
Ligustilide Inhibits Tumor Angiogenesis by Downregulating VEGFA Secretion from Cancer-Associated Fibroblasts in Prostate Cancer via TLR4


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Results
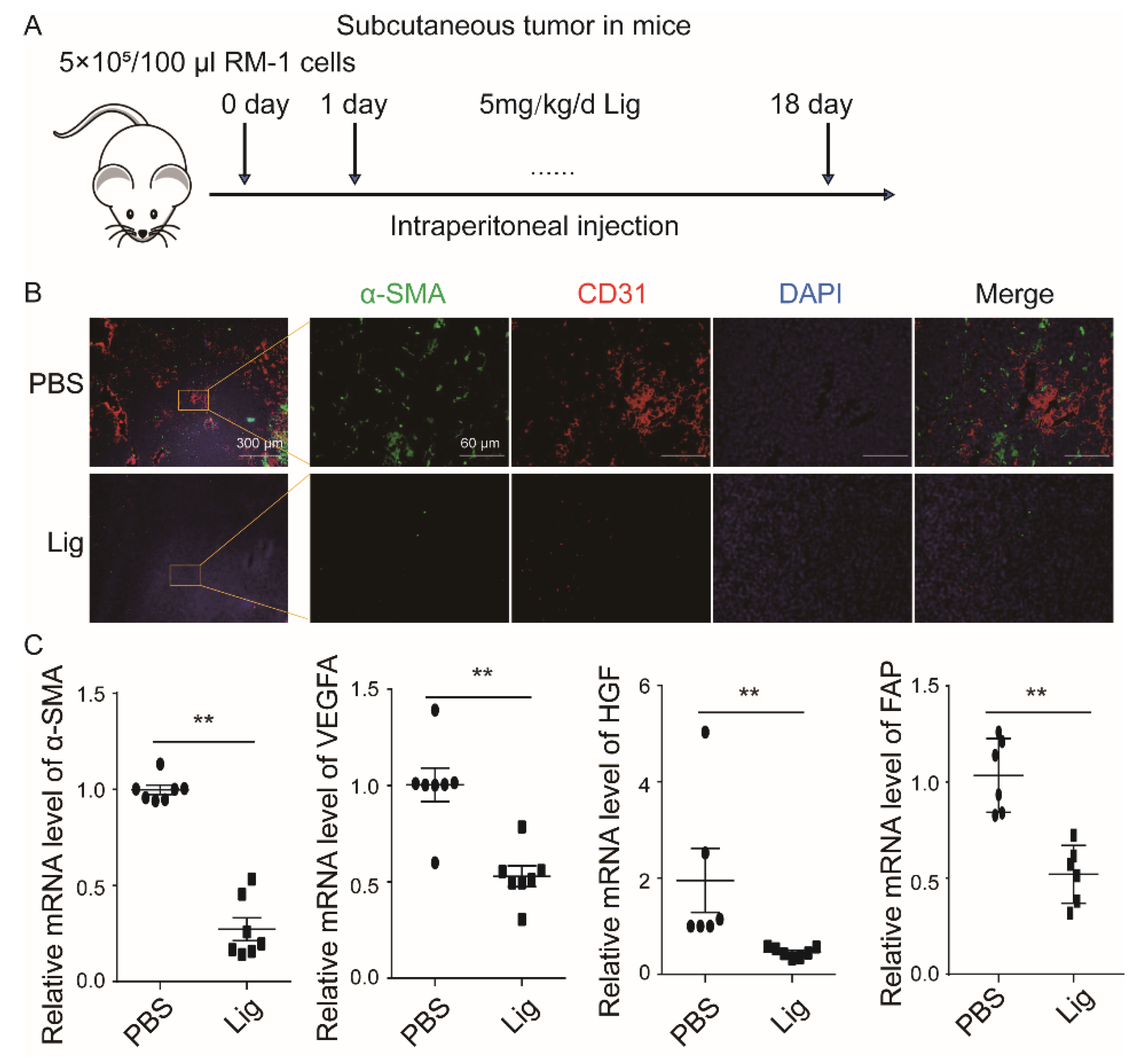
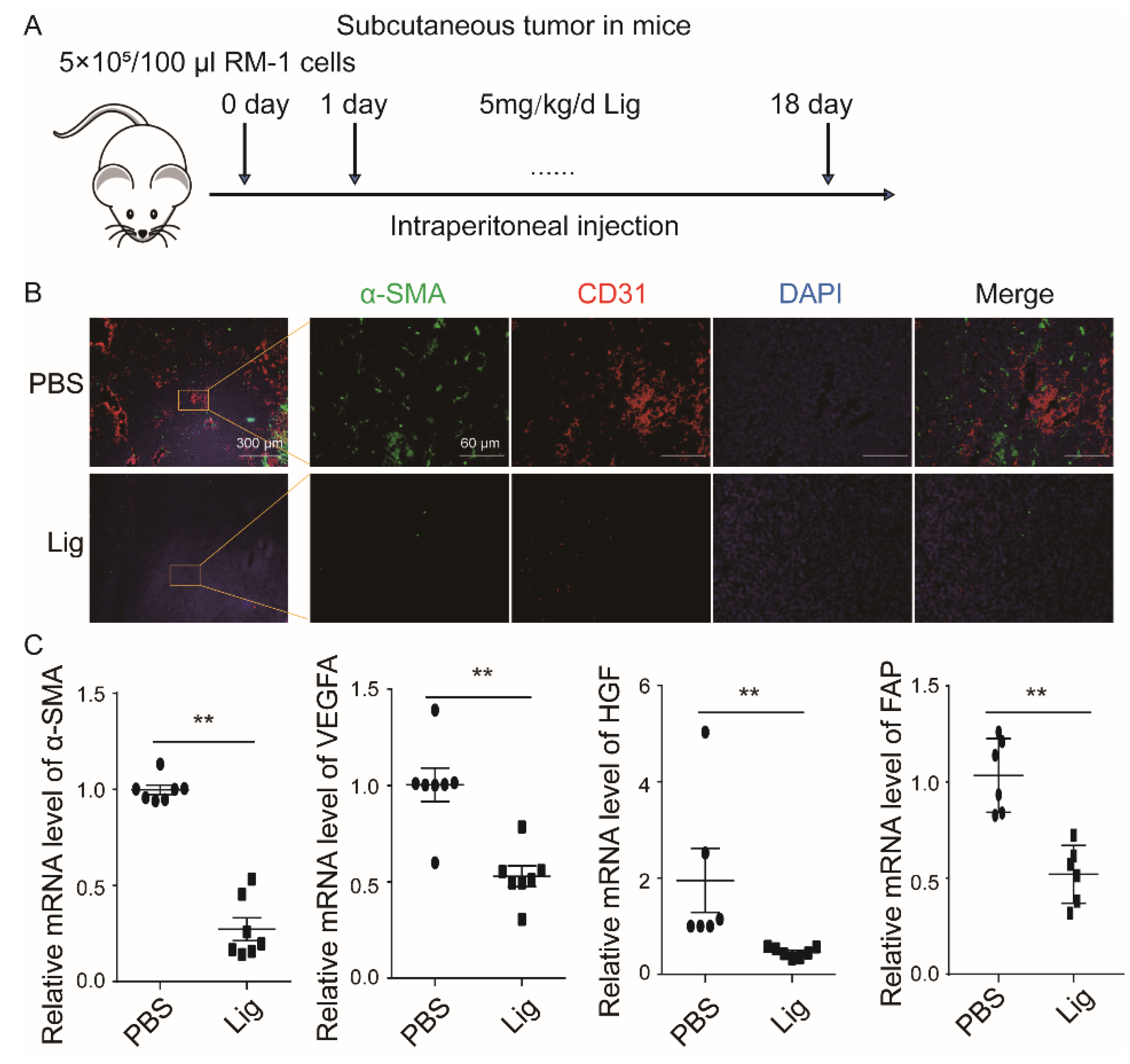
2.1. Ligustilide Reduces Blood Vessel Density in Prostate Cancer Tissue
2.2. Ligustilide Significantly Inhibits the Pro-Angiogenesis Effect of CAF Supernatant
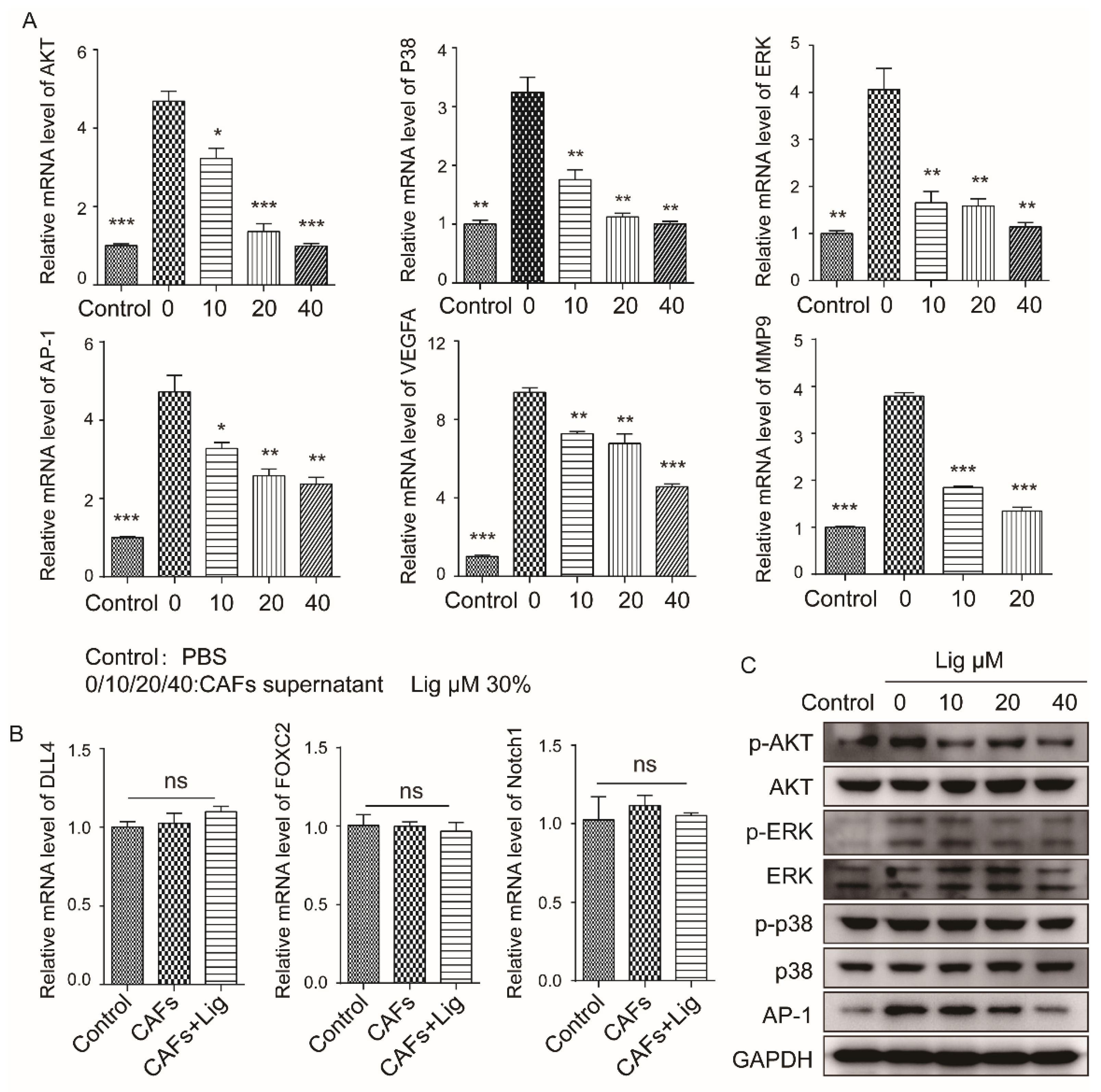
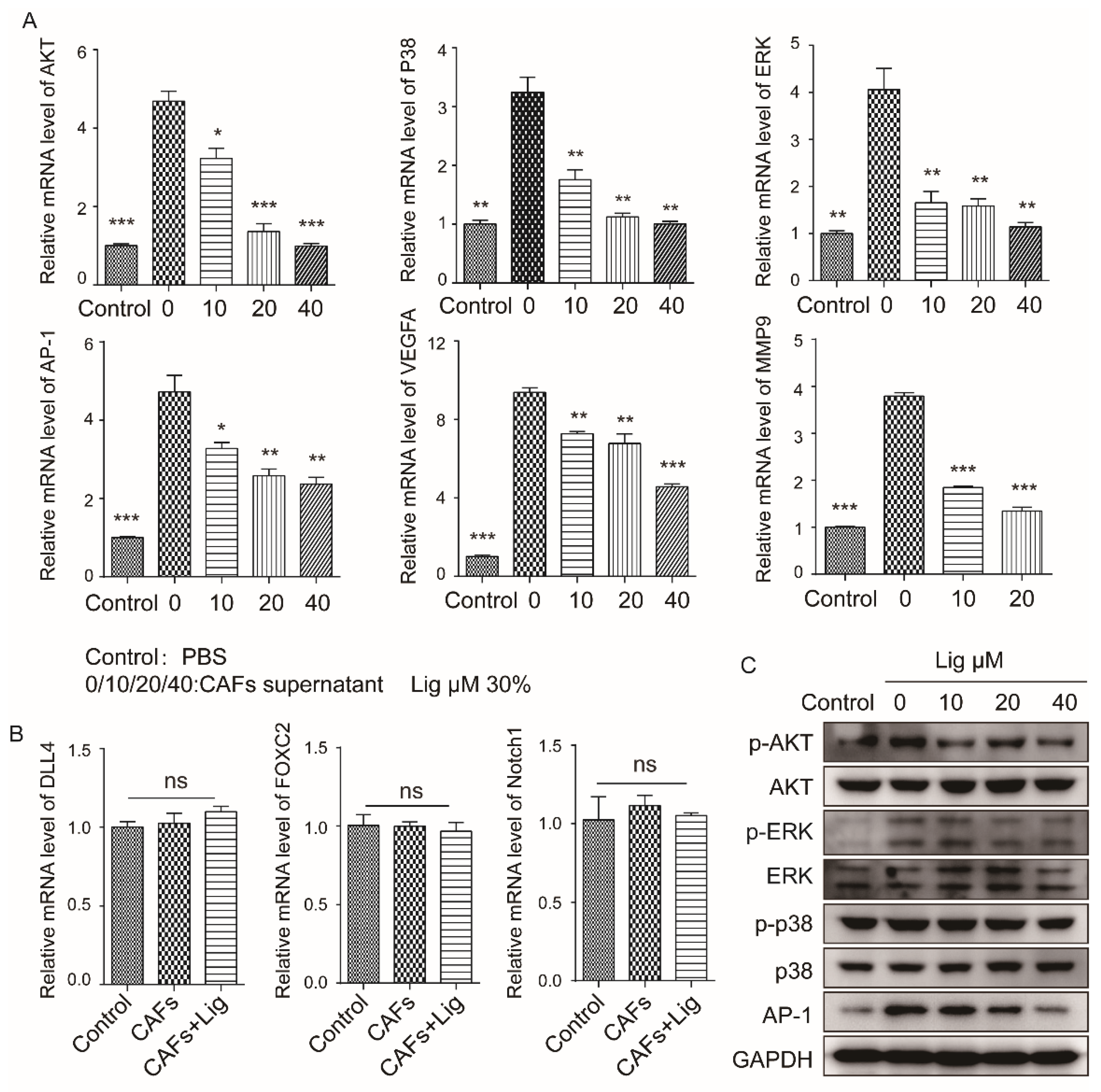
2.3. Ligustilide Inhibits the Pro-Angiogenic Effect of CAFs via the TLR4-AP-1 Signaling Pathway
2.4. Ligustilide DownRegulates the Expression Level of VEGFA in CAFs via the TLR4-AP-1 Signaling Pathway
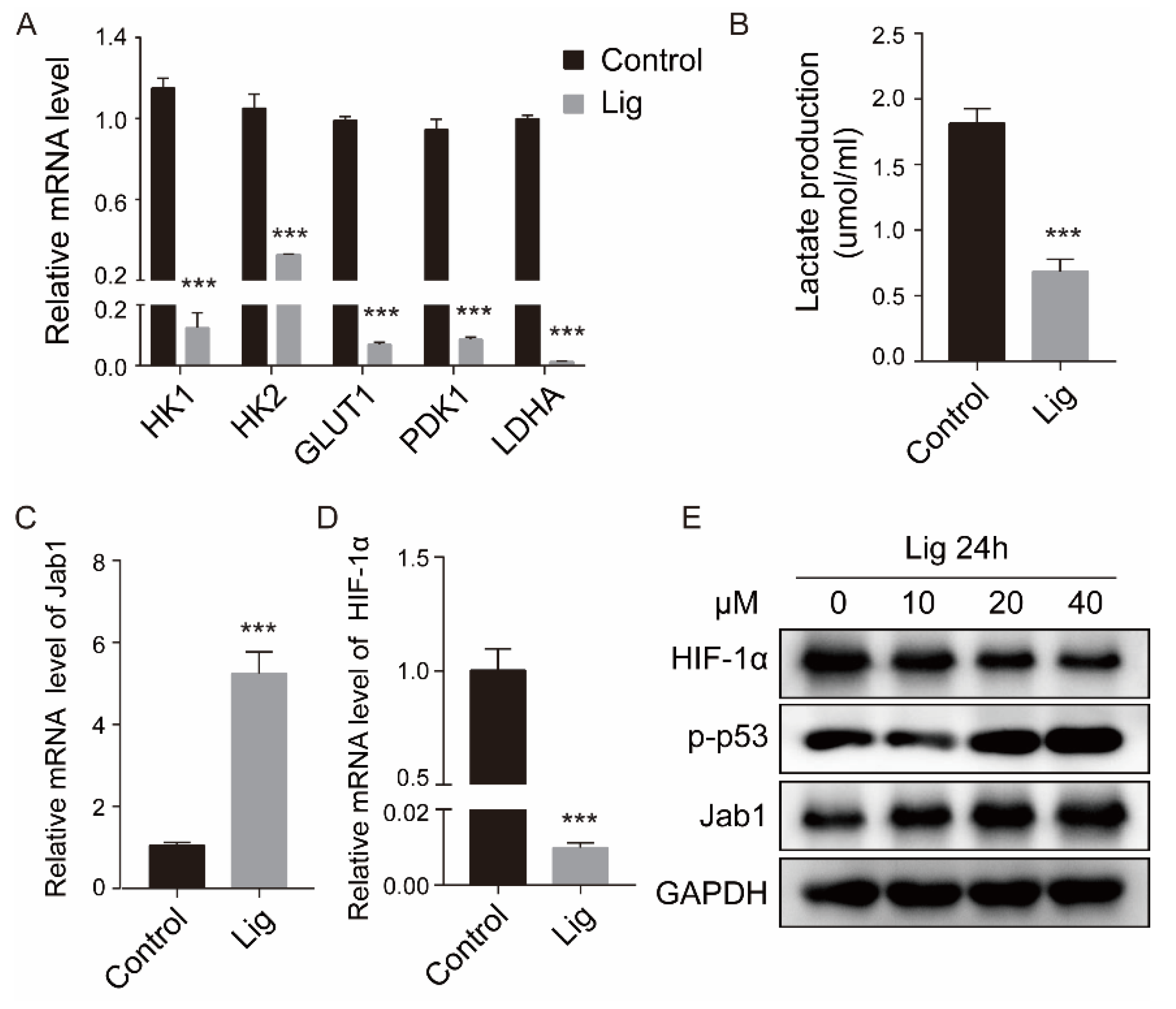
2.5. Ligustilide Inhibits Glycolysis and HIF-1 Expression in CAFs
2.6. Ligustilide Attenuates the Signaling Pathway Involved in the Proliferation of HUVECs Induced by CAF Supernatant
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Preparation of Ligustilide Solution and CAF Supernatant
4.3. Cell Viability Assay (MTT)
4.4. Real-Time Quantitative Polymerase Chain Reaction (RT-qPCR)
4.5. Western Blot Assay
4.6. ELISA
4.7. Cell Migration Assays
4.8. Tube Formation Assay
4.9. Immunohistochemical Staining (IHC)
4.10. Immunofluorescence
4.11. Lactate Detection
4.12. Mouse Tumor Models
4.13. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carmeliet, P.; Jain, R.K. Molecular mechanisms and clinical applications of angiogenesis. Nature 2011, 473, 298–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Folkman, J. Tumor angiogenesis: Therapeutic implications. N. Engl. J. Med. 1971, 285, 1182–1186. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, N. Vascular endothelial growth factor. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 789–791. [Google Scholar] [CrossRef] [Green Version]
