
Rheumatoid Factor: A Novel Determiner in Cancer History


Abstract
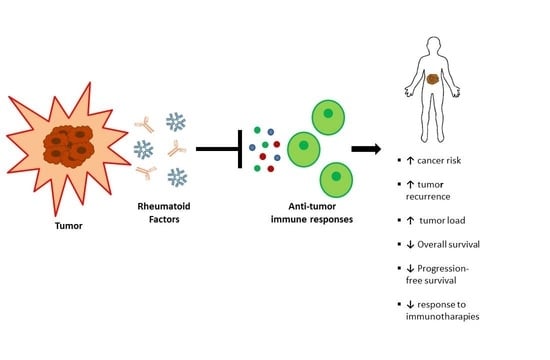
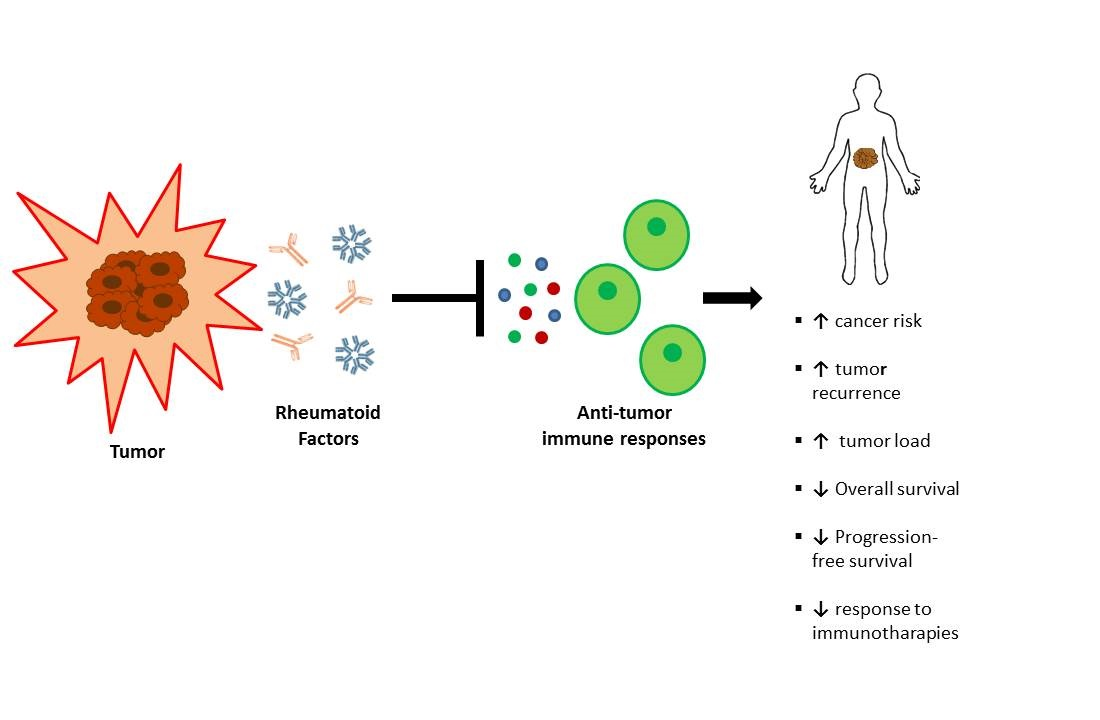
Simple Summary
Abstract
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Rheumatoid Factors Isotypes and Affinities
3. Physiological Rheumatoid Factor Production and Its Presence in Different Clinical Conditions
3.1. RFs in Patients with Non-Autoimmune Conditions
3.2. RFs in the General Healthy Population
3.3. RFs in Rheumatoid Arthritis and Other Autoimmune Diseases
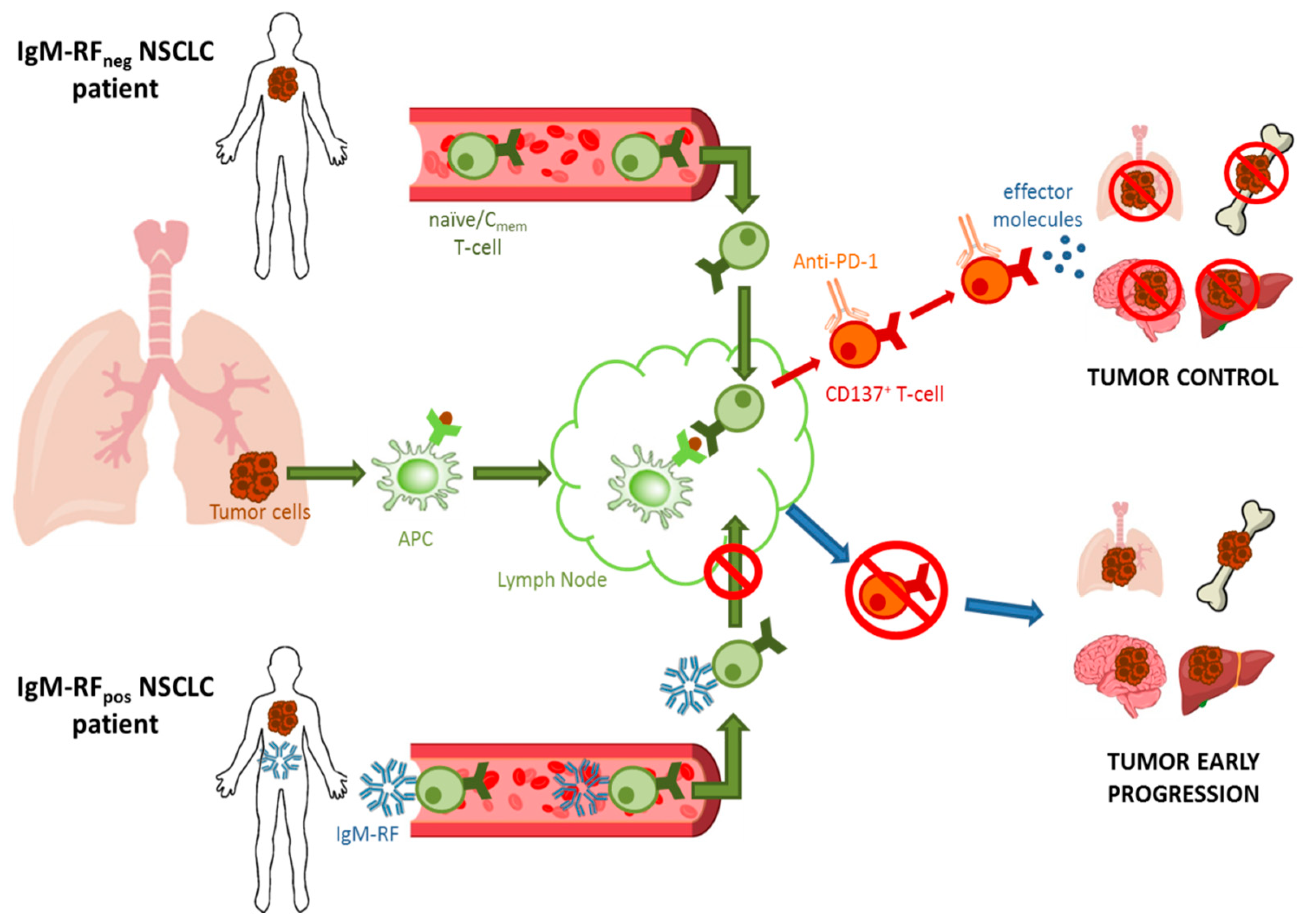
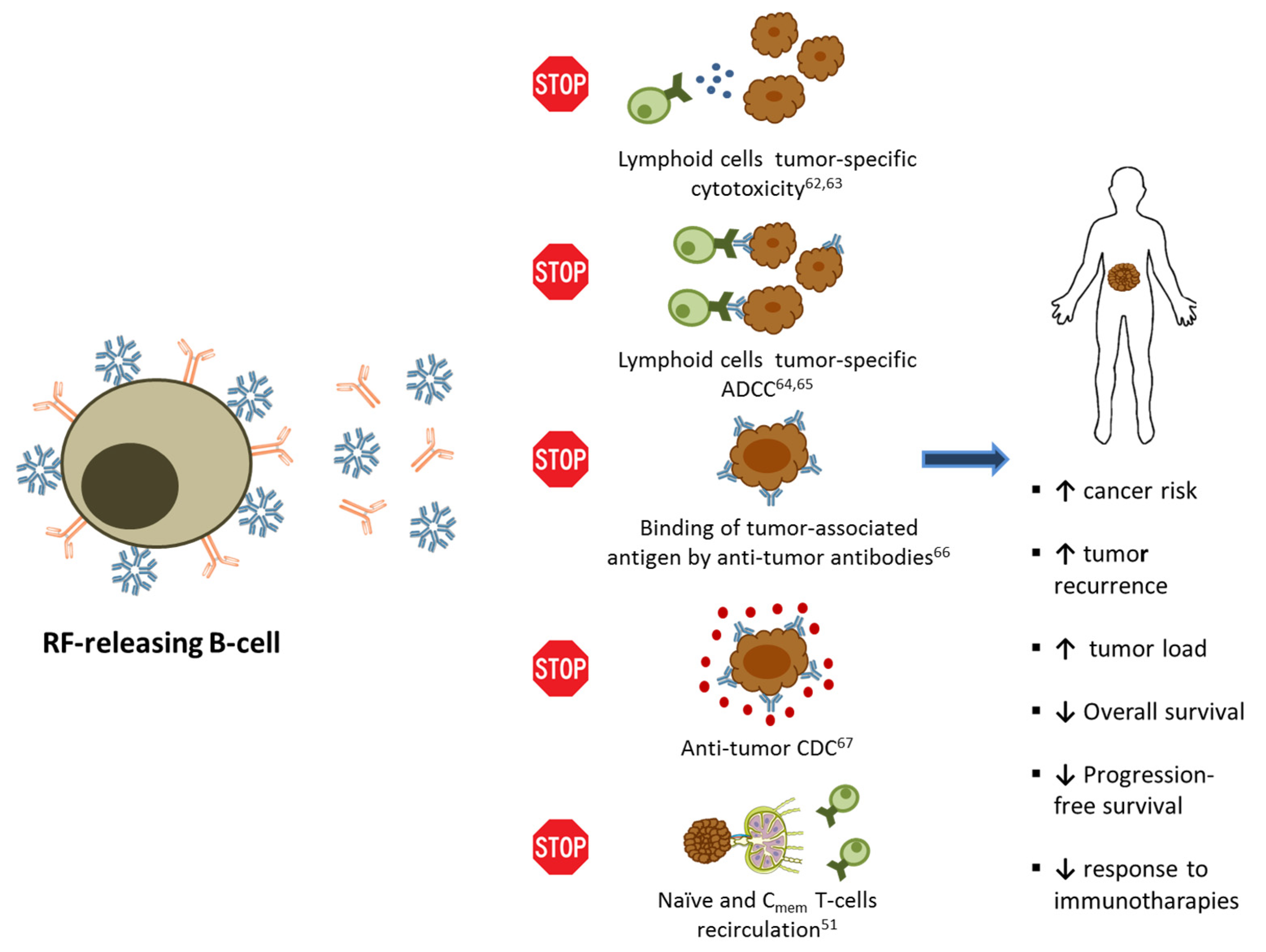
4. Rheumatoid Factor and Cancer History
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Van Zeben, D.; Hazes, J.M.W.; Zwinderman, A.H.; Cats, A.; Van Der Voort, E.A.M.; Breedveld, F.C.; Van Zeben, D.; Hazes, J.M.W.; Cats, A.; Van Der Voort, E.A.M.; et al. Clinical significance of rheumatoid factors in early rheumatoid arthritis: Results of a follow up study. Ann. Rheum. Dis. 1992, 51, 1029–1035. [Google Scholar] [CrossRef] [PubMed]
- Steiner, G.; Smolen, J.S. Autoantibodies in rheumatoid arthritis: Prevalence and clinical significance. In Rheumatoid Arthritis; Oxford University Press: Oxford, UK, 2006; pp. 193–198. ISBN 9780198566304. [Google Scholar]
- Dörner, T.; Egerer, K.; Feist, E.; Burmester, G.R. Rheumatoid factor revisited. Curr. Opin. Rheumatol. 2004, 16, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Ingegnoli, F.; Castelli, R.; Gualtierotti, R. Rheumatoid factors: Clinical applications. Dis. Markers 2013, 35, 727–734. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, J.H. Pathogenetic concepts and origins of rheumatoid factor in rheumatoid arthritis. Arthritis Rheum. 1993, 36, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Carson, D.A.; Chen, P.P.; Fox, R.I.; Kipps, T.J.; Jirik, F.; Goldfien, R.D.; Silverman, G.; Radoux, V.; Fong, S. Rheumatoid factor and immune networks. Annu. Rev. Immunol. 1987, 5, 109–126. [Google Scholar] [CrossRef]
- Carson, D.A.; Chen, P.P.; Kipps, T.J. New roles for rheumatoid factor. J. Clin. Investig. 1991, 87, 379–383. [Google Scholar] [CrossRef]
