
Impact of Epithelial–Mesenchymal Transition on the Immune Landscape in Breast Cancer



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients and Tissue Samples
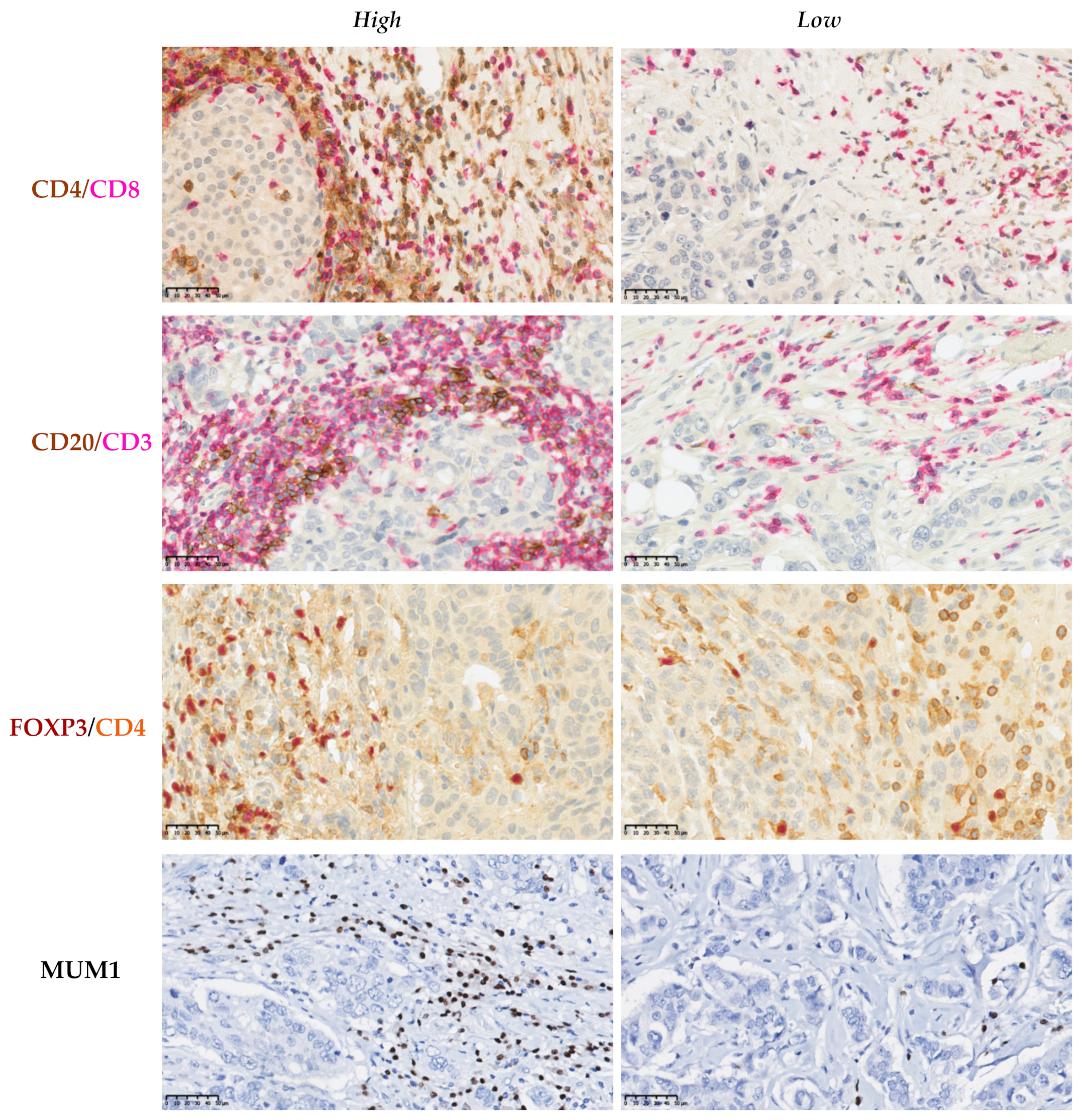
2.2. Assessment of the Inflammatory Infiltrate in the Tissue Samples
2.3. Characterization of the Inflammatory Infiltrate in the TME
2.4. Tissue Microarray
2.5. Immunohistochemistry
2.6. Evaluation of the Immunohistochemical Labeling
2.7. Statistical Analysis
3. Results
3.1. Clinicopathological Characteristics of the Cohort
3.2. Association between the Levels of Expression of EMT-Related Markers and the Clinicopathological Criteria
3.3. The Mesenchymal Markers N-Cadherin and Vimentin Were Associated with the Histological Grade and the Molecular Subtype
3.4. The Epithelial Markers E-Cadherin, GRHL2, and EpCAM Were Associated with the Histological Grade and the Molecular Subtype
3.5. Poorly Differentiated, HER2-Positive and Triple-Negative Carcinomas Were More Often Associated with Inflammatory Infiltrate
3.6. The Expression of Epithelial Markers Related to EMT Was Associated with the Degree of Inflammatory Infiltrate
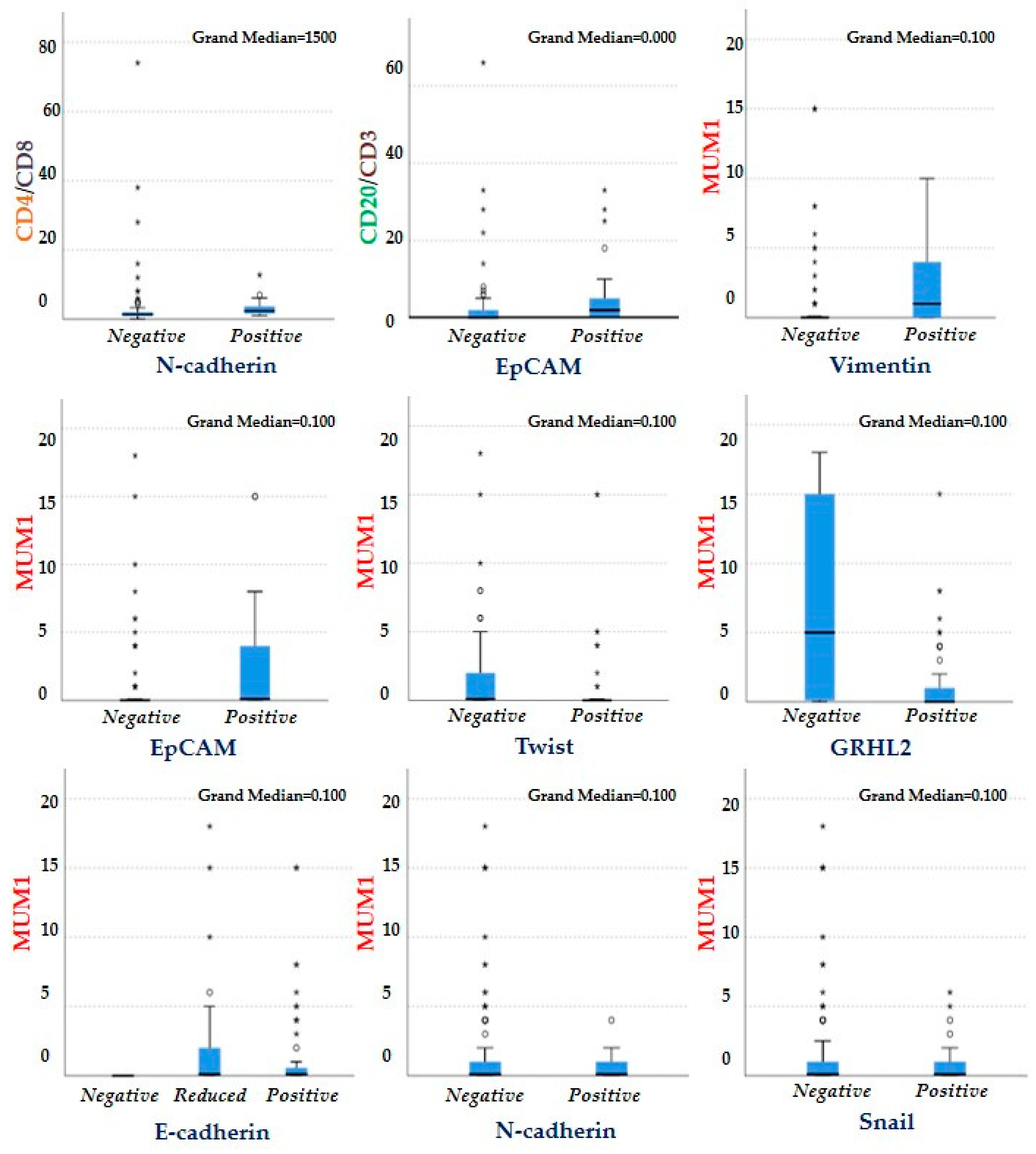
3.7. Association between the EMT-Related Markers and the Different Cells Which Constitute the Immune Infiltrate of the TME
3.8. Association between the Combined Epithelial and Mesenchymal Markers and the Inflammatory Infiltrate
3.9. The Staining Patterns of E-Cadherin and β-Catenin Are Associated and Positively Correlated
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Viale, G. The current state of breast cancer classification. Ann. Oncol. 2012, 23 (Suppl. 10), x207–x210. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Chakrabarti, R. Consequences of EMT-Driven Changes in the Immune Microenvironment of Breast Cancer and Therapeutic Response of Cancer Cells. J. Clin. Med. 2019, 8, 642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.-T.; Liu, H.; Mao, M.-J.; Tan, Y.; Mo, X.-Q.; Meng, X.-J.; Cao, M.-T.; Zhong, C.-Y.; Liu, Y.; Shan, H.; et al. Crosstalk between autophagy and epithelial-mesenchymal transition and its application in cancer therapy. Mol. Cancer 2019, 18, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Sarkissyan, M.; Vadgama, J.V. Epithelial-Mesenchymal Transition and Breast Cancer. J. Clin. Med. 2016, 5, 13. [Google Scholar] [CrossRef] [Green Version]
- Lamouille, S.; Xu, J.; Derynck, R. Molecular mechanisms of epithelial–mesenchymal transition. Nat. Rev. Mol. Cell Biol. 2014, 15, 178–196. [Google Scholar] [CrossRef] [Green Version]
- Bronsert, P.; Enderle-Ammour, K.; Bader, M.; Timme, S.; Kuehs, M.; Csanadi, A.; Kayser, G.; Kohler, I.; Bausch, D.; Hoeppner, J.; et al. Cancer cell invasion and EMT marker expression: A three-dimensional study of the human cancer-host interface. J. Pathol. 2014, 234, 410–422. [Google Scholar] [CrossRef]
- Kröger, C.; Afeyan, A.; Mraz, J.; Eaton, E.N.; Reinhardt, F.; Khodor, Y.L.; Thiru, P.; Bierie, B.; Ye, X.; Burge, C.B.; et al. Acquisition of a hybrid E/M state is essential for tumorigenicity of basal breast cancer cells. Proc. Natl. Acad. Sci. USA 2019, 116, 7353–7362. [Google Scholar] [CrossRef] [Green Version]
- Godin, L.; Balsat, C.; Van Eycke, Y.-R.; Allard, J.; Royer, C.; Remmelink, M.; Pastushenko, I.; D’Haene, N.; Blanpain, C.; Salmon, I.; et al. A Novel Approach for Quantifying Cancer Cells Showing Hybrid Epithelial/Mesenchymal States in Large Series of Tissue Samples: Towards a New Prognostic Marker. Cancers 2020, 12, 906. [Google Scholar] [CrossRef] [Green Version]
- Celià-Terrassa, T.; Jolly, M.K. Cancer Stem Cells and Epithelial-to-Mesenchymal Transition in Cancer Metastasis. Cold Spring Harb. Perspect. Med. 2020, 10, a036905. [Google Scholar] [CrossRef]
- Gupta, P.B.; Pastushenko, I.; Skibinski, A.; Blanpain, C.; Kuperwasser, C. Phenotypic Plasticity: Driver of Cancer Initiation, Progression, and Therapy Resistance. Cell Stem Cell 2019, 24, 65–78. [Google Scholar] [CrossRef] [Green Version]
