
The Human TOR Signaling Regulator Is the Key Indicator of Liver Cancer Patients’ Overall Survival: TIPRL/LC3/CD133/CD44 as Potential Biomarkers for Early Liver Cancers

Abstract
Simple Summary
Abstract
1. Introduction
2. Results
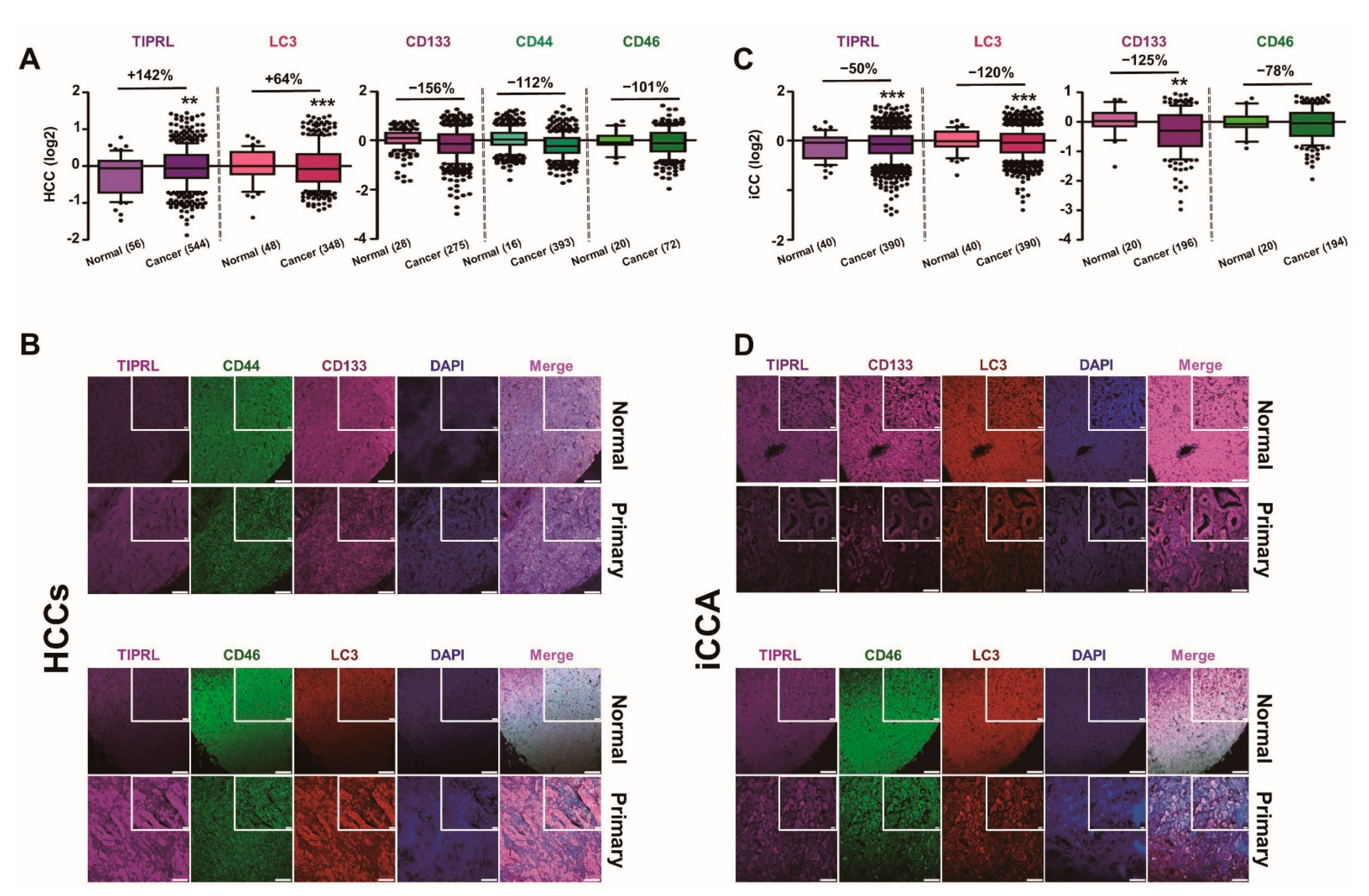
2.1. Inverse Expression of TIPRL and LC3 in HCCs and iCCA
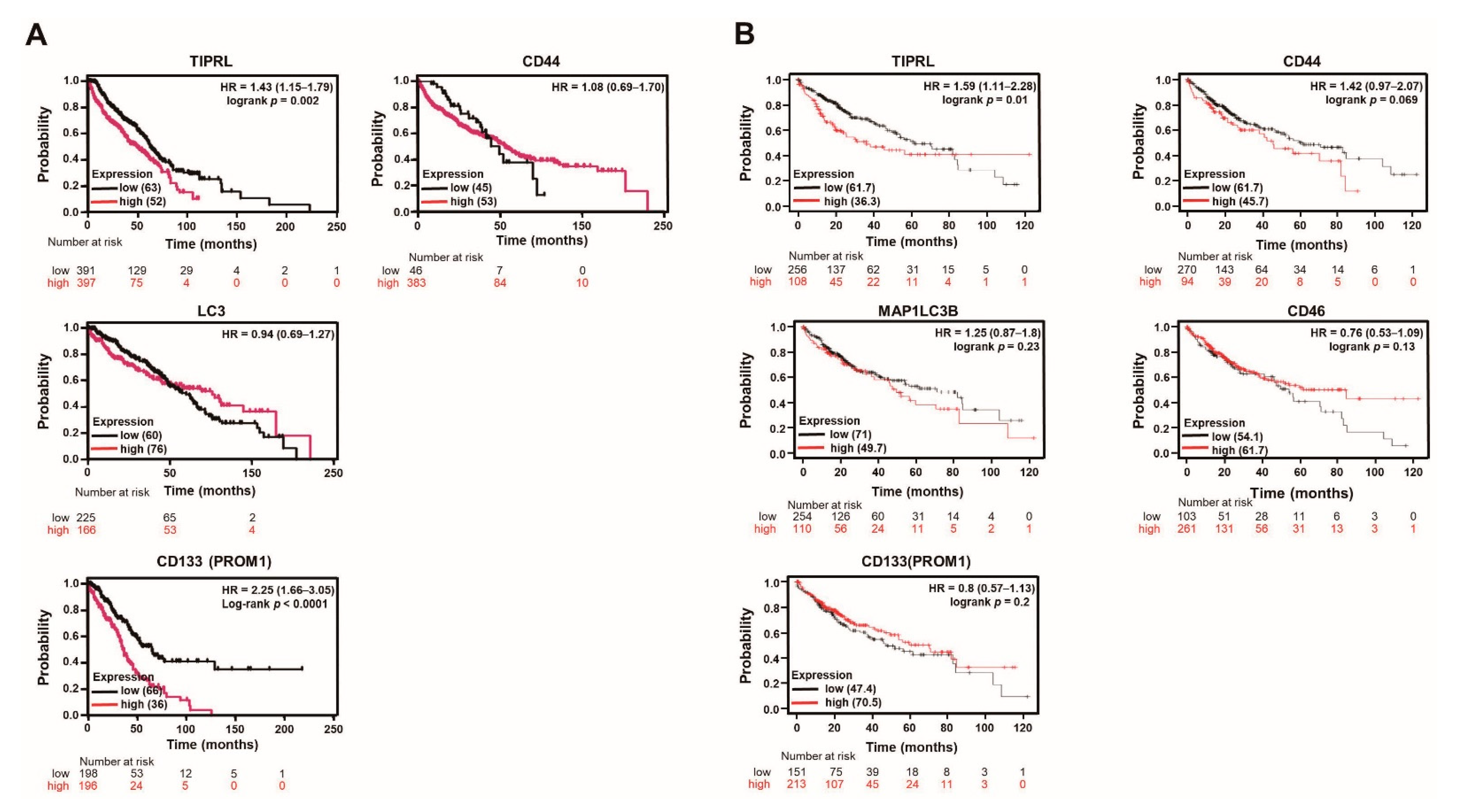
2.2. The Significant Association between TIPRL and Liver Disease Patients’ Survival
2.3. The Significant Predictive Ability of TIPRL on Liver Disease Patients
2.4. TIPRL as a Key Player for Liver Cancer Survival
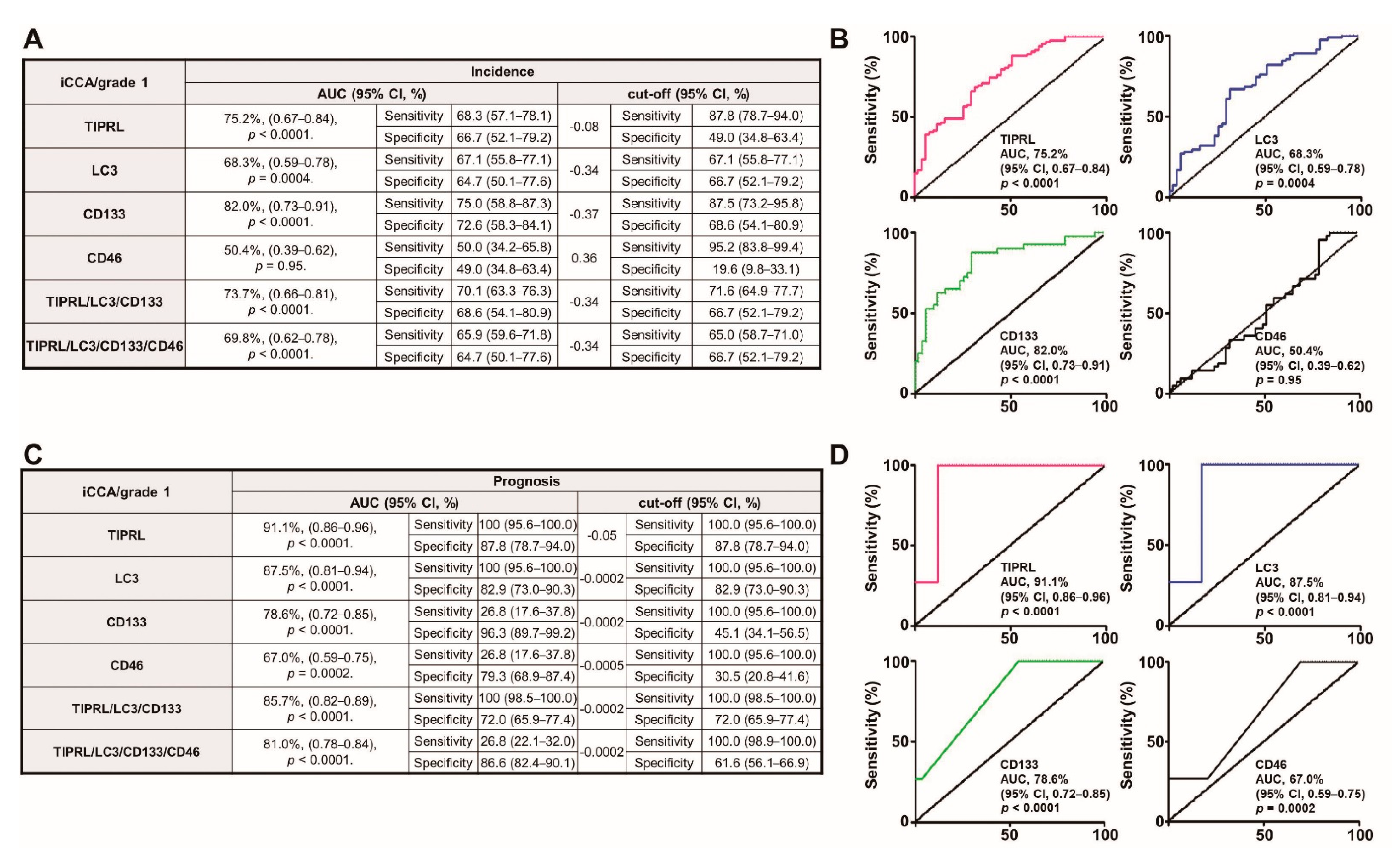
2.5. TIPRL, LC3, CD133, and CD44 as Liver Cancer Biomarkers for Early Diagnosis and Prognosis
3. Discussion
4. Materials and Methods
4.1. Patients’ Tissues and Information
4.2. Immunohistochemistry and Histopathology
4.3. Statistical Analysis
4.4. Cell Culture and Small-Interfering RNAs (siRNAs) Transfection
4.5. Cell Proliferation and Survival Assays (MTT Assay)
4.6. Reverse-Transcriptase Quantitative Polymerase-Chain Reaction
4.7. Tumorspheres Formation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| TIPRL | Human TOR signaling regulator |
| LC3 | Microtubule-associated light chain 3 |
| CD133 | Prominin-1 |
| CD44 | Cluster of differentiation 44 |
