
Interaction of Adenosine, Modified Using Carborane Clusters, with Ovarian Cancer Cells: A New Anticancer Approach against Chemoresistance

 , ,
, ,  , ,
, ,  and
and 
Abstract
Simple Summary
Abstract

1. Introduction
2. Results
2.1. Principle of the Study
- modifications at the N6 position of purine, compounds 14, 15, and 16;
- modification at the C8 of the nucleobase, compounds 18, 19, and 20; and
- modification at the C2′ position of the ribose moiety, compounds 22, 23, and 24.
- 3-cobalt-bis(1,2-dicarba-closo-undecaborate](−1) (3) (1),
- 3-ferra-bis(1,2-dicarba-closo-undecaborate](−1) (2),
- 3-chromma-bis(1,2-dicarba-closo-undecaborate](−1) (3).
2.2. Cytotoxicity of the Compounds in Relation to the Resistance of the Cells to Cisplatin
2.2.1. Sensitive A2780 Cell Line
2.2.2. Resistant A2780cis Cell Line and Highly Resistant A2780cisR and A2780cisKB Cells
2.2.3. OVCAR-3 and SKOV-3 Cell Lines
2.3. Structure–Activity Relationship. The Presence of Nucleoside, Role of Metal Ions, and Substitution Site
2.3.1. Role of Nucleoside and Metal Ion
2.3.2. Role of Substitution Site
2.4. Accumulation of the Compounds in the Cells
2.5. Apoptosis, Necrosis, and Reactive Oxygen Species (ROS) Production
2.6. Phases of Cell Cycle
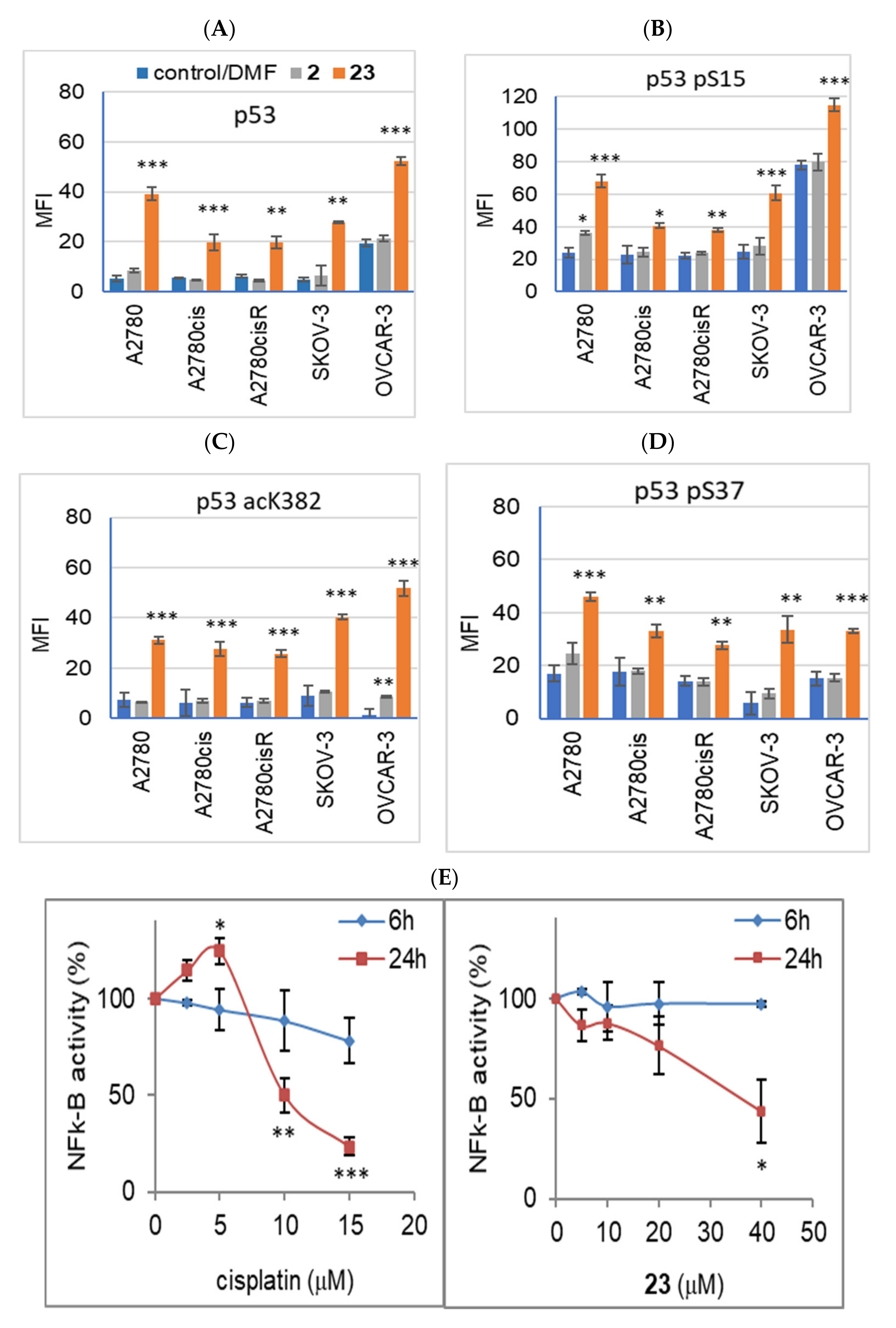
2.7. Effect of Compound 23 on Tumor Suppressor Protein p53 Expression and its Phosphorylated and Acetylated Forms
2.8. Activation of Transcription Factors—AP-1 and NF-κB
2.9. Long-Term Cell Cultures
Growth Rate and Doubling Time
2.10. Three-Dimensional Spheroid Cell Cultures
2.11. Preparation of Metallacarborane- and Nido-7,8-Dicarbaundecaborate Derivatives
- 2′-deoxyadenosine derivatives containing modification at the N6 position of adenine, 6-N-{5-[3,3′-metalla-bis(1,2-dicarba-closo-undekaborate)-8-yl]-3-oxa-pentoxy}-2′-deoxyadenosine, compounds 14–16;
- derivatives modified at C8 position of 2′-deoxyadenosine purine system, 8-{5-[3,3′-metalla-bis(1,2-dicarba-closo-undekaborate)-8-yl]-3-oxa-pentoxy}-1N-1,2,3-triazol-4-yl}-2′-deoxyadenosine, compounds 18–20; and
- derivatives of adenosine modified at C2′ of ribose moiety, 2′-O-{{5-[3,3′-cobalta-bis(1,2-dicarba-closo-undekaborate)-8-yl]-3-oxa-pentoxy}-1N-1,2,3-triazol-4-yl}propyloadenosine compounds 22–24.
2.11.1. Preparation of Dioxane Adducts
2.11.2. Synthesis of 2′-Deoxyadenosine and Adenosine- Metallacarborane Conjugates 14–24
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Cell Culture Reagents
4.2.1. P53 Antibodies
4.2.2. Drugs
4.2.3. Preparation of Stock Solutions of Drugs and Compounds for Biological Study
4.3. Cell Lines and Growth Conditions
- A2780 cell line (ECACC 93112519, Sigma-Aldrich), parental cisplatin-sensitive;
- A2780cis cell line (ECACC 93112517, Sigma-Aldrich), cisplatin-resistant;
- A2780cisR and A2780cisKB cell lines of increased platinum resistance or refractory, obtained experimentally for the purpose of this study;
- SKOV-3 stable cell line, cisplatin-resistant; and
- NIH:OVCAR-3 ovarian adenocarcinoma cell line (ATCC-HTB-161; LGC Standards, Teddington, UK), relatively cisplatin-resistant/sensitive.
4.4. Determination of Compound Cytotoxicity
4.5. Accumulation of Compounds in Cells