- Riabov, V.; Gudima, A.; Wang, N.; Mickley, A.; Orekhov, A.; Kzhyshkowska, J. Role of tumor associated macrophages in tumor angiogenesis and lymphangiogenesis. Front. Physiol. 2014, 5, 75. [Google Scholar] [CrossRef] [Green Version]
- Ferrara, N. Vascular endothelial growth factor: Basic science and clinical progress. Endocr. Rev. 2004, 25, 581–611. [Google Scholar] [CrossRef]
- Hurwitz, H.; Fehrenbacher, L.; Novotny, W.; Cartwright, T.; Hainsworth, J.; Heim, W.; Berlin, J.; Baron, A.; Griffing, S.; Holmgren, E.; et al. Bevacizumab plus irinotecan, fluorouracil, and leucovorin for metastatic colorectal cancer. N. Engl. J. Med. 2004, 350, 2335–2342. [Google Scholar] [CrossRef] [Green Version]
- Ferrara, N.; Adamis, A.P. Ten years of anti-vascular endothelial growth factor therapy. Nat. Rev. Drug Discov. 2016, 15, 385–403. [Google Scholar] [CrossRef] [Green Version]
- Ferrara, N.; Mass, R.D.; Campa, C.; Kim, R. Targeting VEGF-A to treat cancer and age-related macular degeneration. Annu. Rev. Med. 2007, 58, 491–504. [Google Scholar] [CrossRef]
- Ferrara, N.; Kerbel, R.S. Angiogenesis as a therapeutic target. Nature 2005, 438, 967–974. [Google Scholar] [CrossRef]
- Bennouna, J.; Sastre, J.; Arnold, D.; Österlund, P.; Greil, R.; Van Cutsem, E.; von Moos, R.; Viéitez, J.M.; Bouché, O.; Borg, C.; et al. Continuation of bevacizumab after first progression in metastatic colorectal cancer (ML18147): A randomised phase 3 trial. Lancet Oncol. 2013, 14, 29–37. [Google Scholar] [CrossRef]
