Rho GTPases in Gynecologic Cancers: In-Depth Analysis toward the Paradigm Change from Reactive to Predictive, Preventive, and Personalized Medical Approach Benefiting the Patient and Healthcare

 , ,
, ,  ,
,
Abstract
1. Introduction
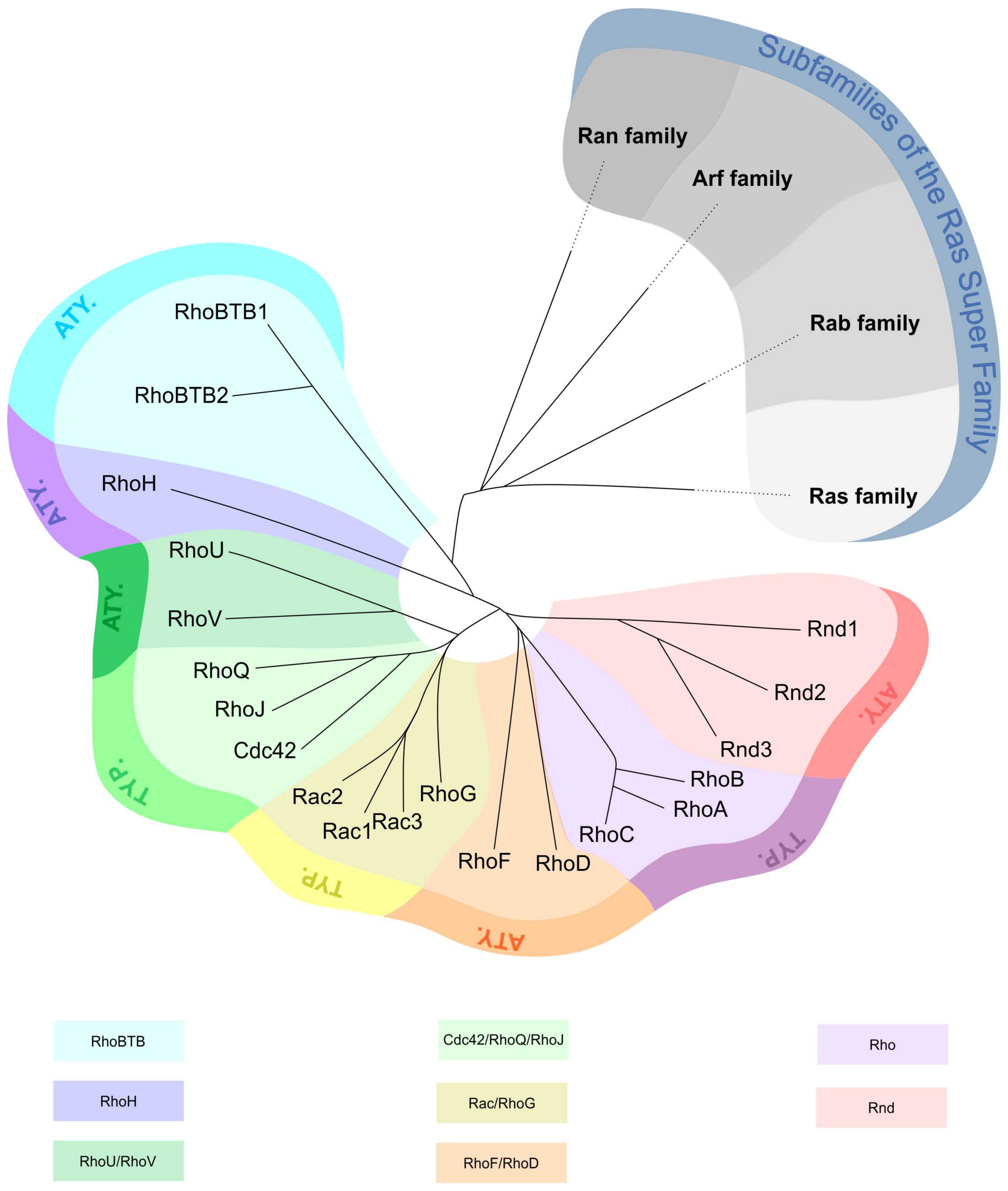
1.1. Rho GTPase Characteristics and Classification
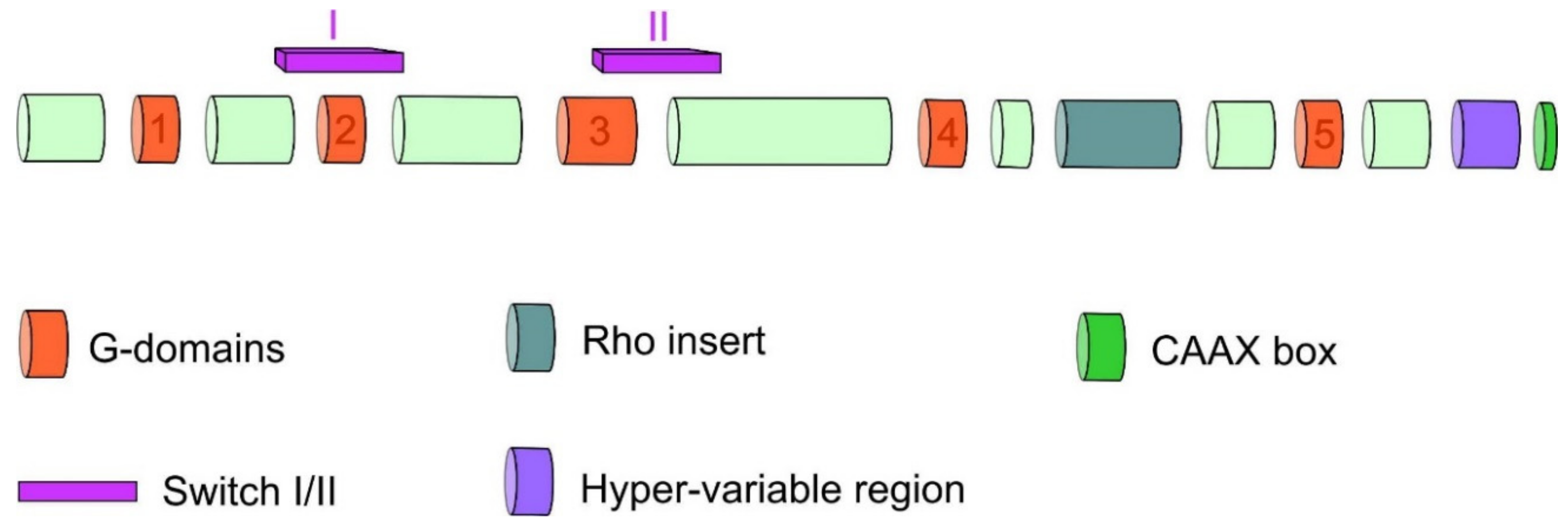
1.2. Rho GTPases Structure
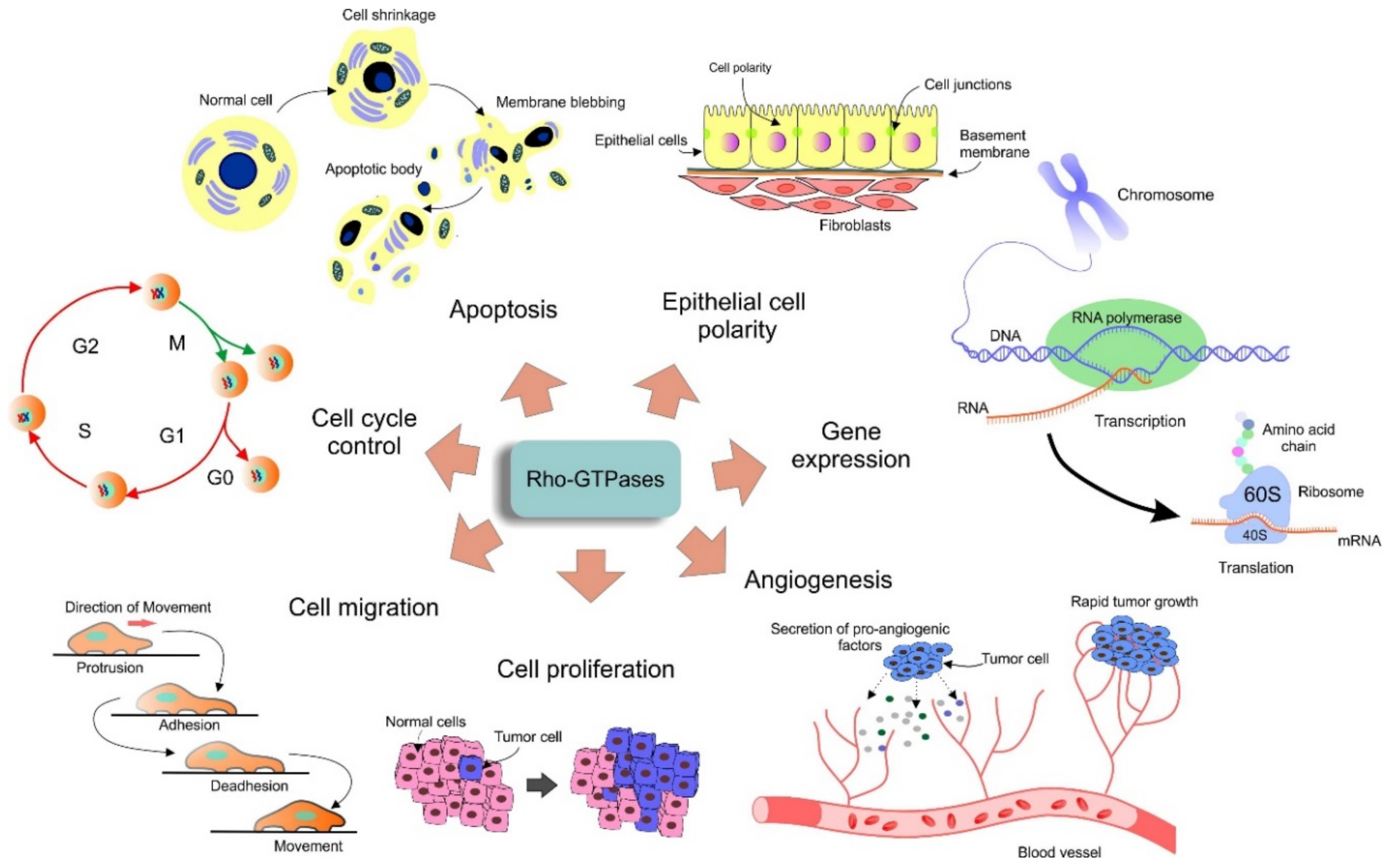
1.3. Rho GTPases Function
1.4. Rho GTPases Regulation
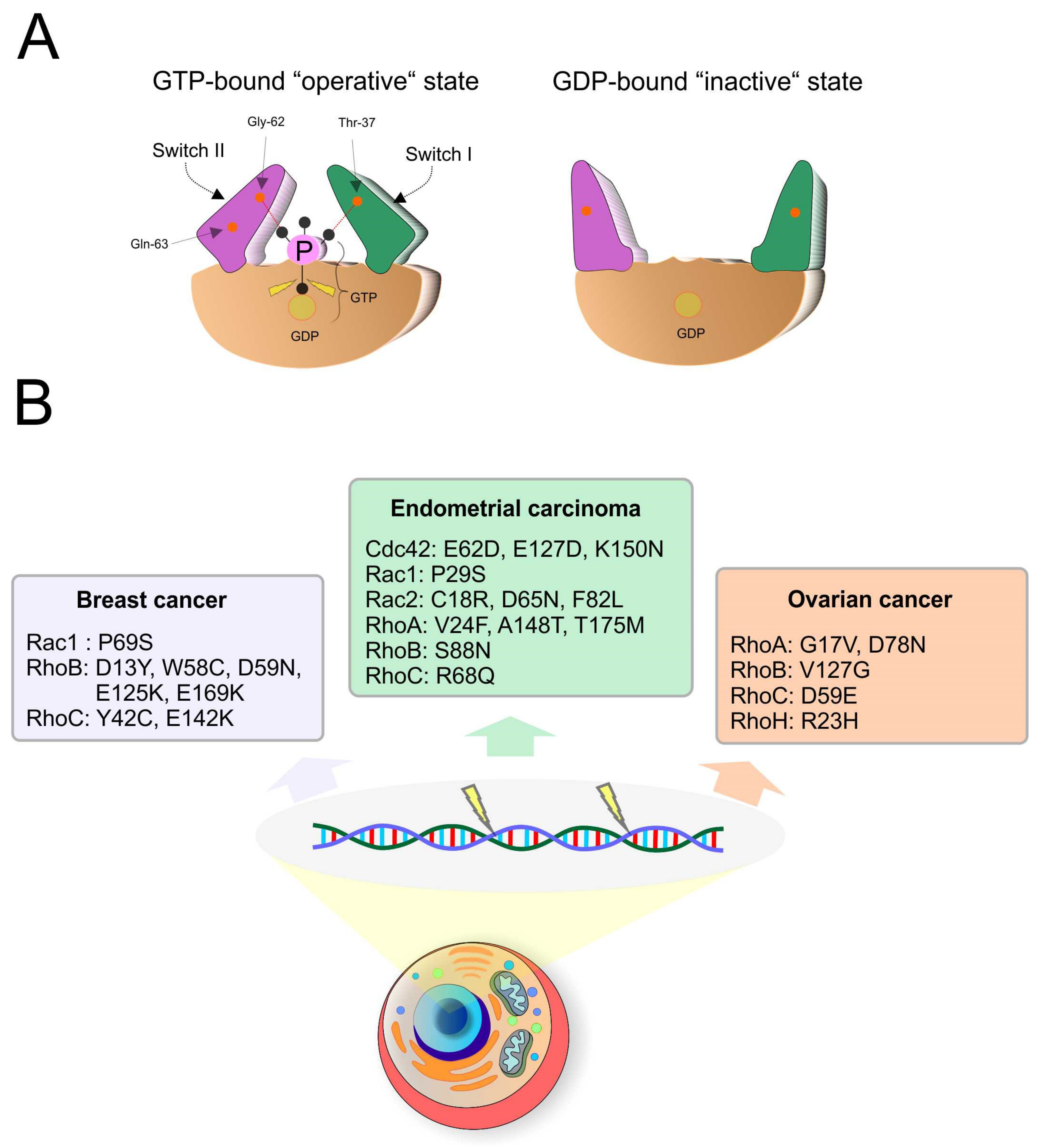
1.5. The Role of GTPases in Tumorigenesis
2. Rho GTPases in Gynecological Cancer
2.1. Ovarian Cancer
2.2. Endometrial Cancer
2.3. Cervical Cancer
2.4. Breast Cancer
3. A Role of Rho GTPases in Cancer Therapy
3.1. ROCK Inhibitors
3.1.1. Y-27632
3.1.2. Fasudil
3.1.3. Phytochemicals
3.2. RhoA, Rac1, and Cdc42 Inhibitors
3.2.1. Ketorolac
3.2.2. NSC23766
3.2.3. Berberine
3.3. Farnesyltransferase Inhibitors and Geranylgeranyltransferase Inhibitor
3.4. Rho Regulators Inhibitors
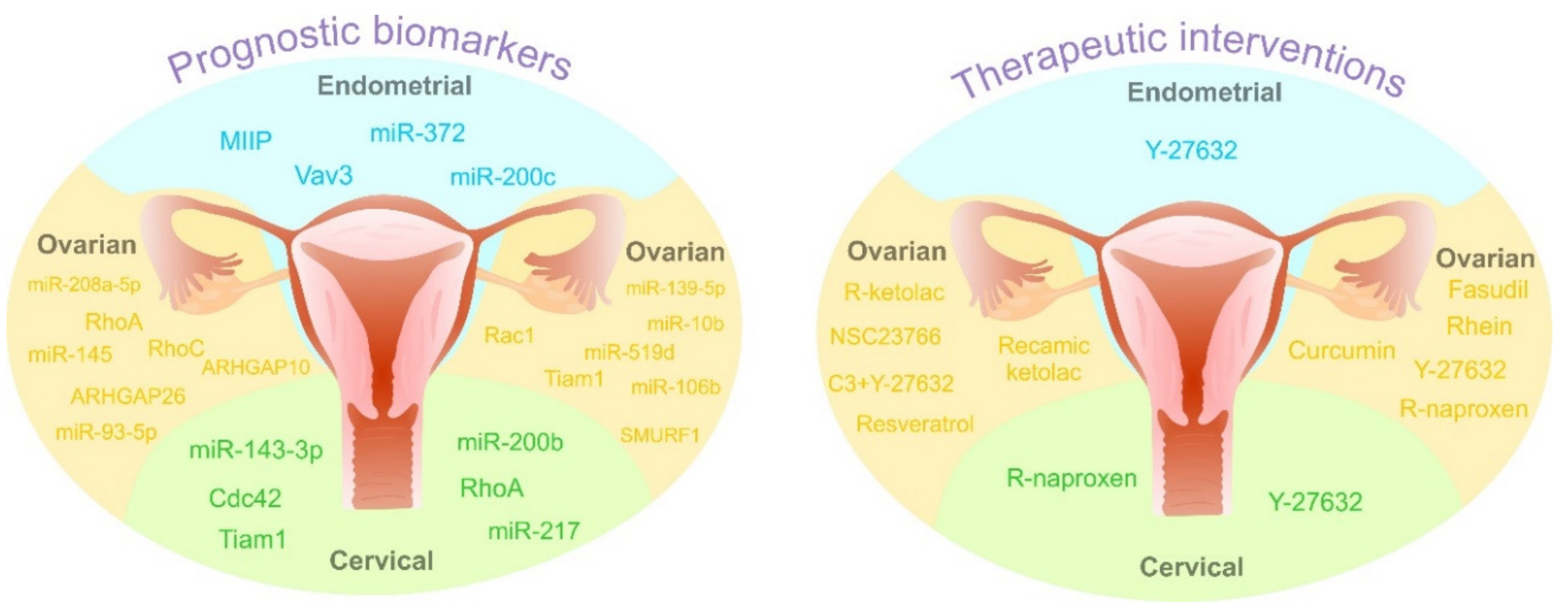
4. Prognostic Value of Rho GTPases: Spectacular Biomarkers or “Blind Alley” of Personalized Medicine
5. Towards Predictive, Preventive, and Personalized Medical (PPPM/3PM) Approaches in Gynecological Oncology: Prominent Examples Involving Rho GTPases as a Target
5.1. Rho GTPases in Prediction of Aggressive Gynecologic Cancers and Metastatic Disease: Prominent Examples
5.1.1. Rho GTPases as a Target for Early Detection of Pre/Cancerous Lesions in High Mammographic Density Breast
5.1.2. Predictive Diagnosis of Breast Cancer Based on RhoA Patterns: Multiomics Approach
5.2. Targeted Prevention
5.2.1. Inflammation
5.2.2. Chronic Wounds and Impaired Healing
5.2.3. Rho GTPases-Based Prevention of Endometrial Carcinoma in Obese Phenotype
5.2.4. Small GTPases are Regulated by Nutrients—An Approach for Dietary Cancer Prevention
5.2.5. Targeted Chemoprevention
5.3. Personalized Treatment Algorithms
- -
- Personalized preventive measures applied to suboptimal health;
- -
- Personalized primary cancer prevention and treatments at the level of pre-cancerous lesions;
- -
- Personalized treatment of cancer;
- -
- Personalized secondary prevention (e.g., prevention of cancer in obese and diabetic patients);
- -
6. Conclusions
Funding
Conflicts of Interest
References
- Madaule, P.; Axel, R. A novel ras-related gene family. Cell 1985, 41, 31–40. [Google Scholar] [CrossRef]
- Haga, R.B.; Ridley, A.J. Rho GTPases: Regulation and roles in cancer cell biology. Small GTPases 2016, 7, 207–221. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.S.; Chen, Y.H. Ras family of small GTPases in immunity and inflammation. Curr. Opin. Pharmacol. 2012, 12, 458–463. [Google Scholar] [CrossRef] [PubMed]
- Boureux, A.; Vignal, E.; Faure, S.; Fort, P. Evolution of the Rho family of ras-like GTPases in eukaryotes. Mol. Biol. Evol. 2007, 24, 203–216. [Google Scholar] [CrossRef] [PubMed]
- Hall, A. Rho family GTPases. Biochem. Soc. Trans. 2012, 40, 1378–1382. [Google Scholar] [CrossRef] [PubMed]
- Heasman, S.J.; Ridley, A.J. Mammalian Rho GTPases: new insights into their functions from in vivo studies. Nat. Rev. Mol. Cell Biol. 2008, 9, 690–701. [Google Scholar] [CrossRef]
- Wennerberg, K.; Der, C.J. Rho-family GTPases: it’s not only Rac and Rho (and I like it). J. Cell Sci. 2004, 117, 1301–1312. [Google Scholar] [CrossRef]
- Etienne-Manneville, S.; Hall, A. Rho GTPases in cell biology. Nature 2002, 420, 629–635. [Google Scholar] [CrossRef]
- Schwartz, M. Rho signalling at a glance. J. Cell Sci. 2004, 117, 5457–5458. [Google Scholar] [CrossRef]
- Vega, F.M.; Ridley, A.J. Rho GTPases in cancer cell biology. FEBS Lett. 2008, 582, 2093–2101. [Google Scholar] [CrossRef]
- Wittinghofer, A.; Vetter, I.R. Structure-Function Relationships of the G Domain, a Canonical Switch Motif. Annu. Rev. Biochem. 2011, 80, 943–971. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, A.; Reinhard, N.R.; Hordijk, P.L. Toward understanding RhoGTPase specificity: structure, function and local activation. Small GTPases 2014, 5, e968004. [Google Scholar] [CrossRef]
- Thomas, C.; Fricke, I.; Scrima, A.; Berken, A.; Wittinghofer, A. Structural evidence for a common intermediate in small G protein-GEF reactions. Mol. Cell 2007, 25, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Bourne, H.R.; Sanders, D.A.; McCormick, F. The GTPase superfamily: a conserved switch for diverse cell functions. Nature 1990, 348, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Bourne, H.R.; Sanders, D.A.; McCormick, F. The GTPase superfamily: conserved structure and molecular mechanism. Nature 1991, 349, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Wennerberg, K.; Rossman, K.L.; Der, C.J. The Ras superfamily at a glance. J. Cell Sci. 2005, 118, 843–846. [Google Scholar] [CrossRef]
- ten Klooster, J.P.; Hordijk, P.L. Targeting and localized signalling by small GTPases. Biol. Cell 2007, 99, 1–12. [Google Scholar] [CrossRef]
- Cherfils, J.; Zeghouf, M. Regulation of Small GTPases by GEFs, GAPs, and GDIs. Physiol. Rev. 2013, 93, 269–309. [Google Scholar] [CrossRef]
- Vetter, I.R. The Guanine Nucleotide-Binding Switch in Three Dimensions. Science 2001, 294, 1299–1304. [Google Scholar] [CrossRef]
- Porter, A.P.; Papaioannou, A.; Malliri, A. Deregulation of Rho GTPases in cancer. Small GTPases 2016, 7, 123–138. [Google Scholar] [CrossRef]