4.6. Apoptosis, Necrosis, and ROS Production
4.6.1. PI/Annexin V Assay
4.6.2. DCF-DA Cellular-Based Assay
4.6.3. Plasma Membrane Potential
4.7. Cell Cycle
4.8. Expression Level of Tumor Suppressor p53 Protein
4.9. Reporter Gene Cell Lines and AP-1 and NF-κB Activity Assays
4.10. Long-Term Cultures
4.10.1. Long-Term Treatment with the Compounds
4.10.2. Cell Response to Cisplatin, Carboplatin, Paclitaxel, Doxorubicin, or Gemcitabine
4.11. Three-Dimensional Cultures, Spheroids
4.11.1. Spheroid Formation
4.11.2. Treatment of Spheroids with Compounds
4.11.3. Determination of Spheroid Viability
4.11.4. Microscopic Imaging of Spheroids
4.12. Statistical Analysis
4.13. Physicochemical Characterization of Compounds
4.14. X-ray Crystallography
4.15. Synthesis of Compounds
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lheureux, S.; Gourley, C.; Vergote, I.; Oza, A.M. Epithelial ovarian cancer. Lancet 2019, 393, 1240–1253. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2018. CA Cancer J. Clin. 2018, 68, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Bowtell, D.D.; Böhm, S.; Ahmed, A.A.; Aspuria, P.-J.; Bast, R.C.; Beral, V.; Berek, J.S.; Birrer, M.J.; Blagden, S.; Bookman, M.A.; et al. Rethinking ovarian cancer II: Reducing mortality from high-grade serous ovarian cancer. Nat. Rev. Cancer 2015, 15, 668–679. [Google Scholar] [CrossRef]
- Jayson, G.C.; Kohn, E.C.; Kitchener, H.C.; Ledermann, J.A. Ovarian cancer. Lancet 2014, 9951, 1376–1388. [Google Scholar] [CrossRef]
- Blay, J.; White, T.D.; Hoskin, D.W. The Extracellular Fluid of Solid Carcinomas Contains Immunosuppressive Concentrations of Adenosine. Cancer Res. 1997, 57, 2602–2605. [Google Scholar]
- Von Strandmann, E.P.; Reinartz, S.; Wager, U.; Müller, R. Tumor–Host Cell Interactions in Ovarian Cancer: Pathways to Therapy Failure. Trends Cancer 2017, 3, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Zou, X.; Zhao, Y.; Liang, X.; Wang, H.; Zhu, Y.; Shao, Q. Double Insurance for OC: miRNA-Mediated Platinum Resistance and Immune Escape. Front. Immunol. 2021, 12, 641937. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, H.; Maruyama, T.; Urade, Y.; Nagata, S. Immunosuppression via adenosine receptor activation by adenosine monophosphate released from apoptotic cells. eLife 2014, 3, e02172. [Google Scholar] [CrossRef]
- Allard, B.; Longhi, M.S.; Robson, S.C.; Stagg, J. The ectonucleotidases CD39 and CD73: Novel checkpoint inhibitor targets. Immunol. Rev. 2017, 276, 121–144. [Google Scholar] [CrossRef] [PubMed]
- Azambuja, J.H.; Ludwig, N.; Braganhol, E.; Whiteside, T.L. Inhibition of the Adenosinergic Pathway in Cancer Rejuvenates Innate and Adaptive Immunity. Int. J. Mol. Sci. 2019, 20, 5698. [Google Scholar] [CrossRef]
- Sadej, R.; Skladanowski, A.C. Dual, enzymatic and non-enzymatic, function of ecto-5′-nucleotidase (eN, CD73) in migration and invasion of A375 melanoma cells. Acta Biochim. Pol. 2012, 59, 7–52. [Google Scholar] [CrossRef]