- Chung, A.S.; Lee, J.; Ferrara, N. Targeting the tumour vasculature: Insights from physiological angiogenesis. Nat. Rev. Cancer 2010, 10, 505–514. [Google Scholar] [CrossRef] [PubMed]
- De Palma, M.; Biziato, D.; Petrova, T.V. Microenvironmental regulation of tumour angiogenesis. Nat. Rev. Cancer 2017, 17, 457–474. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Yang, W.; Qin, C.; Zhang, L.; Deng, J.; Liu, S.; Qin, Z. Responsiveness of stromal fibroblasts to IFN-γ blocks tumor growth via angiostasis. J. Immunol. 2009, 183, 6413–6421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mojic, M.; Takeda, K.; Hayakawa, Y. The Dark Side of IFN-γ: Its Role in Promoting Cancer Immunoevasion. Int. J. Mol. Sci. 2017, 19, 89. [Google Scholar] [CrossRef] [Green Version]
- Kaul, P.N.; Mallavarapu, G.R.; Chamoli, R.P. The essential oil composition of Angelica glauca roots. Planta Med. 1996, 62, 80–81. [Google Scholar] [CrossRef]
- Jiang, X.; Zhao, W.; Zhu, F.; Wu, H.; Ding, X.; Bai, J.; Zhang, X.; Qian, M. Ligustilide inhibits the proliferation of non-small cell lung cancer via glycolytic metabolism. Toxicol. Appl. Pharm. 2021, 410, 115336. [Google Scholar] [CrossRef]
- Zhang, B.; Wu, D.; Hu, L.; Cha, X.; Liu, Y.; Li, J.; Xie, B.; Li, B.; Zheng, L. Ligustilide inhibits the proliferation of human osteoblastoma MG63 cells through the TLR4-ERK pathway. Life Sci. 2022, 288, 118993. [Google Scholar] [CrossRef]
- Lang, F.; Qu, J.; Yin, H.; Li, L.; Zhi, Y.; Liu, Y.; Fang, Z.; Hao, E. Apoptotic cell death induced by Z-Ligustilidein human ovarian cancer cells and role of NRF2. Food Chem. Toxicol. 2018, 121, 631–638. [Google Scholar] [CrossRef]