| CD46 | Cluster of differentiation 46 |
| ROC | Receiver-operating characteristic |
| HCCs | Hepatocellular carcinomas |
| iCCA | Intrahepatic cholangiocarcinomas |
| OS | Overall survival |
| DSS | Disease-specific survival |
| HR | Hazard ratio |
| CI | 95% confidence ratio |
| HR | Hazard ratio |
References
- World Health Organization. Available online: https://www.who.int/news-room/fact-sheets/detail/cancer (accessed on 22 April 2019).
- Anstee, Q.M.; Reeves, H.L.; Kotsiliti, E.; Govaere, O.; Heikenwalder, M. From NASH to HCC: Current concepts and future challenges. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 411–428. [Google Scholar] [CrossRef]
- Song, I.S.; Jun, S.Y.; Na, H.J.; Kim, H.T.; Jung, S.Y.; Ha, G.H.; Park, Y.H.; Long, L.Z.; Yu, D.Y.; Kim, J.M.; et al. Inhibition of MKK7-JNK by the TOR signaling pathway regulator-like protein contributes to resistance of HCC cells to TRAIL-induced apoptosis. Gastroenterology 2012, 143, 1341–1351. [Google Scholar] [CrossRef]
- Jacinto, E.; Guo, B.; Arndt, K.T.; Schmelzle, T.; Hall, M.N. TIP4 interacts with TAP42 and negatively regulates the TOR signaling pathway. Mol. Cell 2001, 8, 1017–1026. [Google Scholar] [CrossRef]
