Targeted Gene Delivery Therapies for Cervical Cancer



Abstract
1. Introduction
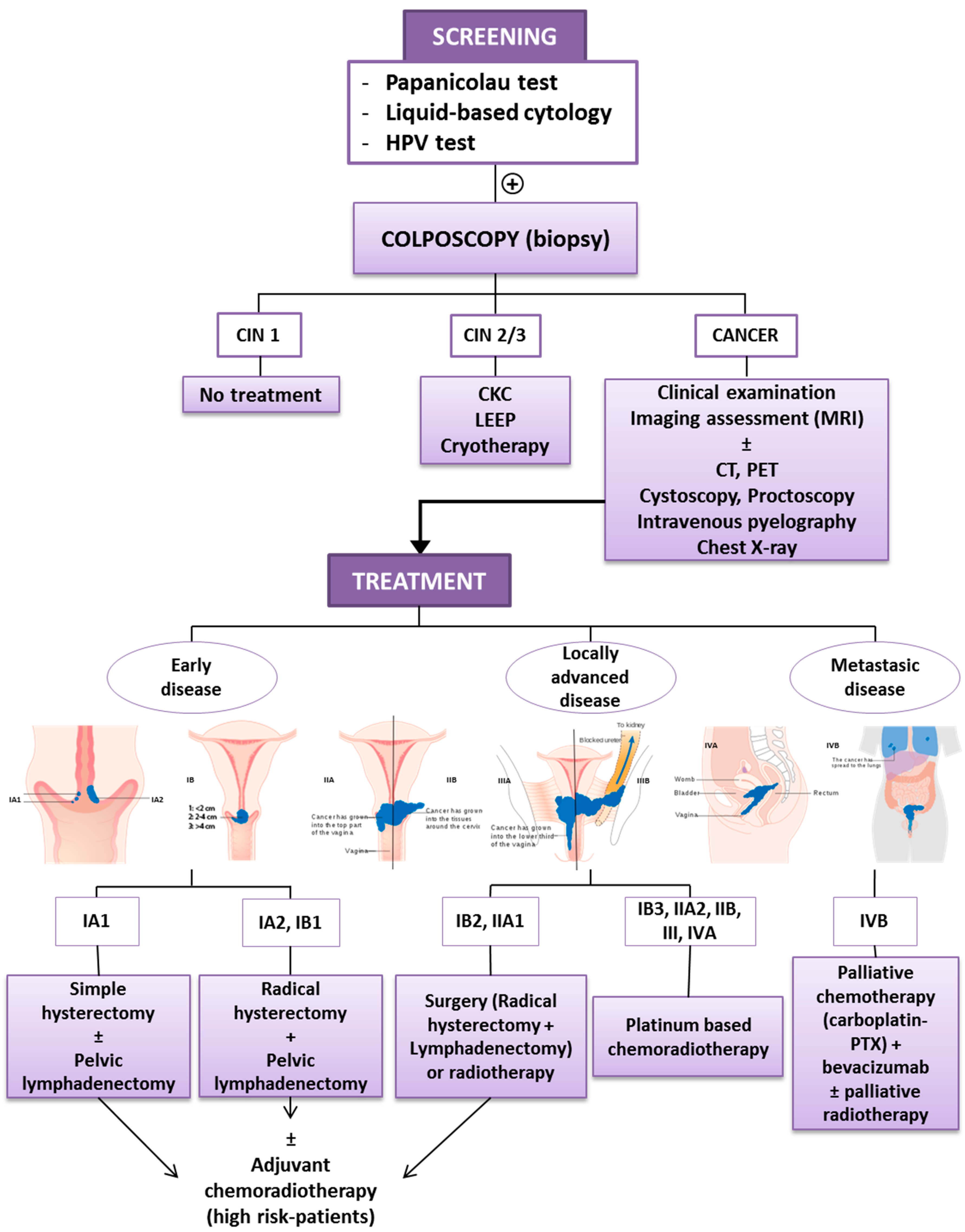
1.1. Diagnosis and Early Detection
1.2. Current Treatment of Cervical Cancer
1.3. New Approaches for the Treatment of Advanced Cervical Cancer
2. What’s New in Gene Targeted Therapies for Cervical Cancer?
2.1. Vectors Targeting Cervical Cancer
2.2. Targeted Therapy Strategies for Cervical Cancer Treatment
2.2.1. Therapy Targeting Compensation of Mutations
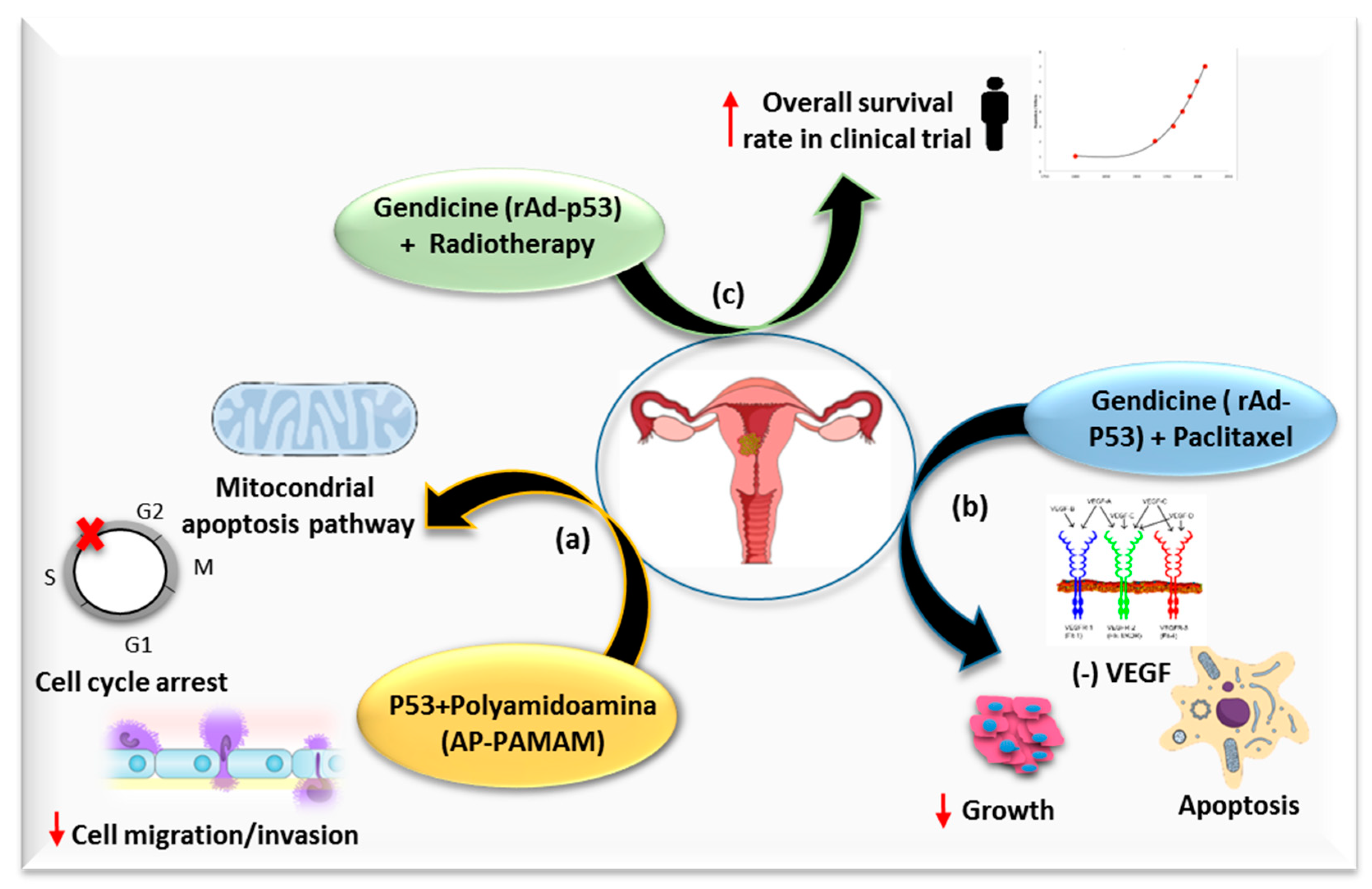
Tumor Suppressor Gene Restoration
Blocking Oncogenic Expression
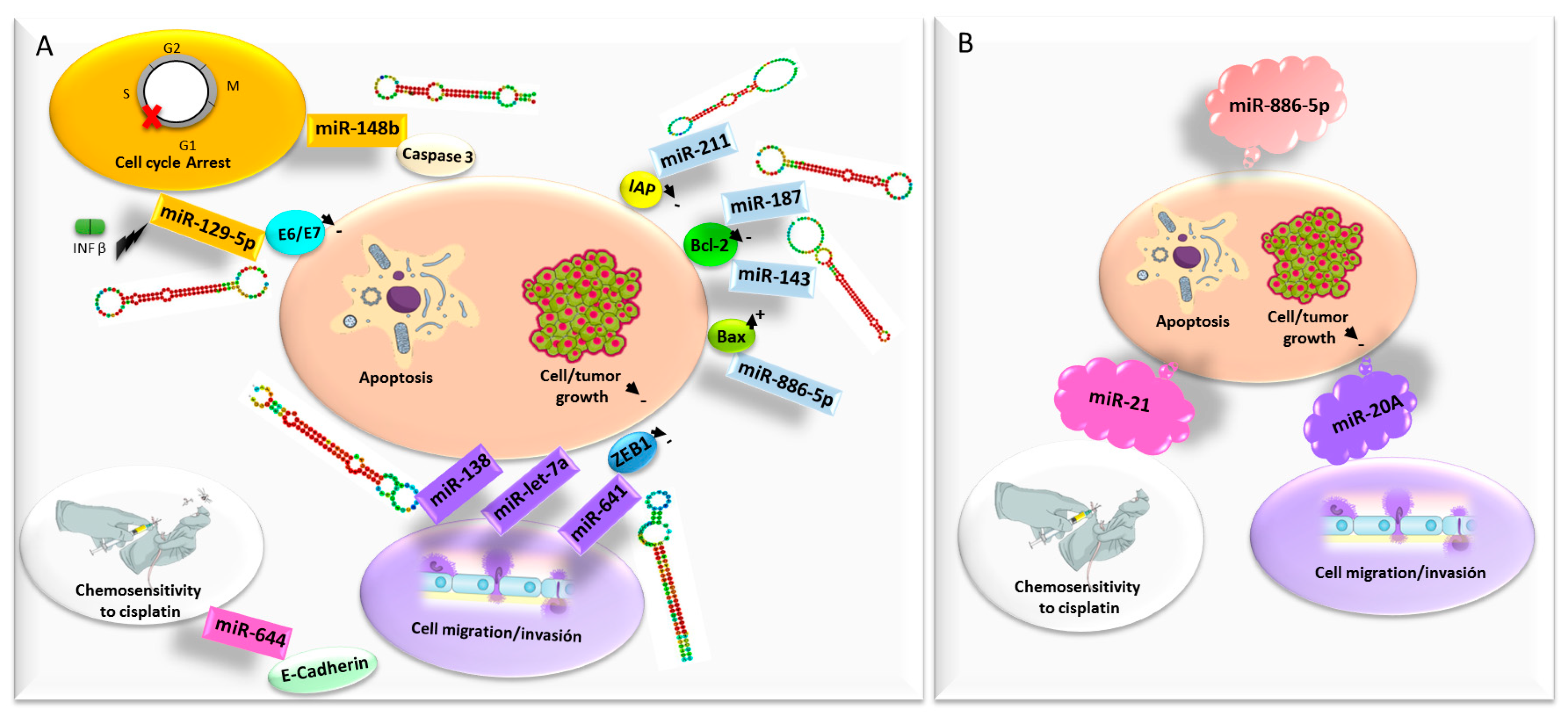
microRNAs
2.2.2. Suicide Gene Therapy
2.2.3. Oncolytic Virotherapy
2.2.4. Antiangiogenic Strategies
2.2.5. Genetic Immunopotentiation Therapies
- (i)
- Passive immunotherapy; consisting of ex-vivo activated cells or molecules that, once inside the body, compensate for impaired immune functions. This category includes the administration of elements of the immune system such as tumor-specific antibodies, recombinant cytokines or pre-activated effector immune cells.
- (ii)
- Active immunotherapy; which focuses on stimulation of host immune response in vivo to generate an antitumor response using vaccines, immunostimulatory cytokine among others [104].