- Jiang, T.; Xu, X.; Qiao, M.; Li, X.; Zhao, C.; Zhou, F.; Gao, G.; Wu, F.; Chen, X.; Su, C.; et al. Comprehensive evaluation of NT5E/CD73 expression and its prognostic significance in distinct types of cancers. BMC Cancer 2018, 18, 50. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Ramírez, A.S.; Díaz-Muñoz, M.; Battastini, A.M.; Campos-Contreras, A.; Olvera, A.; Bergamin, L.; Glaser, T.; Moritz, C.E.J.; Ulrich, H.; Vázquez-Cuevas, F.G. Cellular Migration Ability Is Modulated by Extracellular Purines in Ovarian Carcinoma SKOV-3 Cells. J. Cell. Biochem. 2017, 118, 4468–4478. [Google Scholar] [CrossRef] [PubMed]
- Robak, T.; Błoński, J.; Urbańska-Ryś, H.; Błasińska-Morawiec, M.; Skotnicki, A. 2-Chlorodeoxyadenosine (Cladribine) in the treatment of patients with chronic lymphocytic leukemia 55 years old and younger. Leukemia 1999, 13, 518–523. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sandoval, A.; Consoli, U.; Plunkett, W. Fludarabine-mediated inhibition of nucleotide excision repair induces apoptosis in quiescent human lymphocytes. Clin. Cancer Res. 1996, 2, 1731–1741. [Google Scholar]
- Bontemps, F.; Delacauw, A.; Cardoen, S.; Van Den Neste, E.; Van Den Berghe, G. Metabolism and cytotoxic effects of 2-chloroadenine, the major catabolite of 2-chloro-2′-deoxyadenosine. Biochem. Pharmacol. 2000, 59, 1237–1243. [Google Scholar] [CrossRef]
- Bednarska-Szczepaniak, K.; Krzyżanowski, D.; Klink, M.; Nowak, M. Adenosine Analogues as Opposite Modulators of the Cisplatin Resistance of Ovarian Cancer Cells. Anti-Cancer Agents Med. Chem. 2019, 19, 473–486. [Google Scholar] [CrossRef] [PubMed]
- Leśnikowski, Z.J. What are the current challenges with the application of boron clusters to drug design? Expert Opin. Drug Discov. 2020, 16, 481–483. [Google Scholar] [CrossRef] [PubMed]
- Stockmann, P.; Gozzi, M.; Kuhnert, R.; Sárosi, M.B.; Hey-Hawkins, E. New keys for old locks: Carborane-containing drugs as platforms for mechanism-based therapies. Chem. Soc. Rev. 2019, 48, 3497–3512. [Google Scholar] [CrossRef] [PubMed]
- Cígler, P.; Kozísek, M.; Rezácová, P.; Brynda, J.; Otwinowski, Z.; Pokorná, J.; Plešek, J.; Grüner, B.; Dolecková-Maresová, L.; Mása, M.; et al. From nonpeptide toward noncarbon protease inhibitors: Metallacarboranes as specific and potent inhibitors of HIV protease. Proc. Natl. Acad. Sci. USA 2005, 102, 15394–15399. [Google Scholar] [CrossRef]
- Grüner, B.; Brynda, J.; Das, V.; Šícha, V.; Štěpánková, J.; Nekvinda, J.; Holub, J.; Pospíšilová, K.; Fábry, M.; Pachl, P.; et al. Metallacarborane Sulfamides: Unconventional, Specific, and Highly Selective Inhibitors of Carbonic Anhydrase IX. J. Med. Chem. 2019, 62, 9560–9575. [Google Scholar] [CrossRef]
- Grüner, B.; Kugler, M.; El Anwar, S.; Holub, J.; Nekvinda, J.; Bavol, D.; Růžičková, Z.; Pospíšilová, K.; Fábry, M.; Král, V.; et al. Cobalt Bis(dicarbollide) Alkylsulfonamides: Potent and Highly Selective Inhibitors of Tumor Specific Carbonic Anhydrase IX. ChemPlusChem. 2020, 86, 351. [Google Scholar] [CrossRef] [PubMed]
- Nekvinda, J.; Różycka, D.; Rykowski, S.; Wyszko, E.; Fedoruk-Wyszomirska, A.; Gurda, D.; Orlicka-Płocka, M.; Giel-Pietraszuk, M.; Kiliszek, A.; Rypniewski, W.; et al. Synthesis of naphthalimide-carborane and metallacarborane conjugates: Anticancer activity, DNA binding ability. Bioorganic Chem. 2020, 94, 103432. [Google Scholar] [CrossRef]