- Berthold, J.; Schenková, K.; Ramos, S.; Miura, Y.; Furukawa, M.; Aspenström, P.; Rivero, F. Characterization of RhoBTB-dependent Cul3 ubiquitin ligase complexes--evidence for an autoregulatory mechanism. Exp. Cell Res. 2008, 314, 3453–3465. [Google Scholar] [CrossRef] [PubMed]
- Ji, W.; Rivero, F. Atypical Rho GTPases of the RhoBTB Subfamily: Roles in Vesicle Trafficking and Tumorigenesis. Cells 2016, 5, 28. [Google Scholar] [CrossRef] [PubMed]
- Aspenström, P.; Fransson, A.; Saras, J. Rho GTPases have diverse effects on the organization of the actin filament system. Biochem. J. 2004, 377, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, A.; Ping, Q.; Carpenter, C.L. RhoBTB2 is a substrate of the mammalian Cul3 ubiquitin ligase complex. Genes Dev. 2004, 18, 856–861. [Google Scholar] [CrossRef]
- Hudson, L.G.; Gillette, J.M.; Kang, H.; Rivera, M.R.; Wandinger-Ness, A. Ovarian Tumor Microenvironment Signaling: Convergence on the Rac1 GTPase. Cancers 2018, 10, 358. [Google Scholar] [CrossRef]
- Ridley, A.J. Rho GTPase signalling in cell migration. Curr. Opin. Cell Biol. 2015, 36, 103–112. [Google Scholar] [CrossRef]
- Rossman, K.L.; Der, C.J.; Sondek, J. GEF means go: turning on RHO GTPases with guanine nucleotide-exchange factors. Nat. Rev. Mol. Cell Biol. 2005, 6, 167–180. [Google Scholar] [CrossRef]
- Jaffe, A.B.; Hall, A. Rho GTPases: biochemistry and biology. Annu. Rev. Cell Dev. Biol. 2005, 21, 247–269. [Google Scholar] [CrossRef]
- Cardama, G.A.; Gonzalez, N.; Maggio, J.; Menna, P.L.; Gomez, D.E. Rho GTPases as therapeutic targets in cancer (Review). Int. J. Oncol. 2017, 51, 1025–1034. [Google Scholar] [CrossRef]
- Bishop, A.L.; Hall, A. Rho GTPases and their effector proteins. Biochem. J. 2000, 348(Pt. 2), 241–255. [Google Scholar] [CrossRef]
- Miyano, K.; Sumimoto, H. Assessment of the role for Rho family GTPases in NADPH oxidase activation. Methods Mol. Biol. 2012, 827, 195–212. [Google Scholar] [CrossRef] [PubMed]
- David, M.; Petit, D.; Bertoglio, J. Cell cycle regulation of Rho signaling pathways. Cell Cycle 2012, 11, 3003–3010. [Google Scholar] [CrossRef] [PubMed]
- Hodge, R.G.; Ridley, A.J. Regulating Rho GTPases and their regulators. Nat. Rev. Mol. Cell Biol. 2016, 17, 496–510. [Google Scholar] [CrossRef] [PubMed]
- Meller, N.; Merlot, S.; Guda, C. CZH proteins: a new family of Rho-GEFs. J. Cell Sci. 2005, 118, 4937–4946. [Google Scholar] [CrossRef] [PubMed]
- Fort, P.; Blangy, A. The Evolutionary Landscape of Dbl-Like RhoGEF Families: Adapting Eukaryotic Cells to Environmental Signals. Genome Biol. Evol. 2017, 9, 1471–1486. [Google Scholar] [CrossRef] [PubMed]
- Cook, D.R.; Rossman, K.L.; Der, C.J. Rho guanine nucleotide exchange factors: regulators of Rho GTPase activity in development and disease. Oncogene 2014, 33, 4021–4035. [Google Scholar] [CrossRef]
- Laurin, M.; Cote, J.-F. Insights into the biological functions of Dock family guanine nucleotide exchange factors. Genes Dev. 2014, 28, 533–547. [Google Scholar] [CrossRef]
- Amin, E.; Jaiswal, M.; Derewenda, U.; Reis, K.; Nouri, K.; Koessmeier, K.T.; Aspenström, P.; Somlyo, A.V.; Dvorsky, R.; Ahmadian, M.R. Deciphering the Molecular and Functional Basis of RHOGAP Family Proteins: : A systematic approach toward selective inactivation of rho family proteins. J. Biol. Chem. 2016, 291, 20353–20371. [Google Scholar] [CrossRef]
- Garcia-Mata, R.; Boulter, E.; Burridge, K. The “invisible hand”: regulation of RHO GTPases by RHOGDIs. Nat. Rev. Mol. Cell Biol. 2011, 12, 493–504. [Google Scholar] [CrossRef]
- DerMardirossian, C.; Bokoch, G.M. GDIs: central regulatory molecules in Rho GTPase activation. Trends Cell Biol. 2005, 15, 356–363. [Google Scholar] [CrossRef]
- Liu, M.; Bi, F.; Zhou, X.; Zheng, Y. Rho GTPase regulation by miRNAs and covalent modifications. Trends Cell Biol. 2012, 22, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Croft, D.R.; Olson, M.F. Transcriptional regulation of Rho GTPase signaling. Transcription 2011, 2, 211–215. [Google Scholar] [CrossRef] [PubMed]
- Aspenström, P.; Ruusala, A.; Pacholsky, D. Taking Rho GTPases to the next level: the cellular functions of atypical Rho GTPases. Exp. Cell Res. 2007, 313, 3673–3679. [Google Scholar] [CrossRef] [PubMed]
- Aspenström, P. Fast-cycling Rho GTPases. Small GTPases 2018, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Hodge, R.G.; Ridley, A.J. Regulation and functions of RhoU and RhoV. Small GTPases 2020, 11, 8–15. [Google Scholar] [CrossRef]
- Traut, T.W. Physiological concentrations of purines and pyrimidines. Mol. Cell. Biochem. 1994, 140, 1–22. [Google Scholar] [CrossRef]
- Bustelo, X.R. RHO GTPases in cancer: known facts, open questions, and therapeutic challenges. Biochem. Soc. Trans. 2018, 46, 741–760. [Google Scholar] [CrossRef]
- Huang, M.; Prendergast, G.C. RhoB in cancer suppression. Histol. Histopathol. 2006, 21, 213–218. [Google Scholar] [CrossRef]
- Li, N.; Zhan, X. Signaling pathway network alterations in human ovarian cancers identified with quantitative mitochondrial proteomics. EPMA J. 2019, 10, 153–172. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2018. CA Cancer J. Clin. 2018, 68, 7–30. [Google Scholar] [CrossRef]