- Nieto, M.A.; Huang, R.Y.-J.; Jackson, R.A.; Thiery, J.P. EMT: 2016. Cell 2016, 166, 21–45. [Google Scholar] [CrossRef] [Green Version]
- Terry, S.; Savagner, P.; Ortiz-Cuaran, S.; Mahjoubi, L.; Saintigny, P.; Thiery, J.-P.; Chouaib, S. New insights into the role of EMT in tumor immune escape. Mol. Oncol. 2017, 11, 824–846. [Google Scholar] [CrossRef] [Green Version]
- Fischer, K.R.; Durrans, A.; Lee, S.; Sheng, J.; Li, F.; Wong, S.T.C.; Choi, H.; El Rayes, T.; Ryu, S.; Troeger, J.; et al. Epithelial-to-mesenchymal transition is not required for lung metastasis but contributes to chemoresistance. Nature 2015, 527, 472–476. [Google Scholar] [CrossRef]
- Brenot, A.; Knolhoff, B.L.; DeNardo, D.G.; Longmore, G.D. SNAIL1 action in tumor cells influences macrophage polarization and metastasis in breast cancer through altered GM-CSF secretion. Oncogenesis 2018, 7, 32. [Google Scholar] [CrossRef]
- Cortés, M.; Sanchez-Moral, L.; de Barrios, O.; Fernández-Aceñero, M.J.; Martínez-Campanario, M.; Esteve-Codina, A.; Darling, D.S.; Győrffy, B.; Lawrence, T.; Dean, D.C.; et al. Tumor-associated macrophages (TAMs) depend on ZEB1 for their cancer-promoting roles. EMBO J. 2017, 36, 3336–3355. [Google Scholar] [CrossRef]
- Quail, D.F.; Joyce, J.A. Microenvironmental regulation of tumor progression and metastasis. Nat. Med. 2013, 19, 1423–1437. [Google Scholar] [CrossRef]
- Dongre, A.; Rashidian, M.; Reinhardt, F.; Bagnato, A.; Keckesova, Z.; Ploegh, H.L.; Weinberg, R.A. Epithelial-to-Mesenchymal Transition Contributes to Immunosuppression in Breast Carcinomas. Cancer Res. 2017, 77, 3982–3989. [Google Scholar] [CrossRef] [Green Version]
- Mrozik, K.M.; Blaschuk, O.W.; Cheong, C.M.; Zannettino, A.C.W.; Vandyke, K. N-cadherin in cancer metastasis, its emerging role in haematological malignancies and potential as a therapeutic target in cancer. BMC Cancer 2018, 18, 939. [Google Scholar] [CrossRef]
- Wang, Y.; Shi, J.; Chai, K.; Ying, X.; Zhou, B.P. The Role of Snail in EMT and Tumorigenesis. Curr. Cancer Drug Targets 2013, 13, 963–972. [Google Scholar] [CrossRef]
- Qin, Q.; Xu, Y.; He, T.; Qin, C.; Xu, J. Normal and disease-related biological functions of Twist1 and underlying molecular mechanisms. Cell Res. 2012, 22, 90–106. [Google Scholar] [CrossRef] [Green Version]
- Lindner, P.; Paul, S.; Eckstein, M.; Hampel, C.; Muenzner, J.; Erlenbach-Wuensch, K.; Ahmed, H.P.; Mahadevan, V.; Brabletz, T.; Hartmann, A.; et al. EMT transcription factor ZEB1 alters the epigenetic landscape of colorectal cancer cells. Cell Death Dis. 2020, 11, 147. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xu, L.; Li, A.; Han, X. The roles of ZEB1 in tumorigenic progression and epigenetic modifications. Biomed. Pharmacother. 2019, 110, 400–408. [Google Scholar] [CrossRef] [PubMed]
- Cieply, B.; Farris, J.; Denvir, J.; Ford, H.L.; Frisch, S.M. Epithelial–Mesenchymal Transition and Tumor Suppression Are Controlled by a Reciprocal Feedback Loop between ZEB1 and Grainyhead-like-2. Cancer Res. 2013, 73, 6299–6309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spizzo, G.; Fong, D.; Wurm, M.; Ensinger, C.; Obrist, P.; Hofer, C.; Mazzoleni, G.; Gastl, G.; Went, P. EpCAM expression in primary tumour tissues and metastases: An immunohistochemical analysis. J. Clin. Pathol. 2011, 64, 415–420. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Fang, Z.; Ma, J. Regulatory mechanisms and clinical significance of vimentin in breast cancer. Biomed. Pharmacother. 2021, 133, 111068. [Google Scholar] [CrossRef]
- Wellenstein, M.D.; De Visser, K.E. Cancer-Cell-Intrinsic Mechanisms Shaping the Tumor Immune Landscape. Immunity 2018, 48, 399–416. [Google Scholar] [CrossRef] [Green Version]
- Kononen, J.; Bubendorf, L.; Kallioniemi, O.; Bärlund, M.; Schraml, P.; Leighton, S.; Torhorst, J.; Mihatsch, M.J.; Sauter, G.; Kallionimeni, O.-P. Tissue microarrays for high-throughput molecular profiling of tumor specimens. Nat. Med. 1998, 4, 844–847. [Google Scholar] [CrossRef]
