Human FBXL8 Is a Novel E3 Ligase Which Promotes BRCA Metastasis by Stimulating Pro-Tumorigenic Cytokines and Inhibiting Tumor Suppressors



 , and
, and 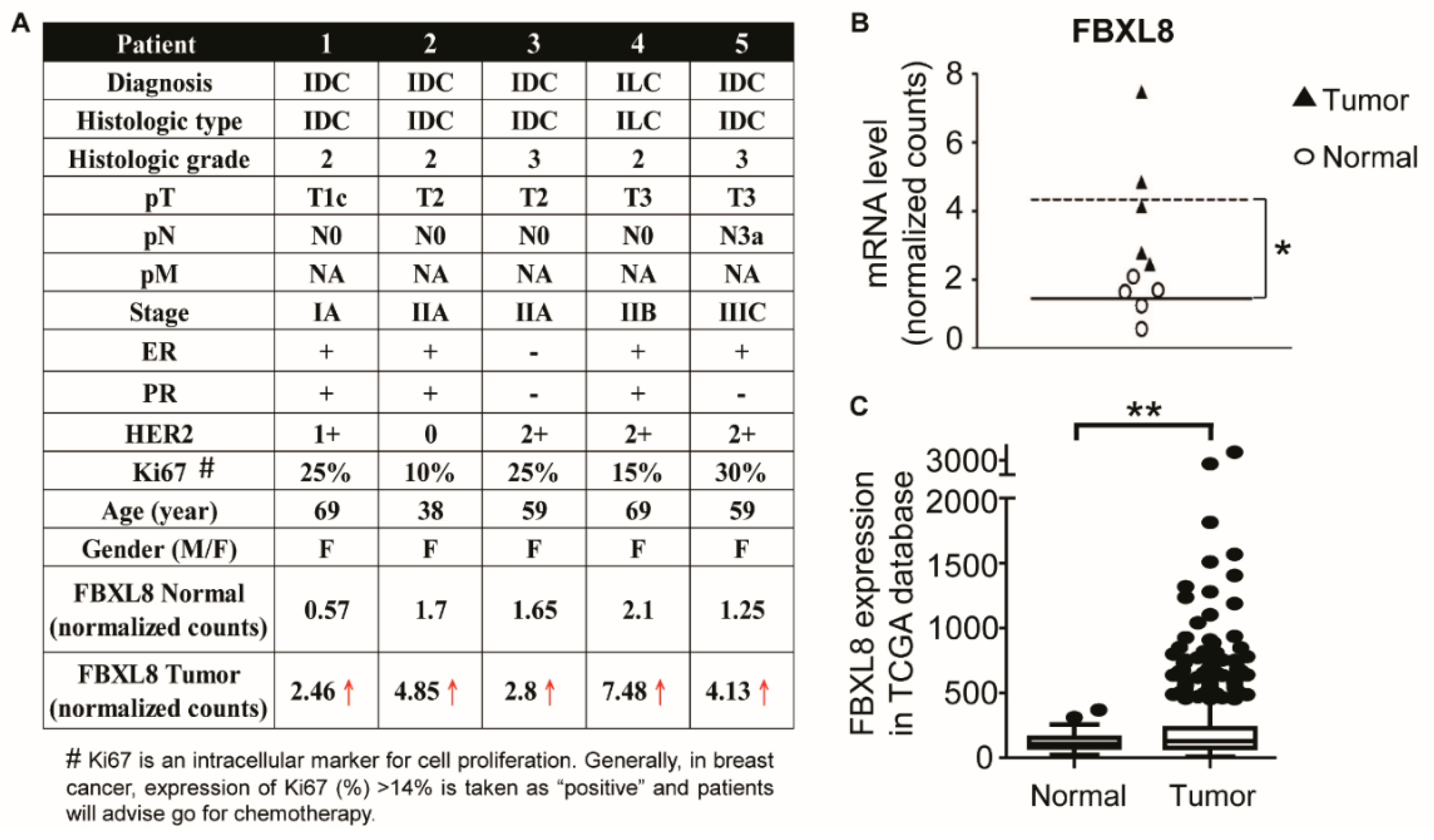{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Clinicopathological Parameters of BRCA Patient Tissues Used For the Global RNA Sequencing
2.2. FBXL8 Is Significantly Upregulated in Human Primary BRCA Tissues
2.3. FBXL8 Is Upregulated with the Advancement of BRCA Stages
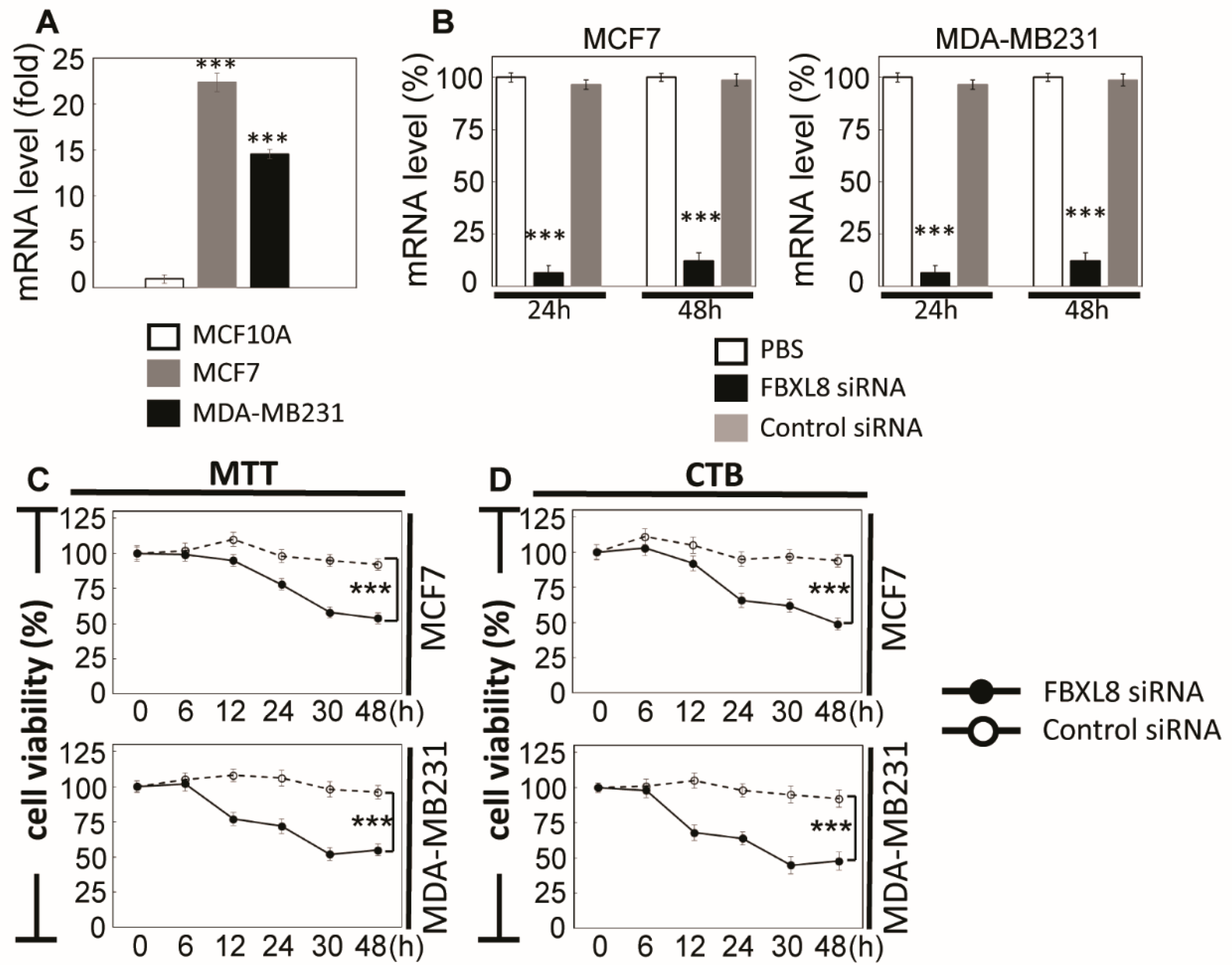
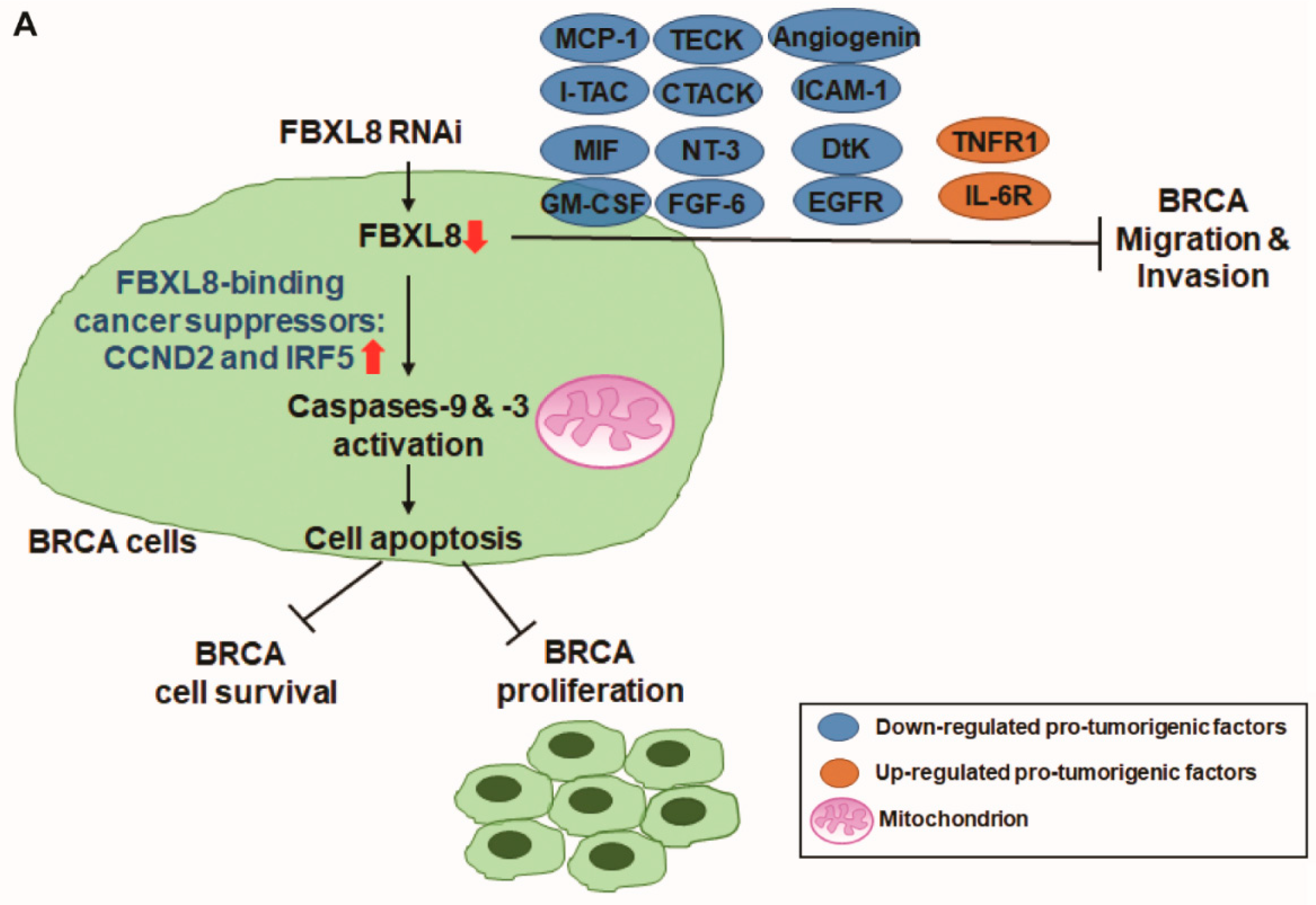
2.4. FBXL8 is a Novel Anti-Apoptosis Factor in BRCA
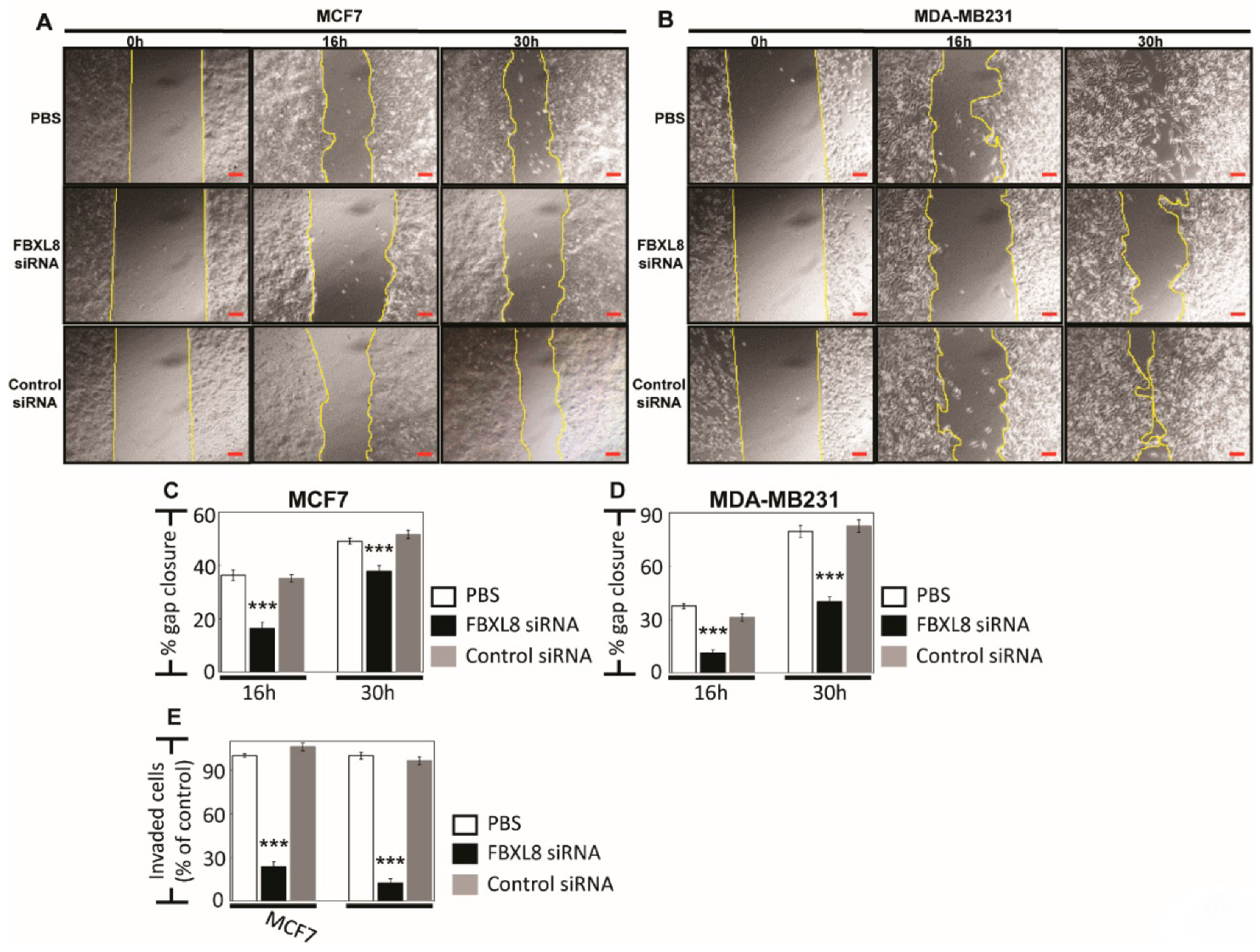
2.5. Knockdown of FBXL8 Inhibited BRCA Cell Migration and Invasion
2.6. Knockdown of FBXL8 Suppressed Pro-Inflammatory Cytokines/Chemokines in BRCA
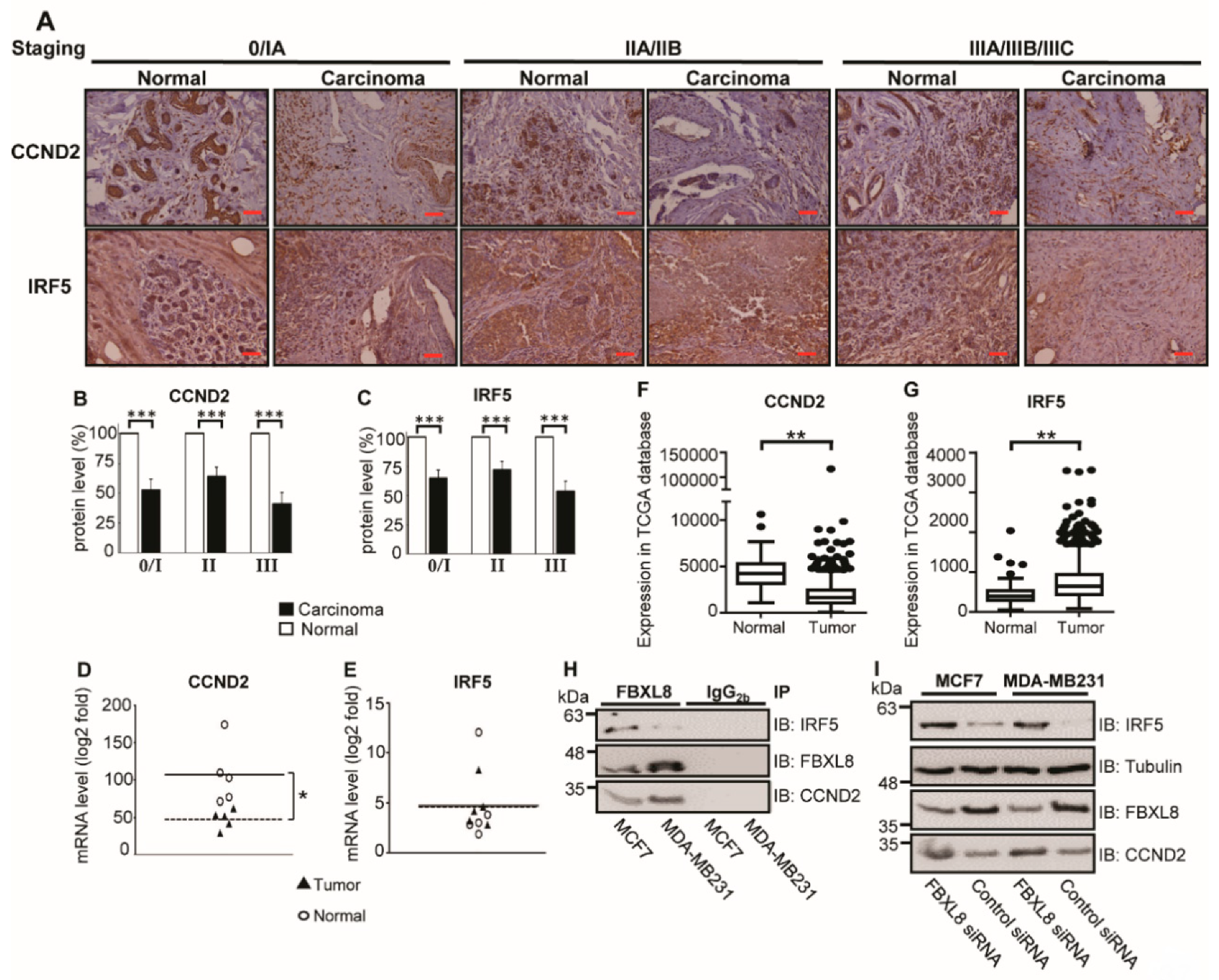
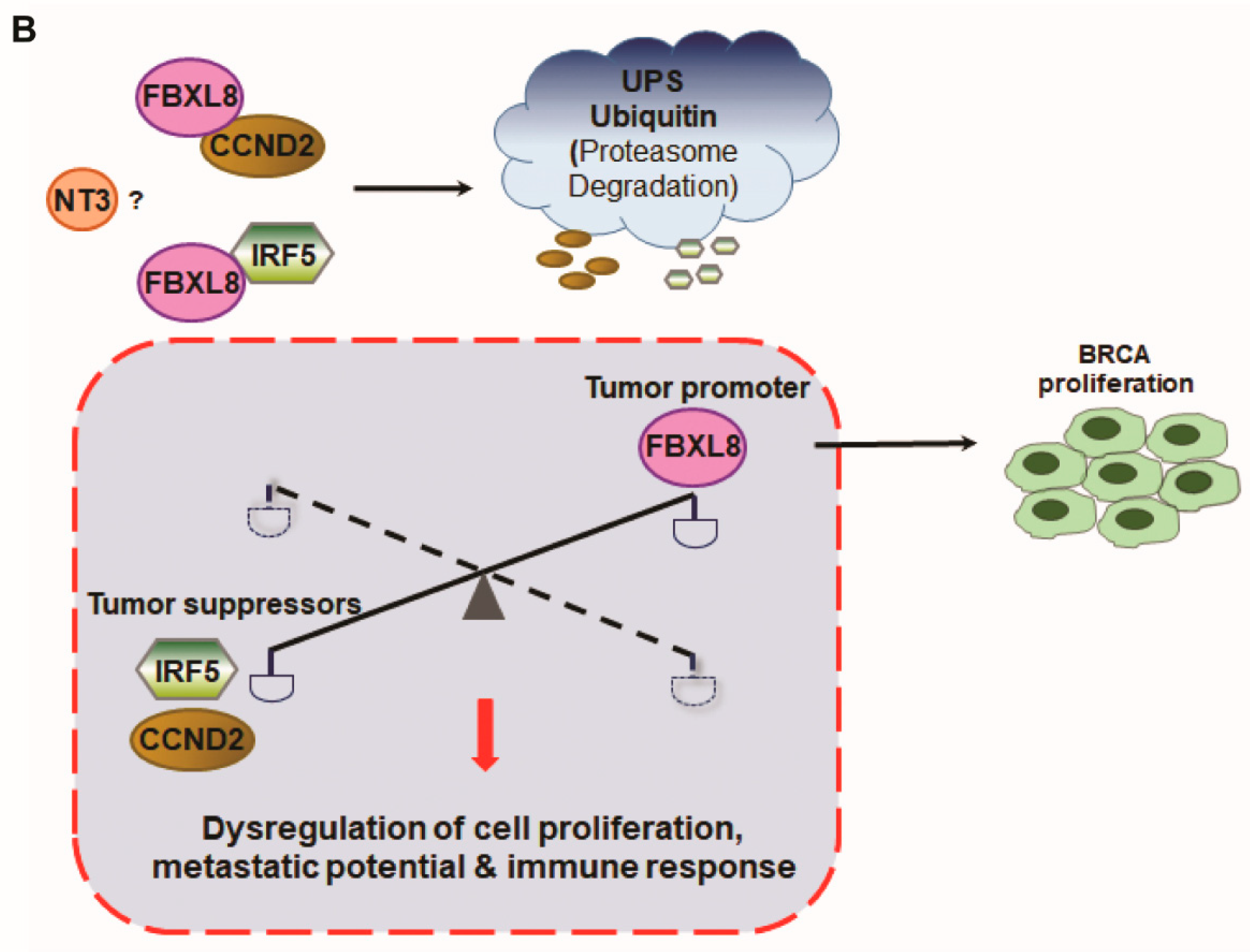
2.7. Cancer-Suppressors, CCND2 and IRF5, Are FBXL8-Binding Partners in BRCA
3. Discussion
4. Materials and Methods
4.1. Tissue Samples
4.2. Cell Lines and Reagents
4.3. Next-Generation Sequencing (NGS)-Based RNA-Seq Analysis
4.3.1. RNA Isolation for NGS-Based RNA-Seq
4.3.2. Library Preparation & Sequencing for NGS-Based RNA-Seq Application
4.3.3. RNA Sequence Analysis
4.4. TCGA Database Analysis
4.5. Quantitative Real-Time PCR
4.6. FBXL8 Knockdown in BRCA Cells
4.7. Cell Viability Assay
4.8. Cell Proliferation Assay
4.9. Cellular Apoptosis Assay
4.10. Caspases-9 and -3 Assays
4.11. Cell Migration and Invasion Assays
4.12. Cytokine Arrays
4.13. ELISA Quantification of Cytokines
4.14. Co-IP and Immunodetection
4.15. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Coughlin, S.S.; Ekwueme, N.U. Breast cancer as a global health concern. Cancer Epidemiol. 2009, 33, 315–318. [Google Scholar] [CrossRef] [PubMed]