- Bos, W.H.; van de Stadt, L.A.; Sohrabian, A.; Rönnelid, J.; Van Schaardenburg, D. Development of anti-citrullinated protein antibody and rheumatoid factor isotypes prior to the onset of rheumatoid arthritis. Arthritis Res. Ther. 2014, 16, 1–2. [Google Scholar] [CrossRef][Green Version]
- Dresser, D.W.; Popham, A.M. Induction of an IgM anti-(bovine)-IgG response in mice by bacterial lipopolysaccharide. Nature 1976, 264, 552–554. [Google Scholar] [CrossRef]
- Falkenburg, W.J.J.; Van Schaardenburg, D.; Ooijevaar-De Heer, P.; Wolbink, G.; Rispens, T. IgG subclass specificity discriminates restricted IgM rheumatoid factor responses from more mature anti-citrullinated protein antibody-associated or isotype-switched IgA responses. Arthritis Rheumatol. 2015, 67, 3124–3134. [Google Scholar] [CrossRef]
- Bonagura, V.R.; Artandi, S.E.; Agostino, N.; Tao, M.H.; Morrison, S.L. Mapping rheumatoid factor binding sites using genetically engineered, chimeric IgG antibodies. DNA Cell Biol. 1992, 11, 245–252. [Google Scholar] [CrossRef]
- Hirohata, S.; Inoue, T.; Ito, K. Frequency analysis of human peripheral blood B cells producing autoantibodies: Differential activation requirements of precursors for B cells producing IgM-RF and anti-DNA antibody. Cell. Immunol. 1991, 138, 445–455. [Google Scholar] [CrossRef]
- He, X.; Goronzy, J.J.; Weyand, C.M. The repertoire of rheumatoid factor-producing B cells in normal subjects and patients with rheumatoid arthritis. Arthritis Rheum. 1993, 36, 1061–1069. [Google Scholar] [CrossRef] [PubMed]
- Carson, D.A.; Chen, P.P.; Kipps, T.J.; Roudier, J.; Silverman, G.J.; Tighe, H. Regulation of rheumatoid factor synthesis. Clin. Exp. Rheumatol. 1989, 7, S69–S73. [Google Scholar] [PubMed]
- Van Esch, W.J.; Reparon-Schuijt, C.C.; Levarht, E.W.; Van Kooten, C.; Breedveld, F.C.; Verweij, C.L. Differential requirements for induction of total immunoglobulin and physiological rheumatoid factor production by human peripheral blood B cells. Clin. Exp. Immunol. 2001, 123, 496–504. [Google Scholar] [CrossRef]
- Reparon-Schuijt, C.C.; Van Esch, W.J.E.; Van Kooten, C.; Levarht, E.W.N.; Breedveld, F.C.; Verweij, C.L. Functional analysis of rheumatoid factor-producing B cells from the synovial fluid of rheumatoid arthritis patients. Arthritis Rheum. 1998, 41, 2211–2220. [Google Scholar] [CrossRef]