- Eble, J.N.; Tavassoli, F.A.; Devilee, P. World health organization classification of tumours. In Pathology and Genetics Tumours of the Breast and Female Genital Organs; IARC Press: Lyon, France, 2003; pp. 19–23. [Google Scholar]
- Kondov, B.; Milenkovikj, Z.; Kondov, G.; Petrushevska, G.; Basheska, N.; Bogdanovska-Todorovska, M.; Tolevska, N.; Ivkovski, L. Presentation of the Molecular Subtypes of Breast Cancer Detected By Immunohistochemistry in Surgically Treated Patients. Open Access Maced. J. Med. Sci. 2018, 6, 961–967. [Google Scholar] [CrossRef] [Green Version]
- Goldhirsch, A.; Winer, E.P.; Coates, A.S.; Gelber, R.D.; Piccart-Gebhart, M.; Thürlimann, B.; Senn, H.J.; Panel Members. Personalizing the treatment of women with early breast cancer: Highlights of the St Gallen International Expert Consensus on the Primary Therapy of Early Breast Cancer 2013. Ann Oncol. 2013, 24, 2206–2223. [Google Scholar] [CrossRef]
- Denkert, C.; Wienert, S.; Poterie, A.; Loibl, S.; Budczies, J.; Badve, S.; Bago-Horvath, Z.; Bane, A.; Bedri, S.; Brock, J.; et al. Standardized evaluation of tumor-infiltrating lymphocytes in breast cancer: Results of the ring studies of the international immuno-oncology biomarker working group. Mod. Pathol. 2016, 29, 1155–1164. [Google Scholar] [CrossRef]
- Nederlof, I.; De Bortoli, D.; Bareche, Y.; Nguyen, B.; De Maaker, M.; Hooijer, G.K.J.; Buisseret, L.; Kok, M.; Smid, M.; Van den Eynden, G.G.; et al. Comprehensive evaluation of methods to assess overall and cell-specific immune infiltrates in breast cancer. Breast Cancer Res. 2019, 21, 151. [Google Scholar] [CrossRef] [Green Version]
- Elmoneim, H.M.A.; Zaghloul, N.M. Expression of e-cadherin, n-cadherin and snail and their correlation with clinicopathologicalvariants: An immunohistochemical study of 132 invasive ductal breast carcinomas in Egypt. Clinics 2011, 66, 1765–1771. [Google Scholar] [CrossRef]
- Siitonen, S.M.; Kononen, J.T.; Helin, H.J.; Rantala, I.S.; Holli, K.A.; Isola, J.J. Reduced E-Cadherin Expression is Associated With Invasiveness and Unfavorable Prognosis in Breast Cancer. Am. J. Clin. Pathol. 1996, 105, 394–402. [Google Scholar] [CrossRef] [Green Version]
- Soysal, S.; Muenst, S.; Barbie, T.; Fleming, T.; Gao, F.; Spizzo, G.; Oertli, D.; Viehl, C.T.; Obermann, E.C.; Gillanders, W.E. EpCAM expression varies significantly and is differentially associated with prognosis in the luminal B HER2+, basal-like, and HER2 intrinsic subtypes of breast cancer. Br. J. Cancer 2013, 108, 1480–1487. [Google Scholar] [CrossRef]
- Hemalatha, A.; Suresh, T.N.; Kumar, M.H. Expression of vimentin in breast carcinoma, its correlation with Ki67 and other histopathological parameters. Indian J. Cancer 2013, 50, 189. [Google Scholar] [CrossRef]
- Borcherding, N.; Cole, K.; Kluz, P.; Jorgensen, M.; Kolb, R.; Bellizzi, A.; Zhang, W. Re-Evaluating E-Cadherin and β-Catenin: A Pan-Cancer Proteomic Approach with an Emphasis on Breast Cancer. Am. J. Pathol. 2018, 188, 1910–1920. [Google Scholar] [CrossRef]
- Varma, K.; Chauhan, A.; Bhargava, M.; Misra, V.; Srivastava, S. Association of different patterns of expression of beta-catenin and cyclin D1 with pathogenesis of breast carcinoma. Indian J. Pathol. Microbiol. 2020, 63, 13–18. [Google Scholar] [CrossRef]
- Stanton, S.E.; Disis, M.L. Clinical significance of tumor-infiltrating lymphocytes in breast cancer. J. Immunother. Cancer 2016, 4, 59. [Google Scholar] [CrossRef] [Green Version]
- Jézéquel, P.; Kerdraon, O.; Hondermarck, H.; Guérin-Charbonnel, C.; Lasla, H.; Gouraud, W.; Canon, J.-L.; Gombos, A.; Dalenc, F.; Delaloge, S.; et al. Identification of three subtypes of triple-negative breast cancer with potential therapeutic implications. Breast Cancer Res. 2019, 21, 65. [Google Scholar] [CrossRef]
- Barnes, T.A.; Amir, E. HYPE or HOPE: The prognostic value of infiltrating immune cells in cancer. Br. J. Cancer 2017, 117, 451–460. [Google Scholar] [CrossRef] [Green Version]