- Jemal, A.; Siegel, R.; Xu, J.; Ward, E. Cancer statistics, 2010. CA Cancer J. Clin. 2010, 60, 277–300. [Google Scholar] [CrossRef] [PubMed]
- Orlowski, R.Z.; Dees, E.C. The role of the ubiquitination-proteasome pathway in breast cancer: Applying drugs that affect the ubiquitin-proteasome pathway to the therapy of breast cancer. Breast Cancer Res. 2002, 5, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Randle, S.J.; Laman, H. F-box protein interactions with the hallmark pathways in cancer. Semin. Cancer Biol. 2016, 36, 3–17. [Google Scholar] [CrossRef]
- Soucy, T.A.; Smith, P.G.; Rolfe, M. Targeting NEDD8-activated Cullin-RING ligases for the treatment of cancer. Clin. Cancer Res. 2009, 15, 3912–3916. [Google Scholar] [CrossRef]
- Soucy, T.A.; Smith, P.G.; Milhollen, M.A.; Berger, A.J.; Gavin, J.M.; Adhikari, S.; Brownell, J.E.; Burke, K.E.; Cardin, D.P.; Critchley, S.; et al. An inhibitor of NEDD8-activating enzyme as a new approach to treat cancer. Nature 2009, 458, 732–736. [Google Scholar] [CrossRef]
- Chen, X.; Cui, D.; Bi, Y.; Shu, J.; Xiong, X.; Zhao, Y.; Shu, J. AKT inhibitor MK-2206 sensitizes breast cancer cells to MLN4924, a first-in-class NEDD8-activating enzyme (NAE) inhibitor. Cell Cycle 2018, 17, 2069–2079. [Google Scholar] [CrossRef]
- Chang, S.C.; Ding, J.L. Ubiquitination and SUMOylation in the chronic inflammatory tumor microenvironment. Biochim. Biophys. Acta 2018, 1870, 165–175. [Google Scholar] [CrossRef]
- Mathewson, N.; Toubai, T.; Kapeles, S.; Sun, Y.; Oravecz-Wilson, K.; Tamaki, H.; Wang, Y.; Hou, G.; Sun, Y.; Reddy, P. Neddylation plays an important role in the regulation of murine and human dendritic cell function. Blood 2013, 122, 2062–2073. [Google Scholar] [CrossRef]