- Ray-Coquard, I.; Trama, A.; Seckl, M.J.; Fotopoulou, C.; Pautier, P.; Pignata, S.; Kristensen, G.; Mangili, G.; Falconer, H.; Massuger, L.; et al. Rare ovarian tumours: Epidemiology, treatment challenges in and outside a network setting. Eur. J. Surg. Oncol. 2019, 45, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Coleman, R.L.; Monk, B.J.; Sood, A.K.; Herzog, T.J. Latest research and treatment of advanced-stage epithelial ovarian cancer. Nat. Rev. Clin. Oncol. 2013, 10, 211–224. [Google Scholar] [CrossRef] [PubMed]
- Paulsen, T.; Kjaerheim, K.; Kaern, J.; Tretli, S.; Tropé, C. Improved short-term survival for advanced ovarian, tubal, and peritoneal cancer patients operated at teaching hospitals. Int. J. Gynecol. Cancer 2006, 16 (Suppl. 1), 11–17. [Google Scholar] [CrossRef]
- Noer, M.C.; Leandersson, P.; Paulsen, T.; Rosthøj, S.; Antonsen, S.L.; Borgfeldt, C.; Høgdall, C. Confounders other than comorbidity explain survival differences in Danish and Swedish ovarian cancer patients - a comparative cohort study. Acta Oncol. 2018, 57, 1100–1108. [Google Scholar] [CrossRef]
- Paulsen, T.; Kaern, J.; Kjaerheim, K.; Haldorsen, T.; Tropé, C. Influence of interval between primary surgery and chemotherapy on short-term survival of patients with advanced ovarian, tubal or peritoneal cancer. Gynecol. Oncol. 2006, 102, 447–452. [Google Scholar] [CrossRef]
- Paulsen, T.; Kærn, J.; Tropé, C. Improved 5-year disease-free survival for FIGO stage I epithelial ovarian cancer patients without tumor rupture during surgery. Gynecol. Oncol. 2011, 122, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Vilming Elgaaen, B.; Olstad, O.K.; Haug, K.B.F.; Brusletto, B.; Sandvik, L.; Staff, A.C.; Gautvik, K.M.; Davidson, B. Global miRNA expression analysis of serous and clear cell ovarian carcinomas identifies differentially expressed miRNAs including miR-200c-3p as a prognostic marker. BMC Cancer 2014, 14, 80. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Wang, J.; Gou, W.-F.; Xiu, Y.-L.; Zheng, H.-C.; Zong, Z.-H.; Takano, Y.; Zhao, Y. The Involvement of RhoA and Wnt-5a in the Tumorigenesis and Progression of Ovarian Epithelial Carcinoma. Int. J. Mol. Sci. 2013, 14, 24187–24199. [Google Scholar] [CrossRef]
- Ridley, A.J. Rho GTPases and cell migration. J. Cell Sci. 2001, 114, 2713–2722. [Google Scholar]
- Zaoui, K.; Boudhraa, Z.; Khalifé, P.; Carmona, E.; Provencher, D.; Mes-Masson, A.-M. Ran promotes membrane targeting and stabilization of RhoA to orchestrate ovarian cancer cell invasion. Nat. Commun. 2019, 10, 1–12. [Google Scholar] [CrossRef]
- Horiuchi, A.; Imai, T.; Wang, C.; Ohira, S.; Feng, Y.; Nikaido, T.; Konishi, I. Up-Regulation of Small GTPases, RhoA and RhoC, Is Associated with Tumor Progression in Ovarian Carcinoma. Lab. Investig. 2003, 83, 861–870. [Google Scholar] [CrossRef] [PubMed]
- Adamson, P.; Paterson, H.F.; Hall, A. Intracellular localization of the P21rho proteins. J. Cell Biol. 1992, 119, 617–627. [Google Scholar] [CrossRef] [PubMed]
- Cavazza, T.; Vernos, I. The RanGTP Pathway: From Nucleo-Cytoplasmic Transport to Spindle Assembly and Beyond. Front. Cell. Dev. Biol. 2015, 3, 82. [Google Scholar] [CrossRef] [PubMed]
- Ouellet, V.; Guyot, M.-C.; Le Page, C.; Filali-Mouhim, A.; Lussier, C.; Tonin, P.N.; Provencher, D.M.; Mes-Masson, A.-M. Tissue array analysis of expression microarray candidates identifies markers associated with tumor grade and outcome in serous epithelial ovarian cancer. Int. J. Cancer 2006, 119, 599–607. [Google Scholar] [CrossRef]
- Cheung, L.W.T.; Leung, P.C.K.; Wong, A.S.T. Cadherin switching and activation of p120 catenin signaling are mediators of gonadotropin-releasing hormone to promote tumor cell migration and invasion in ovarian cancer. Oncogene 2010, 29, 2427–2440. [Google Scholar] [CrossRef]
- Reynolds, A.B.; Roesel, D.J.; Kanner, S.B.; Parsons, J.T. Transformation-specific tyrosine phosphorylation of a novel cellular protein in chicken cells expressing oncogenic variants of the avian cellular src gene. Mol. Cell. Biol. 1989, 9, 629–638. [Google Scholar] [CrossRef]
- Chen, X.; Li, X.; Wang, X.; Zhu, Q.; Wu, X.; Wang, X. MUC16 impacts tumor proliferation and migration through cytoplasmic translocation of P120-catenin in epithelial ovarian cancer cells: an original research. BMC Cancer 2019, 19, 171. [Google Scholar] [CrossRef]
- Thériault, C.; Pinard, M.; Comamala, M.; Migneault, M.; Beaudin, J.; Matte, I.; Boivin, M.; Piché, A.; Rancourt, C. MUC16 (CA125) regulates epithelial ovarian cancer cell growth, tumorigenesis and metastasis. Gynecol. Oncol. 2011, 121, 434–443. [Google Scholar] [CrossRef]
- Earp, M.; Tyrer, J.P.; Winham, S.J.; Lin, H.-Y.; Chornokur, G.; Dennis, J.; Aben, K.K.H.; Anton-Culver, H.; Antonenkova, N.; Bandera, E.V.; et al. Variants in genes encoding small GTPases and association with epithelial ovarian cancer susceptibility. PLoS ONE 2018, 13, e0197561. [Google Scholar] [CrossRef]
- Guo, Y.; Kenney, S.R.; Cook, L.; Adams, S.F.; Rutledge, T.; Romero, E.; Oprea, T.I.; Sklar, L.A.; Bedrick, E.; Wiggins, C.L.; et al. A Novel Pharmacologic Activity of Ketorolac for Therapeutic Benefit in Ovarian Cancer Patients. Clin. Cancer Res. 2015, 21, 5064–5072. [Google Scholar] [CrossRef]