- Yin, J.; Wang, C.; Mody, A.; Bao, L.; Hung, S.-H.; Svoronos, S.A.; Tseng, Y. The Effect of Z-Ligustilide on the Mobility of Human Glioblastoma T98G Cells. PLoS ONE 2013, 8, e66598. [Google Scholar] [CrossRef]
- Ma, J.; Mei, J.; Lu, J.; Wang, Y.; Hu, M.; Ma, F.; Long, H.; Qin, Z.; Tao, N. Ligustilide promotes apoptosis of cancer-associated fibroblasts via the TLR4 pathways. Food Chem. Toxicol. 2020, 135, 110991. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Xu, Y.; Zheng, Q.; Wang, Y.; Hu, M.; Ma, F.; Long, H.; Qin, Z.; Tao, N. Ligustilide inhibits the activation of cancer-associated fibroblasts. Life Sci. 2019, 218, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Veglia, F.; Perego, M.; Gabrilovich, D. Myeloid-derived suppressor cells coming of age. Nat. Immunol. 2018, 19, 108–119. [Google Scholar] [CrossRef] [PubMed]
- Takeya, M.; Komohara, Y. Role of tumor-associated macrophages in human malignancies: Friend or foe? Pathol. Int. 2016, 66, 491–505. [Google Scholar] [CrossRef] [Green Version]
- Facciabene, A.; Peng, X.; Hagemann, I.S.; Balint, K.; Barchetti, A.; Wang, L.-P.; Gimotty, P.A.; Gilks, C.B.; Lal, P.; Zhang, L.; et al. Tumour hypoxia promotes tolerance and angiogenesis via CCL28 and T(reg) cells. Nature 2011, 475, 226–230. [Google Scholar] [CrossRef]
- Sarvaria, A.; Madrigal, J.A.; Saudemont, A. B cell regulation in cancer and anti-tumor immunity. Cell. Mol. Immunol. 2017, 14, 662–674. [Google Scholar] [CrossRef] [Green Version]