- Nakayama, K.I.; Nakayama, K. Ubiquitin ligases: Cell-cycle control and cancer. Nat. Rev. Cancer 2006, 6, 369–381. [Google Scholar] [CrossRef]
- Chang, S.C.; Ding, J.L. Ubiquitination by SAG regulates macrophage survival/death and immune response during infection. Cell Death Differ. 2014, 21, 1388–1398. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.C.; Choo, W.Q.W.; Toh, H.C.; Ding, J.L. SAG-UPS attenuates proapoptotic SARM and Noxa to confer survival advantage to early hepatocellular carcinoma. Cell Death Discov. 2015, 1, 15032. [Google Scholar] [CrossRef] [PubMed]
- Pierce, B.L.; Ballard-Barbash, R.; Bernstein, L.; Baumgartner, R.N.; Neuhouser, M.L.; Wener, M.H.; Baumgartner, K.B.; Gilliland, F.D.; Sorensen, B.E.; McTiernan, A.; et al. Elevated biomarkers of inflammation are associated with reduced survival among breast cancer patients. J. Clin. Oncol. 2009, 27, 3437–3444. [Google Scholar] [CrossRef]
- Song, Y.; Lin, M.; Liu, Y.; Wang, Z.; Zhu, X. Emerging role of F-box proteins in the regulation of epithelial-mesenchymal transition and stem cells in human cancers. Stem Cell Res. Ther. 2019, 10, 124. [Google Scholar] [CrossRef]
- Wang, Z.; Liu, P.; Inuzuka, H.; Wei, W. Roles of F-box proteins in cancer. Nat. Rev. Cancer 2014, 14, 233–247. [Google Scholar] [CrossRef]
- Kipreos, E.T.; Pagano, M. The F-box protein family. Genome Biol. 2000, 1, 3001–3002. [Google Scholar] [CrossRef]
- Skaar, J.R.; Pagan, J.; Pagano, M. Mechanisms and function of substrate recruitment by F-box proteins. Nat. Rev. Mol. Cell Biol. 2013, 14, 369–381. [Google Scholar] [CrossRef]
- Sakuma, M.; Akahira, J.-I.; Ito, K.; Niikura, H.; Moriya, T.; Okamura, K.; Sasano, H.; Yaegashi, N. Promoter methylation status of the Cyclin D2 gene is associated with poor prognosis in human epithelial ovarian cancer. Cancer Sci. 2007, 98, 380–386. [Google Scholar] [CrossRef]