2.2.6. Therapies Targeting Drug Resistance
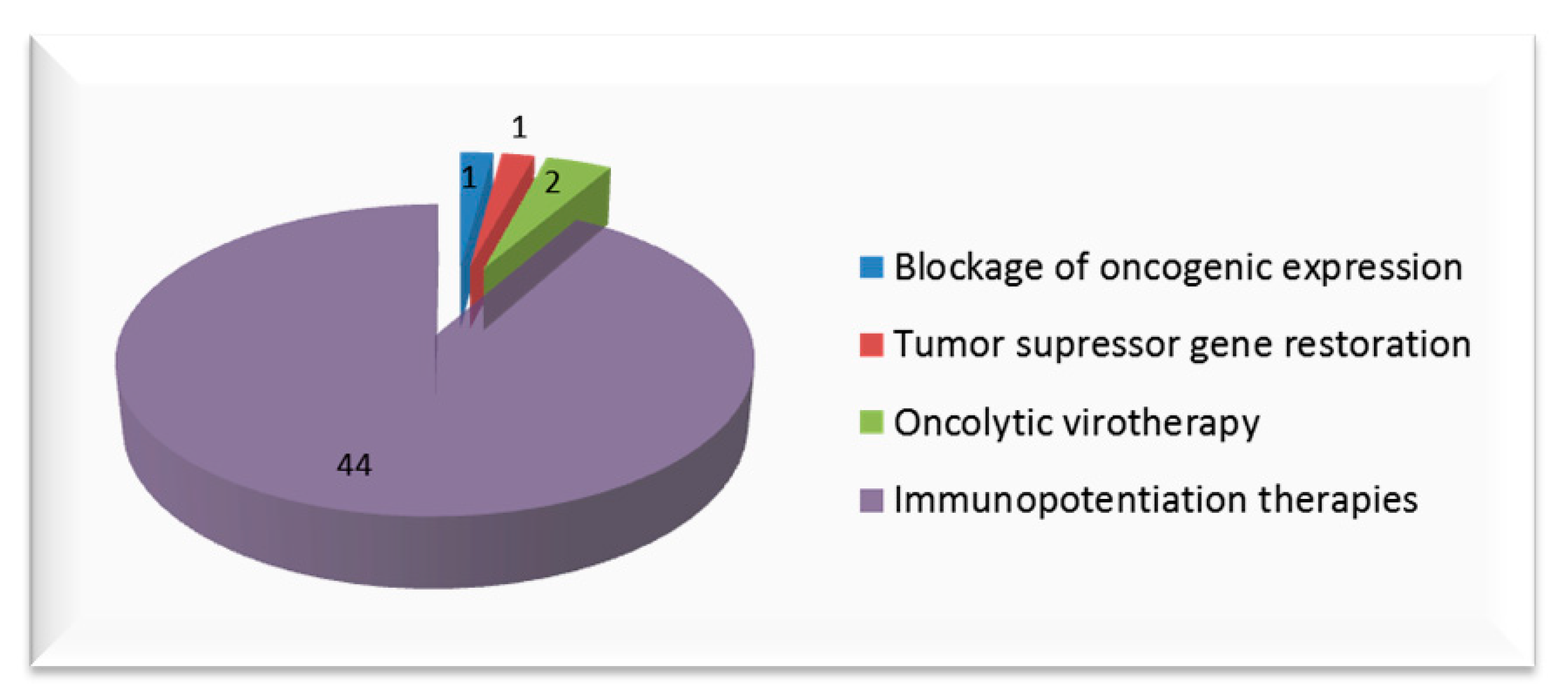
3. Clinical Trials
4. Future Directions
- (a)
- Apigenin, a dietary flavonoid that inhibits the self-renewal capacity of HeLa CSCs by the inactivation of casein kinase 2α [138];
- (b)
- Morusin, a natural compound isolated from the root bark of Morus australis with the ability to inhibit cervical CSCs growth and migration through NF-κB attenuation mediated apoptosis induction [139];
- (c)
- Phenethyl isothiocyanate, a dietary constituent with cytotoxic activity through TRAIL-mediated apoptotic pathways [140];
- (d)
- Doxycycline, a tetracycline used to treat a variety of infections that has the ability to inhibit proliferation and migration in HeLa-CSCs in addition to inducing apoptosis in vivo and in vitro [141];
- (e)
- Zolendronic acid, that has been found to induce apoptosis and arrest cell cycle in CSCs, in addition to achieving attenuation of the stemness phenotype, leading to the inhibition of cervical CSCs proliferation in vitro and in vivo [142];
- (f)
- A1E, a composition of 11 oriental medicinal plants that can inhibit CSCs and reduce the expression of stemness markers [143].