- Masucci, M.T.; Minopoli, M.; Carriero, M.V. Tumor Associated Neutrophils. Their Role in Tumorigenesis, Metastasis, Prognosis and Therapy. Front. Oncol. 2019, 9, 1146. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Song, E. Turning foes to friends: Targeting cancer-associated fibroblasts. Nat. Rev. Drug Discov. 2019, 18, 99–115. [Google Scholar] [CrossRef]
- Calon, A.; Tauriello, D.V.; Batlle, E. TGF-beta in CAF-mediated tumor growth and metastasis. Semin. Cancer Biol. 2014, 25, 15–22. [Google Scholar] [CrossRef]
- Fukumura, D.; Jain, R.K. Tumor microvasculature and microenvironment: Targets for anti-angiogenesis and normalization. Microvasc. Res. 2007, 74, 72–84. [Google Scholar] [CrossRef] [Green Version]
- Ozawa, C.R.; Banfi, A.; Glazer, N.L.; Thurston, G.; Springer, M.L.; Kraft, P.E.; McDonald, D.M.; Blau, H.M. Microenvironmental VEGF concentration, not total dose, determines a threshold between normal and aberrant angiogenesis. J. Clin. Investig. 2004, 113, 516–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schreiber, H.; Rowley, D.A. Awakening immunity. Science 2010, 330, 761–762. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, N. VEGF and Intraocular Neovascularization: From Discovery to Therapy. Transl. Vis. Sci. Technol. 2016, 5, 10. [Google Scholar] [CrossRef] [Green Version]
- Park, B.S.; Song, D.H.; Kim, H.M.; Choi, B.S.; Lee, H.; Lee, J.O. The structural basis of lipopolysaccharide recognition by the TLR4-MD-2 complex. Nature 2009, 458, 1191–1195. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zeng, Y.; Zhu, L.; Wan, J.; Lei, N.; Yao, X.; Duan, X.; Zhang, Y.; Cheng, Y.; Tao, N.; et al. Polysaccharides from Lentinus Edodes Inhibits Lymphangiogenesis via the Toll-Like Receptor 4/JNK Pathway of Cancer-Associated Fibroblasts. Front. Oncol. 