymphomas. Cancers, 14(11), 2799. https://doi.org/10.3390/cancers14112799