- Guo, Y.; Kenney, S.R.; Muller, C.Y.; Adams, S.; Rutledge, T.; Romero, E.; Murray-Krezan, C.; Prekeris, R.; Sklar, L.A.; Hudson, L.G.; et al. R-Ketorolac Targets Cdc42 and Rac1 and Alters Ovarian Cancer Cell Behaviors Critical for Invasion and Metastasis. Mol. Cancer Ther. 2015, 14, 2215–2227. [Google Scholar] [CrossRef] [PubMed]
- Winkler, S.; Mohl, M.; Wieland, T.; Lutz, S. GrinchGEF--a novel Rho-specific guanine nucleotide exchange factor. Biochem. Biophys. Res. Commun. 2005, 335, 1280–1286. [Google Scholar] [CrossRef] [PubMed]
- Mokady, D.; Meiri, D. RhoGTPases - A novel link between cytoskeleton organization and cisplatin resistance. Drug Resist. Update 2015, 19, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Lazer, G.; Katzav, S. Guanine nucleotide exchange factors for RhoGTPases: Good therapeutic targets for cancer therapy? Cell. Signal. 2011, 23, 969–979. [Google Scholar] [CrossRef] [PubMed]
- Sahai, E.; Marshall, C.J. RHO–GTPases and cancer. Nat. Rev. Cancer 2002, 2, 133–142. [Google Scholar] [CrossRef]
- Sharma, S.; Santiskulvong, C.; Bentolila, L.A.; Rao, J.; Dorigo, O.; Gimzewski, J.K. Correlative nanomechanical profiling with super-resolution F-actin imaging reveals novel insights into mechanisms of cisplatin resistance in ovarian cancer cells. Nanomedicine 2012, 8, 757–766. [Google Scholar] [CrossRef]
- McGrail, D.J.; Kieu, Q.M.N.; Dawson, M.R. The malignancy of metastatic ovarian cancer cells is increased on soft matrices through a mechanosensitive Rho-ROCK pathway. J. Cell Sci. 2014, 127, 2621–2626. [Google Scholar] [CrossRef]
- Torre, L.A.; Islami, F.; Siegel, R.L.; Ward, E.M.; Jemal, A. Global Cancer in Women: Burden and Trends. Cancer Epidemiol. Biomark. Prev. 2017, 26, 444–457. [Google Scholar] [CrossRef]
- Lindemann, K.; Eskild, A.; Vatten, L.J.; Bray, F. Endometrial cancer incidence trends in Norway during 1953-2007 and predictions for 2008-2027. Int. J. Cancer 2010, 127, 2661–2668. [Google Scholar] [CrossRef]
- Felix, A.S.; Brinton, L.A. Cancer progress and priorities: Uterine cancer. Cancer Epidemiol. Biomark. Prev. 2018, 27, 985–994. [Google Scholar] [CrossRef]
- Charo, L.M.; Plaxe, S.C. Recent advances in endometrial cancer: a review of key clinical trials from 2015 to 2019. F1000Research 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Suri, V.; Arora, A. Management of Endometrial Cancer: A Review. Rev. Recent Clin. Trials 2015, 10, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Brooks, R.A.; Fleming, G.F.; Lastra, R.R.; Lee, N.K.; Moroney, J.W.; Son, C.H.; Tatebe, K.; Veneris, J.L. Current recommendations and recent progress in endometrial cancer. CA Cancer J. Clin. 2019. [Google Scholar] [CrossRef] [PubMed]
- Lindemann, K.; Vatten, L.J.; Ellstrøm-Engh, M.; Eskild, A. Body mass, diabetes and smoking, and endometrial cancer risk: a follow-up study. Br. J. Cancer 2008, 98, 1582–1585. [Google Scholar] [CrossRef]
- Lindemann, K.; Vatten, L.J.; Ellstrøm-Engh, M.; Eskild, A. Serum lipids and endometrial cancer risk: results from the HUNT-II study. Int. J. Cancer 2009, 124, 2938–2941. [Google Scholar] [CrossRef]
- Sherman, M.E.; Devesa, S.S. Analysis of racial differences in incidence, survival, and mortality for malignant tumors of the uterine corpus. Cancer 2003, 98, 176–186. [Google Scholar] [CrossRef]
- Smith, D.; Stewart, C.J.R.; Clarke, E.M.; Lose, F.; Davies, C.; Armes, J.; Obermair, A.; Brennan, D.; Webb, P.M.; Nagle, C.M.; et al. ER and PR expression and survival after endometrial cancer. Gynecol. Oncol. 2018, 148, 258–266. [Google Scholar] [CrossRef]
- Lobo, F.; Thomas, E. Type II endometrial cancers: A case series. J. Midlife Health 2016, 7, 69–72. [Google Scholar] [CrossRef]
- Zahl Eriksson, A.G.; Ducie, J.; Ali, N.; McGree, M.E.; Weaver, A.L.; Bogani, G.; Cliby, W.A.; Dowdy, S.C.; Bakkum-Gamez, J.N.; Abu-Rustum, N.R.; et al. Comparison of a sentinel lymph node and a selective lymphadenectomy algorithm in patients with endometrioid endometrial carcinoma and limited myometrial invasion. Gynecol. Oncol. 2016, 140, 394–399. [Google Scholar] [CrossRef]
- Leitao, M.M.; Narain, W.R.; Boccamazzo, D.; Sioulas, V.; Cassella, D.; Ducie, J.A.; Eriksson, A.G.Z.; Sonoda, Y.; Chi, D.S.; Brown, C.L.; et al. Impact of Robotic Platforms on Surgical Approach and Costs in the Management of Morbidly Obese Patients with Newly Diagnosed Uterine Cancer. Ann. Surg. Oncol. 2016, 23, 2192–2198. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Y.; Li, J.; Cragun, J.; Hatch, K.; Chambers, S.K.; Zheng, W. Lynch syndrome related endometrial cancer: clinical significance beyond the endometrium. J. Hematol. Oncol. 2013, 6, 22. [Google Scholar] [CrossRef] [PubMed]
- Gammon, A.; Jasperson, K.; Champine, M. Genetic basis of Cowden syndrome and its implications for clinical practice and risk management. Appl. Clin. Genet. 2016, 9, 83–92. [Google Scholar] [CrossRef] [PubMed]
- O’Hara, A.J.; Bell, D.W. The genomics and genetics of endometrial cancer. Adv. Genom. Genet. 2012, 2012, 33–47. [Google Scholar] [CrossRef]
- Wang, Y.; Hu, L.; Ji, P.; Teng, F.; Tian, W.; Liu, Y.; Cogdell, D.; Liu, J.; Sood, A.K.; Broaddus, R.; et al. MIIP remodels Rac1-mediated cytoskeleton structure in suppression of endometrial cancer metastasis. J. Hematol. Oncol. 2016, 9. [Google Scholar] [CrossRef]
- Ridley, A.J. Rho GTPases and actin dynamics in membrane protrusions and vesicle trafficking. Trends Cell Biol. 2006, 16, 522–529. [Google Scholar] [CrossRef]