2020, 10, 547683. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Ma, J.; Zheng, Q.; Wang, Y.; Hu, M.; Ma, F.; Qin, Z.; Lei, N.; Tao, N. MPSSS impairs the immunosuppressive function of cancer-associated fibroblasts via the TLR4-NF-kappaB pathway. Biosci. Rep. 2019, 39, BSR20182171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mei, J.; Ma, J.; Xu, Y.; Wang, Y.; Hu, M.; Ma, F.; Qin, Z.; Xue, R.; Tao, N. Cinnamaldehyde Treatment of Prostate Cancer-Associated Fibroblasts Prevents Their Inhibitory Effect on T Cells through Toll-Like Receptor 4. Drug Des. Dev. Ther. 2020, 14, 3363–3372. [Google Scholar] [CrossRef]
- Zhang, W.; He, W.; Shi, X.; Li, X.; Wang, Y.; Hu, M.; Ma, F.; Tao, N.; Wang, G.; Qin, Z. An Asparagus polysaccharide fraction inhibits MDSCs by inducing apoptosis through toll-like receptor 4. Phytother. Res. 2018, 32, 1297–1303. [Google Scholar] [CrossRef]
- Ding, Y.; Yang, Z.; Zhang, W.; Xu, Y.; Wang, Y.; Hu, M.; Ma, F.; Long, H.; Tao, N.; Qin, Z. Eugenol triggers CD11b+Gr1+ myeloid-derived suppressor cell apoptosis via endogenous apoptosis pathway. RSC Adv. 2018, 8, 3833–3838. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Y.; Du, Q.; Zhang, Z.; Ma, J.; Han, L.; Wang, Y.; Yang, L.; Tao, N.; Qin, Z. Curcumin promotes cancer-associated fibroblasts apoptosis via ROS-mediated endoplasmic reticulum stress. Arch. Biochem. Biophys. 2020, 694, 108613. [Google Scholar] [CrossRef]