- Reparon-Schuijt, C.C.; Van Esch, W.J.E.; Van Kooten, C.; Rozier, B.C.D.; Levarht, E.W.N.; Breedveld, F.C.; Verweij, C.L. Regulation of synovial B cell survival nrheumatoid arthritis by vascular cell adhesion molecule 1 (CD106) expressed on fibroblast-like synoviocytes. Arthritis Rheum. 2000, 43, 1115–1121. [Google Scholar] [CrossRef]
- Meyer, M.P.; Malan, A.F. Rheumatoid factor in congenital syphilis. Genitourin. Med. 1989, 65, 304–307. [Google Scholar] [CrossRef]
- Svec, K.H.; Dingle, J.H. The Occurence of Rheumatoid Factor in Association with Antibody Response to Influenza A2(Asian) Virus. Arthritis Rheum. 1965, 8, 524–529. [Google Scholar] [CrossRef]
- Carson, D.A.; Bayer, A.S.; Eisenberg, R.A.; Lawrance, S.; Theofilopoulos, A. IgG rheumatoid factor in subacute bacterial endocarditis: Relationship to IgM rheumatoid factor and circulating immune complexes. Clin. Exp. Immunol. 1978, 31, 100–103. [Google Scholar]
- Harboe, M. Rheumatoid factors in leprosy and parasitic diseases. Scand. J. Rheumatol. 1988, 17, 309–313. [Google Scholar] [CrossRef]
- Slaughter, L.; Carson, D.A.; Jensen, F.C.; Holbrook, T.L.; Vaughan, J.H. In vitro effects of Epstein-Barr virus on peripheral blood mononuclear cells from patients with rheumatoid arthritis and normal subjects. J. Exp. Med. 1978, 148, 1429–1434. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.C. Rheumatoid factors in subacute bacterial endocarditis and other infectious diseases. Scand. J. Rheumatol. Suppl. 1988, 75, 300–308. [Google Scholar] [CrossRef] [PubMed]
- Roosnek, E.; Lanzavecchia, A. Efficient and selective presentation of antigen-antibody complexes by rheumatoid factor B cells. J. Exp. Med. 1991, 173, 487–489. [Google Scholar] [CrossRef] [PubMed]
- Tighe, H.; Chen, P.P.; Tucker, R.; Kipps, T.J.; Roudier, J.; Jirik, F.R.; Carson, D.A. Function of B cells expressing a human immunoglobulin M rheumatoid factor autoantibody in transgenic mice. J. Exp. Med. 1993, 177, 109–118. [Google Scholar] [CrossRef]
- Newkirk, M.M. Rheumatoid factors: Host resistance or autoimmunity? Clin. Immunol. 2002, 104, 1–13. [Google Scholar] [CrossRef]