- Hu, G.; Mancl, M.E.; Barnes, B.J. Signaling through IFN regulatory factor-5 Sensitizes p53-deficient tumors to DNA damage–Induced apoptosis and cell death. Cancer Res. 2005, 65, 7403–7412. [Google Scholar] [CrossRef]
- Bi, X.; Hameed, M.; Mirani, N.; Pimenta, E.M.; Anari, J.; Barnes, B.J. Loss of interferon regulatory factor 5 (IRF5) expression in human ductal carcinoma correlates with disease stage and contributes to metastasis. Breast Cancer Res. 2011, 13, R111. [Google Scholar] [CrossRef]
- Feng, C.; Yang, F.; Wang, J. FBXO4 inhibits lung cancer cell survival by targeting Mcl-1 for degradation. Cancer Gene Ther. 2017, 24, 342–347. [Google Scholar] [CrossRef] [PubMed]
- Yeh, C.H.; Bellon, M.; Nicot, C. FBXW7: A critical tumor suppressor of human cancers. Mol. Cancer 2018, 17, 115. [Google Scholar] [CrossRef] [PubMed]
- Duan, H.; Wang, Y.; Aviram, M.; Swaroop, M.; Loo, J.A.; Bian, J.; Tian, Y.; Mueller, T.; Bisgaier, C.L.; Sun, Y. SAG, a novel zinc RING finger protein that protects cells from apoptosis induced by redox agents. Mol. Cell. Biol. 1999, 19, 3145–3155. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.; Yang, J.; Hao, X.; Zheng, M.; He, H.; Xiong, X.; Xu, L.; Sun, Y. Validation of sag/rbx2/roc2 e3 ubiquitin ligase as an anticancer and radiosensitizing target. Clin. Cancer Res. 2010, 16, 814–824. [Google Scholar] [CrossRef]
- Chang, S.C.; Ding, J.L. Sag-ups regulates malignant transformation—From chronic inflammation to pro-tumorigenesis to liver cancer. Cell Death Dis. 2015, 6, e1941. [Google Scholar] [CrossRef][Green Version]