- Gruosso, T.; Gigoux, M.; Manem, V.S.K.; Bertos, N.; Zuo, D.; Perlitch, I.; Saleh, S.M.I.; Zhao, H.; Souleimanova, M.; Johnson, R.M.; et al. Spatially distinct tumor immune microenvironments stratify triple-negative breast cancers. J. Clin. Investig. 2019, 129, 1785–1800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Rooijen, J.M.; Stutvoet, T.S.; Schröder, C.P.; de Vries, E. Immunotherapeutic options on the horizon in breast cancer treatment. Pharmacol. Ther. 2015, 156, 90–101. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Ellison, S.; Alarid, E.T.; Shapiro, D.J. Interplay between the levels of estrogen and estrogen receptor controls the level of the granzyme inhibitor, proteinase inhibitor 9 and susceptibility to immune surveillance by natural killer cells. Oncogene 2007, 26, 4106–4114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mostafa, A.; Codner, D.; Hirasawa, K.; Komatsu, Y.; Young, M.N.; Steimle, V.; Drover, S. Activation of ERα Signaling Differentially Modulates IFN-γ Induced HLA-Class II Expression in Breast Cancer Cells. PLoS ONE 2014, 9, e87377. [Google Scholar] [CrossRef]
- Sahoo, S.; Nayak, S.P.; Hari, K.; Purkait, P.; Mandal, S.; Kishore, A.; Levine, H.; Jolly, M.K. Immunosuppressive traits of the hybrid epithelial/mesenchymal phenotype. bioRxiv 2021. [Google Scholar] [CrossRef]
- Binnewies, M.; Roberts, E.W.; Kersten, K.; Chan, V.; Fearon, D.F.; Merad, M.; Coussens, L.M.; Gabrilovich, D.I.; Ostrand-Rosenberg, S.; Hedrick, C.C.; et al. Understanding the tumor immune mi-croenvironment (TIME) for effective therapy. Nat. Med. 2018, 24, 541–550. [Google Scholar] [CrossRef]
- Lim, B.; Woodward, W.; Wang, X.; Reuben, J.M.; Ueno, N.T. Inflammatory breast cancer biology: The tumour microenvironment is key. Nat. Rev. Cancer 2018, 18, 485–499. [Google Scholar] [CrossRef]
- Yeong, J.; Lim, J.C.; Lee, B.; Li, H.; Chia, N.; Ong, C.C.; Lye, W.K.; Putti, T.C.; Dent, R.; Lim, E.; et al. High densities of tumor-associated plasma cells predict improved prognosis in triple negative breast cancer. Front. Immunol. 2018, 9, 1209. [Google Scholar] [CrossRef]
- Brown, J.R.; Wimberly, H.; Lannin, D.R.; Nixon, C.; Rimm, D.L.; Bossuyt, V. Multiplexed Quantitative Analysis of CD3, CD8, and CD20 Predicts Response to Neoadjuvant Chemotherapy in Breast Cancer. Clin. Cancer Res. 2014, 20, 5995–6005. [Google Scholar] [CrossRef] [Green Version]
- Bar, I.; Theate, I.; Haussy, S.; Beniuga, G.; Carrasco, J.; Canon, J.-L.; Delrée, P.; Merhi, A. MiR-210 Is Overexpressed in Tumor-infiltrating Plasma Cells in Triple-negative Breast Cancer. J. Histochem. Cytochem. 2020, 68, 25–32. [Google Scholar] [CrossRef]
- Wouters, M.C.A.; Nelson, B.H. Prognostic significance of tumor-infiltrating B cells and plasma cells in human cancer. Clin. Cancer Res. 2018, 24, 6125–6135. [Google Scholar] [CrossRef] [Green Version]
- Shen, M.; Wang, J.; Ren, X. New Insights into Tumor-Infiltrating B Lymphocytes in Breast Cancer: Clinical Impacts and Regulatory Mechanisms. Front. Immunol. 2018, 9, 470. [Google Scholar] [CrossRef] [Green Version]
- MadorskyRowdo, F.P.; Baron, A.; Urrutia, M.; Mordoh, J. Immunotherapy in cancer: A combat between tumors and the immune system; you win some, you lose some. Front. Immunol. 2015, 6, 127. [Google Scholar] [CrossRef] [Green Version]
- Ding, Z.C.; Zhou, G. Cytotoxic chemotherapy and CD4+ effector T cells: An emerging alliance for durable antitumor effects. Clin. Dev. Immunol. 2012, 2012, 890178. [Google Scholar] [CrossRef]
- Kristensen, V.N.; Vaske, C.; Ursini-Siegel, J.; Van Loo, P.; Nordgard, S.H.; Sachidanandam, R.; Sorlie, T.; Wärnberg, F.; Haakensen, V.D.; Helland, Å.; et al. Integrated molecular profiles of invasive breast tumors and ductal carcinoma in situ (DCIS) reveal differential vascular and interleukin signaling. Proc. Natl. Acad. Sci. USA 2012, 109, 2802–2807. [Google Scholar] [CrossRef] [Green Version]
- Emens, L.A. Breast cancer immunobiology driving immunotherapy: Vaccines and immune checkpoint blockade. Expert Rev. Anticancer. Ther. 2012, 12, 1597–1611. [Google Scholar] [CrossRef] [Green Version]
