
CX3CL1 Overexpression Prevents the Formation of Lung Metastases in Trastuzumab-Treated MDA-MB-453-Based Humanized Tumor Mice (HTM)

 ,
, 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. BC Cell Lines and Treatments
2.2. Western Blotting
2.3. Mice
2.4. Ethic Statements
2.5. Immunohistochemistry
2.6. Flow Cytometry Analysis
- (a)
- Tumor cell phenotyping: Samples were stained using the following antibodies: anti-HER2-FITC (clone 24D2, BioLegend), anti-CD47-PE (clone B6H12, BD Biosciences), anti-CD44 Pe-Cy7 (clone G44-26, BD Biosciences), anti-c-MET APC (clone # 95106, R&D Systems), anti-CD24 APC Vio770 (clone 32D12, Miltenyi Biotec, Bergisch Gladbach, Germany) anti-CD45 BV510 (clone H130, BioLegend, San Diego, CA, USA) and anti-EPCAM BV421 (clone 9C4, BioLegend).
- (b)
- Human immune cell reconstitution in spleen and tumor were analyzed with anti-CD3-FITC (clone UCHT1), anti-CD19-PE (clone HIB19) and anti-CD45-APC (clone HI30) from BD Bioscience and anti-CD33-PerCPCy5.5 (clone WM53) from BioLegend.
- (c)
- T cell phenotyping was analyzed by anti-CD3-FITC (clone UCHT1, BD Bioscience), anti-CD127-PE (clone A019D5, BioLegend), anti-CD27-PeCy7 (clone O323, eBioscience (Thermo Fisher Scientific), Darmstadt, Germany), anti-PD-1-AF647 (clone EH12.2H7, BioLegend), anti-CD4-APC-H7 (clone SK3, BD Bioscience), anti-CD8a-BV510 (clone RPA-T8, BioLegend) and anti-CD45RA-BV421 (clone HI100, BioLegend)
- (d)
- NK cells were characterized by anti-NKp46-APC (clone 9-E2, BD Bioscience), anti-CD16-PE (clone 3G8, BioLegend) and anti-CD56-Horizon™V450 (clone B159, BD Bioscience).
- (e)
- CX3CR3 expression on immune cells and tumor cells were analyzed by anti-HER2-FITC (clone 24D2, BioLegend), anti-CD8-PE (clone HIT8a, BD Bioscience), anti-CD45-APC (clone HI30, BD Bioscience), anti-CXCR3-BV421 (clone 2A9-1, BioLegend), anti-CD4-APC-H7 (clone SK3, BD Bioscience), anti-CD19-BV510 (clone HIB19, BioLegend) and anti-NKp46-APC (clone 9-E2, BD Bioscience). CX3CR1 expression on tumor cells were performed by anti-CXCR3-BV421 (clone 2A9-1, BioLegend).
- (f)