- Lv, Z.; Hu, M.; Zhen, J.; Lin, J.; Wang, Q.; Wang, R. Rac1/PAK1 signaling promotes epithelial-mesenchymal transition of podocytes in vitro via triggering β-catenin transcriptional activity under high glucose conditions. Int. J. Biochem. Cell Biol. 2013, 45, 255–264. [Google Scholar] [CrossRef]
- Izdebska, M.; Zielińska, W.; Grzanka, D.; Gagat, M. The Role of Actin Dynamics and Actin-Binding Proteins Expression in Epithelial-to-Mesenchymal Transition and Its Association with Cancer Progression and Evaluation of Possible Therapeutic Targets. Biomed Res. Int. 2018, 2018, 4578373. [Google Scholar] [CrossRef]
- Yamazaki, D.; Kurisu, S.; Takenawa, T. Regulation of cancer cell motility through actin reorganization. Cancer Sci. 2005, 96, 379–386. [Google Scholar] [CrossRef]
- Sales, K.J.; Boddy, S.C.; Jabbour, H.N. F-prostanoid receptor alters adhesion, morphology and migration of endometrial adenocarcinoma cells. Oncogene 2008, 27, 2466–2477. [Google Scholar] [CrossRef]
- Burgos-Panadero, R.; Noguera, I.; Cañete, A.; Navarro, S.; Noguera, R. Vitronectin as a molecular player of the tumor microenvironment in neuroblastoma. BMC Cancer 2019, 19, 479. [Google Scholar] [CrossRef]
- Ke, J.; Yang, Y.; Che, Q.; Jiang, F.; Wang, H.; Chen, Z.; Zhu, M.; Tong, H.; Zhang, H.; Yan, X.; et al. Prostaglandin E2 (PGE2) promotes proliferation and invasion by enhancing SUMO-1 activity via EP4 receptor in endometrial cancer. Tumour Biol. 2016, 37, 12203–12211. [Google Scholar] [CrossRef] [PubMed]
- Sales, K.J.; Jabbour, H.N. Cyclooxygenase enzymes and prostaglandins in pathology of the endometrium. Reproduction 2003, 126, 559–567. [Google Scholar] [CrossRef] [PubMed]
- Salker, M.S.; Schierbaum, N.; Alowayed, N.; Singh, Y.; Mack, A.F.; Stournaras, C.; Schäffer, T.E.; Lang, F. LeftyA decreases Actin Polymerization and Stiffness in Human Endometrial Cancer Cells. Sci. Rep. 2016, 6, 29370. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.; Shi, L.; Li, M.; Jiang, N.; Fu, L.; Guo, J. Lefty inhibits glioma growth by suppressing Nodal-activated Smad and ERK1/2 pathways. J. Neurol. Sci. 2014, 347, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Salker, M.S.; Zhou, Y.; Singh, Y.; Brosens, J.; Lang, F. LeftyA sensitive cytosolic pH regulation and glycolytic flux in Ishikawa human endometrial cancer cells. Biochem. Biophys. Res. Commun. 2015, 460, 845–849. [Google Scholar] [CrossRef] [PubMed]
- Damaghi, M.; Wojtkowiak, J.W.; Gillies, R.J. pH sensing and regulation in cancer. Front. Physiol. 2013, 4, 370. [Google Scholar] [CrossRef]
- Stock, C.; Pedersen, S.F. Roles of pH and the Na+/H+ exchanger NHE1 in cancer: From cell biology and animal models to an emerging translational perspective? Semin. Cancer Biol. 2017, 43, 5–16. [Google Scholar] [CrossRef]
- Paradiso, A.; Cardone, R.A.; Bellizzi, A.; Bagorda, A.; Guerra, L.; Tommasino, M.; Casavola, V.; Reshkin, S.J. The Na+-H+ exchanger-1 induces cytoskeletal changes involving reciprocal RhoA and Rac1 signaling, resulting in motility and invasion in MDA-MB-435 cells. Breast Cancer Res. 2004, 6, R616–R628. [Google Scholar] [CrossRef]
- Denker, S.P.; Barber, D.L. Cell migration requires both ion translocation and cytoskeletal anchoring by the Na-H exchanger NHE1. J. Cell Biol. 2002, 159, 1087–1096. [Google Scholar] [CrossRef]
- Reshkin, S.J.; Bellizzi, A.; Albarani, V.; Guerra, L.; Tommasino, M.; Paradiso, A.; Casavola, V. Phosphoinositide 3-kinase is involved in the tumor-specific activation of human breast cancer cell Na+/H+ exchange, motility, and invasion induced by serum deprivation. J. Biol. Chem. 2000, 275, 5361–5369. [Google Scholar] [CrossRef]
- Lin, T.Y.; Wu, F.J.; Chang, C.L.; Li, Z.; Luo, C.W. NMU signaling promotes endometrial cancer cell progression by modulating adhesion signaling. Oncotarget 2016, 7, 10228–10242. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.-Y.; Wu, F.-J.; Lee, W.-Y.; Hsiao, C.-L.; Luo, C.-W. Ovarian regulation of neuromedin U and its local actions in the ovary, mediated through neuromedin U receptor 2. Am. J. Physiol. Endocrinol. Metab. 2013, 304, E800–E809. [Google Scholar] [CrossRef] [PubMed]
- Huveneers, S.; Danen, E.H.J. Adhesion signaling - crosstalk between integrins, Src and Rho. J. Cell Sci. 2009, 122, 1059–1069. [Google Scholar] [CrossRef] [PubMed]
- Prifti, S.; Zourab, Y.; Koumouridis, A.; Bohlmann, M.; Strowitzki, T.; Rabe, T. Role of integrins in invasion of endometrial cancer cell lines. Gynecol. Oncol. 2002, 84, 12–20. [Google Scholar] [CrossRef]
- Li, T.; Pan, H.; Li, R. The dual regulatory role of miR-204 in cancer. Tumour Biol. 2016, 37, 11667–11677. [Google Scholar] [CrossRef]
- Montagnana, M.; Benati, M.; Danese, E.; Giudici, S.; Perfranceschi, M.; Ruzzenenete, O.; Salvagno, G.L.; Bassi, A.; Gelati, M.; Paviati, E.; et al. Aberrant MicroRNA Expression in Patients With Endometrial Cancer. Int. J. Gynecol. Cancer 2017, 27, 459–466. [Google Scholar] [CrossRef]
- Imam, J.S.; Plyler, J.R.; Bansal, H.; Prajapati, S.; Bansal, S.; Rebeles, J.; Chen, H.-I.H.; Chang, Y.-F.; Panneerdoss, S.; Zoghi, B.; et al. Genomic loss of tumor suppressor miRNA-204 promotes cancer cell migration and invasion by activating AKT/mTOR/Rac1 signaling and actin reorganization. PLoS ONE 2012, 7, e52397. [Google Scholar] [CrossRef]