- McConnel, J.L.; Gomez, R.J.; McCorvey, L.R.; Law, B.K.; Wadzinski, B.E. Identification of a PP2A-interacting protein that functions as a negative regulator of phosphatase activity in the ATM/ATR signaling pathway. Oncogene 2007, 26, 6021–6030. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, A.; Tanimura-Ito, K.; Oshiro, N.; Eguchi, S.; Miyamoto, T.; Momonami, A.; Kamada, S.; Yonezawa, K.; Kikkawa, U. A positive role of mammalian Tip41-like protein, TIPRL, in the amino-acid dependent mTORC1-signaling pathway through interaction with PP2A. FEBS Lett. 2013, 587, 2924–2929. [Google Scholar] [CrossRef] [PubMed]
- Jeon, S.J.; Ahn, J.H.; Halder, D.; Cho, H.S.; Lim, J.H.; Jun, S.Y.; Lee, J.J.; Yoon, J.Y.; Choi, M.H.; Jung, C.R.; et al. TIPRL potentiates survival of lung cancer by inducing autophagy through the eif2α-ATF4 pathway. Cell Death Dis. 2019, 10, 959. [Google Scholar] [CrossRef] [PubMed]
- Jun, S.Y.; Jeon, S.J.; Yoon, J.Y.; Lee, J.J.; Yoon, H.R.; Choi, M.H.; Halder, D.; Lee, K.H.; Kim, N.S. The positive correlation of TIPRL with LC3 and CD133 contributes to cancer aggressiveness: Potential biomarkers for early liver cancer. Sci. Rep. 2019, 9, 16802. [Google Scholar] [CrossRef]