- Olejniczak, A.; Nawrot, B.; Leśnikowski, Z. DNA Modified with Boron–Metal Cluster Complexes [M(C2B9H11)2]—Synthesis, Properties, and Applications. Int. J. Mol. Sci. 2018, 19, 3501. [Google Scholar] [CrossRef]
- Białek-Pietras, M.; Olejniczak, A.B.; Tachikawa, S.; Nakamura, H.; Leśnikowski, Z.J. Towards new boron carriers for boron neutron capture therapy: Metallacarboranes bearing cobalt, iron and chromium and their cholesterol conjugates. Bioorganic Med. Chem. 2013, 21, 1136–1142. [Google Scholar] [CrossRef]
- Byun, Y.; Narayanasamy, S.; Johnsamuel, J.; Bandyopadhyaya, A.K.; Tiwari, R.; Al-Madhoun, A.S.; Barth, R.F.; Eriksson, S.; Tjarks, W. 3-Carboranyl Thymidine Analogues (3CTAs) and Other Boronated Nucleosides for Boron Neutron Capture Therapy. Anticancer Agents Med. Chem. 2006, 6, 127–144. [Google Scholar] [CrossRef] [PubMed]
- Kaniowski, D.; Ebenryter-Olbińska, K.; Sobczak, M.; Wojtczak, B.; Janczak, S.; Leśnikowski, Z.; Nawrot, B. High Boron-loaded DNA-Oligomers as Potential Boron Neutron Capture Therapy and Antisense Oligonucleotide Dual-Action Anticancer Agents. Molecules 2017, 22, 1393. [Google Scholar] [CrossRef]
- Leśnikowski, Z.J.; Paradowska, E.; Olejniczak, A.B.; Studzińska, M.; Seekamp, P.; Schüßler, U.; Gabel, D.; Schinazi, R.F.; Plešek, J. Towards new boron carriers for boron neutron capture therapy: Metallacarboranes and their nucleoside conjugates. Bioorganic Med. Chem. 2005, 13, 4168–4175. [Google Scholar] [CrossRef] [PubMed]
- Adamska-Bartłomiejczyk, A.; Bednarska, K.; Białek-Pietras, M.; Kiliańska, Z.M.; Mieczkowski, A.; Olejniczak, A.B.; Paradowska, E.; Studzińska, M.; Sułowska, Z.; Żołnierczyk, J.D.; et al. Boron Cluster Modifications with Antiviral, Anticancer, and Modulation of Purinergic Receptors’ Activities Based on Nucleoside Structures. In Boron-Based Compounds: Potential and Emerging Applications in Medicine; John Wiley & Sons: Hoboken, NJ, USA, 2018. [Google Scholar]
- Bednarska, K.; Olejniczak, A.B.; Wojtczak, B.A.; Sułowska, Z.; Leśnikowski, Z.J. Adenosine and 2′-Deoxyadenosine Modified with Boron Cluster Pharmacophores as New Classes of Human Blood Platelet Function Modulators. ChemMedChem 2010, 5, 749–756. [Google Scholar] [CrossRef] [PubMed]
- Bednarska, K.; Olejniczak, A.B.; Piskala, A.; Klink, M.; Sulowska, Z.; Lesnikowski, Z.J. Effect of adenosine modified with a boron cluster pharmacophore on reactive oxygen species production by human neutrophils. Bioorganic Med. Chem. 2012, 20, 6621–6629. [Google Scholar] [CrossRef]
- Bednarska, K.; Olejniczak, A.B.; Klink, M.; Sułowska, Z.; Leśnikowski, Z.J. Modulation of human neutrophil activity by adenosine modified with a carborane pharmacophore. Bioorganic Med. Chem. Lett. 2014, 24, 3073–3078. [Google Scholar] [CrossRef] [PubMed]
- Olejniczak, A.B.; Semenuk, A.; Kwiatkowski, M.; Lesnikowski, Z.J. Synthesis of adenosine containing carborane modification. J. Organomet. Chem. 2003, 680, 124–126. [Google Scholar] [CrossRef]
- Olejniczak, A.B.; Plešek, J.; Leśnikowski, Z.J. Nucleoside-Metallacarborane Conjugates for Base-Specific Metal Labeling of DNA. Chem. Eur. J. 2006, 13, 311–318. [Google Scholar] [CrossRef]