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018. [Google Scholar] [CrossRef] [PubMed]
- Small, W.; Bacon, M.A.; Bajaj, A.; Chuang, L.T.; Fisher, B.J.; Harkenrider, M.M.; Jhingran, A.; Kitchener, H.C.; Mileshkin, L.R.; Viswanathan, A.N.; et al. Cervical cancer: A global health crisis. Cancer 2017, 123, 2404–2412. [Google Scholar] [CrossRef] [PubMed]
- Schiffman, M.; Castle, P.E.; Jeronimo, J.; Rodriguez, A.C.; Wacholder, S. Human papillomavirus and cervical cancer. Lancet 2007, 370, 890–907. [Google Scholar] [CrossRef]
- Woodman, C.B.J.; Collins, S.I.; Young, L.S. The natural history of cervical HPV infection: Unresolved issues. Nat. Rev. Cancer 2007, 7, 11–22. [Google Scholar] [CrossRef]
- Petrosky, E.; Bocchini, J.A.; Hariri, S.; Chesson, H.; Curtis, C.R.; Saraiya, M.; Unger, E.R.; Markowitz, L.E. Centers for Disease Control and Prevention (CDC) Use of 9-valent human papillomavirus (HPV) vaccine: Updated HPV vaccination recommendations of the advisory committee on immunization practices. Morb. Mortal. Wkly. Rep. 2015, 64, 300–304. [Google Scholar]
- Sawaya, G.F.; Smith-McCune, K.; Kuppermann, M. Cervical Cancer Screening. JAMA 2019, 321, 2018. [Google Scholar] [CrossRef]
- Santesso, N.; Mustafa, R.A.; Schünemann, H.J.; Arbyn, M.; Blumenthal, P.D.; Cain, J.; Chirenje, M.; Denny, L.; De Vuyst, H.; Eckert, L.O.; et al. World Health Organization Guidelines for treatment of cervical intraepithelial neoplasia 2-3 and screen-and-treat strategies to prevent cervical cancer. Int. J. Gynaecol. Obstet. 2016, 132, 252–258. [Google Scholar] [CrossRef]
- Bhatla, N.; Aoki, D.; Sharma, D.N.; Sankaranarayanan, R. Cancer of the cervix uteri. Int. J. Gynaecol. Obstet. 2018, 143, 22–36. [Google Scholar] [CrossRef]
- Marth, C.; Landoni, F.; Mahner, S.; McCormack, M.; Gonzalez-Martin, A.; Colombo, N. ESMO Guidelines Committee Cervical cancer: ESMO Clinical Practice Guidelines for diagnosis, treatment and follow-up. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2017, 28, iv72–iv83. [Google Scholar] [CrossRef]
- Crusz, S.M.; Miller, R.E. Targeted therapies in gynaecological cancers. Histopathology 2020, 76, 157–170. [Google Scholar] [CrossRef]
- Chung, H.C.; Ros, W.; Delord, J.-P.; Perets, R.; Italiano, A.; Shapira-Frommer, R.; Manzuk, L.; Piha-Paul, S.A.; Xu, L.; Zeigenfuss, S.; et al. Efficacy and Safety of Pembrolizumab in Previously Treated Advanced Cervical Cancer: Results From the Phase II KEYNOTE-158 Study. J. Clin. Oncol. 2019, 37, 1470–1478. [Google Scholar] [CrossRef] [PubMed]
- Hollebecque, A.; Meyer, T.; Moore, K.N.; Machiels, J.-P.H.; De Greve, J.; López-Picazo, J.M.; Oaknin, A.; Kerger, J.N.; Boni, V.; Evans, T.R.J.; et al. An open-label, multicohort, phase I/II study of nivolumab in patients with virus-associated tumors (CheckMate 358): Efficacy and safety in recurrent or metastatic (R/M) cervical, vaginal, and vulvar cancers. J. Clin. Oncol. 2017, 35, 5504. [Google Scholar] [CrossRef]
- Monk, B.J.; Mayadev, J.; Nunes, A.T.; Dabrowska Brown, A.; Marcovitz, M.; Lanasa, M.C. CALLA: Efficacy and safety of durvalumab with and following concurrent chemoradiotherapy (CCRT) versus CCRT alone in women with locally advanced cervical cancer: A phase III, randomized, double-blind, multicenter study. J. Clin. Oncol. 2019, 37, TPS5597. [Google Scholar] [CrossRef]
- Mackay, H.J.; Tinker, A.; Winquist, E.; Thomas, G.; Swenerton, K.; Oza, A.; Sederias, J.; Ivy, P.; Eisenhauer, E.A. A phase II study of sunitinib in patients with locally advanced or metastatic cervical carcinoma: NCIC CTG Trial IND.184. Gynecol. Oncol. 2010, 116, 163–167. [Google Scholar] [CrossRef]
- Monk, B.J.; Mas Lopez, L.; Zarba, J.J.; Oaknin, A.; Tarpin, C.; Termrungruanglert, W.; Alber, J.A.; Ding, J.; Stutts, M.W.; Pandite, L.N. Phase II, Open-Label Study of Pazopanib or Lapatinib Monotherapy Compared With Pazopanib Plus Lapatinib Combination Therapy in Patients with Advanced and Recurrent Cervical Cancer. J. Clin. Oncol. 2010, 28, 3562–3569. [Google Scholar] [CrossRef]
- Symonds, R.P.; Gourley, C.; Davidson, S.; Carty, K.; McCartney, E.; Rai, D.; Banerjee, S.; Jackson, D.; Lord, R.; McCormack, M.; et al. Cediranib combined with carboplatin and paclitaxel in patients with metastatic or recurrent cervical cancer (CIRCCa): A randomised, double-blind, placebo-controlled phase 2 trial. Lancet Oncol. 2015, 16, 1515–1524. [Google Scholar] [CrossRef]
- American Cancer Society Survival Rates for Cervical Cancer. Available online: http://www.cancer.org/Cancer/CervicalCancer/DetailedGuide/cervical-cancer-survival (accessed on 26 April 2020).
- Áyen, Á.; Jiménez Martínez, Y.; Marchal, J.; Boulaiz, H. Recent Progress in Gene Therapy for Ovarian Cancer. Int. J. Mol. Sci. 2018, 19, 1930. [Google Scholar] [CrossRef]
- Kay, M.A. State-of-the-art gene-based therapies: The road ahead. Nat. Rev. Genet. 2011, 12, 316–328. [Google Scholar] [CrossRef]
- Sun, W.; Shi, Q.; Zhang, H.; Yang, K.; Ke, Y.; Wang, Y.; Qiao, L. Advances in the techniques and methodologies of cancer gene therapy. Discov. Med. 2019, 27, 45–55. [Google Scholar]
- Ibraheem, D.; Elaissari, A.; Fessi, H. Gene therapy and DNA delivery systems. Int. J. Pharm. 2014, 459, 70–83. [Google Scholar] [CrossRef]
- Boulaiz, H.; Marchal, J.A.; Prados, J.; Melguizo, C.; Aránega, A. Non-viral and viral vectors for gene therapy. Cell. Mol. Biol. (Noisy-le-grand) 2005, 51, 3–22. [Google Scholar] [PubMed]
- Boulaiz, H.; Alvarez, P.J.; Ramirez, A.; Marchal, J.A.; Prados, J.; Rodríguez-Serrano, F.; Perán, M.; Melguizo, C.; Aranega, A. Nanomedicine: Application Areas and Development Prospects. Int. J. Mol. Sci. 2011, 12, 3303–3321. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Moreno, P.; Ortega-Vinuesa, J.L.; Peula-Garcia, J.M.; Marchal, J.A.; Boulaiz, H. Smart Drug-Delivery Systems for Cancer Nanotherapy. Curr. Drug Targets 2018, 19, 339–359. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.; Rajasekaran, N.; Ju, W.; Shin, Y. Human Papillomavirus: Current and Future RNAi Therapeutic Strategies for Cervical Cancer. J. Clin. Med. 2015, 4, 1126–1155. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; He, L.; Liu, Y.; Xia, S.; Fang, A.; Xie, Y.; Gan, L.; He, Z.; Tan, X.; Jiang, C.; et al. Promising Nanocarriers for PEDF Gene Targeting Delivery to Cervical Cancer Cells Mediated by the Over-expressing FRα. Sci. Rep. 2016, 6, 32427. [Google Scholar] [CrossRef]
- Kim, H.-S.; Yi, B.-R.; Hwang, K.-A.; Kim, S.U.; Choi, K.-C. Anticancer effects of the engineered stem cells transduced with therapeutic genes via a selective tumor tropism caused by vascular endothelial growth factor toward HeLa cervical cancer cells. Mol. Cells 2013, 36, 347–354. [Google Scholar] [CrossRef]
- Bai, L.-X.; Ding, L.; Jiang, S.-W.; Kang, H.-J.; Gao, C.-F.; Chen, C.; Zhou, Q.; Wang, J.-T. Down-regulation of FRα Inhibits Proliferation and Promotes Apoptosis of Cervical Cancer Cells in Vitro. Asian Pac. J. Cancer Prev. 2014, 15, 5667–5672. [Google Scholar] [CrossRef]