- Westwood, O.M.R.; Nelson, P.N.; Hay, F.C. Rheumatoid factors: What’s new? Rheumatology 2006, 45, 379–385. [Google Scholar] [CrossRef]
- Sinclair, N.R.S.; Panoskaltsis, A. Immunoregulation by Fc signals. A mechanism for self-nonself discrimination. Immunol. Today 1987, 8, 76–79. [Google Scholar] [CrossRef]
- Van Snick, J.L.; Van Roost, E.; Markowetz, B.; Cambiaso, C.L.; Masson, P.L. Enhancement by IgM rheumatoid factor of in vitro ingestion by macrophages and in vivo clearance of aggregated IgG or antigen-antibody complexes. Eur. J. Immunol. 1978, 8, 279–285. [Google Scholar] [CrossRef]
- Palazzi, C.; Buskila, D.; D’Angelo, S.; D’Amico, E.; Olivieri, I. Autoantibodies in patients with chronic hepatitis C virus infection: Pitfalls for the diagnosis of rheumatic diseases. Autoimmun. Rev. 2012, 11, 659–663. [Google Scholar] [CrossRef]
- Charles, E.D.; Orloff, M.I.M.; Nishiuchi, E.; Marukian, S.; Rice, C.M.; Dustin, L.B. Somatic hypermutations confer rheumatoid factor activity in hepatitis C virus-associated mixed cryoglobulinemia. Arthritis Rheum. 2013, 65, 2430–2440. [Google Scholar] [CrossRef]
- Børretzen, M.; Chapman, C.; Natvig, J.B.; Thompson, K.M. Differences in mutational patterns between rheumatoid factors in health and disease are related to variable heavy chain family and germ-line gene usage. Eur. J. Immunol. 1997, 27, 735–741. [Google Scholar] [CrossRef] [PubMed]
- Simard, J.F.; Holmqvist, M. Rheumatoid factor positivity in the general population. BMJ 2012, 345, e5841. [Google Scholar] [CrossRef] [PubMed]
- Tasliyurt, T.; Kisacik, B.; Kaya, S.U.; Yildirim, B.; Pehlivan, Y.; Kutluturk, F.; Ozyurt, H.; Sahin, S.; Onat, A.M. The frequency of antibodies against cyclic citrullinated peptides and rheumatoid factor in healthy population: A field study of rheumatoid arthritis from northern turkey. Rheumatol. Int. 2013, 33, 939–942. [Google Scholar] [CrossRef] [PubMed]
- Newkirk, M.M. Rheumatoid factors: What do they tell us? J. Rheumatol. 2002, 29, 2034–2040. [Google Scholar]
- Jacobsson, L.T.H.; Knowler, W.C.; Pillemer, S.; Hanson, R.L.; Pettitt, D.J.; Nelson, R.G.; Puente, A.; Del Mccance, D.R.; Charles, M.A.; Bennett, P.H. Rheumatoid arthritis and mortality. A longitudinal study in pima indians. Arthritis Rheum. 1993, 36, 1045–1053. [Google Scholar] [CrossRef] [PubMed]