- DeNardo, D.G.; Brennan, D.J.; Rexhepaj, E.; Ruffell, B.; Shiao, S.L.; Madden, S.F.; Gallagher, W.M.; Wadhwani, N.; Keil, S.D.; Junaid, S.A.; et al. Leukocyte Complexity Predicts Breast Cancer Survival and Functionally Regulates Response to Chemotherapy. Cancer Discov. 2011, 1, 54–67. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Gibbons, D.L.; Goswami, S.; Cortez, M.A.; Ahn, Y.-H.; Byers, L.A.; Zhang, X.; Yi, X.; Dwyer, D.; Lin, W.; et al. Metastasis is regulated via microRNA-200/ZEB1 axis control of tumour cell PD-L1 expression and intratumoral immunosuppression. Nat. Commun. 2014, 5, 5241. [Google Scholar] [CrossRef]
- Aiello, N.; Maddipati, R.; Norgard, R.J.; Balli, D.; Li, J.; Yuan, S.; Yamazoe, T.; Black, T.; Sahmoud, A.; Furth, E.E.; et al. EMT Subtype Influences Epithelial Plasticity and Mode of Cell Migration. Dev. Cell 2018, 45, 681–695.e4. [Google Scholar] [CrossRef] [Green Version]
- Sahoo, S.; Mishra, A.; Kaur, H.; Hari, K.; Muralidharan, S.; Mandal, S.; Jolly, M.K. A mechanistic model captures the emergence and implications of non-genetic heterogeneity and reversible drug resistance in ER+ breast cancer cells. NAR Cancer 2021, 3, zcab027. [Google Scholar] [CrossRef]
- Kai, K.; Masuda, M.; Aishima, S. Inverse correlation between CD8+ inflammatory cells and E-cadherin expression in gallbladder cancer: Tissue microarray and imaging analysis. World J. Clin. Cases 2017, 5, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.-Y.; Tseng, Y.-K.; Chen, Y.-C.; Shu, C.-W.; Lin, M.-I.; Liou, H.-H.; Fu, T.-Y.; Lin, Y.-C.; Ger, L.-P.; Yeh, M.-H.; et al. High snail expression predicts a poor prognosis in breast invasive ductal carcinoma patients with HER2/EGFR-positive subtypes. Surg. Oncol. 2018, 27, 314–320. [Google Scholar] [CrossRef] [PubMed]
- Blanco, M.J.; Moreno-Bueno, G.; Sarrio, D.; Locascio, A.; Cano, A.; Palacios, J.; Nieto, M.Á. Correlation of Snail expression with histological grade and lymph node status in breast carcinomas. Oncogene 2002, 21, 3241–3246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, T.; Tan, R.; Wang, L.; Song, J.; Li, J. Expression and significance of Twist, estrogen receptor, and E-cadherin in human breast cancer cells and tissues. J. Cancer Res. Ther. 2017, 13, 707–714. [Google Scholar] [CrossRef]
- Berx, G.; Van Roy, F. The E-cadherin/catenin complex: An important gatekeeper in breast cancer tumorigenesis and malignant progression. Breast Cancer Res. 2001, 3, 289. [Google Scholar] [CrossRef]
- Acs, G.; Lawton, T.J.; Rebbeck, T.R.; Livolsi, V.A.; Zhang, P.J. Differential Expression of E-Cadherin in Lobular and Ductal Neoplasms of the Breast and Its Biologic and Diagnostic Implications. Am. J. Clin. Pathol. 2001, 115, 85–98. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, R.; Kimura, N.; Harada, Y.; Ohuchi, N. The loss of E-cadherin, alpha- and beta-catenin expression is associated with metastasis and poor prognosis in invasive breast cancer. Int. J. Oncol. 2001, 18, 513–520. [Google Scholar]
- Xiang, X.; Deng, Z.; Zhuang, X.; Ju, S.; Mu, J.; Jiang, H.; Zhang, L.; Yan, J.; Miller, D.; Zhang, H.-G. Grhl2 Determines the Epithelial Phenotype of Breast Cancers and Promotes Tumor Progression. PLoS ONE 2012, 7, e50781. [Google Scholar] [CrossRef]
- Sorlie, T.; Perou, C.M.; Tibshirani, R.; Aas, T.; Geisler, S.; Johnsen, H.; Hastie, T.; Eisen, M.B.; van de Rijn, M.; Jeffrey, S.S.; et al. Gene expression patterns of breast carcinomas distinguish tumor subclasses with clinical implications. Proc. Natl. Acad. Sci. USA 2001, 98, 10869–10874. [Google Scholar] [CrossRef] [Green Version]
- Münz, M.; Kieu, C.; Mack, B.; Schmitt, B.; Zeidler, R.; Gires, O. The carcinoma-associated antigen EpCAM upregulates c-myc and induces cell proliferation. Oncogene 2004, 23, 5748–5758. [Google Scholar] [CrossRef]
- Winter, M.J.; Nagelkerken, B.; Mertens, A.E.; Rees-Bakker, H.A.; Bruijn, I.H.B.-D.; Litvinov, S.V. Expression of Ep-CAM shifts the state of cadherin-mediated adhesions from strong to weak. Exp. Cell Res. 2003, 285, 50–58. [Google Scholar] [CrossRef]
- Huelsken, J.; Behrens, J. The Wntsignalling pathway. J. Cell Sci. 2002, 115, 3977–3978. [Google Scholar] [CrossRef] [Green Version]