- Dutta, S.; Bandyopadhyay, C.; Bottero, V.; Veettil, M.V.; Wilson, L.; Pins, M.R.; Johnson, K.E.; Warshall, C.; Chandran, B. Angiogenin interacts with the plasminogen activation system at the cell surface of breast cancer cells to regulate plasmin formation and cell migration. Mol. Oncol. 2014, 8, 483–507. [Google Scholar] [CrossRef]
- Reggiani, F.; Bertolini, F. GM-CSF promotes a supportive adipose and lung microenvironment in metastatic breast cancer. Oncoscience 2017, 4, 126–127. [Google Scholar] [CrossRef]
- Köstler, W.J.; Tomek, S.; Brodowicz, T.; Budinsky, A.C.; Flamm, M.; Hejna, M.; Krainer, M.; Wiltschke, C.; Zielinski, C.C. Soluble ICAM-1 in breast cancer: Clinical significance and biological implications. Cancer Immunol. Immunother. 2001, 50, 483–490. [Google Scholar] [CrossRef]
- Dickler, M.N.; Cobleigh, M.A.; Miller, K.D.; Klein, P.M.; Winer, E.P. Efficacy and safety of erlotinib in patients with locally advanced or metastatic breast cancer. Breast Cancer Res. Treat. 2008, 115, 115–121. [Google Scholar] [CrossRef]
- Louie, E.; Chen, X.F.; Coomes, A.; Ji, K.; Tsirka, S.; Chen, E.I. Neurotrophin-3 modulates breast cancer cells and the microenvironment to promote the growth of breast cancer brain metastasis. Oncogene 2012, 32, 4064–4077. [Google Scholar] [CrossRef]
- Dutta, P.; Sarkissyan, M.; Paico, K.; Wu, Y.; Vadgama, J.V. MCP-1 is overexpressed in triple-negative breast cancers and drives cancer invasiveness and metastasis. Breast Cancer Res. Treat. 2018, 170, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Balogh, K.N.; Templeton, D.J.; Cross, J.V. Macrophage migration inhibitory factor protects cancer cells from immunogenic cell death and impairs anti-tumor immune responses. PLoS ONE 2018, 13, e0197702. [Google Scholar] [CrossRef] [PubMed]