- Dhar, D.; Antonucci, L.; Nakagawa, H.; Kim, J.Y.; Glitzner, E.; Caruso, S.; Shalapour, S.; Yang, L.; Valasek, M.A.; Lee, S.; et al. Liver cancer initiation requires p53 inhibition by CD44-enhanced growth factor signaling. Cancer Cell 2018, 33, 1061–1077. [Google Scholar] [CrossRef]
- Hou, Y.; Zou, Q.; Ge, R.; Shen, F.; Wang, Y. The critical role of CD133+CD44+/high tumor cells in hematogenous metastasis of liver cancers. Cell Res. 2012, 22, 259–272. [Google Scholar] [CrossRef]
- Qiu, L.; Li, H.; Fu, S.; Chen, X.; Lu, L. Surface markers of liver cancer stem cells and innovative targeted-therapy strategies for HCC. Oncol. Lett. 2018, 15, 2039–2048. [Google Scholar] [CrossRef]
- Ma, S.; Chan, K.W.; Hu, L.; Lee, T.K.W.; Wo, J.Y.H.; Ng, I.O.L.; Zheng, B.J.; Guan, X.Y. Identification and characterization of tumorigenic liver cancer stem/progenitor cells. Gastroenterology 2007, 132, 2542–2556. [Google Scholar] [CrossRef]
- Chen, H.; Luo, Z.; Dong, L.; Tan, Y.; Yang, J.; Feng, G.; Wu, M.; Li, Z.; Wang, H. CD133/prominin-1-mediated autophagy and glucose uptake beneficial for hepatoma cell survival. PLoS ONE 2013, 8, e56878. [Google Scholar] [CrossRef]
- Chun, Y.; Kim, J. Autophagy: An essential degradation program for cellular homeostasis and life. Cells 2018, 7, 278. [Google Scholar] [CrossRef] [PubMed]