- Wu, M.; Gunning, W.; Ratnam, M. Expression of folate receptor type alpha in relation to cell type, malignancy, and differentiation in ovary, uterus, and cervix. Cancer Epidemiol. Biomark. Prev. 1999, 8, 775–782. [Google Scholar] [PubMed]
- Rubinsak, L.A.; Cohen, C.; Khanna, N.; Horowitz, I.R.; Hanley, K.Z. Folate Receptor Alpha Expression in Platinum Resistant/Refractory Ovarian Carcinomas and Primary Endocervical Adenocarcinomas. Appl. Immunohistochem. Mol. Morphol. 2018, 26, 567–572. [Google Scholar] [CrossRef]
- Liu, B.; Han, S.-M.; Tang, X.-Y.; Han, L.; Li, C.-Z. Cervical cancer gene therapy by gene loaded PEG-PLA nanomedicine. Asian Pac. J. Cancer Prev. 2014, 15, 4915–4918. [Google Scholar] [CrossRef]
- Pillai, M.R. Expression of folate receptors and heterogeneous nuclear ribonucleoprotein E1 in women with human papillomavirus mediated transformation of cervical tissue to cancer. J. Clin. Pathol. 2003, 56, 569–574. [Google Scholar] [CrossRef] [PubMed]
- Bermúdez-Morales, V.H.; Fierros-Zarate, G.; García-Meléndrez, C.; Alcocer-Gonzalez, J.M.; Morales-Ortega, A.; Peralta-Zaragoza, O.; Torres-Poveda, K.; Burguete-García, A.I.; Hernández-Márquez, E.; Madrid-Marina, V. In vivo Antitumor Effect of an HPV-specific Promoter driving IL-12 Expression in an HPV 16-positive Murine Model of Cervical Cancer. J. Cancer 2016, 7, 1950–1959. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Mao, Y.; Zhang, F.; Ye, C.; Tong, H.; Su, Y.; Zhu, J. The inhibitory effect of a new scFv/tP protein as siRNA delivery system to target hWAPL in cervical carcinoma. Mol. Cell. Biochem. 2014, 391, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Pal, A.; Kundu, R. Human Papillomavirus E6 and E7: The Cervical Cancer Hallmarks and Targets for Therapy. Front. Microbiol. 2020, 10, 3116. [Google Scholar] [CrossRef]
- Moody, C.A.; Laimins, L.A. Human papillomavirus oncoproteins: Pathways to transformation. Nat. Rev. Cancer 2010, 10, 550–560. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Chen, W.; Yang, J.; Liang, X.; Wang, Y.; Li, Q.; Yang, Y.; Li, K. Inhibition of cell proliferation and migration through nucleobase-modified polyamidoamine-mediated p53 delivery. Int. J. Nanomed. 2018, 13, 1297–1311. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-G.; Zheng, X.-L.; Liu, F.-M. The mechanism and inhibitory effect of recombinant human P53 adenovirus injection combined with paclitaxel on human cervical cancer cell HeLa. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 1037–1042. [Google Scholar] [PubMed]
- Su, X.; Chen, W.-J.; Xiao, S.-W.; Li, X.-F.; Xu, G.; Pan, J.-J.; Zhang, S.-W. Effect and Safety of Recombinant Adenovirus-p53 Transfer Combined with Radiotherapy on Long-Term Survival of Locally Advanced Cervical Cancer. Hum. Gene Ther. 2016, 27, 1008–1014. [Google Scholar] [CrossRef]
- Cheng, H.Y.; Zhang, T.; Qu, Y.; Shi, W.J.; Lou, G.; Liu, Y.X.; Zhang, Y.Y.; Cheng, L. Synergism between RIZ1 gene therapy and paclitaxel in SiHa cervical cancer cells. Cancer Gene Ther. 2016, 23, 392–395. [Google Scholar] [CrossRef]
- Yang, S.; Xing, L.; Gu, L.; Cheng, H.; Feng, Y.; Zhang, Y. Combination of RIZ1 Overexpression and Radiotherapy Contributes to Apoptosis and DNA Damage of HeLa and SiHa Cervical Cancer Cells. Basic Clin. Pharmacol. Toxicol. 2018, 123, 137–146. [Google Scholar] [CrossRef]
- Yan, C.-M.; Zhao, Y.-L.; Cai, H.-Y.; Miao, G.-Y.; Ma, W. Blockage of PTPRJ promotes cell growth and resistance to 5-FU through activation of JAK1/STAT3 in the cervical carcinoma cell line C33A. Oncol. Rep. 2015, 33, 1737–1744. [Google Scholar] [CrossRef] [PubMed]
- Bonetta, A.C.; Mailly, L.; Robinet, E.; Travé, G.; Masson, M.; Deryckere, F. Artificial microRNAs against the viral E6 protein provoke apoptosis in HPV positive cancer cells. Biochem. Biophys. Res. Commun. 2015, 465, 658–664. [Google Scholar] [CrossRef] [PubMed]
- Sato, N.; Saga, Y.; Uchibori, R.; Tsukahara, T.; Urabe, M.; Kume, A.; Fujiwara, H.; Suzuki, M.; Ozawa, K.; Mizukami, H. Eradication of cervical cancer in vivo by an AAV vector that encodes shRNA targeting human papillomavirus type 16 E6/E7. Int. J. Oncol. 2018, 52, 687–696. [Google Scholar] [CrossRef]
- Reschner, A.; Bontems, S.; Le Gac, S.; Lambermont, J.; Marcélis, L.; Defrancq, E.; Hubert, P.; Moucheron, C.; Kirsch-De Mesmaeker, A.; Raes, M.; et al. Ruthenium oligonucleotides, targeting HPV16 E6 oncogene, inhibit the growth of cervical cancer cells under illumination by a mechanism involving p53. Gene Ther. 2013, 20, 435–443. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Yu, L.; Zhu, D.; Ding, W.; Wang, X.; Zhang, C.; Wang, L.; Jiang, X.; Shen, H.; He, D.; et al. Disruption of HPV16-E7 by CRISPR/Cas system induces apoptosis and growth inhibition in HPV16 positive human cervical cancer cells. Biomed. Res. Int. 2014, 2014, 612823. [Google Scholar] [CrossRef]
- Hu, Z.; Ding, W.; Zhu, D.; Yu, L.; Jiang, X.; Wang, X.; Zhang, C.; Wang, L.; Ji, T.; Liu, D.; et al. TALEN-mediated targeting of HPV oncogenes ameliorates HPV-related cervical malignancy. J. Clin. Invest. 2015, 125, 425–436. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Peng, C.; Li, B.; Wang, F.; Zhou, C.; Hong, D.; Ye, F.; Cheng, X.; Lü, W.; Xie, X. Transcriptional gene silencing of HPV16 E6/E7 induces growth inhibition via apoptosis in vitro and in vivo. Gynecol. Oncol. 2012, 124, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Lechanteur, A.; Furst, T.; Evrard, B.; Delvenne, P.; Piel, G.; Hubert, P. Promoting Vaginal Distribution of E7 and MCL-1 siRNA-Silencing Nanoparticles for Cervical Cancer Treatment. Mol. Pharm. 2017, 14, 1706–1717. [Google Scholar] [CrossRef]
- Au, W.W.; Abdou-Salama, S.; Al-Hendy, A. Inhibition of growth of cervical cancer cells using a dominant negative estrogen receptor gene. Gynecol. Oncol. 2007, 104, 276–280. [Google Scholar] [CrossRef]
- Heo, M.Y.; Salama, S.A.; Khatoon, N.; Al-Hendy, A.; Au, W.W. Abrogation of estrogen receptor signaling augments cytotoxicity of anticancer drugs on CaSki cervical cancer cells. Anticancer Res. 2008, 28, 2181–2187. [Google Scholar]
- Wang, P.; Yin, T.; Li, J.; Zheng, B.; Wang, X.; Wang, Y.; Zheng, J.; Zheng, R.; Shuai, X. Ultrasound-responsive microbubbles for sonography-guided siRNA delivery. Nanomedicine 2016, 12, 1139–1149. [Google Scholar] [CrossRef] [PubMed]
- Nowak, E.; Galilejczyk, A.; Sypniewski, D.; Bednarek, I. MMP-9 directed shRNAs as relevant inhibitors of matrix metalloproteinase 9 activity and signaling. Postepy Hig. Med. Dosw. (Online) 2013, 67, 742–749. [Google Scholar] [CrossRef] [PubMed]
- Ying, T.-H.; Lee, C.-H.; Chiou, H.-L.; Yang, S.-F.; Lin, C.-L.; Hung, C.-H.; Tsai, J.-P.; Hsieh, Y.-H. Knockdown of Pentraxin 3 suppresses tumorigenicity and metastasis of human cervical cancer cells. Sci. Rep. 2016, 6, 29385. [Google Scholar] [CrossRef] [PubMed]
- Lv, X.-F.; Hong, H.-Q.; Liu, L.; Cui, S.-H.; Ren, C.-C.; Li, H.-Y.; Zhang, X.-A.; Zhang, L.-D.; Wei, T.-X.; Liu, J.-J.; et al. RNAi-mediated downregulation of asparaginase-like protein 1 inhibits growth and promotes apoptosis of human cervical cancer line SiHa. Mol. Med. Rep. 2018, 18, 931–937. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Xing, L. RNAi gene therapy of SiHa cells via targeting human TERT induces growth inhibition and enhances radiosensitivity. Int. J. Oncol. 2013, 43, 1228–1234. [Google Scholar] [CrossRef][Green Version]