- Osta, W.A.; Chen, Y.; Mikhitarian, K.; Mitas, M.; Salem, M.; Hannun, Y.A.; Cole, D.J.; Gillanders, W.E. EpCAM Is Overexpressed in Breast Cancer and Is a Potential Target for Breast Cancer Gene Therapy. Cancer Res. 2004, 64, 5818–5824. [Google Scholar] [CrossRef] [Green Version]
- Balzar, M.; Prins, F.A.; Bakker, H.A.; Fleuren, G.J.; Warnaar, S.O.; Litvinov, S.V. The Structural Analysis of Adhesions Mediated by Ep-CAM. Exp. Cell Res. 1999, 246, 108–121. [Google Scholar] [CrossRef]
- Sankpal, N.V.; Fleming, T.P.; Sharma, P.K.; Wiedner, H.; Gillanders, W.E. A double-negative feedback loop between EpCAM and ERK contributes to the regulation of epithelial–mesenchymal transition in cancer. Oncogene 2017, 36, 3706–3717. [Google Scholar] [CrossRef] [Green Version]
- Driemel, C.; Kremling, H.; Schumacher, S.; Will, D.; Wolters, J.; Lindenlauf, N.; Mack, B.; Baldus, S.A.; Hoya, V.; Pietsch, J.M.; et al. Context-dependent adaption of EpCAM expression in early systemic esophageal cancer. Oncogene 2013, 33, 4904–4915. [Google Scholar] [CrossRef] [Green Version]
- Markiewicz, A.; Wełnicka-Jaśkiewicz, M.; Seroczyńska, B.; Skokowski, J.; Majewska, H.; Szade, J.; Żaczek, A.J. Epithelial-mesenchymal transition markers in lymph node metastases and primary breast tumors-relation to dissemination and proliferation. Am. J. Transl. Res. 2014, 6, 793–808. [Google Scholar]
- Savci-Heijink, C.D.; Halfwerk, H.; Hooijer, G.K.J.; Koster, J.; Horlings, H.M.; Meijer, S.L.; Van De Vijver, M.J. Epithelial-to-mesenchymal transition status of primary breast carcinomas and its correlation with metastatic behavior. Breast Cancer Res. Treat. 2019, 174, 649–659. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Source | Clone | Dilution | Unmasking | Incubation (min) * | Detection | Location of the Staining |
|---|---|---|---|---|---|---|---|
| Snail (AMAb91215) | Atlas antibody Bromma, Sweden | CL3700 | 1/3000 | Citrate | 30/15/15 | Peroxidase polymer (HRP-DAB) | Nuclear |
| Twist (ab50581) | Abcam Cambridge, UK | 1/1000 | Citrate | 15/0/8 | Peroxidase polymer (HRP-DAB) | Nuclear | |
| ZEB1 (ab180905) | Abcam Cambridge, UK | OTI3G6 | 1/1000 | Citrate | 15/8/8 | Peroxidase polymer (HRP-DAB) | Nuclear |
| E-cadherin (M3612) | DAKO Agilent Santa Clara, CA, USA | NCH-38 | 1/50 | EDTA | 60/30/30 | Peroxidase polymer (HRP-DAB) | Membrane |
| N-cadherin (ab225719) | Abcam Cambridge, UK | SP90 | 1/50 | EDTA | 30/0/15 | Peroxidase polymer (HRP-DAB) | Membrane |
| EpCAM (ab223582) | Abcam Cambridge, UK | EPR20532-225 | 1/500 | EDTA | 30/0/15 | Peroxidase polymer (HRP-DAB) | Membrane |
| GRHL2 | Sigma-Aldrich St. Louis, MO, USA | HPA004820 | 1/100 | EDTA | 30/0/15 | Peroxidase polymer (HRP-DAB) | Nuclear |
| Vimentin Bs-0756R | Bioss Antibodies Inc. Woburn, MA, USA | 1/500 | Citrate | 30/0/15 | Peroxidase polymer (HRP-DAB) | Cytoplasm | |
| β-catenin | Ventana Medical Systems, Roche Diagnostics, Canada | 14 | RTU | EDTA | 24/28 | Peroxidase polymer (HRP-DAB) | Membrane |
| CD3 (PA0553) | Leica Biosystems, Newcastle, UK | LN10 | BOND RTU | EDTA | Protocol F | Red polymer (AP-Fast Red) | Membrane |
| CD4 (M7310) | DAKO Agilent Santa Clara, CA, USA | 4B12 | 1/50 | EDTA | 30/15/15 | Peroxidase polymer (HRP-DAB) | Membrane |
| CD8 (GA623) | DAKO Agilent Santa Clara, CA, USA | C8/144B | 1/200 | EDTA | 30/15/15 | Red polymer (AP-Fast Red) | Membrane |
| CD20 (PA0200) | Leica Biosystems, Newcastle, UK | L26 | BOND RTU | Citrate | Peroxidase polymer (HRP-DAB) | Membrane | |
| FoxP3 (ab20034) | Abcam Cambridge, UK | 236A/E7 | 1/100 | EDTA | 30/15/15 | Red polymer (AP-Fast Red) | Nuclear |
| MUM1 (GA644) | DAKO Agilent Santa Clara, CA, USA | MUM1p | 1/200 | EDTA | 48/32 | Peroxidase polymer (HRP-DAB) | Nuclear |
| Clinicopathological Characteristics | Number (Percentage) |
|---|---|
| Histological subtype | |
| Infiltrating ductal carcinoma (IDC) | 126 (87.5%) |
| Infiltrating lobular carcinoma (ILC) | 14 (9.7%) |
| Undifferentiated medullary-like carcinoma | 4 (2.8%) |
| Histological grade | |
| Grade I | 31 (21.5%) |