- Korpilähde, T.; Heliövaara, M.; Kaipiainen-Seppänen, O.; Knekt, P.; Aho, K. Regional differences in Finland in the prevalence of rheumatoid factor in the presence and absence of arthritis. Ann. Rheum. Dis. 2003, 62, 353–355. [Google Scholar] [CrossRef]
- Dresser, D.W. Most IgM-producing cells in the mouse secrete auto-antibodies (rheumatoid factor). Nature 1978, 274, 480–483. [Google Scholar] [CrossRef]
- Goodwin, J.S.; Searles, R.P.; Tung, K.S.K. Immunological responses of a healthy elderly population. Clin. Exp. Immunol. 1982, 48, 403–410. [Google Scholar]
- Van Schaardenburg, D.; Lagaay, A.M.; Breedveld, F.C.; Hijmans, W.; Vandebroucke, J.P. Rheumatoid arthritis in a population of persons aged 85 years and over. Rheumatology 1993, 32, 104–108. [Google Scholar] [CrossRef]
- Ursum, J.; Bos, W.H.; van de Stadt, R.J.; Dijkmans, B.A.C.; van Schaardenburg, D. Different properties of ACPA and IgM-RF derived from a large dataset: Further evidence of two distinct autoantibody systems. Arthritis Res. Ther. 2009, 11. [Google Scholar] [CrossRef]
- Schroeder, H.W.; Cavacini, L.; Schroeder, H.W., Jr.; Cavacini, L.; Schroeder, H.W.; Cavacini, L. Structure and function of immunoglobulins. J. Allergy Clin. Immunol. 2010, 125, S41–S52. [Google Scholar] [CrossRef] [PubMed]
- Meek, B.; Kelder, J.C.; Claessen, A.M.E.; van Houte, A.J.; ter Borg, E.J. Rheumatoid factor isotype and Ro epitope distribution in primary Sjögren syndrome and rheumatoid arthritis with keratoconjunctivitis sicca. Rheumatol. Int. 2018, 38, 1487–1493. [Google Scholar] [CrossRef] [PubMed]
- Sieghart, D.; Platzer, A.; Studenic, P.; Alasti, F.; Grundhuber, M.; Swiniarski, S.; Horn, T.; Haslacher, H.; Blüml, S.; Smolen, J.; et al. Determination of autoantibody isotypes increases the sensitivity of serodiagnostics in rheumatoid arthritis. Front. Immunol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Carson, D.A.; Pasquali, J.L.; Tsoukas, C.D.; Fong, S.; Slovin, S.F.; Lawrance, S.K.; Slaughter, L.; Vaughan, J.H. Physiology and pathology of rheumatoid factors. Springer Semin. Immunopathol. 1981, 4, 161–179. [Google Scholar] [CrossRef]