- Proliferation assessment: Cells were harvested by trypsinization and washed twice with PBS, followed by fixation and permeabilization with cooled MeOH (70%) overnight. Afterward the cells were washed and incubated for 20 min in the presence of RNase (Roche Molecular Systems) at 37 °C and finally stained with 1 μg/mL DAPI prior to analysis. Cell doublets, aggregates and ¬debris were excluded by pulse processing and DNA histograms of the gated population were plotted on a linear scale. Cell cycle fractions (% of cells in G0/G1-, S- and G2/M-phase) were analyzed using the ModFit LT 3.2 software (Verity Software House, Topsham, ME, USA).
- (g)
- Annexin V-FITC/DAPI Assay: Cells were cultured for 72 h in 10% FCS/DMEM and treated for 48 h with trastuzumab, ADAM Inhibitor TMI-1 and GI254023X. Untreated cells served as negative control. Cells were harvested by trypsinization without discarding the supernatant and stained using the TACS Annexin-V-FITC Apoptosis Detection Kit (ImmunoTools, Friesoythe, Germany) according to the manufacturer’s instructions.
2.7. Statistical Analyses
2.8. List of Abbreviations
3. Results
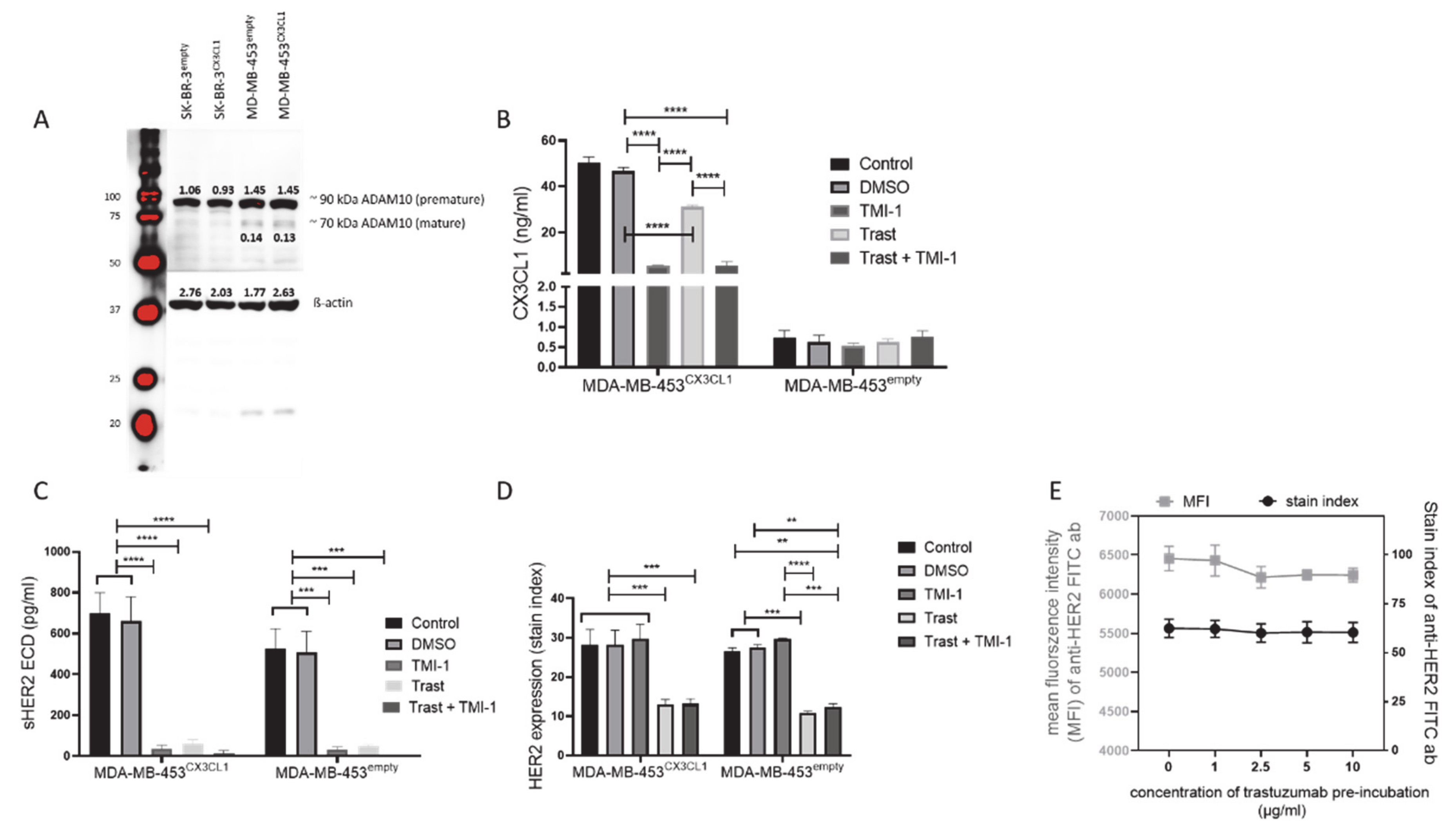
3.1. TMI-1 and GI254023X Inhibit CX3CL1 and HER2 Shedding In vitro without Affecting Tumor Cell Proliferation
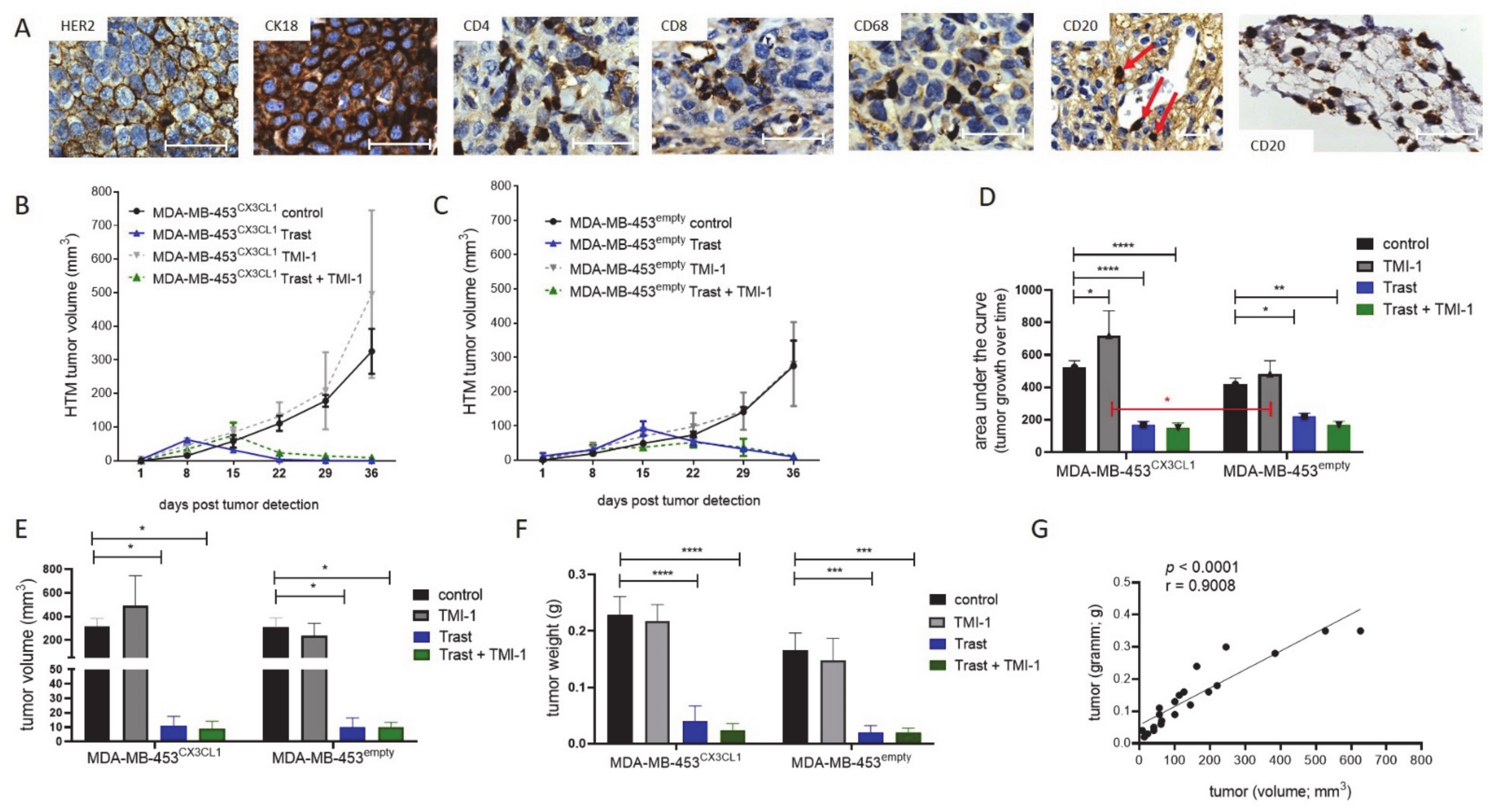
3.2. Trastuzumab Efficiently Blocked Tumor Growth with Enhanced Efficiency in CX3CL1 Overexpressing MDA-MB-453-Based HTM
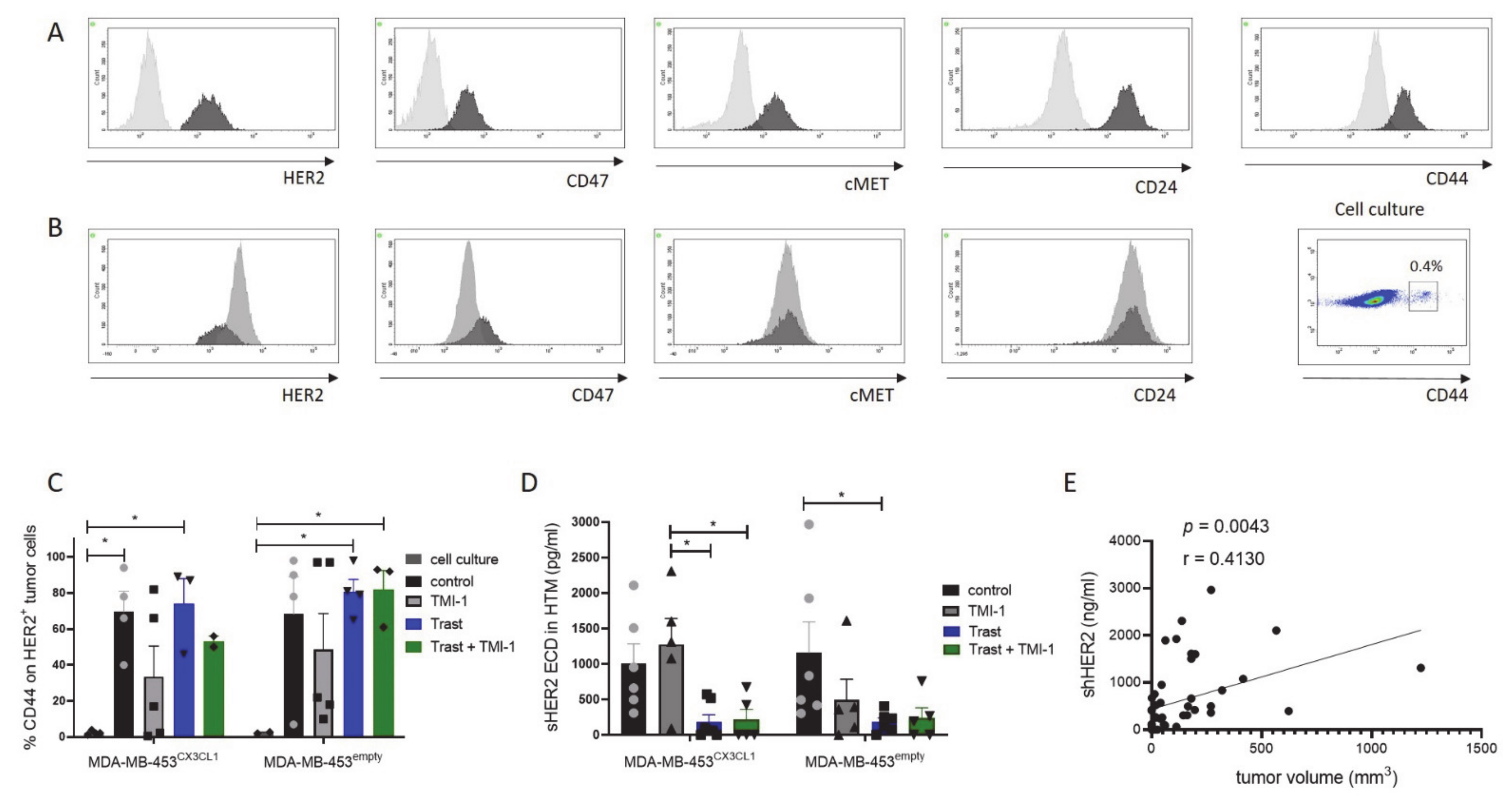
3.3. Increased CD44 Expression In vivo on MDA-MB-453CX3CL1 and MDA-MB-453empty Primary Tumor Cells
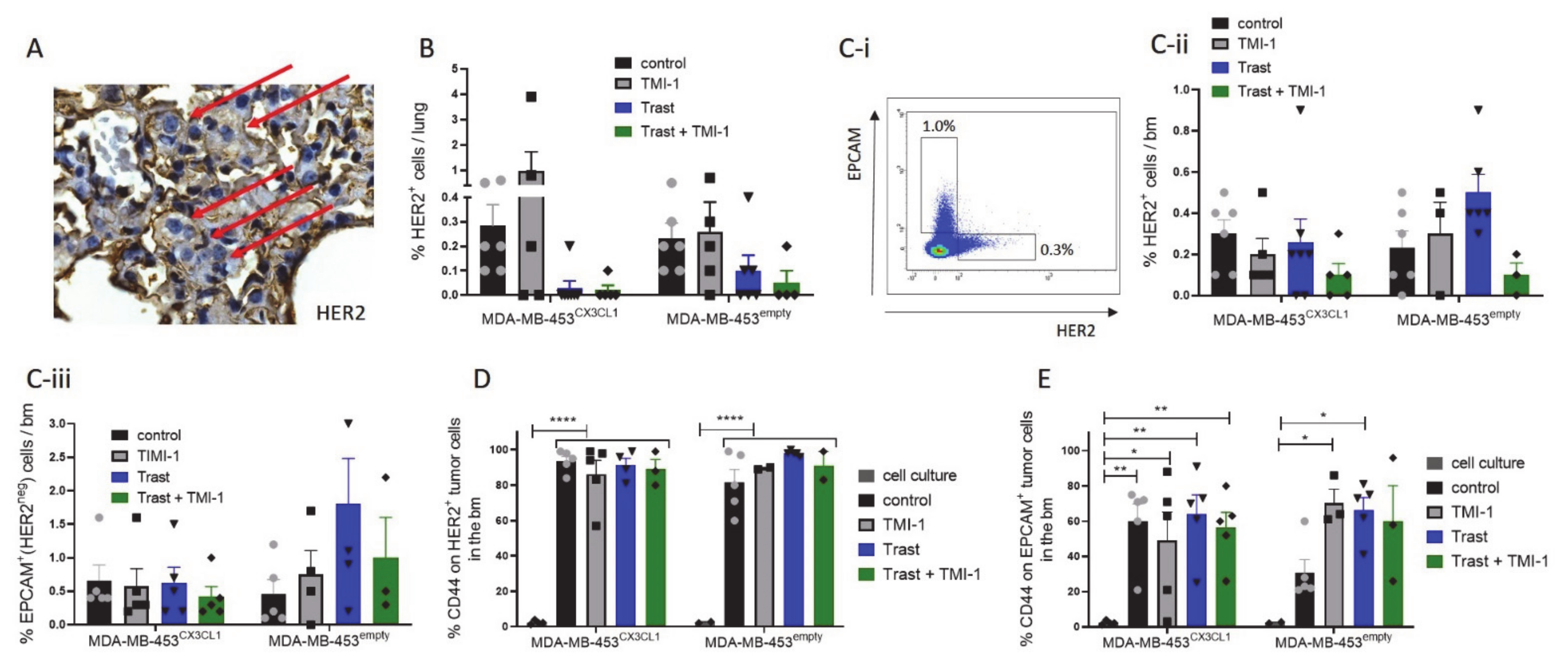
3.4. Trastuzumab Treatment Significantly Reduced Lung Metastases in MDA-MB-453CX3CL1- but Not in MDA-MB-453empty-Based HTM
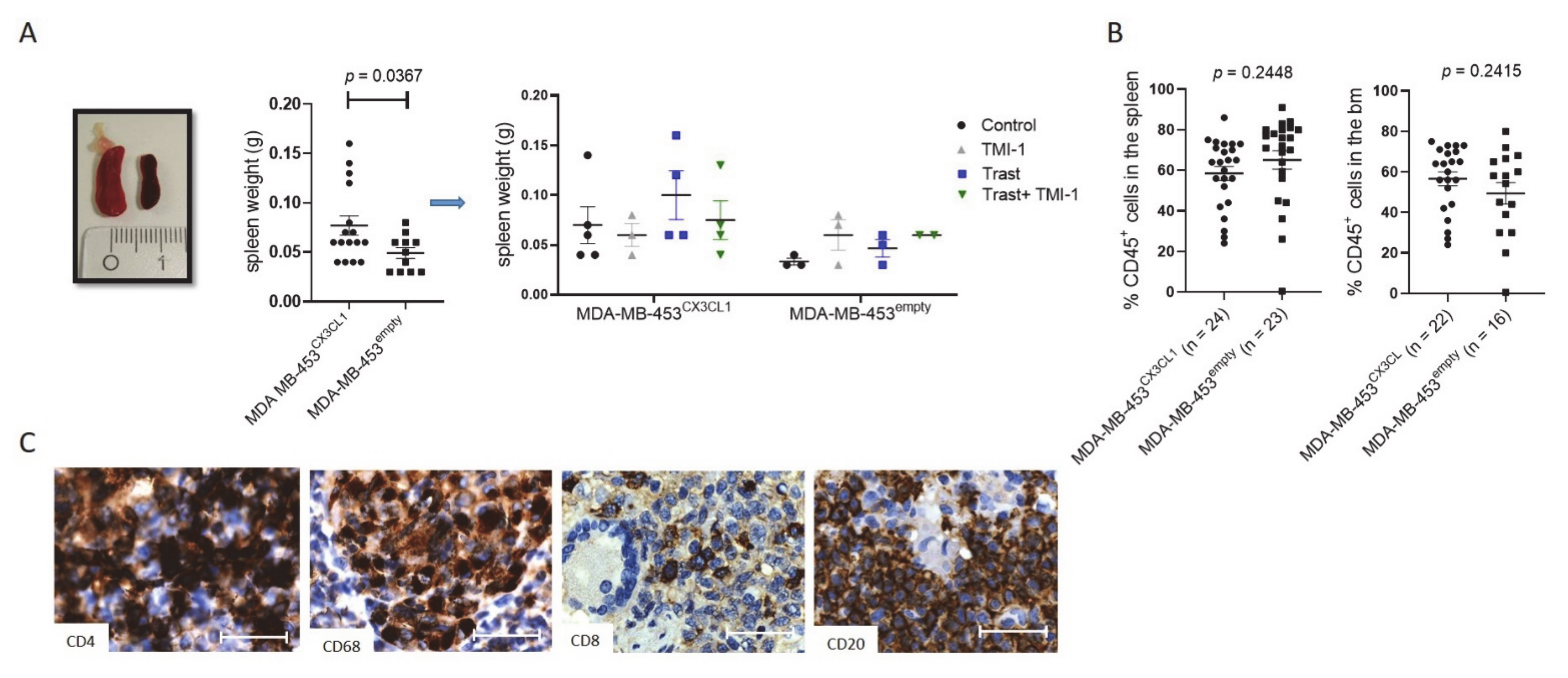
3.5. MDA-MB-453CX3CL1-Transplanted HTM Developed Enlarged Spleens but Showed Similar High Human Immune Cell Reconstitution as MDA-MB-453empty-Transplanted Mice
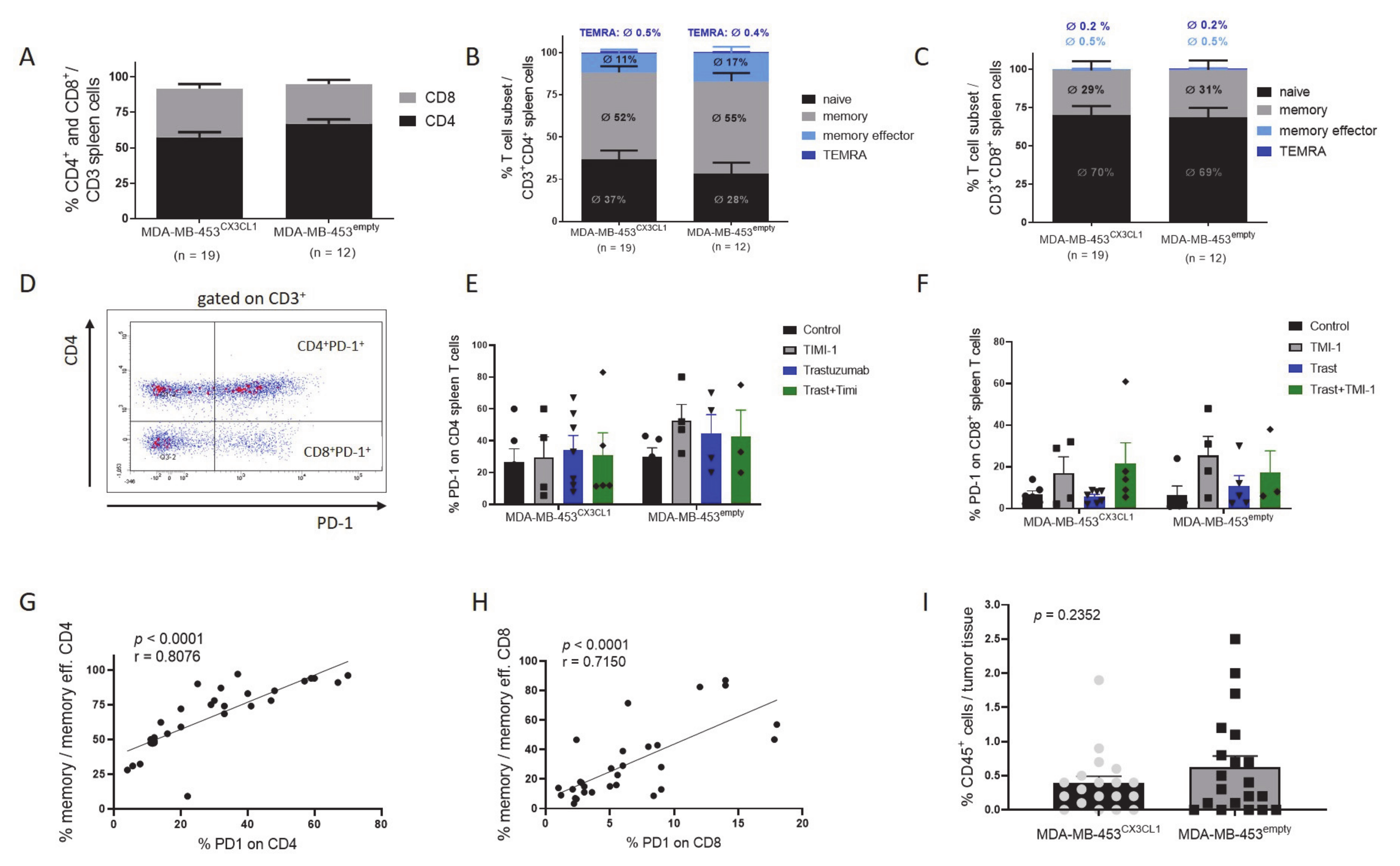
3.6. Rejection of Tumor Was Associated with Increased CD4+ T Cell Fraction and Maturation in HTM
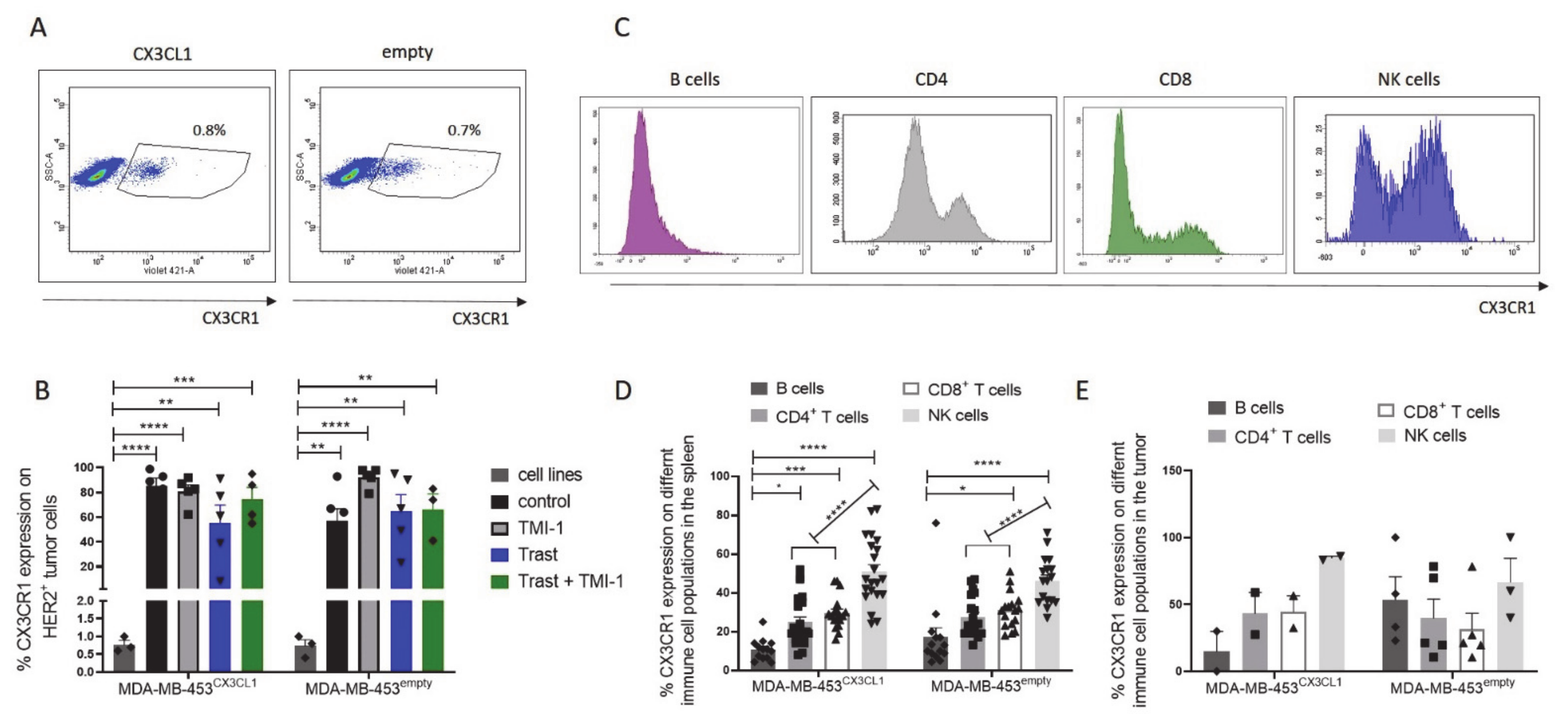
3.7. CX3CR1 Expression on Tumor Cells Was Significantly Increased In Vivo
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A. Current Updates on Trastuzumab Resistance in HER2 Overexpressing Breast Cancers. Adv. Exp. Med. Biol. 2019, 1152, 217–228. [Google Scholar] [CrossRef] [PubMed]
- Denkert, C.; Von Minckwitz, G.; Brase, J.C.; Sinn, B.V.; Gade, S.; Kronenwett, R.; Pfitzner, B.M.; Salat, C.; Loi, S.; Schmitt, W.D.; et al. Tumor-Infiltrating Lymphocytes and Response to Neoadjuvant Chemotherapy With or Without Carboplatin in Human Epidermal Growth Factor Receptor 2–Positive and Triple-Negative Primary Breast Cancers. J. Clin. Oncol. 2015, 33, 983–991. [Google Scholar] [CrossRef] [PubMed]