- Comel, A.; Sorrentino, G.; Capaci, V.; Sal, G.D. The cytoplasmic side of p53’s oncosuppressive activities. FEBS Lett. 2014, 588, 2600–2609. [Google Scholar] [CrossRef]
- Li, J.; Hu, S.B.; Wang, L.Y.; Zhang, X.; Zhou, X.; Yang, B.; Li, J.H.; Xiong, J.; Liu, N.; Li, Y.; et al. Autophagy-dependent generation of Axin2+ cancer stem-like cells promotes hepatocarcinogenesis in liver cirrhosis. Oncogene 2017, 36, 6725–6737. [Google Scholar] [CrossRef] [PubMed]
- National Cancer Institute. Available online: https://www.cancer.gov/publications/dictionaries/cancer-terms/def/disease-specific-survival-rate (accessed on 8 November 2020).
- Nazio, F.; Bordi, M.; Cianfanelli, V.; Locatelli, F.; Cecconi, F. Autophagy and cancer stem cells: Molecular mechanisms and therapeutic applications. Cell Death Differ. 2019, 26, 690–702. [Google Scholar] [CrossRef] [PubMed]
- Safari, S.; Baratloo, A.; Elfil, M.; Negida, A. Evidence based emergency medicine; part 5 receiver operating curve and area under the curve. Emergency 2016, 4, 111–113. [Google Scholar]
- Hawkes, N. Cancer survival data emphasis importance of early diagnosis. BMJ 2019, 364, I408. [Google Scholar] [CrossRef] [PubMed]
- Cancer.Net. Doctor-Approved Patient Information from ACSO. Available online: https://www.cancer.net/cancer-types/liver-cancer/statistics (accessed on 15 December 2020).