- Olejniczak, A.B.; Grűner, B.; Šicha, V.; Broniarek, S.; Lesnikowski, Z.J. Metallacarboranes as Labels for Multipotential Electrochemical Coding of DNA. [3-Chromium bis(dicarbollide)](-1)ate and Its Nucleoside Conjugates. Electroanalysis 2009, 21, 501–506. [Google Scholar] [CrossRef]
- Olejniczak, A.B.; Mucha, P.; Grüner, B.; Lesnikowski, Z.J. DNA-Dinucleotides Bearing a 3′,3′-Cobalt- or 3′,3′-Iron-1,2,1′,2′-dicarbollide Complex. Organometallics 2007, 26, 3272–3274. [Google Scholar] [CrossRef]
- Wojtczak, B.A.; Andrysiak, A.; Grüner, B.; Lesnikowski, Z.J. “Chemical Ligation”: A Versatile Method for Nucleoside Modification with Boron Clusters. Chem. Eur. J. 2008, 14, 10675–10682. [Google Scholar] [CrossRef] [PubMed]
- Wojtczak, B.A.; Olejniczak, A.B.; Lesnikowski, Z.J. Nucleoside Modification with Boron Clusters and Their Metal Complexes. Curr. Protoc. Nucleic Acid Chem. 2009, 36, 4–37. [Google Scholar] [CrossRef]
- Hamilton, T.C.; Young, R.C.; McKoy, W.M.; Grotzinger, K.R.; Green, J.A.; Chu, E.W.; Whang-Peng, J.; Rogan, A.M.; Green, W.R.; Ozols, R.F. Characterization of a Human Ovarian Carcinoma Cell Line (NIH:OVCAR-3) with Androgen and Estrogen Receptors. Cancer Res. 1983, 43, 5379–5389. [Google Scholar]
- Egawa-Takata, T.; Endo, H.; Fujita, M.; Ueda, Y.; Miyatake, T.; Okuyama, H.; Yoshino, K.; Kamiura, S.; Enomoto, T.; Kimura, T.; et al. Early reduction of glucose uptake after cisplatin treatment is a marker of cisplatin sensitivity in ovarian cancer. Cancer Sci. 2010, 101, 2171–2178. [Google Scholar] [CrossRef]
- Bieging, K.T.; Mello, S.S.; Attardi, L.D. Unravelling mechanisms of p53-mediated tumour suppression. Nat. Rev. Cancer 2014, 14, 359–370. [Google Scholar] [CrossRef]
- Brooks, C.L.; Gu, W. The impact of acetylation and deacetylation on the p53 pathway. Protein Cell 2011, 2, 456–462. [Google Scholar] [CrossRef]
- Xia, C.; Meng, Q.; Liu, L.-Z.; Rojanasakul, Y.; Wang, X.-R.; Jiang, B.-H. Reactive Oxygen Species Regulate Angiogenesis and Tumor Growth through Vascular Endothelial Growth Factor. Cancer Res. 2007, 67, 10823–10830. [Google Scholar] [CrossRef]
- Rushworth, S.A.; Zaitseva, L.; Murray, M.Y.; Shah, N.M.; Bowles, K.M.; MacEwan, D.J. The high Nrf2 expression in human acute myeloid leukemia is driven by NF-κB and underlies its chemo-resistance. Blood 2012, 120, 5188–5198. [Google Scholar] [CrossRef]
- Chowanadisai, W.; Messerli, S.M.; Miller, D.H.; Medina, J.E.; Hamilton, J.W.; Messerli, M.A.; Brodsky, A.S. Cisplatin Resistant Spheroids Model Clinically Relevant Survival Mechanisms in Ovarian Tumors. PLoS ONE 2016, 11, e0151089. [Google Scholar] [CrossRef] [PubMed]
- Weiswald, L.-B.; Bellet, D.; Dangles-Marie, V. Spherical Cancer Models in Tumor Biology. Neoplasia 2015, 17, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Plešek, J.; Heřmánek, S.; Franken, A.; Císařová, I.; Nachtigal, C. Dimethyl Sulfate Induced Nucleophilic Substitution of the [Bis(1,2-dicarbollido)-3-cobalt(1-)]ate Ion. Syntheses, Properties and Structures of Its 8,8′-μ-Sulfato, 8-Phenyland 8-Dioxane Derivatives. Collect. Czechoslov. Chem. Commun. 1997, 62, 47–56. [Google Scholar] [CrossRef]
- Plešek, J.; Grüner, B.; Macháček, J.; Císařová, I.; Čáslavský, J. 8-Dioxane ferra(III) bis(dicarbollide): A paramagnetic functional molecule as versatile building block for introduction of a Fe(III) centre into organic molecules. J. Organomet. Chem. 2007, 692, 4801–4804. [Google Scholar] [CrossRef]