- Salgado, R.; Denkert, C.; Demaria, S.; Sirtaine, N.; Klauschen, F.; Pruneri, G.; Wienert, S.; van den Eynden, G.; Baehner, F.L.; Penault-Llorca, F.; et al. The evaluation of tumor-infiltrating lymphocytes (TILs) in breast cancer: Recommendations by an International TILs Working Group 2014. Ann. Oncol. 2015, 26, 259–271. [Google Scholar] [CrossRef]
- Kroemer, G.; Senovilla, L.; Galluzzi, L.; André, F.; Zitvogel, L. Natural and therapy-induced immunosurveillance in breast cancer. Nat. Med. 2015, 21, 1128–1138. [Google Scholar] [CrossRef]
- Clynes, R.A.; Towers, T.L.; Presta, L.G.; Ravetch, J.V. Inhibitory Fc receptors modulate in vivo cytoxicity against tumor targets. Nat. Med. 2000, 6, 443–446. [Google Scholar] [CrossRef]
- Barok, M.; Isola, J.; Pályi-Krekk, Z.; Nagy, P.; Juhász, I.; Vereb, G.; Kauraniemi, P.; Kapanen, A.I.; Tanner, M.; Szöllösi, J. Trastuzumab causes antibody-dependent cellular cytotoxicity–mediated growth inhibition of submacroscopic JIMT-1 breast cancer xenografts despite intrinsic drug resistance. Mol. Cancer Ther. 2007, 6, 2065–2072. [Google Scholar] [CrossRef]
- Korbecki, J.; Simińska, D.; Kojder, K.; Grochans, S.; Gutowska, I.; Chlubek, D.; Baranowska-Bosiacka, I. Fractalkine/CX3CL1 in Neoplastic Processes. Int. J. Mol. Sci. 2020, 21, 3723. [Google Scholar] [CrossRef]
- Haskell, C.A.; Cleary, M.D.; Charo, I.F. Unique role of the chemokine domain of fractalkine in cell capture. Kinetics of re-ceptor dissociation correlate with cell adhesion. J. Biol. Chem. 2000, 275, 34183–34189. [Google Scholar] [CrossRef]
- Franciszkiewicz, K.; Boissonnas, A.; Boutet, M.; Combadière, C.; Mami-Chouaib, F. Role of chemokines and chemokine receptors in shaping the effector phase of the antitumor immune response. Cancer Res. 2012, 72, 6325–6332. [Google Scholar] [CrossRef]
- Conroy, M.J.; Lysaght, J. CX3CL1 Signaling in the Tumor Microenvironment. Adv. Exp. Med. Biol. 2020, 1231, 1–12. [Google Scholar] [CrossRef]
- Wendel, M.; Galani, I.E.; Suri-Payer, E.; Cerwenka, A. Natural Killer Cell Accumulation in Tumors Is Dependent on IFN-γ and CXCR3 Ligands. Cancer Res. 2008, 68, 8437–8445. [Google Scholar] [CrossRef]