- Sourbier, C. Plasma HSP90α and liver cancer: A potential biomarker? EBioMedicine 2017, 25, 7–8. [Google Scholar] [CrossRef]
- Sia, D.; Villanueva, A.; Friedman, S.; LIovet, J.M. Liver cancer cell of origin, molecular class, and effects on patient prognosis. Gastroenterology 2017, 152, 745–761. [Google Scholar] [CrossRef]
- Sekiya, S.; Suzuki, A. Intrahepatic cholangiocarcinoma can arise from Notch-mediated conversion of hepatocytes. J. Clin. Investig. 2012, 122, 3914–3918. [Google Scholar] [CrossRef]
- Fan, B.; Malato, Y.; Calvisi, D.F.; Nagvi, S.; Rzumilava, N.; Ribback, S.; Gores, G.J.; Dombrowski, F.D.; Evert, M.; Chen, X.; et al. Cholangiocarcinomas can originate from hepatocytes in mice. J. Clin. Investig. 2012, 122, 2911–2915. [Google Scholar] [CrossRef]
- Liovet, J.M.; Bruix, J.F. Intention-to-treat analysis of surgical treatment for early hepatocellular carcinoma: Resection versus transplantation. Hepatology 1999, 30, 1434–1440. [Google Scholar] [CrossRef]
- Rhee, H.; Park, J.H.; Park, Y.N. Update on pathologic and radiologic diagnosis of combined hepatocellular-cholangiocarcinoma. J. Liver Cancer 2021, 21, 12–24. [Google Scholar] [CrossRef]
- Brunt, E.M. Histological assessment of nonalcoholic fatty liver disease in adults and children. Clin. Liver Dis. 2012, 1, 108–111. [Google Scholar] [CrossRef]
- Singh, G.; Yoshida, E.M.; Rathi, S.; Marquez, V.; Kim, P.; Erb, S.R.; Salh, B.S. Biomarkers for hepatocellular cancer. World J. Hepatol. 2020, 12, 558–573. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A. Training | B. Validation | ||||
|---|---|---|---|---|---|
| Variables | Overall Survival | Variables | Overall Survival | ||
| Univariate | Univariate | ||||
| HR (95 % CI) | p-values | HR (95 % CI) | p-values | ||
| Age | Age | ||||
| ≤50 | ref | ≤50 | ref | ||
| >50 | 0.57 (0.40–0.80) | 0.001 | >50 | 0.88 (0.64–1.22) | 0.44 |
| Sex | Sex | ||||
| Female | ref | Female | ref | ||
| Male | 0.58 (0.40–0.83) | 0.003 | Male | 3.0 × 10−7 (5.47 × 10−14-1.65) | 0.96 |
| Subtypes | Subtypes | ||||
| Hepatocellular carcinoma | 0.79 (0.47–1.33) | 0.37 | Hepatocellular carcinoma | 4.72 (1.72–13.01) | 0.003 |
| Chronic hepatitis | 0.57 (0.33–0.97) | 0.04 | Intrahepatic cholangiocarcinoma | 3.91 (1.44–10.63) | 0.008 |
| Cirrhosis | 1.72 (0.96–3.08) | 0.07 | Mixed carcinoma | 1.93 (1.47–16.53) | 0.01 |
| Hepatic steatosis | 2.38 (1.18–4.79) | 0.02 | Carcinoid | 55.72 (12.89–240.92) | 7.36 × 10−8 |
| Liver cell/tissue degeneration, Inflammation of portal area | NA | Adenosquamous carcinoma | 4.30 (1.07–17.39) | 0.04 | |