| Grade II | 50 (34.7%) |
| Grade III | 63 (43.8%) |
| Molecular subtype | |
| Luminal (A/B) | 91 (63.2%) |
| HER2-positive | 10 (6.9%) |
| Triple-negative | 43 (29.9%) |
| Lymph node metastases | |
| Yes | 42 (29.2%) |
| No | 97 (67.3%) |
| Not documented | 5 (3.5%) |
| Clinicopathological Criteria | Snail | Twist | N-Cadherin | Vimentin | ||||
|---|---|---|---|---|---|---|---|---|
| Positive N (%) | Negative N (%) | Positive N (%) | Negative N (%) | Positive N (%) | Negative N (%) | Positive N (%) | Negative N (%) | |
| Histological subtype | ||||||||
| IDC ILC UC-MLC | 25 (21.7%) 1 (9%) 0 (0%) | 90 (78.3%) 11 (91%) 4 (100%) | 52 (45.2%) 8 (61.5%) 0 (0%) | 63 (54.8%) 5 (38.5%) 4 (100%) | 15 (12.8%) 0 (0%) 1 (25%) | 102 (87.2%) 13 (100%) 3 (75%) | 28 (25.2%) 0 (0%) 2 (50%) | 83 (74.8%) 14 (100%) 2 (50%) |
| p = 0.325 | p = 0.95 | p = 0.285 | p = 0.048 | |||||
| Histological grade | ||||||||
| Grade I Grade II Grade III | 3 (12.5%) 7 (14.5%) 16 (38%) | 21 (87.5%) 41 (85.5%) 42 (62%) | 12 (44.4%) 25 (53.2%) 23 (39.6%) | 15 (55.6%) 22 (46.8%) 35 (60.4%) | 1 (3.8%) 3 (6.7%) 12 (20%) | 25 (96.2%) 45 (93.3%) 48 (80%) | 3 (10.4%) 1 (2.2%) 26 (48.2%) | 26 (89.6%) 45 (97.8%) 28 (51.8%) |
| p = 0.149 | p = 0.380 | p = 0.033 | p < 0.0001 | |||||
| Molecular subtype | ||||||||
| Luminal A andB Her2+ Triple-negative | 12 (15%) 2 (22.2%) 12 (29.3%) | 68 (85%) 7 (77.8%) 29 (70.6%) | 40 (48.8%) 4 (40%) 16 (40%) | 42 (51.2%) 6 (60%) 24 (60%) | 3 (3.7%) 2 (20%) 11 (26.8%) | 79 (96.3%) 8 (80%) 30 (73.2%) | 4 (4.8%) 1 (12.5%) 25 (65.8%) | 79 (95.2%) 7 (87.5%) 13 (34.2%) |
| p = 0.176 | p = 0.617 | p = 0.001 | p < 0.0001 | |||||
| Lymph-node metastasis | ||||||||
| Yes No | 9 (21.9%) 16 (18.8%) | 32 (78.1%) 69 (81.2%) | 21 (51.2%) 36 (41.9%) | 20 (48.8%) 50 (58.1%) | 2 (4.8%) 14 (17.9%) | 40 (95.2%) 73 (82.1%) | 7 (18.4%) 22 (24.7%) | 31 (81.6%) 67 (75.3%) |
| p = 0.86 | p = 0.42 | p = 0.12 | p = 0.58 | |||||
| Clinicopathological Criteria | E-Cadherin | GRHL2 | EpCAM | ||||
|---|---|---|---|---|---|---|---|
| Positive N (%) | Reduced N (%) | Negative N (%) | Positive N (%) | Negative N (%) | Positive N (%) | Negative N (%) | |
| Histological subtype | |||||||
| IDC ILC UC-MLC | 75 (61.5%) 0 (0%) 1 (25%) | 44 (36%) 4 (28.6%) 3 (75%) | 3 (2.5%) 10 (71.4%) 0 (0%) | 109 (90.8%) 14 (100%) 4 (100%) | 11 (9.2%) 0 (0%) 0 (0%) | 40 (33.3%) 0 (%) 2 (50%) | 80 (66.7%) 13 (100%) 2 (50%) |
| p < 0.001 | p = 0.408 | p = 0.032 | |||||
| Histological grade | |||||||
| Grade I Grade II Grade III | 20 (71.4%) 29 (59.2%) 27 (42.8%) | 6 (21.4%) 11 (22.4%) 34 (54%) | 2 (7.2%) 9 (18.4%) 2 (3.2%) | 29 (100%) 48 (100%) 50 (82%) | 0 (0%) 0 (0%) 11 (18%) | 4 (14.3%) 4 (8.3%) 34 (54%) | 24 (85.7%) 44 (91.7%) 27 (46%) |
| p = 0.001 | p = 0.001 | p < 0.001 | |||||
| Molecular subtype | |||||||
| Luminal A and B Her2+ Triple-negative | 55 (63.2%) 6 (60%) 15 (34.9%) | 21 (24.2%) 4 (40%) 26 (60.5%) | 11 (12.6%) 0 (0%) 2 (4.6%) | 85 (98.8%) 9 (90%) 33 (78.6%) | 1 (1.2%) 1 (10%) 9 (21.4%) | 11 (12.9%) 3 (30%) 28 (66.7%) | 74 (87.1%) 7 (70%) 14 (33.3%) |
| p = 0.001 | p < 0.001 | p < 0.001 | |||||
| Lymph-node metastasis | |||||||
| Yes No | 20 (47.6%) 53 (57%) | 15 (35.7%) 34 (36.6%) | 7 (16.7%) 6 (6.4%) | 37 (88.1%) 88 (94.6%) | 5 (11.9%) 5 (5.4%) | 10 (24.4%) 53 (46.5%) | 31 (75.6%) 61 (53.5%) |
| p = 0.16 | p = 0.32 | p = 0.02 | |||||
| Clinicopathological Criteria | Inflammatory Infiltrate (H&E-Stained WS) | Inflammatory Infiltrate (IHC-Stained TMAs) | ||
|---|---|---|---|---|
| Mild N (%) | Intense N (%) | Mild N (%) | Intense N (%) | |
| Histological subtype | ||||
| IDC | 100 (79.4%) | 26 (20.6%) | 84 (75.7%) | 27 (24.3%) |
| ILC | 13 (92.8%) | 1 (7.2%) | 13 (92.8%) | 1 (7.2%) |
| UC-MLC | 1 (25%) | 3 (75%) | 1 (25%) | 3 (75%) |
| p = 0.013 | p = 0.02 | |||
| Histological Grade | ||||
| Grade I | 31 (100%) | 0 (0) | 24 (100%) | 0 (0) |
| Grade II | 46 (92%) | 4 (8%) | 44 (93.6%) | 3 (6.4%) |
| Grade III | 37 (58.7%) | 26 (41.3%) | 29 (50.9%) | 28 (49.1%) |
| p < 0.0001 | p < 0.0001 | |||
| Molecular subtype | ||||