- Dong, F.; Zhang, Q.; Kong, W.; Chen, J.; Ma, J.; Wang, L.; Wang, Y.; Liu, Y.; Li, Y.; Wen, J. Regulation of endometrial cell proliferation by estrogen-induced BDNF signaling pathway. Gynecol. Endocrinol. 2017, 33, 485–489. [Google Scholar] [CrossRef]
- Kataoka, K.; Ogawa, S. Variegated RHOA mutations in human cancers. Exp. Hematol. 2016, 44, 1123–1129. [Google Scholar] [CrossRef]
- Althubiti, M.A. Mutation Frequencies in Endometrial Cancer Patients of Different Ethnicities and Tumor Grades: An Analytical Study. Saudi J. Med. Med. Sci. 2019, 7, 16–21. [Google Scholar] [CrossRef]
- Yotova, I.Y.; Quan, P.; Leditznig, N.; Beer, U.; Wenzl, R.; Tschugguel, W. Abnormal activation of Ras/Raf/MAPK and RhoA/ROCKII signalling pathways in eutopic endometrial stromal cells of patients with endometriosis. Hum. Reprod. 2011, 26, 885–897. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.-Y.; Yang, X.-M.; Cheng, M.-J.; Zhang, R.; Ye, J.; Yi, H.; Ao, J.-P.; Zhang, Z.-G.; Xu, C.-J. Dysregulated cell mechanical properties of endometrial stromal cells from endometriosis patients. Int. J. Clin. Exp. Pathol. 2014, 7, 648–655. [Google Scholar] [PubMed]
- Jiang, C.; Gong, W.; Chen, R.; Ke, H.; Qu, X.; Yang, W.; Cheng, Z. RhoA/ROCK/ARHGAP26 signaling in the eutopic and ectopic endometrium is involved in clinical characteristics of adenomyosis. J. Int. Med. Res. 2018, 46, 5019–5029. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Duan, H.; Zhang, Y.; Sun, F.Q. Abnormal activation of RhoA/ROCK-I signaling in junctional zone smooth muscle cells of patients with adenomyosis. Reprod. Sci. 2016, 23, 333–341. [Google Scholar] [CrossRef]
- Arbyn, M.; Weiderpass, E.; Bruni, L.; de Sanjosé, S.; Saraiya, M.; Ferlay, J.; Bray, F. Estimates of incidence and mortality of cervical cancer in 2018: a worldwide analysis. Lancet Glob Health 2020, 8, e191–e203. [Google Scholar] [CrossRef]
- Denny, L.; Herrero, R.; Levin, C.; Kim, J.J. Cervical Cancer. In Cancer: Disease Control Priorities, 3rd ed.; Gelband, H., Jha, P., Sankaranarayanan, R., Horton, S., Eds.; The International Bank for Reconstruction and Development/The World Bank: Washington, DC, USA, 2015; Volume 3, ISBN 978-1-4648-0349-9. [Google Scholar]
- Rerucha, C.M.; Caro, R.J.; Wheeler, V.L. Cervical Cancer Screening. Am. Fam. Phys. 2018, 97, 441–448. [Google Scholar]
- Enerly, E.; Bonde, J.; Schee, K.; Pedersen, H.; Lönnberg, S.; Nygård, M. Self-Sampling for Human Papillomavirus Testing among Non-Attenders Increases Attendance to the Norwegian Cervical Cancer Screening Programme. PLoS ONE 2016, 11, e0151978. [Google Scholar] [CrossRef]
- Bulten, J.; Horvat, R.; Jordan, J.; Herbert, A.; Wiener, H.; Arbyn, M. European guidelines for quality assurance in cervical histopathology. Acta Oncol. 2011, 50, 611–620. [Google Scholar] [CrossRef]
- Cibula, D.; Planchamp, F.; Fischerova, D.; Fotopoulou, C.; Kohler, C.; Landoni, F.; Mathevet, P.; Naik, R.; Ponce, J.; Raspagliesi, F.; et al. European Society of Gynaecological Oncology quality indicators for surgical treatment of cervical cancer. Int. J. Gynecol. Cancer 2020, 30, 3–14. [Google Scholar] [CrossRef]
- Cibula, D.; Pötter, R.; Planchamp, F.; Avall-Lundqvist, E.; Fischerova, D.; Haie Meder, C.; Köhler, C.; Landoni, F.; Lax, S.; Lindegaard, J.C.; et al. The European Society of Gynaecological Oncology/European Society for Radiotherapy and Oncology/European Society of Pathology Guidelines for the Management of Patients With Cervical Cancer. Int. J. Gynecol. Cancer 2018, 28, 641–655. [Google Scholar] [CrossRef]
- Liu, X.; Chen, D.; Liu, G. Overexpression of RhoA promotes the proliferation and migration of cervical cancer cells. Biosci. Biotechnol. Biochem. 2014, 78, 1895–1901. [Google Scholar] [CrossRef]
- Mendoza-Catalán, M.A.; Cristóbal-Mondragón, G.R.; Adame-Gómez, J.; del Valle-Flores, H.N.; Coppe, J.F.; Sierra-López, L.; Romero-Hernández, M.A.; del Carmen Alarcón-Romero, L.; Illades-Aguiar, B.; Castañeda-Saucedo, E. Nuclear expression of Rac1 in cervical premalignant lesions and cervical cancer cells. BMC Cancer 2012, 12, 116. [Google Scholar] [CrossRef]
- Ma, D.; Cheng, Y.; Zhang, Y.; Guo, Y.; Li, Z.; Li, G. Expression of CDC42 in cervical squamous cell carcinoma and its correlation with clinicopathologic characteristics. Chin. J. Cancer Res. 2013, 25, 6. [Google Scholar] [CrossRef]
- Ruiz-Lafuente, N.; Alcaraz-García, M.-J.; García-Serna, A.-M.; Sebastián-Ruiz, S.; Moya-Quiles, M.-R.; García-Alonso, A.-M.; Parrado, A. Dock10, a Cdc42 and Rac1 GEF, induces loss of elongation, filopodia, and ruffles in cervical cancer epithelial HeLa cells. Biol. Open 2015, 4, 627–635. [Google Scholar] [CrossRef] [PubMed]
- Katoh, H.; Hiramoto, K.; Negishi, M. Activation of Rac1 by RhoG regulates cell migration. J. Cell Sci. 2006, 119, 56–65. [Google Scholar] [CrossRef] [PubMed]
- McKinnon, C.M.; Lygoe, K.A.; Skelton, L.; Mitter, R.; Mellor, H. The atypical Rho GTPase RhoBTB2 is required for expression of the chemokine CXCL14 in normal and cancerous epithelial cells. Oncogene 2008, 27, 6856–6865. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.; Ramdass, B.; Nagarajan, S.; Rehman, M.; Mukherjee, G.; Krishna, S. Notch1 regulates the functional contribution of RhoC to cervical carcinoma progression. Br. J. Cancer 2010, 102, 196–205. [Google Scholar] [CrossRef]