- Liang, Y.; Yi, L.; Liu, P.; Jiang, L.; Wang, H.; Hu, A.; Sun, C.; Dong, J. CX3CL1 involves in breast cancer metastasizing to the spine via the Src/FAK signaling pathway. J. Cancer 2018, 9, 3603–3612. [Google Scholar] [CrossRef]
- Jamieson-Gladney, W.L.; Zhang, Y.; Fong, A.M.; Meucci, O.; Fatatis, A. The chemokine receptor CX3CR1 is directly involved in the arrest of breast cancer cells to the skeleton. Breast Cancer Res. 2011, 13, R91. [Google Scholar] [CrossRef]
- Andre, F.; Cabioglu, N.; Assi, H.; Sabourin, J.C.; Delaloge, S.; Sahin, A.; Broglio, K.; Spano, J.P.; Combadiere, C.; Bucana, C.; et al. Expression of chemokine receptors predicts the site of metastatic relapse in patients with axillary node positive primary breast cancer. Ann. Oncol. 2006, 17, 945–951. [Google Scholar] [CrossRef]
- White, G.E.; Tan, T.C.; John, A.E.; Whatling, C.; McPheat, W.L.; Greaves, D.R. Fractalkine has anti-apoptotic and proliferative effects on human vascular smooth muscle cells via epidermal growth factor receptor signalling. Cardiovasc. Res. 2009, 85, 825–835. [Google Scholar] [CrossRef]
- Tardáguila, M.; Mira, E.; García-Cabezas, M.A.; Feijoo, A.M.; Quintela-Fandino, M.; Azcoitia, I.; Lira, S.A.; Mañes, S. CX3CL1 Promotes Breast Cancer via Transactivation of the EGF Pathway. Cancer Res. 2013, 73, 4461–4473. [Google Scholar] [CrossRef]
- Bazan, J.F.; Bacon, K.B.; Hardiman, G.; Wang, W.; Soo, K.; Rossi, D.; Greaves, D.R.; Zlotnik, A.; Schall, T.J. A new class of membrane-bound chemokine with a CX3C motif. Nature 1997, 385, 640–644. [Google Scholar] [CrossRef]
- Bronger, H.; Magdolen, V.; Goettig, P.; Dreyer, T. Proteolytic chemokine cleavage as a regulator of lymphocytic infiltration in solid tumors. Cancer Metastasis Rev. 2019, 38, 417–430. [Google Scholar] [CrossRef]
- Mullooly, M.; McGowan, P.M.; Kennedy, S.A.; Madden, S.F.; Crown, J.; O’Donovan, N.; Duffy, M.J. ADAM10: A new player in breast cancer progression? Br. J. Cancer 2015, 113, 945–951. [Google Scholar] [CrossRef]
- Hundhausen, C.; Misztela, D.; Berkhout, T.A.; Broadway, N.; Saftig, P.; Reiss, K.; Hartmann, D.; Fahrenholz, F.; Postina, R.; Matthews, V.; et al. The disintegrin-like metalloproteinase ADAM10 is involved in constitutive cleavage of CX3CL1 (fractalkine) and regulates CX3CL1-mediated cell-cell adhesion. Blood 2003, 102, 1186–1195. [Google Scholar] [CrossRef] [PubMed]