- Seligmann, M.; Brouet, J.C. Antibody activity of human myeloma globulins. Semin. Hematol. 1973, 10, 163–177. [Google Scholar]
- Shmerling, R.H.; Delbanco, T.L. The rheumatoid factor: An analysis of clinical utility. Am. J. Med. 1991, 91, 528–534. [Google Scholar] [CrossRef]
- Diaz-Lopez, C.; Geli, C.; Corominas, H.; Malat, N.; Diaz-Torner, C.; Llobet, J.M.; De La Serna, A.R.; Laiz, A.; Moreno, M.; Vazquez, G. Are there clinical or serological differences between male and female patients with primary Sjogren’s syndrome? J. Rheumatol. 2004, 31, 1352–1355. [Google Scholar]
- Sansonno, D.; Lauletta, G.; Nisi, L.; Gatti, P.; Pesola, F.; Pansini, N.; Dammacco, F. Non-enveloped HCV core protein as constitutive antigen of cold-precipitable immune complexes in type II mixed cryoglobulinaemia. Clin. Exp. Immunol. 2003, 133, 275–282. [Google Scholar] [CrossRef]
- Hurri, L.; Perttala, Y. Observations on non-specific Waaler-Rose and latex reactions in cancer patients. Ann. Med. Intern. Fenn. 1965, 54, 181–183. [Google Scholar]
- Ugolini, A.; Zizzari, I.G.; Ceccarelli, F.; Botticelli, A.; Colasanti, T.; Strigari, L.; Rughetti, A.; Rahimi, H.; Conti, F.; Valesini, G.; et al. IgM-Rheumatoid factor confers primary resistance to anti-PD-1 immunotherapies in NSCLC patients by reducing CD137+T-cells. EBioMedicine 2020, 62, 103098. [Google Scholar] [CrossRef]
- Allander, E.; Björnsson, O.J.; Kolbeinsson, A.; Olafsson, O.; Sigfússon, N.; Thorsteinsson, J. Rheumatoid factor in Iceland: A population study. Int. J. Epidemiol. 1972, 1, 211–223. [Google Scholar] [CrossRef] [PubMed]
- Thorsteinsson, J.; Björnsson, O.J.; Kolbeinsson, A.; Allander, E.; Sigfússon, N.; Olafsson, O. A population study of rheumatoid factor in Iceland. A 5-year follow-up of 50 women with rheumatoid factor (RF). Ann. Clin. Res. 1975, 7, 183–194. [Google Scholar] [PubMed]