| Normal liver tissue | ref | Cancer adjacent liver tissue | ref | ||
| TNM | TNM | ||||
| T1N0M0 | 2.13 (1.49–3.05) | 3.83 × 10−5 | T2N0M0 | 2.84 (1.06–7.65) | 0.04 |
| T2N0M0 | 1.24 (0.97–1.58) | 0.08 | T2N1M0 | 2.32 (1.09–4.92) | 0.03 |
| T2N1M0 | NA | T3N0M0 | 2.28 (0.68–7.67) | 0.18 | |
| T2N0M1 | 8.5 × 10−7 (0.0-Inf) | 0.99 | T3N1M0 | 1.95 (0.95–4.02) | 0.07 |
| T3N0M0 | 0.38 (0.24–0.61) | 5.43 × 10−5 | T3N0M1 | 5.12 (1.89–13.88) | 0.001 |
| T4N0M0 | NA | T3N1M1 | 3.52 (0.44–28.29) | 0.24 | |
| Grade | T4N1M0 | 1.36 (0.41–4.56) | 0.62 | ||
| Grade1 | 0.32 (0.19–0.53) | 1.58 × 10−5 | T4N1M1 | 3.62 (1.24–10.59) | 0.02 |
| Grade 1-2 | 0.28 (0.16–0.47) | 1.47 × 10−6 | Grade | ||
| Grade2 | 0.19 (0.13–0.29) | 1.89 × 10−15 | Grade1 | 0.89 (0.52–1.52) | 0.67 |
| Grade 2-3 | 0.11 (0.06–0.19) | 3.06 × 10−16 | Grade2 | 0.92 (0.62–1.37) | 0.67 |
| Grade3 | NA | Grade3 | 1.84 (1.21–2.80) | 0.005 | |
| Stage | Stage | ||||
| stage1 | 0.37 (0.18–0.78) | 0.009 | stage1 | NA | |
| stage2 | 0.20 (0.09–0.41) | 1.38 × 10−5 | stage2 | 2.76 (1.03–7.42) | 0.04 |
| stage3 | 0.02 (0.009–0.06) | <2 × 10−16 | stage3 | 2.17 (0.65–7.30) | 0.21 |
| stage4 | 7.844 × 10−8 (0.0-Inf) | 0.99 | stage4 | 2.11 (1.03–4.30) | 0.04 |
| Markers | Markers | ||||
| TIPRL | 1.36 (1.04–1.78) | 0.02 | TIPRL | 1.53 (1.26–1.87) | 2.46 × 10−5 |
| LC3 | 0.60 (0.42–0.86) | 0.005 | LC3 | 1.04 (0.83–1.31) | 0.72 |
| CD133 | 1.50 (0.96–2.34) | 0.07 | CD133 | 1.52 (1.01–2.10) | 0.01 |
| CD44 | 1.21 (0.91–1.61) | 0.19 | CD46 | 0.92 (0.63–1.34) | 0.67 |
| A. Training | B. Validation | ||||
|---|---|---|---|---|---|
| Variables | Overall Survival | Variables | Overall Survival | ||
| Univariate | Univariate | ||||
| HR (95 % CI) | p-values | HR (95 % CI) | p-values | ||
| Subtypes | Subtypes | ||||
| Hepatocellular carcinoma | 0.21 (0.10–0.41) | 8.31 × 10−6 | Hepatocellular carcinoma | 6.89 (2.18–21.79) | 0.001 |
| Chronic hepatitis | 0.43 (0.25–0.74) | 0.002 | Intrahepatic cholangiocarcinoma | 5.25 (1.74–15.85) | 0.003 |
| Cirrhosis | 1.91 (1.05–3.47) | 0.04 | Mixed carcinoma | 5.13 (1.53–17.23) | 0.008 |
| Hepatic steatosis | 2.32 (1.14–4.71) | 0.02 | Carcinoid | 59.86 (13.81–259.43) | 4.53 × 10−8 |
| Liver cell/tissue degeneration, Inflammation of portal area | NA | NA | Adenosquamous carcinoma | 4.46 (1.10–18.08) | 0.04 |
| Normal liver tissue | ref | Cancer adjacent liver tissue | ref | ||
| TNM | TNM | ||||
| T1N0M0 | 5.68 (3.10–10.40) | 1.88 × 10−8 | T2N0M0 | 2.25 (0.83–6.11) | 0.11 |