- He, X.; Qian, Y.; Cai, H.; Yang, S.; Cai, J.; Wang, Z. RhoC is essential in TGF-β1 induced epithelial-mesenchymal transition in cervical cancer cells. Oncol. Lett. 2015, 10, 985–989. [Google Scholar] [CrossRef]
- Nishizuka, M.; Komada, R.; Imagawa, M. Knockdown of RhoE Expression Enhances TGF-β-Induced EMT (epithelial-to-mesenchymal transition) in Cervical Cancer HeLa Cells. Int. J. Mol. Sci. 2019, 20, 4697. [Google Scholar] [CrossRef]
- Münger, K.; Baldwin, A.; Edwards, K.M.; Hayakawa, H.; Nguyen, C.L.; Owens, M.; Grace, M.; Huh, K. Mechanisms of human papillomavirus-induced oncogenesis. J. Virol. 2004, 78, 11451–11460. [Google Scholar] [CrossRef]
- Matarrese, P.; Abbruzzese, C.; Mileo, A.M.; Vona, R.; Ascione, B.; Visca, P.; Rollo, F.; Benevolo, M.; Malorni, W.; Paggi, M.G. Interaction between the human papillomavirus 16 E7 oncoprotein and gelsolin ignites cancer cell motility and invasiveness. Oncotarget 2016, 7, 50972–50985. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Oliver, A.W.; He, X.; Borthwick, K.; Donne, A.J.; Hampson, L.; Hampson, I.N. The HPV16 E6 binding protein Tip-1 interacts with ARHGEF16, which activates Cdc42. Br. J. Cancer 2011, 104, 324–331. [Google Scholar] [CrossRef] [PubMed]
- Krishna Subbaiah, V.; Massimi, P.; Boon, S.S.; Myers, M.P.; Sharek, L.; Garcia-Mata, R.; Banks, L. The invasive capacity of HPV transformed cells requires the hDlg-dependent enhancement of SGEF/RhoG activity. PLoS Pathog. 2012, 8, e1002543. [Google Scholar] [CrossRef] [PubMed]
- Todorovic, B.; Nichols, A.C.; Chitilian, J.M.; Myers, M.P.; Shepherd, T.G.; Parsons, S.J.; Barrett, J.W.; Banks, L.; Mymryk, J.S. The human papillomavirus E7 proteins associate with p190RhoGAP and alter its function. J. Virol. 2014, 88, 3653–3663. [Google Scholar] [CrossRef][Green Version]
- Charette, S.T.; McCance, D.J. The E7 protein from human papillomavirus type 16 enhances keratinocyte migration in an Akt-dependent manner. Oncogene 2007, 26, 7386–7390. [Google Scholar] [CrossRef]
- Xu, A.L.; Yu, G.Q.; Kong, X.C.; Qiu, X.H.; Li, P.L. Effect of Rac1 downregulation mediated by shRNA on the biological behaviour of human cervical cancer cells. J. Int. Med. Res. 2013, 41, 1037–1048. [Google Scholar] [CrossRef]
- Chakerzehi, A.; Arvanagh, N.; Saedi, S.; Hematti, M.; Ardakani, J.; Moradi, A.; Shokouhi, A. Effect of Quercetin on RAC1 Gene Expression as a Marker of Metastasis in Cervical Cancer Cells. Zahedan J. Res. Med. Sci. 2015, 17. [Google Scholar] [CrossRef]
- Becker, M.S.; Müller, P.M.; Bajorat, J.; Schroeder, A.; Giaisi, M.; Amin, E.; Ahmadian, M.R.; Rocks, O.; Köhler, R.; Krammer, P.H.; et al. The anticancer phytochemical rocaglamide inhibits Rho GTPase activity and cancer cell migration. Oncotarget 2016, 7, 51908–51921. [Google Scholar] [CrossRef]
- Osaki, J.H.; Espinha, G.; Magalhaes, Y.T.; Forti, F.L. Modulation of RhoA GTPase Activity Sensitizes Human Cervix Carcinoma Cells to γ-Radiation by Attenuating DNA Repair Pathways. Oxid. Med. Cell Longev. 2016, 2016, 6012642. [Google Scholar] [CrossRef]
- Pranatharthi, A.; Thomas, P.; Udayashankar, A.H.; Bhavani, C.; Suresh, S.B.; Krishna, S.; Thatte, J.; Srikantia, N.; Ross, C.R.; Srivastava, S. RhoC regulates radioresistance via crosstalk of ROCK2 with the DNA repair machinery in cervical cancer. J. Exp. Clin. Cancer Res. 2019, 38, 392. [Google Scholar] [CrossRef]
- Zendehdel, M.; Niakan, B.; Keshtkar, A.; Rafiei, E.; Salamat, F. Subtypes of Benign Breast Disease as a Risk Factor for Breast Cancer: A Systematic Review and Meta-Analysis Protocol. Iran J. Med. Sci. 2018, 43, 1–8. [Google Scholar] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- Neil-Sztramko, S.E.; Winters-Stone, K.M.; Bland, K.A.; Campbell, K.L. Updated systematic review of exercise studies in breast cancer survivors: attention to the principles of exercise training. Br. J. Sports Med. 2019, 53, 504–512. [Google Scholar] [CrossRef] [PubMed]
- Momenimovahed, Z.; Salehiniya, H. Epidemiological characteristics of and risk factors for breast cancer in the world. Breast Cancer 2019, 11, 151–164. [Google Scholar] [CrossRef] [PubMed]
- Little, A.C.; Pathanjeli, P.; Wu, Z.; Bao, L.; Goo, L.E.; Yates, J.A.; Oliver, C.R.; Soellner, M.B.; Merajver, S.D. IL-4/IL-13 Stimulated Macrophages Enhance Breast Cancer Invasion Via Rho-GTPase Regulation of Synergistic VEGF/CCL-18 Signaling. Front. Oncol. 2019, 9, 456. [Google Scholar] [CrossRef] [PubMed]
- Gómez del Pulgar, T.; Benitah, S.A.; Valerón, P.F.; Espina, C.; Lacal, J.C. Rho GTPase expression in tumourigenesis: evidence for a significant link. Bioessays 2005, 27, 602–613. [Google Scholar] [CrossRef]
- O’Hayre, M.; Inoue, A.; Kufareva, I.; Wang, Z.; Mikelis, C.M.; Drummond, R.A.; Avino, S.; Finkel, K.; Kalim, K.W.; DiPasquale, G.; et al. Inactivating mutations in GNA13 and RHOA in Burkitt’s lymphoma and diffuse large B-cell lymphoma: a tumor suppressor function for the Gα13/RhoA axis in B cells. Oncogene 2016, 35, 3771–3780. [Google Scholar] [CrossRef]
- Zandvakili, I.; Lin, Y.; Morris, J.C.; Zheng, Y. Rho GTPases: Anti- or pro-neoplastic targets? Oncogene 2017, 36, 3213–3222. [Google Scholar] [CrossRef]
- Lawson, C.D.; Fan, C.; Mitin, N.; Baker, N.M.; George, S.D.; Graham, D.M.; Perou, C.M.; Burridge, K.; Der, C.J.; Rossman, K.L. Rho GTPase Transcriptome Analysis Reveals Oncogenic Roles for Rho GTPase-Activating Proteins in Basal-like Breast Cancers. Cancer Res. 2016, 76, 3826–3837. [Google Scholar] [CrossRef]