- Ajeganova, S.; Humphreys, J.H.; Verheul, M.K.; Van Steenbergen, H.W.; Van Nies, J.A.B.; Svensson, B.; Hafström, I.J.; Huizinga, T.W.; Trouw, L.A.; Verstappen, S.M.M.; et al. Factor are associated with increased mortality but with different causes of death in patients with rheumatoid arthritis: A longitudinal study in three European cohorts. Ann. Rheum. Dis. 2016, 75. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.K.; Hwang, J.; Chang, Y.; Ryu, S. Rheumatoid factor positivity increases all-cause and cancer mortality: A cohort study. Rheumatol. Int. 2017. [Google Scholar] [CrossRef]
- Gupta, N.P.; Malaviya, A.N.; Singh, S.M. Rheumatoid factor: Correlation with recurrence in transitional cell carcinoma of the bladder. J. Urol. 1979, 121, 417–418. [Google Scholar] [CrossRef]
- Schattner, A.; Shani, A.; Talpaz, M.; Bentwich, Z. Rheumatoid factors in the sera of patients with gastrointestinal carcinoma. Cancer 1983, 52. [Google Scholar] [CrossRef]
- Lochman, I.; Tvrdik, J.; Lochmanova, A.; Machálek, J.; Vrtná, L.; Kyselá, T.; Beska, F.; Repisták, J.; Konrád, B.; Cesaný, P. Antiimmunoglobulins of rheumatoid factor (RF) type in prediction of melanoma patients. Neoplasma 1986, 33, 737–741. [Google Scholar]
- Turnbull, A.R.; Turner, D.T.; Fraser, J.D.; Lloyd, R.S.; Lang, C.J.; Wright, R. Autoantibodies in early breast cancer: A stage-related phenomenon? Br. J. Cancer 1978, 38, 461–463. [Google Scholar] [CrossRef]
- Lewis, M.G.; Hartman, D.; Jerry, L.M. Antibodies and anti-antibodies in human malignancy: An expression of deranged immune regulation. Ann. N. Y. Acad. Sci. 1976, 276, 316–327. [Google Scholar] [CrossRef]
- Ugolini, A.; Zizzari, I.G.; Ceccarelli, F.; Botticelli, A.; Colasanti, T.; Strigari, L.; Rughetti, A.; Rahimi, H.; Conti, F.; Valesini, G.; et al. 4P IgM-rheumatoid factor as a novel biomarker for a reduced survival in anti-PD-1 treated NSCLC patients through the decrease of CD137+ T-cells. Ann. Oncol. 2020, 31, S1418. [Google Scholar] [CrossRef]
- Saksela, E.; Pyrhøonen, S.; Timonen, T.; Teppo, A.-M.; Wager, O.; Penttinen, K. Blocking effect of rheumatoid factor on the in vitro cytotoxicity of lymphoid cells from carcinoma patients. Scand. J. Immunol. 1976, 5, 1075–1080. [Google Scholar] [CrossRef]
- Pyrhönen, S.; Timonen, T.; Heikkinen, A.; Penttinen, K.; Alftan, O.; Saksela, E.; Wager, O. Rheumatoid factor as an indicator of serum blocking activity and tumour recurrences in bladder tumours. Eur. J. Cancer 1976, 12, 87–94. [Google Scholar] [CrossRef]