- Shi, Y.-A.; Zhao, Q.; Zhang, L.-H.; Du, W.; Wang, X.-Y.; He, X.; Wu, S.; Li, Y.-L. Knockdown of hTERT by siRNA inhibits cervical cancer cell growth in vitro and in vivo. Int. J. Oncol. 2014, 45, 1216–1224. [Google Scholar] [CrossRef]
- Zhou, S.; Xiao, Y.; Zhuang, Y.; Liu, Y.; Zhao, H.; Yang, H.; Xie, C.; Zhou, F.; Zhou, Y. Knockdown of homeobox containing 1 increases the radiosensitivity of cervical cancer cells through telomere shortening. Oncol. Rep. 2017, 38, 515–521. [Google Scholar] [CrossRef]
- Matsushita, K.; Shimada, H.; Ueda, Y.; Inoue, M.; Hasegawa, M.; Tomonaga, T.; Matsubara, H.; Nomura, F. Non-transmissible Sendai virus vector encoding c-myc suppressor FBP-interacting repressor for cancer therapy. World J. Gastroenterol. 2014, 20, 4316–4328. [Google Scholar] [CrossRef]
- Hollstein, M.; Sidransky, D.; Vogelstein, B.; Harris, C.C. p53 mutations in human cancers. Science 1991, 253, 49–53. [Google Scholar] [CrossRef]
- Doorbar, J. Molecular biology of human papillomavirus infection and cervical cancer. Clin. Sci. 2006, 110, 525–541. [Google Scholar] [CrossRef]
- Tan, S.; Hougardy, B.M.T.; Meersma, G.J.; Schaap, B.; de Vries, E.G.E.; van der Zee, A.G.J.; de Jong, S. Human papilloma virus 16 E6 RNA interference enhances cisplatin and death receptor-mediated apoptosis in human cervical carcinoma cells. Mol. Pharmacol. 2012, 81, 701–709. [Google Scholar] [CrossRef]
- Ajay, A.K.; Upadhyay, A.K.; Singh, S.; Vijayakumar, M.V.; Kumari, R.; Pandey, V.; Boppana, R.; Bhat, M.K. Cdk5 phosphorylates non-genotoxically overexpressed p53 following inhibition of PP2A to induce cell cycle arrest/apoptosis and inhibits tumor progression. Mol. Cancer 2010, 9, 204. [Google Scholar] [CrossRef] [PubMed]
- Tornesello, M.L.; Faraonio, R.; Buonaguro, L.; Annunziata, C.; Starita, N.; Cerasuolo, A.; Pezzuto, F.; Tornesello, A.L.; Buonaguro, F.M. The Role of microRNAs, Long Non-coding RNAs, and Circular RNAs in Cervical Cancer. Front. Oncol. 2020, 10. [Google Scholar] [CrossRef]
- He, C.Y.; Yang, J. miR-187 induces apoptosis of SiHa cervical carcinoma cells by downregulating Bcl-2. Genet. Mol. Res. 2017, 16. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.; Luo, R.; Chen, X.; Zhao, Y.; Tan, A. miR-187 inhibits the growth of cervical cancer cells by targeting FGF9. Oncol. Rep. 2017, 38, 1977–1984. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Yu, X.; Guo, X.; Tian, Z.; Su, M.; Long, Y.; Huang, C.; Zhou, F.; Liu, M.; Wu, X.; et al. miR-143 is downregulated in cervical cancer and promotes apoptosis and inhibits tumor formation by targeting Bcl-2. Mol. Med. Rep. 2012, 5, 753–760. [Google Scholar] [CrossRef] [PubMed]
- Mou, Z.; Xu, X.; Dong, M.; Xu, J. MicroRNA-148b Acts as a Tumor Suppressor in Cervical Cancer by Inducing G1/S-Phase Cell Cycle Arrest and Apoptosis in a Caspase-3-Dependent Manner. Med. Sci. Monit. 2016, 22, 2809–2815. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-L.; Wang, G.-Q.; Cui, H.-X.; Li, X.-X.; Xu, Z.-L.; Wang, X.-Y. miRNA211 induces apoptosis of cervical cancer SiHa cells via down-regulation of inhibitor of apoptosis proteins. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 336–342. [Google Scholar]
- Zhou, N.; Fei, D.; Zong, S.; Zhang, M.; Yue, Y. MicroRNA-138 inhibits proliferation, migration and invasion through targeting hTERT in cervical cancer. Oncol. Lett. 2016, 12, 3633–3639. [Google Scholar] [CrossRef]
- Yao, R.; Zheng, H.; Wu, L.; Cai, P. miRNA-641 inhibits the proliferation, migration, and invasion and induces apoptosis of cervical cancer cells by directly targeting ZEB1. OncoTargets Ther. 2018, 11, 8965–8976. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, H.; Wang, X.; Chen, L. Up-regulation of microRNA-664 inhibits cell growth and increases cisplatin sensitivity in cervical cancer. Int. J. Clin. Exp. Med. 2015, 8, 18123–18129. [Google Scholar] [PubMed]
- Guo, M.; Zhao, X.; Yuan, X.; Jiang, J.; Li, P. MiR-let-7a inhibits cell proliferation, migration, and invasion by down-regulating PKM2 in cervical cancer. Oncotarget 2017, 8, 28226–28236. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, S.; Yan, Q.; Chen, X.; Yang, Y.; Liu, X.; Wan, X. Interferon-β induced microRNA-129-5p down-regulates HPV-18 E6 and E7 viral gene expression by targeting SP1 in cervical cancer cells. PLoS ONE 2013, 8, e81366. [Google Scholar] [CrossRef] [PubMed]
- Peralta-Zaragoza, O.; Deas, J.; Meneses-Acosta, A.; De la O-Gómez, F.; Fernández-Tilapa, G.; Gómez-Cerón, C.; Benítez-Boijseauneau, O.; Burguete-García, A.; Torres-Poveda, K.; Bermúdez-Morales, V.H.; et al. Relevance of miR-21 in regulation of tumor suppressor gene PTEN in human cervical cancer cells. BMC Cancer 2016, 16, 215. [Google Scholar] [CrossRef]
- Chen, B.; Chen, X.; Wu, X.; Wang, X.; Wang, Y.; Lin, T.-Y.; Kurata, J.; Wu, J.; Vonderfecht, S.; Sun, G.; et al. Disruption of microRNA-21 by TALEN leads to diminished cell transformation and increased expression of cell-environment interaction genes. Cancer Lett. 2015, 356, 506–516. [Google Scholar] [CrossRef]
- Li, J.-H.; Xiao, X.; Zhang, Y.-N.; Wang, Y.-M.; Feng, L.-M.; Wu, Y.-M.; Zhang, Y.-X. MicroRNA miR-886-5p inhibits apoptosis by down-regulating Bax expression in human cervical carcinoma cells. Gynecol. Oncol. 2011, 120, 145–151. [Google Scholar] [CrossRef]
- Kang, H.-W.; Wang, F.; Wei, Q.; Zhao, Y.-F.; Liu, M.; Li, X.; Tang, H. miR-20a promotes migration and invasion by regulating TNKS2 in human cervical cancer cells. FEBS Lett. 2012, 586, 897–904. [Google Scholar] [CrossRef]
- Panghal, A.; Sharma, H.; Flora, S.; Naqvi, S. Suicide gene therapy: A promising approach towards gene delivery. Front. Nanosci. Nanotechnol. 2019, 5, 1–9. [Google Scholar] [CrossRef]
- Navarro, S.A.; Carrillo, E.; Griñán-Lisón, C.; Martín, A.; Perán, M.; Marchal, J.A.; Boulaiz, H. Cancer suicide gene therapy: A patent review. Expert Opin. Ther. Pat. 2016, 26, 1095–1104. [Google Scholar] [CrossRef]
- Hao, Y.; Guo, L.; Abudula, A.; Saidoula, W.; Guo, X. Proliferation inhibition and apoptosis enhancement of human cervical cancer cells by ultrasound-targeted microbubble destruction delivered double suicide genes. Int. J. Clin. Exp. Med. 2014, 7, 5330–5335. [Google Scholar]
- Teng, G.; Ju, Y.; Yang, Y.; Hua, H.; Chi, J.; Mu, X. Combined antitumor activity of the nitroreductase/CB1954 suicide gene system and γ-rays in HeLa cells in vitro. Mol. Med. Rep. 2016, 14, 5164–5170. [Google Scholar] [CrossRef]
- Narayanan, S.; Sanpui, P.; Sahoo, L.; Ghosh, S.S. Unravelling the potential of a new uracil phosphoribosyltransferase (UPRT) from Arabidopsis thaliana in sensitizing HeLa cells towards 5-fluorouracil. Int. J. Biol. Macromol. 2016, 91, 310–316. [Google Scholar] [CrossRef] [PubMed]
- Preston, M.A.; Pimentel, B.; Bermejo-Rodríguez, C.; Dionne, I.; Turnbull, A.; de la Cueva-Méndez, G. Repurposing a Prokaryotic Toxin-Antitoxin System for the Selective Killing of Oncogenically Stressed Human Cells. ACS Synth. Biol. 2016, 5, 540–546. [Google Scholar] [CrossRef] [PubMed]