- Forward, J.M.; Michael, D.; Mingos, P.; Müller, T.E.; Williams, D.J.; Yan, Y.-K. Synthesis and structural characterization of metallacarborane sandwich salts with tetrathiafulvalene (ttf) [M(C2B9H11)2][ttf] (M = Cr, Fe, Ni). J. Organomet. Chem. 1994, 467, 207–216. [Google Scholar] [CrossRef]
- Čižmár, E.; Šoltésová, D.; Kazheva, O.N.; Alexandrov, G.G.; Kravchenko, A.V.; Chekulaeva, L.A.; Kosenko, I.D.; Sivaev, I.B.; Bregadze, V.I.; Fedorchenko, A.V.; et al. Large magnetic anisotropy of chromium(III) ions in a bis(ethylenedithio)tetrathiafulvalenium salt of chromium bis(dicarbollide), (ET)2[3,3′-Cr(1,2-C2B9H11)2]. Transit. Met. Chem. 2018, 43, 647–655. [Google Scholar] [CrossRef]
- Bandman, M.A.; Knobler, C.B.; Hawthorne, M.F. The synthesis and structural characterization of the first gallacarborane sandwich. Inorg. Chem. 1989, 28, 1204–1206. [Google Scholar] [CrossRef]
- Semioshkin, A.A.; Sivaev, I.B.; Bregadze, V.I. Cyclic oxonium derivatives of polyhedral boron hydrides and their synthetic applications. Dalton Trans. 2008, 8, 977–992. [Google Scholar] [CrossRef] [PubMed]
- Olejniczak, A.; Wojtczak, B.; Lesnikowski, Z.J. 2′-Deoxyadenosine Bearing Hydrophobic Carborane Pharmacophore. Nucleosides Nucleotides Nucleic Acids 2007, 26, 1611–1613. [Google Scholar] [CrossRef]
- Sagi, G.; Otvos, L.; Ikeda, S.; Andrei, G.; Snoeck, R.; De Clercq, E. Synthesis and Antiviral Activities of 8-Alkynyl-, 8-Alkenyl-, and 8-Alkyl-2′-deoxyadenosine Analogs. J. Med. Chem. 1994, 37, 1307–1311. [Google Scholar] [CrossRef] [PubMed]
- Jawalekar, A.M.; Meeuwenoord, N.; Cremers, J.S.G.; Overkleeft, H.S.; van der Marel, G.A.; Rutjes, F.P.J.T.; van Delft, F.L. Conjugation of Nucleosides and Oligonucleotides by [3+2] Cycloaddition. J. Org. Chem. 2007, 73, 287–290. [Google Scholar] [CrossRef]
- Cortez, A.J.; Tudrej, P.; Kujawa, K.A.; Lisowska, K.M. Advances in ovarian cancer therapy. Cancer Chemother. Pharmacol. 2017, 81, 17–38. [Google Scholar] [CrossRef] [PubMed]
- Barth, R.F.; Vicente, M.H.; Harling, O.K.; Kiger, W.; Riley, K.J.; Binns, P.J.; Wagner, F.M.; Suzuki, M.; Aihara, T.; Kato, I.; et al. Current status of boron neutron capture therapy of high grade gliomas and recurrent head and neck cancer. Radiat. Oncol. 2012, 7, 146. [Google Scholar] [CrossRef]
- Grimes, R.N. Carboranes in the chemist’s toolbox. Dalton Trans. 2015, 44, 5939–5956. [Google Scholar] [CrossRef]
- Murphy, N.; McCarthy, E.; Dwyer, R.; Farràs, P. Boron clusters as breast cancer therapeutics. J. Inorg. Biochem. 2021, 218, 111412. [Google Scholar] [CrossRef] [PubMed]
- Tarrés, M.; Canetta, E.; Paul, E.; Forbes, J.; Azzouni, K.; Viñas, C.; Teixidor, F.; Harwood, A.J. Biological interaction of living cells with COSAN-based synthetic vesicles. Chem. Commun. 2014, 50, 3370–3372. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, I.; García-Mendiola, T.; Sato, S.; Pita, M.; Nakamura, H.; Lorenzo, E.; Teixidor, F.; Marques, F.; Viñas, C. Metallacarboranes on the Road to Anticancer Therapies: Cellular Uptake, DNA Interaction, and Biological Evaluation of Cobaltabisdicarbollide [COSAN]–. Chem. A Eur. J. 2018, 24, 17239–17254. [Google Scholar] [CrossRef]