- Lin, S.C.; Lo, Y.C.; Wu, H. Helical assembly in the MyD88-IRAK4-IRAK2 complex in TLR/IL-1R signalling. Nature 2010, 465, 885–890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen recognition and innate immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, M.; Beane, T.J.; Quillien, A.; Male, I.; Zhu, L.H.J.; Lawson, N.D. Vegfa signals through ERK to promote angiogenesis, but not artery differentiation. Development 2016, 143, 3796–3805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bae, M.-K.; Ahn, M.-Y.; Jeong, J.-W.; Bae, M.-H.; Lee, Y.M.; Bae, S.-K.; Park, J.-W.; Kim, K.-R.; Kim, K.-W. Jab1 interacts directly with HIF-1 alpha and regulates its stability. J. Biol. Chem. 2002, 277, 9–12. [Google Scholar] [CrossRef] [Green Version]
- Kleemann, R.; Hausser, A.; Geiger, G.; Mischke, R.; Burger-Kentischer, A.; Flieger, O.; Johannes, F.-J.; Roger, T.; Calandra, T.; Kapurniotu, A.; et al. Intracellular action of the cytokine MIF to modulate AP-1 activity and the cell cycle through Jab1. Nature 2000, 408, 211–216. [Google Scholar] [CrossRef]
- Wang, H.; Sun, X.; Luo, Y.; Lin, Z.; Wu, J. Adapter protein NRBP associates with Jab1 and negatively regulates AP-1 activity. FEBS Lett. 2006, 580, 6015–6021. [Google Scholar] [CrossRef] [Green Version]
- Gerald, D.; Berra, E.; Frapart, Y.M.; Chan, D.A.; Giaccia, A.J.; Mansuy, D.; Pouysségur, J.; Yaniv, M.; Mechta-Grigoriou, F. JunD reduces tumor angiogenesis by protecting cells from oxidative stress. Cell 2004, 118, 781–794. [Google Scholar] [CrossRef]
- Tang, N.; Wang, L.; Esko, J.; Giordano, F.J.; Huang, Y.; Gerber, H.-P.; Ferrara, N.; Johnson, R.S.; Tang, N.; Wang, L.; et al. Loss of HIF-1 alpha in endothelial cells disrupts a hypoxia-driven VEGF autocrine loop necessary for tumorigenesis. Cancer Cell 2004, 6, 485–495. [Google Scholar] [CrossRef] [Green Version]