- Qiu, N.; Gao, J.; Liu, Q.; Wang, J.; Shen, Y. Enzyme-Responsive Charge-Reversal Polymer-Mediated Effective Gene Therapy for Intraperitoneal Tumors. Biomacromolecules 2018, 19, 2308–2319. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Chen, H.; Zeng, X.; Liu, Z.; Xiao, X.; Zhu, Y.; Gu, D.; Mei, L. Surface modification of TPGS-b-(PCL-ran-PGA) nanoparticles with polyethyleneimine as a co-delivery system of TRAIL and endostatin for cervical cancer gene therapy. Nanoscale Res. Lett. 2013, 8, 161. [Google Scholar] [CrossRef]
- Chen, Y.; Zhuang, H.; Chen, X.; Shi, Z.; Wang, X. Spermidine-induced growth inhibition and apoptosis via autophagic activation in cervical cancer. Oncol. Rep. 2018, 39, 2845–2854. [Google Scholar] [CrossRef]
- Li, W.Y.; Li, Q.W.; Han, Z.S.; Jiang, Z.L.; Yang, H.; Li, J.; Zhang, X.B. Growth suppression effects of recombinant adenovirus expressing human lactoferrin on cervical cancer in vitro and in vivo. Cancer Biother. Radiopharm. 2011, 26, 477–483. [Google Scholar] [CrossRef]
- Singh, P.K.; Tiwari, A.K.; Rajmani, R.S.; Kumar, G.R.; Chaturvedi, U.; Saxena, L.; Saxena, S.; Doley, J.; Sahoo, A.P.; Santra, L.; et al. Apoptin as a potential viral gene oncotherapeutic agent. Appl. Biochem. Biotechnol. 2015, 176, 196–212. [Google Scholar] [CrossRef]
- Yang, M.; Wang, M.; Li, X.; Xie, Y.; Xia, X.; Tian, J.; Zhang, K.; Chen, F.; Song, H.; Dong, Z.; et al. Inhibition of constructed SEC3-ES lentiviral vector to proliferation, migration of Hela cells. Pathol. Res. Pract. 2019, 215, 315–321. [Google Scholar] [CrossRef]
- Shao, G.; Qian, D.; Wang, H.; Yan, Z.; Hu, M.; Wang, T.; Wang, B. Construction of the plasmid coding for the expression of the EGFP-M-IL-2(88Arg, 125Ala) fusion protein and the anti-tumor effects exerted by the fusion protein in HeLa-60 cells. Oncol. Lett. 2015, 9, 2729–2735. [Google Scholar] [CrossRef]
- Chen, D.; Tang, Q. An experimental study on cervix cancer with combination of HSV-TK/GCV suicide gene therapy system and 60Co radiotherapy. BMC Cancer 2010, 10, 609. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Liao, Z.-K.; Sun, W.-J.; Huang, C.; Xiong, J.; Zhou, F.-X.; Xie, C.-H.; Zhou, Y.-F. Enhanced suicide gene therapy using a tumor-specific promoter in combination with cisplatin. Mol. Med. Rep. 2009, 2, 1017–1022. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bai, Y.; Hui, P.; Du, X.; Su, X. Updates to the antitumor mechanism of oncolytic virus. Thorac. Cancer 2019, 10, 1031–1035. [Google Scholar] [CrossRef] [PubMed]
- Le Boeuf, F.; Niknejad, N.; Wang, J.; Auer, R.; Weberpals, J.I.; Bell, J.C.; Dimitroulakos, J. Sensitivity of cervical carcinoma cells to vesicular stomatitis virus-induced oncolysis: Potential role of human papilloma virus infection. Int. J. cancer 2012, 131, E204–E215. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Xia, X.; Wang, S.; Sima, N.; Li, Y.; Han, Z.; Gao, Q.; Luo, A.; Li, K.; Meng, L.; et al. Oncolytic adenovirus armed with human papillomavirus E2 gene in combination with radiation demonstrates synergistic enhancements of antitumor efficacy. Cancer Gene Ther. 2011, 18, 825–836. [Google Scholar] [CrossRef]
- Xiao, T.; Fan, J.K.; Huang, H.L.; Gu, J.F.; Li, L.-Y.; Liu, X.Y. VEGI-armed oncolytic adenovirus inhibits tumor neovascularization and directly induces mitochondria-mediated cancer cell apoptosis. Cell Res. 2010, 20, 367–378. [Google Scholar] [CrossRef]
- Han, X.; Wang, S.; Zhou, W.; Li, Y.; Lei, W.; Lv, W. Synergistic combination of histone deacetylase inhibitor suberoylanilide hydroxamic acid and oncolytic adenovirus ZD55-TRAIL as a therapy against cervical cancer. Mol. Med. Rep. 2015, 12, 435–441. [Google Scholar] [CrossRef][Green Version]
- Wang, W.; Sima, N.; Kong, D.; Luo, A.; Gao, Q.; Liao, S.; Li, W.; Han, L.; Wang, J.; Wang, S.; et al. Selective targeting of HPV-16 E6/E7 in cervical cancer cells with a potent oncolytic adenovirus and its enhanced effect with radiotherapy in vitro and vivo. Cancer Lett. 2010, 291, 67–75. [Google Scholar] [CrossRef]
- Wang, H.; Song, X.; Zhang, H.; Zhang, J.; Shen, X.; Zhou, Y.; Fan, X.; Dai, L.; Qian, G.; Hoffman, A.R.; et al. Potentiation of tumor radiotherapy by a radiation-inducible oncolytic and oncoapoptotic adenovirus in cervical cancer xenografts. Int. J. Cancer 2012, 130, 443–453. [Google Scholar] [CrossRef]
- Cheng, Y.; Jiang, S.; Yuan, J.; Liu, J.; Simoncini, T. Vascular endothelial growth factor C promotes cervical cancer cell invasiveness via regulation of microRNA-326/cortactin expression. Gynecol. Endocrinol. 2018, 34, 853–858. [Google Scholar] [CrossRef]
- Qi, L.; Xing, L.N.; Wei, X.; Song, S.G. Effects of VEGF suppression by small hairpin RNA interference combined with radiotherapy on the growth of cervical cancer. Genet. Mol. Res. 2014, 13, 5094–5106. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xia, P.; Zhang, W.; Yan, M.; Xiong, X.; Yu, W.; Song, E. Short interfering RNA targeting Net1 reduces the angiogenesis and tumor growth of in vivo cervical squamous cell carcinoma through VEGF down-regulation. Hum. Pathol. 2017, 65, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Papaioannou, N.E.; Beniata, O.V.; Vitsos, P.; Tsitsilonis, O.; Samara, P. Harnessing the immune system to improve cancer therapy. Ann. Transl. Med. 2016, 4, 261. [Google Scholar] [CrossRef] [PubMed]
- Porter, S.S.; Stepp, W.H.; Stamos, J.D.; McBride, A.A. Host cell restriction factors that limit transcription and replication of human papillomavirus. Virus Res. 2017, 231, 10–20. [Google Scholar] [CrossRef]
- Ping, Y.; Liu, C.; Zhang, Y. T-cell receptor-engineered T cells for cancer treatment: Current status and future directions. Protein Cell 2018, 9, 254–266. [Google Scholar] [CrossRef]
- Jin, B.Y.; Campbell, T.E.; Draper, L.M.; Stevanović, S.; Weissbrich, B.; Yu, Z.; Restifo, N.P.; Rosenberg, S.A.; Trimble, C.L.; Hinrichs, C.S. Engineered T cells targeting E7 mediate regression of human papillomavirus cancers in a murine model. JCI Insight 2018, 3, e99488. [Google Scholar] [CrossRef]
- Hamza, T.; Barnett, J.B.; Li, B. Interleukin 12 a Key Immunoregulatory Cytokine in Infection Applications. Int. J. Mol. Sci. 2010, 11, 789–806. [Google Scholar] [CrossRef]
- García Paz, F.; Madrid Marina, V.; Morales Ortega, A.; Santander González, A.; Peralta Zaragoza, O.; Burguete García, A.; Torres Poveda, K.; Moreno, J.; Alcocer González, J.; Hernandez Marquez, E.; et al. The relationship between the antitumor effect of the IL-12 gene therapy and the expression of Th1 cytokines in an HPV16-positive murine tumor model. Mediators Inflamm. 2014, 2014, 510846. [Google Scholar] [CrossRef]
- Wang, L.; Chen, P.; Luo, S.; Li, J.; Liu, K.; Hu, H.; Wei, Y. CXC-chemokine-ligand-10 gene therapy efficiently inhibits the growth of cervical carcinoma on the basis of its anti-angiogenic and antiviral activity. Biotechnol. Appl. Biochem. 2009, 53, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.-L.; Wang, B.-Q.; Chen, L.-L.; Luo, H.-Q.; Wu, J.-B. CXCL10 enhances radiotherapy effects in HeLa cells through cell cycle redistribution. Oncol. Lett. 2012, 3, 383–386. [Google Scholar] [CrossRef]