| T2N0M0 | 3.84 (2.29–6.45) | 3.64 × 10−7 | T2N1M0 | 2.64 (1.22–5.71) | 0.01 |
| T2N1M0 | NA | T3N0M0 | 3.04 (0.86–10.76) | 0.08 | |
| T2N0M1 | 2.2 × 10−6 (0.0-Inf) | 0.99 | T3N1M0 | 2.10 (1.01–4.37) | 0.05 |
| T3N0M0 | NA | T3N0M1 | 7.71 (2.79–21.29) | 8.13 × 10−5 | |
| T4N0M0 | NA | T3N1M1 | 1.70 (0.21–13.70) | 0.62 | |
| T4N1M0 | 1.49 (0.44–5.04) | 0.52 | |||
| T4N1M1 | 4.15 (1.37–12.55) | 0.01 | |||
| Grade | |||||
| Grade1 | 0.63 (0.33–1.21) | 0.17 | |||
| Grade2 | 0.57 (0.32–1.01) | 0.06 | |||
| Grade3 | 1.22 (0.69–2.16) | 0.50 | |||
| Stage | |||||
| stage1 | NA | ||||
| stage2 | 2.13 (0.78–5.78) | 0.14 | |||
| stage3 | 2.67 (0.76–9.39) | 0.13 | |||
| stage4 | 2.27 (1.10–4.69) | 0.03 | |||
| Markers | Markers | ||||
| TIPRL | 2.06 (1.52–2.79) | 2.77 × 10−6 | TIPRL | 14.65 (8.34–25.74) | <2 × 10−16 |
| LC3 | 0.29 (0.17–0.51) | 1.60 × 10−5 | LC3 | 0.03 (0.02–0.07) | <2 × 10−16 |
| CD133 | 0.63 (0.37–1.06) | 0.08 | CD133 | 3.56 (2.17–5.84) | 4.70 × 10−7 |
| CD44 | 2.23 (1.41–3.51) | 0.0006 | CD46 | 2.13 (1.47–3.08) | 6.16 × 10−5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jun, S.Y.; Yoon, H.R.; Yoon, J.-Y.; Jeon, S.-J.; Lee, J.-J.; Halder, D.; Kim, J.-M.; Kim, N.-S. The Human TOR Signaling Regulator Is the Key Indicator of Liver Cancer Patients’ Overall Survival: TIPRL/LC3/CD133/CD44 as Potential Biomarkers for Early Liver Cancers. Cancers 2021, 13, 2925. https://doi.org/10.3390/cancers13122925
Jun SY, Yoon HR, Yoon J-Y, Jeon S-J, Lee J-J, Halder D, Kim J-M, Kim N-S. The Human TOR Signaling Regulator Is the Key Indicator of Liver Cancer Patients’ Overall Survival: TIPRL/LC3/CD133/CD44 as Potential Biomarkers for Early Liver Cancers. Cancers. 2021; 13(12):2925. https://doi.org/10.3390/cancers13122925
Chicago/Turabian StyleJun, Soo Young, Hyang Ran Yoon, Ji-Yong Yoon, Su-Jin Jeon, Jeong-Ju Lee, Debasish Halder, Jin-Man Kim, and Nam-Soon Kim. 2021. "The Human TOR Signaling Regulator Is the Key Indicator of Liver Cancer Patients’ Overall Survival: TIPRL/LC3/CD133/CD44 as Potential Biomarkers for Early Liver Cancers" Cancers 13, no. 12: 2925. https://doi.org/10.3390/cancers13122925
APA StyleJun, S. Y., Yoon, H. R., Yoon, J.-Y., Jeon, S.-J., Lee, J.-J., Halder, D., Kim, J.-M., & Kim, N.-S. (2021). The Human TOR Signaling Regulator Is the Key Indicator of Liver Cancer Patients’ Overall Survival: TIPRL/LC3/CD133/CD44 as Potential Biomarkers for Early Liver Cancers. Cancers, 13(12), 2925. https://doi.org/10.3390/cancers13122925