- Rokitskaya, T.I.; Kosenko, I.D.; Sivaev, I.B.; Antonenko, Y.N.; Bregadze, V.I. Fast flip–flop of halogenated cobalt bis(dicarbollide) anion in a lipid bilayer membrane. Phys. Chem. Chem. Phys. 2017, 19, 25122–25128. [Google Scholar] [CrossRef]
- Siddik, Z.H.; Mims, B.; Lozano, G.; Thai, G. Independent Pathways of p53 Induction by Cisplatin and X-Rays in a Cisplatin-resistant Ovarian Tumor Cell Line. Cancer Res. 1998, 4, 698–703. [Google Scholar]
- Fujioka, S.; Niu, J.; Schmidt, C.; Sclabas, G.M.; Peng, B.; Uwagawa, T.; Li, Z.; Evans, D.B.; Abbruzzese, J.L.; Chiao, P.J. NF-κB and AP-1 Connection: Mechanism of NF-κB-Dependent Regulation of AP-1 Activity. Mol. Cell. Biol. 2004, 24, 7806–7819. [Google Scholar] [CrossRef] [PubMed]
- Sodek, K.L.; Ringuette, M.J.; Brown, T.J. Compact spheroid formation by ovarian cancer cells is associated with contractile behavior and an invasive phenotype. Int. J. Cancer 2009, 124, 2060–2070. [Google Scholar] [CrossRef]
- Repetto, G.; del Peso, A.; Zurita, J.L. Neutral red uptake assay for the estimation of cell viability/cytotoxicity. Nat. Protoc. 2008, 3, 1125–1131. [Google Scholar] [CrossRef]
- Klapperstück, T.; Glanz, D.; Klapperstück, M.; Wohlrab, J. Methodological aspects of measuring absolute values of membrane potential in human cells by flow cytometry. Cytom. Part A J. Int. Soc. Adv. Cytom. 2009, 75, 593–608. [Google Scholar] [CrossRef] [PubMed]
- Wagner, W.; Sachrajda, I.; Pułaski, Ł.; Hałatek, T.; Dastych, J. Application of cellular biosensors for analysis of bioactivity associated with airborne particulate matter. Toxicol. Vitr. 2011, 25, 1132–1142. [Google Scholar] [CrossRef] [PubMed]
- Mankiewicz-Boczek, J.; Karwaciak, I.; Ratajewski, M.; Gągała, I.; Jurczak, T.; Zalewski, M.; Pułaski, Ł. Application of cellular biosensors for detection of atypical toxic bioactivity in microcystin-containing cyanobacterial extracts. Aquat. Toxicol. 2015, 168, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.C.; Kania, K.D.; Wijesuriya, H.; Chawla, S.; Sethi, J.K.; Pulaski, L.; Romero, I.A.; Couraud, P.O.; Weksler, B.B.; Hladky, S.B.; et al. Activation of -catenin signalling by GSK-3 inhibition increases P-glycoprotein expression in brain endothelial cells. J. Neurochem. 2008, 106, 1855–1865. [Google Scholar] [CrossRef]
- Sheldrick, G.M. SHELXT–Integrated space-group and crystal-structure determination. Acta Crystallogr. Sect. A Found. Adv. 2015, 71, 3–8. [Google Scholar] [CrossRef]
- Sheldrick, G.M. Crystal structure refinement with SHELXL. Acta Crystallogr. Sect. C Struct. Chem. 2015, 71, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Dolomanov, O.V.; Bourhis, L.J.; Gildea, R.J.; Howard, J.A.K.; Puschmann, H. OLEX2: A complete structure solution, refinement and analysis program. J. Appl. Crystallogr. 2009, 42, 339–341. [Google Scholar] [CrossRef]
- Diamond—Crystal and Molecular Structure Visualization Crystal Impact, Version 4.6.3. H. Putz & K. Brandenburg GbR. Kreuzherrenstr. 102, 53227 Bonn. Germany. 2020. Available online: http://www.Crystalimpact.Com/Diamond (accessed on 21 January 2021).





