- Zhao, M.; Ma, Q.; Xu, J.; Fu, S.; Chen, L.; Wang, B.; Wu, J.; Yang, L. Combining CXCL10 gene therapy and radiotherapy improved therapeutic efficacy in cervical cancer HeLa cell xenograft tumor models. Oncol. Lett. 2015, 10, 768–772. [Google Scholar] [CrossRef] [PubMed]
- Yiang, G.-T.; Harn, H.-J.; Yu, Y.-L.; Hu, S.-C.; Hung, Y.-T.; Hsieh, C.-J.; Lin, S.-Z.; Wei, C.-W. Immunotherapy: rAAV2 expressing interleukin-15 inhibits HeLa cell tumor growth in mice. J. Biomed. Sci. 2009, 16, 47. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Peng, S.; Qiu, J.; Miao, J.; Yang, B.; Jeang, J.; Hung, C.-F.; Wu, T.-C. Intravaginal HPV DNA vaccination with electroporation induces local CD8+ T-cell immune responses and antitumor effects against cervicovaginal tumors. Gene Ther. 2015, 22, 528–535. [Google Scholar] [CrossRef]
- Garza-Morales, R.; Perez-Trujillo, J.J.; Martinez-Jaramillo, E.; Saucedo-Cardenas, O.; Loera-Arias, M.J.; Garcia-Garcia, A.; Rodriguez-Rocha, H.; Yolcu, E.; Shirwan, H.; Gomez-Gutierrez, J.G.; et al. A DNA Vaccine Encoding SA-4-1BBL Fused to HPV-16 E7 Antigen Has Prophylactic and Therapeutic Efficacy in a Cervical Cancer Mouse Model. Cancers (Basel) 2019, 11, 588. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Peng, S.; Yang, A.; Farmer, E.; Wu, T.-C.; Hung, C.-F. Coinjection of IL2 DNA enhances E7-specific antitumor immunity elicited by intravaginal therapeutic HPV DNA vaccination with electroporation. Gene Ther. 2017, 24, 408–415. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, L.; Teague, J.E.; Morrow, M.P.; Jotova, I.; Wu, T.C.; Wang, C.; Desmarais, C.; Boyer, J.D.; Tycko, B.; Robins, H.S.; et al. Intramuscular therapeutic vaccination targeting HPV16 induces T cell responses that localize in mucosal lesions. Sci. Transl. Med. 2014, 6, 221ra13. [Google Scholar] [CrossRef]
- Çuburu, N.; Khan, S.; Thompson, C.D.; Kim, R.; Vellinga, J.; Zahn, R.; Lowy, D.R.; Scheper, G.; Schiller, J.T. Adenovirus vector-based prime-boost vaccination via heterologous routes induces cervicovaginal CD8+ T cell responses against HPV16 oncoproteins. Int. J. Cancer 2018, 142, 1467–1479. [Google Scholar] [CrossRef]
- Maitituoheti, M.; Li, Y.; Wang, W.; Wang, W.; Han, L.; Yang, R.; Wang, T.; Wu, Z.; Ma, D.; Wang, S. Adeno-associated virus-mediated local delivery of LIGHT suppresses tumorigenesis in a murine cervical cancer model. J. Immunother. 2011, 34, 581–587. [Google Scholar] [CrossRef]
- Jeon, Y.H.; Lee, H.W.; Lee, Y.L.; Kim, J.E.; Hwang, M.-H.; Jeong, S.Y.; Lee, S.-W.; Ahn, B.-C.; Ha, J.-H.; Lee, J. Combined E7-dendritic cell-based immunotherapy and human sodium/iodide symporter radioiodine gene therapy with monitoring of antitumor effects by bioluminescent imaging in a mouse model of uterine cervical cancer. Cancer Biother. Radiopharm. 2011, 26, 671–679. [Google Scholar] [CrossRef]
- Zhang, M.; Shi, S.; Guo, R.; Miao, Y.; Li, B. Use of rhenium-188 for in vivo imaging and treatment of human cervical cancer cells transfected with lentivirus expressing sodium iodide symporter. Oncol. Rep. 2016, 36, 2289–2297. [Google Scholar] [CrossRef]
- Zheng, Y.; Hu, B.; Xie, S.; Chen, X.; Hu, Y.; Chen, W.; Li, S.; Hu, B. Dendritic cells infected by Ad-sh-SOCS1 enhance cytokine-induced killer (CIK) cell immunotherapeutic efficacy in cervical cancer models. Cytotherapy 2017, 19, 617–628. [Google Scholar] [CrossRef] [PubMed]
- Lin, G.; Chen, Q.; Yu, S.; Lin, S.; Yao, H.; Ding, Z.; Chen, S.; Lin, M.C.-M.; Wang, X. Overexpression of human telomerase reverse transcriptase C-terminal polypeptide sensitizes HeLa cells to 5-fluorouracil-induced growth inhibition and apoptosis. Mol. Med. Rep. 2014, 9, 279–284. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Liu, Y.; Li, H.; Zhang, R.; Dang, H.; Sun, P.; Zou, L.; Zhang, Y.; Gao, Y.; Hu, Y. Overexpression of the BRIP1 ameliorates chemosensitivity to cisplatin by inhibiting Rac1 GTPase activity in cervical carcinoma HeLa cells. Gene 2016, 578, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Song, K.-H.; Choi, C.H.; Lee, H.-J.; Oh, S.J.; Woo, S.R.; Hong, S.-O.; Noh, K.H.; Cho, H.; Chung, E.J.; Kim, J.-H.; et al. HDAC1 Upregulation by NANOG Promotes Multidrug Resistance and a Stem-like Phenotype in Immune Edited Tumor Cells. Cancer Res. 2017, 77, 5039–5053. [Google Scholar] [CrossRef]
- Chen, J.; Li, G. MiR-1284 enhances sensitivity of cervical cancer cells to cisplatin via downregulating HMGB1. Biomed. Pharmacother. 2018, 107, 997–1003. [Google Scholar] [CrossRef]
- The Journal of Gene Medicine. Gene Therapy Clinical Trials Worldwide. Available online: http://www.abedia.com/wiley/indications.php (accessed on 30 June 2019).
- Bethesda (MD): National Library of Medicine. ClinicalTrials.org. Available online: https://clinicaltrials.gov/ (accessed on 30 June 2019).
- Alvarez, R.D.; Huh, W.K.; Bae, S.; Lamb, L.S.; Conner, M.G.; Boyer, J.; Wang, C.; Hung, C.-F.; Sauter, E.; Paradis, M.; et al. A pilot study of pNGVL4a-CRT/E7(detox) for the treatment of patients with HPV16+ cervical intraepithelial neoplasia 2/3 (CIN2/3). Gynecol. Oncol. 2016, 140, 245–252. [Google Scholar] [CrossRef]
- Trimble, C.L.; Morrow, M.P.; Kraynyak, K.A.; Shen, X.; Dallas, M.; Yan, J.; Edwards, L.; Parker, R.L.; Denny, L.; Giffear, M.; et al. Safety, efficacy, and immunogenicity of VGX-3100, a therapeutic synthetic DNA vaccine targeting human papillomavirus 16 and 18 E6 and E7 proteins for cervical intraepithelial neoplasia 2/3: A randomised, double-blind, placebo-controlled phase 2b trial. Lancet (Lond. Engl.) 2015, 386, 2078–2088. [Google Scholar] [CrossRef]
- Rosales, R.; López-Contreras, M.; Rosales, C.; Magallanes-Molina, J.-R.; Gonzalez-Vergara, R.; Arroyo-Cazarez, J.M.; Ricardez-Arenas, A.; Del Follo-Valencia, A.; Padilla-Arriaga, S.; Guerrero, M.V.; et al. Regression of human papillomavirus intraepithelial lesions is induced by MVA E2 therapeutic vaccine. Hum. Gene Ther. 2014, 25, 1035–1049. [Google Scholar] [CrossRef]
- Basu, P.; Mehta, A.; Jain, M.; Gupta, S.; Nagarkar, R.V.; John, S.; Petit, R. A Randomized Phase 2 Study of ADXS11-001 Listeria monocytogenes-Listeriolysin O Immunotherapy With or Without Cisplatin in Treatment of Advanced Cervical Cancer. Int. J. Gynecol. Cancer 2018, 28, 764–772. [Google Scholar] [CrossRef]
- Xiao, J.; Zhou, J.; Fu, M.; Liang, L.; Deng, Q.; Liu, X.; Liu, F. Efficacy of recombinant human adenovirus-p53 combined with chemotherapy for locally advanced cervical cancer: A clinical trial. Oncol. Lett. 2017, 13, 3676–3680. [Google Scholar] [CrossRef]
- Huang, R.; Rofstad, E.K. Cancer stem cells (CSCs), cervical CSCs and targeted therapies. Oncotarget 2017, 8, 35351–35367. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Yue, Y.; Wang, R.; Gong, B.; Duan, Z. MicroRNA-145 inhibits tumorigenesis and invasion of cervical cancer stem cells. Int. J. Oncol. 2017, 50, 853–862. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Li, Y.; Liu, N.; Gao, Y.; Li, L. MiR-23b controls ALDH1A1 expression in cervical cancer stem cells. BMC Cancer 2017, 17, 292. [Google Scholar] [CrossRef] [PubMed]