- Park, M.H.; Lee, J.S.; Yoon, J.H. High expression of CX3CL1 by tumor cells correlates with a good prognosis and increased tumor-infiltrating CD8+ T cells, natural killer cells, and dendritic cells in breast carcinoma. J. Surg. Oncol. 2012, 106, 386–392. [Google Scholar] [CrossRef] [PubMed]
- Tsang, J.Y.S.; Ni, Y.-B.; Chan, S.-K.; Shao, M.-M.; Kwok, Y.-K.; Chan, K.-W.; Tan, P.H.; Tse, G.M. CX3CL1 expression is associated with poor outcome in breast cancer patients. Breast Cancer Res. Treat. 2013, 140, 495–504. [Google Scholar] [CrossRef] [PubMed]
- Wege, A.K.; Ernst, W.; Eckl, J.; Frankenberger, B.; Vollmann-Zwerenz, A.; Männel, D.N.; Ortmann, O.; Kroemer, A.; Brockhoff, G. Humanized tumor mice-A new model to study and manipulate the immune response in advanced cancer therapy. Int. J. Cancer 2011, 129, 2194–2206. [Google Scholar] [CrossRef]
- Wege, A.K.; Schmidt, M.; Ueberham, E.; Ponnath, M.; Ortmann, O.; Brockhoff, G.; Lehmann, J. Co-transplantation of human hematopoietic stem cells and human breast cancer cells in NSG mice: A novel approach to generate tumor cell specific human antibodies. mAbs 2014, 6, 968–977. [Google Scholar] [CrossRef]
- Wege, A.K.; Weber, F.; Kroemer, A.; Ortmann, O.; Nimmerjahn, F.; Brockhoff, G. IL-15 enhances the anti-tumor activity of trastuzumab against breast cancer cells but causes fatal side effects in humanized tumor mice (HTM). Oncotarget 2016, 8, 2731–2744. [Google Scholar] [CrossRef]
- Rom-Jurek, E.-M.; Kirchhammer, N.; Ugocsai, P.; Ortmann, O.; Wege, A.K.; Brockhoff, G. Regulation of Programmed Death Ligand 1 (PD-L1) Expression in Breast Cancer Cell Lines In Vitro and in Immunodeficient and Humanized Tumor Mice. Int. J. Mol. Sci. 2018, 19, 563. [Google Scholar] [CrossRef]
- Dreyer, T.F.; Kuhn, S.; Stange, C.; Heithorst, N.; Schilling, D.; Jelsma, J.; Sievert, W.; Seitz, S.; Stangl, S.R.; Hapfelmeier, A.; et al. The chemokine CX3CL1 improves trastuzumab efficacy in HER2-low-expressing cancer in vitro and in vivo. Cancer Immunol. Res. 2021. [Google Scholar] [CrossRef]
- Magnifico, A.; Albano, L.; Campaner, S.; Campiglio, M.; Pilotti, S.; Ménard, S.; Tagliabue, E. Protein Kinase Cα Determines HER2 Fate in Breast Carcinoma Cells with HER2 Protein Overexpression without Gene Amplification. Cancer Res. 2007, 67, 5308–5317. [Google Scholar] [CrossRef]