- Conroy, H.; Mawhinney, L.; Donnelly, S.C. Inflammation and cancer: Macrophage migration inhibitory factor (MIF)--the potential missing link. QJM Int. J. Med. 2010, 103, 831–836. [Google Scholar] [CrossRef] [PubMed]
- Rupertus, K.; Sinistra, J.; Scheuer, C.; Nickels, R.M.; Schilling, M.K.; Menger, M.D.; Kollmar, O. Interaction of the chemokines I-TAC (CXCL11) and SDF-1 (CXCL12) in the regulation of tumor angiogenesis of colorectal cancer. Clin. Exp. Metastasis 2014, 31, 447–459. [Google Scholar] [CrossRef] [PubMed]
- Ejaeidi, A.A.; Craft, B.S.; Puneky, L.V.; Lewis, R.E.; Cruse, J.M. Hormone receptor-independent CXCL10 production is associated with the regulation of cellular factors linked to breast cancer progression and metastasis. Exp. Mol. Pathol. 2015, 99, 163–172. [Google Scholar] [CrossRef]
- Eubank, T.D.; Roberts, R.D.; Khan, M.; Curry, J.M.; Nuovo, G.J.; Kuppusamy, P.; Marsh, C.B. Granulocyte macrophage colony-stimulating factor inhibits breast cancer growth and metastasis by invoking an anti-angiogenic program in tumor-educated macrophages. Cancer Res. 2009, 69, 2133–2140. [Google Scholar] [CrossRef]
- Lillard, J.W.; Johnson-Holiday, C.; Singh, R.; Johnson, E.; Singh, S.; Stockard, C.R.; Grizzle, W.E. CCL25 mediates migration, invasion and matrix metalloproteinase expression by breast cancer cells in a CCR9-dependent fashion. Int. J. Oncol. 2011, 38, 1279–1285. [Google Scholar] [CrossRef][Green Version]
- Sarvaiya, P.J.; Guo, D.; Ulasov, I.V.; Gabikian, P.; Lesniak, M.S. Chemokines in tumor progression and metastasis. Oncotarget 2013, 4, 2171–2185. [Google Scholar] [CrossRef]
- Fortner, R.T.; Sarink, D.; Schock, H.; Johnson, T.; Tjonneland, A.; Olsen, A.; Overvad, K.; Affret, A.; His, M.; Boutron-Ruault, M.; et al. Osteoprotegerin and breast cancer risk by hormone receptor subtype: A nested case-control study in the EPIC cohort. BMC Med. 2017, 15, 26. [Google Scholar] [CrossRef]