- Bigoni-Ordóñez, G.D.; Ortiz-Sánchez, E.; Rosendo-Chalma, P.; Valencia-González, H.A.; Aceves, C.; García-Carrancá, A. Molecular iodine inhibits the expression of stemness markers on cancer stem-like cells of established cell lines derived from cervical cancer. BMC Cancer 2018, 18, 928. [Google Scholar] [CrossRef]
- Liu, J.; Cao, X.-C.; Xiao, Q.; Quan, M.-F. Apigenin inhibits HeLa sphere-forming cells through inactivation of casein kinase 2α. Mol. Med. Rep. 2015, 11, 665–669. [Google Scholar] [CrossRef]
- Wang, L.; Guo, H.; Yang, L.; Dong, L.; Lin, C.; Zhang, J.; Lin, P.; Wang, X. Morusin inhibits human cervical cancer stem cell growth and migration through attenuation of NF-κB activity and apoptosis induction. Mol. Cell. Biochem. 2013, 379, 7–18. [Google Scholar] [CrossRef]
- Wang, D.; Upadhyaya, B.; Liu, Y.; Knudsen, D.; Dey, M. Phenethyl isothiocyanate upregulates death receptors 4 and 5 and inhibits proliferation in human cancer stem-like cells. BMC Cancer 2014, 14, 591. [Google Scholar] [CrossRef]
- Yang, B.; Lu, Y.; Zhang, A.; Zhou, A.; Zhang, L.; Zhang, L.; Gao, L.; Zang, Y.; Tang, X.; Sun, L. Doxycycline Induces Apoptosis and Inhibits Proliferation and Invasion of Human Cervical Carcinoma Stem Cells. PLoS ONE 2015, 10, e0129138. [Google Scholar] [CrossRef]
- Wang, L.; Liu, Y.; Zhou, Y.; Wang, J.; Tu, L.; Sun, Z.; Wang, X.; Luo, F. Zoledronic acid inhibits the growth of cancer stem cell derived from cervical cancer cell by attenuating their stemness phenotype and inducing apoptosis and cell cycle arrest through the Erk1/2 and Akt pathways. J. Exp. Clin. Cancer Res. 2019, 38, 93. [Google Scholar] [CrossRef]
- Kwon, T.; Bak, Y.; Ham, S.-Y.; Yu, D.-Y.; Yoon, D.-Y. A1E reduces stemness and self-renewal in HPV 16-positive cervical cancer stem cells. BMC Complement. Altern. Med. 2016, 16, 42. [Google Scholar] [CrossRef]
- Stevanović, S.; Draper, L.M.; Langhan, M.M.; Campbell, T.E.; Kwong, M.L.; Wunderlich, J.R.; Dudley, M.E.; Yang, J.C.; Sherry, R.M.; Kammula, U.S.; et al. Complete regression of metastatic cervical cancer after treatment with human papillomavirus-targeted tumor-infiltrating T cells. J. Clin. Oncol. 2015, 33, 1543–1550. [Google Scholar] [CrossRef] [PubMed]
- Newick, K.; O’Brien, S.; Moon, E.; Albelda, S.M. CAR T Cell Therapy for Solid Tumors. Annu. Rev. Med. 2017, 68, 139–152. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strategy | Gene | Function in Cervical Cancer | Developed/Inhibition Strategy | Model | Reference |
|---|---|---|---|---|---|
| Tumor suppressor gene restoration | p53 | Cell cycle arrest, apoptosis, autophagy, inhibition of proliferation of tumor cells and chemo/radiosensitivity | AP-PAMAM rAd-p53 | In vitro In vitro, in vivo, in human | [37,38,39] |
| RIZ1 | Cell cycle arrest and apoptosis | pcDNA3.1(+)-RIZ1 plasmid | In vitro, in vivo | [40,41] | |
| PEDF | Antiangiogenic and antitumorigenic properties | FLP, ip administration | In vitro, in vivo | [26] | |
| PTPRJ | Regulate cell growth, differentiation and cell cycle | Lentivirus-pSicoR-PTPRJ | In vitro | [42] | |
| Blocking oncogenic expression | E6/E7 | Polyubiquination of p53, suppressing its function/pRb degradation, leading to S-phase entry, viral replication and maintenance | rAd-artificial miRNAs, intratumoral injection | In vitro, in vivo | [43,44] |
| gene silencing activated under illumination | In vitro | [45] | |||
| CRISPR-Cas9 TALEN | In vitro, in vivo, in human | [46,47] | |||
| siRNA targeting E6/E7 promoter | In vitro, in vivo | [48] | |||
| siRNA delivery by PEG-lipoplexes | In vitro, in vivo | [49] | |||
| Ad-ER-DN | In vitro | [50,51] | |||
| XIAP | Anti-apoptosis | siRNA | In vitro, in vivo | [52] | |
| MMP | Degrade extracellular matrix components, important in cell motility | shRNA Knockdown of PTX3 | In vitro, in vivo | [53,54] | |
| ASRGL1 | Cell cycle and anti-apoptotic factor regulation | shRNA-expressing lentivirus | In vitro | [55] | |
| hTERT | Lengthens telomeres in DNA strands, conferring immortality | siRNA Knockdown of HMBOX1 | In vitro, in vivo | [56,57,58] | |
| C-MYC | Transcriptional factor involved in cell proliferation and tumorigenesis | Sendai virus carrying FIR | In vitro, in vivo | [59] |
| Therapeutic Strategy | Intervention | Reference | Phase | Year (First–Last Posted) |
|---|---|---|---|---|
| Genetically engineered T-Cells and chemotherapy | Fludarabine and Cyclophosphamide + E6 TCR (T-Cells genetically engineered to express T-Cell Receptors targeting HPV-16 E6) + Aldesleukin | NCT02280811 | Phase 1 Phase 2 | 2014–2017 |
| Fludarabine and Cyclophosphamide + Anti-MAGE-A3-DP4 TCR (T-Cells genetically engineered to express T-Cell Receptors targeting the DP0401/0402 restricted MAGE-A3 tumor antigen) + Aldesleukin | NCT02111850 | Phase 2 | 2014–2018 | |
| Aldesleukin + Fludarabine and Cyclophosphamide + Anti-MAGE-A3 HLAA* 01-restricted TCR (T-Cells genetically engineered to express T-Cell Receptors targeting MAGE-A3 tumor antigen) | NCT02153905 | Phase 1 Phase 2 | 2014–2018 | |
| HPV Specific T Cells (modified genetically to be resistant to the TGF-beta) ± lymphodepletion (Cyclophosphamide and Fludarabine) and nivolumab | NCT02379520 | Phase 1 | 2015–2018 | |
| CAR transduced PBL and chemotherapy | Fludarabine + Anti-mesothelin CAR transduced PBL (retroviral vector that contains a chimeric T cell receptor targeting mesothelin) + Cyclophosphamide + Aldesleukin | NCT01583686 | Phase 1 Phase 2 | 2012–2018 |
| Vaccine | pNGVL4a-CRT/E7(detox) vaccine, which targets HPV16 E7 | NCT00988559 | Not Applicable | 2009–2016 |
| Vaccine and chemotherapy | TA-HPV (HPVE6/E7 recombinant vaccine) and pNGVL4a-Sig/E7(detox)/HSP70 DNA vaccines ± imiquimod | NCT00788164 | Phase 1 | 2008–2018 |
| Vigil (vaccine composed of autologous tumor cells which are transfected extracorporeally with a plasmid encoding for the gene for GM-CSF and a bifunctional shRNA that targets furin, a convertase responsible for activation of both TGβ1 and β2) + Atezolizumab | NCT03073525 | Phase 2 | 2017–2018 | |
| DNA plasmid-encoding IL-12/HPV DNA plasmid therapeutic vaccine INO-3112 (MEDI0457) in combination with durvalumab | NCT03439085 | Phase 2 | 2018–2018 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Áyen, Á.; Jiménez Martínez, Y.; Boulaiz, H. Targeted Gene Delivery Therapies for Cervical Cancer. Cancers 2020, 12, 1301. https://doi.org/10.3390/cancers12051301
Áyen Á, Jiménez Martínez Y, Boulaiz H. Targeted Gene Delivery Therapies for Cervical Cancer. Cancers. 2020; 12(5):1301. https://doi.org/10.3390/cancers12051301
Chicago/Turabian StyleÁyen, Ángela, Yaiza Jiménez Martínez, and Houria Boulaiz. 2020. "Targeted Gene Delivery Therapies for Cervical Cancer" Cancers 12, no. 5: 1301. https://doi.org/10.3390/cancers12051301
APA StyleÁyen, Á., Jiménez Martínez, Y., & Boulaiz, H. (2020). Targeted Gene Delivery Therapies for Cervical Cancer. Cancers, 12(5), 1301. https://doi.org/10.3390/cancers12051301