| Luminal A | 64 (98.5%) | 1 (1.5%) | 58 (100%) | 0 (0%) |
| Luminal B | 21 (80.8%) | 5 (19.2 5) | 16 (76.2%) | 5 (23.8) |
| HER2+ | 4 (40%) | 6 (60%) | 3 (33.3%) | 6 (66.7%) |
| Triple-negative | 25 (58.1%) | 18 (41.9%) | 20 (52.6%) | 18 (47.4%) |
| p < 0.0001 | p < 0.0001 | |||
| Lymph-node metastasis | ||||
| Yes | 31 (73.8%) | 11 (26.2%) | 26 (66.7%) | 13 (33.3%) |
| No | 80 (82.5%) | 17 (17.5%) | 66 (79.5%) | 17 (20.5%) |
| p = 0.34 | p = 0.19 | |||
| EMT-Related Markers | Infiltrate (H&E-Stained WS) | Infiltrate (IHC-Stained TMAs) | |||
|---|---|---|---|---|---|
| Mild | Intense | Mild | Intense | ||
| Snail | Negative | 82 (78.8%) | 22 (21.2%) | 75 (78.9%) | 20 (21.1%) |
| Positive | 21 (80.8%) | 5 (19.2%) | 18 (72%) | 7 (28%) | |
| p = 1.000 | p = 0.769 | ||||
| Twist | Negative | 54 (75%) | 18 (25%) | 50 (71.4%) | 20 (28.6%) |
| Positive | 50 (83.3%) | 10 (16.7%) | 44 (83%) | 9 (17%) | |
| p = 0.34 | p = 0.199 | ||||
| N-cadherin | Negative | 94 (79.7%) | 24 (20.3%) | 85 (76.6%) | 26 (23.4%) |
| Positive | 12 (75%) | 4 (25%) | 11 (73.3%) | 4 (26.6%) | |
| p = 0.92 | p = 1.000 | ||||
| Vimentin | Negative | 84 (84.8%) | 15 (15.2%) | 78 (84.8%) | 14 (15.2%) |
| Positive | 20 (66.7%) | 10 (33.3%) | 16 (57.1%) | 12 (42.9%) | |
| p = 0.052 | p = 0.004 | ||||
| E-cadherin | Negative | 13 (100%) | 0 (0%) | 12 (92.3%) | 1 (7.7%) |
| Reduced | 33 (64.7%) | 18 (35.3%) | 29 (63%) | 17 (37%) | |
| Positive | 64 (84.2%) | 12 (15.8%) | 54 (80.5%) | 13 (19.5%) | |
| p = 0.005 | p = 0.03 | ||||
| GRHL2 | Negative | 5 (45.4%) | 6 (54.6%) | 1 (10%) | 9 (90%) |
| Positive | 105 (82%) | 23 (18%) | 94 (81%) | 22 (19%) | |
| p = 0.013 | p = 0.001 | ||||
| EpCAM | Negative | 80 (84.2%) | 15 (15.8%) | 76 (83.5%) | 15 (16.5%) |
| Positive | 28 (66.7%) | 14 (33.3%) | 19 (54.3%) | 16 (45.7%) | |
| p = 0.037 | p < 0.0001 | ||||
| Snail | Twist | N-Cadherin | Vimentin | E-Cadherin | Ep CAM | GRHL2 | |
|---|---|---|---|---|---|---|---|
| CD4/CD8 | p = 0.62 | p = 0.24 | p = 0.004 | p = 0.25 | p = 0.34 | p = 0.52 | p = 0.33 |
| FOXP3/CD4 | p = 0.38 | p = 0.93 | p = 0.09 | p = 0.59 | p = 0.25 | p = 0.82 | p = 0.64 |
| CD20/CD3 | p = 0.15 | p = 0.53 | p = 0.72 | p = 0.05 | p = 0.35 | p = 0.03 | p = 0.29 |
| MUM1 | p = 0.99 | p = 0.039 | p = 0.87 | p = 0.001 | p = 0.005 | p = 0.008 | p = 0.03 |
| Inflammatory Infiltrate | p-Value | ||
|---|---|---|---|
| Mild | Intense | ||
| Group 1: E-cadherin positive + absence of expression of 4 mesenchymal markers | 39 (84.7%) | 7 (15.3%) | p = 0.03 |
| Group 2: E-cadherin positive or reduced + presence of 1 or more mesenchymal markers | 32 (65.3%) | 17 (34.7%) | |
| Group 3: E-cadherin negative+ absence of expression of 4 mesenchymal markers | 11 (100%) | 0 (0%) | |
| Group 4: E-cadherin reduced + absence of expression of 4 mesenchymal markers | 14 (77.8%) | 4 (22.2%) | |
| β-Catenin | |||||
|---|---|---|---|---|---|
| Negative | Membranous | Grade1 | Grade2 | ||
| E-cadherin | Negative | 9 (100%) | 0 | 0 | 0 |
| Reduced | 4 (9.7%) | 1 (2.4%) | 21 (51.2%) | 15(36.6%) | |
| Positive | 0 | 8 (12.3%) | 29 (44.6%) | 28 (43.1%) | |
| p < 0.0001 (p = 0.02) * | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khadri, F.-Z.; Issac, M.S.M.; Gaboury, L.A. Impact of Epithelial–Mesenchymal Transition on the Immune Landscape in Breast Cancer. Cancers 2021, 13, 5099. https://doi.org/10.3390/cancers13205099
Khadri F-Z, Issac MSM, Gaboury LA. Impact of Epithelial–Mesenchymal Transition on the Immune Landscape in Breast Cancer. Cancers. 2021; 13(20):5099. https://doi.org/10.3390/cancers13205099
Chicago/Turabian StyleKhadri, Fatima-Zohra, Marianne Samir Makboul Issac, and Louis Arthur Gaboury. 2021. "Impact of Epithelial–Mesenchymal Transition on the Immune Landscape in Breast Cancer" Cancers 13, no. 20: 5099. https://doi.org/10.3390/cancers13205099
APA StyleKhadri, F.-Z., Issac, M. S. M., & Gaboury, L. A. (2021). Impact of Epithelial–Mesenchymal Transition on the Immune Landscape in Breast Cancer. Cancers, 13(20), 5099. https://doi.org/10.3390/cancers13205099