- Rong, L.; Bian, Y.; Liu, S.; Liu, X.; Li, X.; Liu, H.; Zhou, J.; Peng, J.; Zhang, H.; Chen, H.; et al. Identifying tumor promoting genomic alterations in tumor-associated fibroblasts via retrovirus-insertional mutagenesis. Oncotarget. 2017, 8, 97231–97245. [Google Scholar] [CrossRef] [Green Version]
- Apolloni, E.; Bronte, V.; Mazzoni, A.; Serafini, P.; Cabrelle, A.; Segal, D.M.; Young, H.A.; Zanovello, P. Immortalized myeloid suppressor cells trigger apoptosis in antigen-activated T lymphocytes. J. Immunol. 2000, 165, 6723–6730. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, J.; Chen, X.; Chen, Y.; Tao, N.; Qin, Z. Ligustilide Inhibits Tumor Angiogenesis by Downregulating VEGFA Secretion from Cancer-Associated Fibroblasts in Prostate Cancer via TLR4. Cancers 2022, 14, 2406. https://doi.org/10.3390/cancers14102406
Ma J, Chen X, Chen Y, Tao N, Qin Z. Ligustilide Inhibits Tumor Angiogenesis by Downregulating VEGFA Secretion from Cancer-Associated Fibroblasts in Prostate Cancer via TLR4. Cancers. 2022; 14(10):2406. https://doi.org/10.3390/cancers14102406
Chicago/Turabian StyleMa, Jing, Xu Chen, Yumo Chen, Ning Tao, and Zhihai Qin. 2022. "Ligustilide Inhibits Tumor Angiogenesis by Downregulating VEGFA Secretion from Cancer-Associated Fibroblasts in Prostate Cancer via TLR4" Cancers 14, no. 10: 2406. https://doi.org/10.3390/cancers14102406
APA StyleMa, J., Chen, X., Chen, Y., Tao, N., & Qin, Z. (2022). Ligustilide Inhibits Tumor Angiogenesis by Downregulating VEGFA Secretion from Cancer-Associated Fibroblasts in Prostate Cancer via TLR4. Cancers, 14(10), 2406. https://doi.org/10.3390/cancers14102406