- Martínez-Reza, I.; Díaz, L.; García-Becerra, R. Preclinical and clinical aspects of TNF-α and its receptors TNFR1 and TNFR2 in breast cancer. J. Biomed. Sci. 2017, 24, 90. [Google Scholar] [CrossRef]
- Hung, C.S.; Wang, S.C.; Yen, Y.T.; Lee, T.H.; Wen, W.C.; Lin, R.K. Hypermethylation of CCND2 in Lung and Breast Cancer Is a Potential Biomarker and Drug Target. Int. J. Mol. Sci. 2018, 19, 3096. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Shore, D.L.; Wilson, L.E.; Sanniez, E.I.; Kim, J.H.; Taylor, J.A.; Sandler, D.P. Lifetime use of nonsteroidal anti-inflammatory drugs and breast cancer risk: Results from a prospective study of women with a sister with breast cancer. BMC Cancer 2015, 15, 960. [Google Scholar] [CrossRef] [PubMed]
- Moris, D.; Kontos, M.; Spartalis, E.; Fentiman, I.S. The role of NSAIDs in breast cancer prevention and relapse: Current evidence and future perspectives. Breast Care 2016, 11, 339–344. [Google Scholar] [CrossRef]
- Orlowski, R.Z.; Kuhn, D.J. Proteasome inhibitors in cancer therapy: Lessons from the first decade. Clin. Cancer Res. 2008, 14, 1649–1657. [Google Scholar] [CrossRef]
- Liu, J.; Peng, Y.; Zhang, J.; Long, J.; Liu, J.; Wei, W. Targeting SCF E3 Ligases for Cancer Therapies. Adv. Exp. Med. Biol. 2020, 1217, 123–146. [Google Scholar] [CrossRef]
- Glickman, M.; Ciechanover, A. The ubiquitin-proteasome proteolytic pathway: Destruction for the sake of construction. Physiol. Rev. 2002, 82, 373–428. [Google Scholar] [CrossRef]
- Barnes, B.J.; Kellum, M.J.; Field, A.E.; Pitha, P.M. Multiple regulatory domains of IRF-5 control activation, cellular localization, and induction of chemokines that mediate recruitment of t lymphocytes. Mol. Cell. Biol. 2002, 22, 5721–5740. [Google Scholar] [CrossRef]