- Yan, X.; Lin, J.; Talabattula, V.A.N.; Mußmann, C.; Yang, F.; Wree, A.; Rolfs, A.; Luo, J. ADAM10 Negatively Regulates Neuronal Differentiation during Spinal Cord Development. PLoS ONE 2014, 9, e84617. [Google Scholar] [CrossRef]
- Wege, A.K.; Kirchhammer, N.; Kazandjian, L.V.; Prassl, S.; Brandt, M.; Piendl, G.; Ortmann, O.; Fischer, S.; Brockhoff, G. A novel rabbit derived anti-HER2 antibody with pronounced therapeutic effectiveness on HER2-positive breast cancer cells in vitro and in humanized tumor mice (HTM). J. Transl. Med. 2020, 18, 1–16. [Google Scholar] [CrossRef]
- Ohta, M.; Tanaka, F.; Yamaguchi, H.; Sadanaga, N.; Inoue, H.; Mori, M. The high expression of Fractalkine results in a better prognosis for colorectal cancer patients. Int. J. Oncol. 2005, 26, 41–47. [Google Scholar] [CrossRef]
- Hyakudomi, M.; Matsubara, T.; Hyakudomi, R.; Yamamoto, T.; Kinugasa, S.; Yamanoi, A.; Maruyama, R.; Tanaka, T. Increased Expression of Fractalkine is Correlated with a Better Prognosis and an Increased Number of Both CD8+ T Cells and Natural Killer Cells in Gastric Adenocarcinoma. Ann. Surg. Oncol. 2008, 15, 1775–1782. [Google Scholar] [CrossRef]
- Zhang, X.; Wei, H.; Chen, Q.; Tian, Z. Activation of human natural killer cells by recombinant membrane-expressed frac-talkine on the surface of tumor cells. Oncol. Rep. 2007, 17, 1371–1375. [Google Scholar]
- Zhang, J.; Yang, W.; Luo, B.; Hu, B.; Maheshwari, A.; Fallon, M.B. The role of CX3CL1/CX3CR1 in pulmonary angiogenesis and intravascular monocyte accumulation in rat experimental hepatopulmonary syndrome. J. Hepatol. 2012, 57, 752–758. [Google Scholar] [CrossRef]
- Ryu, J.; Lee, C.-W.; Hong, K.-H.; Shin, J.-A.; Lim, S.-H.; Park, C.-S.; Shim, J.; Nam, K.B.; Choi, K.-J.; Kim, Y.-H.; et al. Activation of fractalkine/CX3CR1 by vascular endothelial cells induces angiogenesis through VEGF-A/KDR and reverses hindlimb ischaemia. Cardiovasc. Res. 2007, 78, 333–340. [Google Scholar] [CrossRef]
- Marchica, V.; Toscani, D.; Corcione, A.; Bolzoni, M.; Storti, P.; Vescovini, R.; Ferretti, E.; Accardalla Palmai, B.D.; Vicario, E.; Accardi, F.; et al. Bone Marrow CX3CL1/Fractalkine is a New Player of the Pro-Angiogenic Microenvironment in Multiple Myeloma Patients. Cancers 2019, 11, 321. [Google Scholar] [CrossRef]
- Park, Y.; Lee, J.; Kwak, J.-Y.; Noh, K.; Yim, E.; Kim, H.-K.; Kim, Y.J.; Broxmeyer, H.E.; Kim, J.-A. Fractalkine induces angiogenic potential in CX3CR1-expressing monocytes. J. Leukoc. Biol. 2017, 103, 53–66. [Google Scholar] [CrossRef]