- Isturiz, M.A.; Maria, M.M.; Pizzi, A.M.; Manni, J.A. Antibody-dependent cell-mediated cytotoxicity in rheumatoid arthritis. Effect of rheumatoid serum fractions on normal lymphocytes. Arthritis Rheum. 1976, 19, 725–730. [Google Scholar] [CrossRef]
- Hallberg, T. In vitro cytotoxicity of human lymphocytes for sensitized chicken erythrocytes is inhibited by sera from rheumatoid arthritis patients. Scand. J. Immunol. 1972, 1, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Giuliano, A.E.; Irie, R.; Morton, D.L. Rheumatoid factor in melanoma patients: Alterations of humoral tumor immunity in vitro. Cancer 1979, 43, 1624–1629. [Google Scholar] [CrossRef]
- Jones, J.D.; Shyu, I.; Newkirk, M.M.; Rigby, W.F.C. A rheumatoid factor paradox: Inhibition of rituximab effector function. Arthritis Res. Ther. 2013, 15. [Google Scholar] [CrossRef]
- Glennie, M.J.; French, R.R.; Cragg, M.S.; Taylor, R.P. Mechanisms of killing by anti-CD20 monoclonal antibodies. Mol. Immunol. 2007, 44, 3823–3837. [Google Scholar] [CrossRef]
- Johnson, P.W.M.; Glennie, M.J. Rituximab: Mechanisms and applications. Br. J. Cancer 2001, 85, 1619–1623. [Google Scholar] [CrossRef]
- King, K.M.; Younes, A. Rituximab: Review and clinical applications focusing on non-Hodgkin’s lymphoma. Expert Rev. Ant Cancer Ther. 2001, 1, 177–186. [Google Scholar] [CrossRef]
- Ugolini, A.; Nuti, M. CD137+ T-Cells: Protagonists of the Immunotherapy Revolution. Cancers 2021, 13, 456. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ugolini, A.; Nuti, M. Rheumatoid Factor: A Novel Determiner in Cancer History. Cancers 2021, 13, 591. https://doi.org/10.3390/cancers13040591
Ugolini A, Nuti M. Rheumatoid Factor: A Novel Determiner in Cancer History. Cancers. 2021; 13(4):591. https://doi.org/10.3390/cancers13040591
Chicago/Turabian StyleUgolini, Alessio, and Marianna Nuti. 2021. "Rheumatoid Factor: A Novel Determiner in Cancer History" Cancers 13, no. 4: 591. https://doi.org/10.3390/cancers13040591
APA StyleUgolini, A., & Nuti, M. (2021). Rheumatoid Factor: A Novel Determiner in Cancer History. Cancers, 13(4), 591. https://doi.org/10.3390/cancers13040591