- Massimino, M.; Vigneri, P.G.; Fallica, M.; Fidilio, A.; Aloisi, A.; Frasca, F.; Manzella, L. IRF5 promotes the proliferation of human thyroid cancer cells. Mol. Cancer 2012, 11, 21. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 3. [Google Scholar] [CrossRef]
- Colaprico, A.; Silva, T.C.; Olsen, C.; Garofano, L.; Cava, C.; Garolini, D.; Sabedot, T.S.; Malta, T.M.; Pagnotta, S.M.; Castiglioni, I.; et al. TCGAbiolinks: An R/Bioconductor package for integrative analysis of TCGA data. Nucleic Acids Res. 2015, 44, e71. [Google Scholar] [CrossRef]











© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, S.-C.; Hsu, W.; Su, E.C.-Y.; Hung, C.-S.; Ding, J.L. Human FBXL8 Is a Novel E3 Ligase Which Promotes BRCA Metastasis by Stimulating Pro-Tumorigenic Cytokines and Inhibiting Tumor Suppressors. Cancers 2020, 12, 2210. https://doi.org/10.3390/cancers12082210
Chang S-C, Hsu W, Su EC-Y, Hung C-S, Ding JL. Human FBXL8 Is a Novel E3 Ligase Which Promotes BRCA Metastasis by Stimulating Pro-Tumorigenic Cytokines and Inhibiting Tumor Suppressors. Cancers. 2020; 12(8):2210. https://doi.org/10.3390/cancers12082210
Chicago/Turabian StyleChang, Shu-Chun, Wayne Hsu, Emily Chia-Yu Su, Chin-Sheng Hung, and Jeak Ling Ding. 2020. "Human FBXL8 Is a Novel E3 Ligase Which Promotes BRCA Metastasis by Stimulating Pro-Tumorigenic Cytokines and Inhibiting Tumor Suppressors" Cancers 12, no. 8: 2210. https://doi.org/10.3390/cancers12082210
APA StyleChang, S.-C., Hsu, W., Su, E. C.-Y., Hung, C.-S., & Ding, J. L. (2020). Human FBXL8 Is a Novel E3 Ligase Which Promotes BRCA Metastasis by Stimulating Pro-Tumorigenic Cytokines and Inhibiting Tumor Suppressors. Cancers, 12(8), 2210. https://doi.org/10.3390/cancers12082210