- Van Der Leun, A.M.; Thommen, D.S.; Schumacher, T.N. CD8+ T cell states in human cancer: Insights from single-cell analysis. Nat. Rev. Cancer 2020, 20, 218–232. [Google Scholar] [CrossRef]
- Gros, A.; Robbins, P.F.; Yao, X.; Li, Y.F.; Turcotte, S.; Tran, E.; Wunderlich, J.R.; Mixon, A.; Farid, S.; Dudley, M.E.; et al. PD-1 identifies the patient-specific CD8+ tumor-reactive repertoire infiltrating human tumors. J. Clin. Investig. 2014, 124, 2246–2259. [Google Scholar] [CrossRef]
- Saftig, P.; Reiss, K. The “A Disintegrin And Metalloproteases” ADAM10 and ADAM17: Novel drug targets with therapeutic potential? Eur. J. Cell Biol. 2011, 90, 527–535. [Google Scholar] [CrossRef]
- Moss, M.L.; Minond, D. Recent Advances in ADAM17 Research: A Promising Target for Cancer and Inflammation. Mediat. Inflamm. 2017, 2017, 1–21. [Google Scholar] [CrossRef]
- Smith, T.M.J.; Tharakan, A.; Martin, R.K. Targeting ADAM10 in Cancer and Autoimmunity. Front. Immunol. 2020, 11, 499. [Google Scholar] [CrossRef]
- McGowan, P.M.; Ryan, B.M.; Hill, A.D.; McDermott, E.; O’Higgins, N.; Duffy, M.J. ADAM-17 Expression in Breast Cancer Correlates with Variables of Tumor Progression. Clin. Cancer Res. 2007, 13, 2335–2343. [Google Scholar] [CrossRef]
- Duffy, M.J.; McKiernan, E.; O’Donovan, N.; McGowan, P.M. Role of ADAMs in Cancer Formation and Progression. Clin. Cancer Res. 2009, 15, 1140–1144. [Google Scholar] [CrossRef]
- McGowan, P.M.; McKiernan, E.; Bolster, F.; Ryan, B.M.; Hill, A.D.K.; McDermott, E.W.; Evoy, D.; O’Higgins, N.; Crown, J.; Duffy, M.J. ADAM-17 predicts adverse outcome in patients with breast cancer. Ann. Oncol. 2008, 19, 1075–1081. [Google Scholar] [CrossRef]
- Mezil, L.; Berruyer-Pouyet, C.; Cabaud, O.; Josselin, E.; Combes, S.; Brunel, J.-M.; Viens, P.; Collette, Y.; Birnbaum, D.; Lopez, M. Tumor Selective Cytotoxic Action of a Thiomorpholin Hydroxamate Inhibitor (TMI-1) in Breast Cancer. PLoS ONE 2012, 7, e43409. [Google Scholar] [CrossRef]
- Venkatesh, H.S.; Tam, L.T.; Woo, P.J.; Lennon, J.; Nagaraja, S.; Gillespie, S.M.; Ni, J.; Duveau, D.Y.; Morris, P.J.; Zhao, J.J.; et al. Targeting neuronal activity-regulated neuroligin-3 dependency in high-grade glioma. Nat. Cell Biol. 2017, 549, 533–537. [Google Scholar] [CrossRef]