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Similarity * % | Cel Line | Cisplatin (μM) | p Value | Resistance Index ** | Carboplatin (μM) | p Value | Resistance Index * |
|---|---|---|---|---|---|---|---|
 | A2780 | 2.8 ± 0.21 | — | — | 51 ± 4.4 | — | — |
| OVCAR-3 | 5.1 ± 0.45 | 0.971 | n.a. | 55 ± 4.4 | 0.945 | n.a. | |
| A2780cis | 15 ± 0.64 | <0.001 | 5.4 | 108 ± 4.9 | <0.001 | 2.1 | |
| SKOV-3 | 21 ± 1.6 | <0.001 | n.a. | 211 ± 11 | <0.001 | n.a. | |
| A2780cisR | 24 ± 0.89 | <0.001 | 8.6 | 162 ± 5.6 | <0.001 | 3.2 | |
| A2780cisKB | 37 ± 2.1 | <0.001 | 13.2 | 230 ± 10 | <0.001 | 4.5 |
| Cell Line | Cisplatin | Compound 23 | Spheroid Structure | ||
|---|---|---|---|---|---|
| Adherent | Spheroids | Adherent | Spheroids | ||
| OVCAR-3 | 40 | 100 | 49 | 36 | compact |
| A2780 | 44 | 49 | 52 | 81 | loose |
| A2780cis | 82 | 88 | 58 | 91 | loose |
| A2780cisR | 89 | 89 | 49 | 83 | loose |
| SKOV-3 | 82 | 81 | 24 | 89 | loose |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bednarska-Szczepaniak, K.; Przelazły, E.; Kania, K.D.; Szwed, M.; Litecká, M.; Grűner, B.; Leśnikowski, Z.J. Interaction of Adenosine, Modified Using Carborane Clusters, with Ovarian Cancer Cells: A New Anticancer Approach against Chemoresistance. Cancers 2021, 13, 3855. https://doi.org/10.3390/cancers13153855
Bednarska-Szczepaniak K, Przelazły E, Kania KD, Szwed M, Litecká M, Grűner B, Leśnikowski ZJ. Interaction of Adenosine, Modified Using Carborane Clusters, with Ovarian Cancer Cells: A New Anticancer Approach against Chemoresistance. Cancers. 2021; 13(15):3855. https://doi.org/10.3390/cancers13153855
Chicago/Turabian StyleBednarska-Szczepaniak, Katarzyna, Ewelina Przelazły, Katarzyna Dominika Kania, Marzena Szwed, Miroslava Litecká, Bohumír Grűner, and Zbigniew J. Leśnikowski. 2021. "Interaction of Adenosine, Modified Using Carborane Clusters, with Ovarian Cancer Cells: A New Anticancer Approach against Chemoresistance" Cancers 13, no. 15: 3855. https://doi.org/10.3390/cancers13153855
APA StyleBednarska-Szczepaniak, K., Przelazły, E., Kania, K. D., Szwed, M., Litecká, M., Grűner, B., & Leśnikowski, Z. J. (2021). Interaction of Adenosine, Modified Using Carborane Clusters, with Ovarian Cancer Cells: A New Anticancer Approach against Chemoresistance. Cancers, 13(15), 3855. https://doi.org/10.3390/cancers13153855