- Perrier, A.; Gligorov, J.; Lefevre, G.; Boissan, M. The extracellular domain of Her2 in serum as a biomarker of breast cancer. Lab. Investig. 2018, 98, 696–707. [Google Scholar] [CrossRef]
- Friedman, S.; Levy, R.; Garrett, W.; Doval, D.; Bondarde, S.; Sahoo, T.; Lokanatha, D.; Julka, P.; Shenoy, K.; Nagarkar, R.; et al. Clinical Benefit of INCB7839, a Potent and Selective Inhibitor of ADAM10 and ADAM17, in Combination with Trastuzumab in Metastatic HER2 Positive Breast Cancer Patients. In Poster Sessio Abstracts, Proceedings of the Abstracts: Thirty-Second Annual CTRC-AACR San Antonio Breast Cancer Symposium San Antonio, TX, USA, 10–13 December 2009; American Association for Cancer Research: Philadelphia, PA, USA, 2009; Volume 69, p. 5056. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HER2+ Lung Metastases | MDA-MB-453CX3CL1 (n/n) | MDA-MB-453empty (n/n) |
|---|---|---|
| Control | 6/6 | 6/6 |
| TMI-1 | 3/5 | 4/5 |
| Trast | 1/7 *A | 3/6 |
| Trast and TMI-1 | 1/5 *B | 1/4 *C |
| HER2+ bm Metastases | MDA-MB-453CX3CL1 (n/n) | MDA-MB-453empty (n/n) |
|---|---|---|
| Control | 6/6 | 5/6 |
| TMI-1 | 5/5 | 2/3 |
| Trast | 5/7 | 6/6 |
| Trast and TMI-1 | 3/5 | 2/3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wege, A.K.; Dreyer, T.F.; Teoman, A.; Ortmann, O.; Brockhoff, G.; Bronger, H. CX3CL1 Overexpression Prevents the Formation of Lung Metastases in Trastuzumab-Treated MDA-MB-453-Based Humanized Tumor Mice (HTM). Cancers 2021, 13, 2459. https://doi.org/10.3390/cancers13102459
Wege AK, Dreyer TF, Teoman A, Ortmann O, Brockhoff G, Bronger H. CX3CL1 Overexpression Prevents the Formation of Lung Metastases in Trastuzumab-Treated MDA-MB-453-Based Humanized Tumor Mice (HTM). Cancers. 2021; 13(10):2459. https://doi.org/10.3390/cancers13102459
Chicago/Turabian StyleWege, Anja Kathrin, Tobias F. Dreyer, Attila Teoman, Olaf Ortmann, Gero Brockhoff, and Holger Bronger. 2021. "CX3CL1 Overexpression Prevents the Formation of Lung Metastases in Trastuzumab-Treated MDA-MB-453-Based Humanized Tumor Mice (HTM)" Cancers 13, no. 10: 2459. https://doi.org/10.3390/cancers13102459
APA StyleWege, A. K., Dreyer, T. F., Teoman, A., Ortmann, O., Brockhoff, G., & Bronger, H. (2021). CX3CL1 Overexpression Prevents the Formation of Lung Metastases in Trastuzumab-Treated MDA-MB-453-Based Humanized Tumor Mice (HTM). Cancers, 13(10), 2459. https://doi.org/10.3390/cancers13102459