- Humphries, B.; Wang, Z.; Li, Y.; Jhan, J.-R.; Jiang, Y.; Yang, C. ARHGAP18 Downregulation by miR-200b Suppresses Metastasis of Triple-Negative Breast Cancer by Enhancing Activation of RhoA. Cancer Res. 2017, 77, 4051–4064. [Google Scholar] [CrossRef]
- Kalpana, G.; Figy, C.; Yeung, M.; Yeung, K.C. Reduced RhoA expression enhances breast cancer metastasis with a concomitant increase in CCR5 and CXCR4 chemokines signaling. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, J.; Lai, X.-N.; Jiao, X.-Q.; Xiong, J.-P.; Xiong, L.-X. Focus on Cdc42 in Breast Cancer: New Insights, Target Therapy Development and Non-Coding RNAs. Cells 2019, 8, 146. [Google Scholar] [CrossRef]
- Yadav, S.; Barton, M.; Nguyen, N.-T. Stretching Induces Overexpression of RhoA and Rac1 GTPases in Breast Cancer Cells. Adv. Biosyst. 2020, 4, e1900222. [Google Scholar] [CrossRef]
- Lin, Y.; Zheng, Y. Approaches of targeting Rho GTPases in cancer drug discovery. Expert Opin. Drug Discov. 2015, 10, 991–1010. [Google Scholar] [CrossRef] [PubMed]
- Ohta, T.; Takahashi, T.; Shibuya, T.; Amita, M.; Henmi, N.; Takahashi, K.; Kurachi, H. Inhibition of the Rho/ROCK pathway enhances the efficacy of cisplatin through the blockage of hypoxia-inducible factor-1α in human ovarian cancer cells. Cancer Biol. Ther. 2012, 13, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Gui, P.; Labrousse, A.; Van Goethem, E.; Besson, A.; Maridonneau-Parini, I.; Le Cabec, V. Rho/ROCK pathway inhibition by the CDK inhibitor p27(kip1) participates in the onset of macrophage 3D-mesenchymal migration. J. Cell Sci. 2014, 127, 4009–4023. [Google Scholar] [CrossRef] [PubMed]
- Rath, N.; Munro, J.; Cutiongco, M.F.; Jagiełło, A.; Gadegaard, N.; McGarry, L.; Unbekandt, M.; Michalopoulou, E.; Kamphorst, J.J.; Sumpton, D.; et al. Rho Kinase Inhibition by AT13148 Blocks Pancreatic Ductal Adenocarcinoma Invasion and Tumor Growth. Cancer Res. 2018, 78, 3321–3336. [Google Scholar] [CrossRef] [PubMed]
- Uehata, M.; Ishizaki, T.; Satoh, H.; Ono, T.; Kawahara, T.; Morishita, T.; Tamakawa, H.; Yamagami, K.; Inui, J.; Maekawa, M.; et al. Calcium sensitization of smooth muscle mediated by a Rho-associated protein kinase in hypertension. Nature 1997, 389, 990–994. [Google Scholar] [CrossRef]
- Chen, R.; Cheng, Y.; Zhang, Y.; Li, Z.; Geng, L. RhoC mediates invasion and migration of CaSki cells through the Rho-associated serine-threonine protein kinase 1 signaling pathway. Int. J. Gynecol. Cancer 2014, 24, 184–191. [Google Scholar] [CrossRef]
- He, M.; Cheng, Y.; Li, W.; Liu, Q.; Liu, J.; Huang, J.; Fu, X. Vascular endothelial growth factor C promotes cervical cancer metastasis via up-regulation and activation of RhoA/ROCK-2/moesin cascade. BMC Cancer 2010, 10, 170. [Google Scholar] [CrossRef]
- Ghasemi, A.; Hashemy, S.I.; Aghaei, M.; Panjehpour, M. RhoA/ROCK pathway mediates leptin-induced uPA expression to promote cell invasion in ovarian cancer cells. Cell. Signal. 2017, 32, 104–114. [Google Scholar] [CrossRef]
- Jeong, K.J.; Park, S.Y.; Cho, K.H.; Sohn, J.S.; Lee, J.; Kim, Y.K.; Kang, J.; Park, C.G.; Han, J.W.; Lee, H.Y. The Rho/ROCK pathway for lysophosphatidic acid-induced proteolytic enzyme expression and ovarian cancer cell invasion. Oncogene 2012, 31, 4279–4289. [Google Scholar] [CrossRef] [PubMed]
- Tiftik, R.N.; Un, I.; Yetkin, D.; Buyukafsar, K. Role of the Rho/Rho-kinase Pathway in Endometrial Adenocarcinoma Cell Proliferation. Int. J. Hemat. Oncol. 2019, 29, 8. [Google Scholar] [CrossRef]
- Tanaka, K.; Minami, H.; Kota, M.; Kuwamura, K.; Kohmura, E. Treatment of cerebral vasospasm with intra-arterial fasudil hydrochloride. Neurosurgery 2005, 56, 214–223. [Google Scholar] [CrossRef] [PubMed]
- Ogata, S.; Morishige, K.-I.; Sawada, K.; Hashimoto, K.; Mabuchi, S.; Kawase, C.; Ooyagi, C.; Sakata, M.; Kimura, T. Fasudil Inhibits Lysophosphatidic Acid-Induced Invasiveness of Human Ovarian Cancer Cells. Int. J. Gynecol. Cancer 2009, 19, 1473–1480. [Google Scholar] [CrossRef]
- Yin, Z.; Sun, J. Curcumin induces human SKOV3 cell apoptosis via the activation of Rho-kinase. Eur. J. Gynaecol. Oncol. 2014, 35, 433–437. [Google Scholar] [PubMed]
- Ferraresi, A.; Phadngam, S.; Morani, F.; Galetto, A.; Alabiso, O.; Chiorino, G.; Isidoro, C. Resveratrol inhibits IL-6-induced ovarian cancer cell migration through epigenetic up-regulation of autophagy. Mol. Carcinog. 2017, 56, 1164–1181. [Google Scholar] [CrossRef]
- Hudson, L.G.; Cook, L.S.; Grimes, M.M.; Muller, C.Y.; Adams, S.F.; Wandinger-Ness, A. Dual Actions of Ketorolac in Metastatic Ovarian Cancer. Cancers 2019, 11, 1049. [Google Scholar] [CrossRef]
- Oprea, T.I.; Sklar, L.A.; Agola, J.O.; Guo, Y.; Silberberg, M.; Roxby, J.; Vestling, A.; Romero, E.; Surviladze, Z.; Murray-Krezan, C.; et al. Novel Activities of Select NSAID R-Enantiomers against Rac1 and Cdc42 GTPases. PLoS ONE 2015, 10, e0142182. [Google Scholar] [CrossRef]
- Hong, L.; Kenney, S.R.; Phillips, G.K.; Simpson, D.; Schroeder, C.E.; Nöth, J.; Romero, E.; Swanson, S.; Waller, A.; Strouse, J.J.; et al. Characterization of a Cdc42 Protein Inhibitor and Its Use as a Molecular Probe. J. Biol. Chem. 2013, 288, 8531–8543. [Google Scholar] [CrossRef]
- Pinzaglia, M.; Montaldo, C.; Polinari, D.; Simone, M.; La Teana, A.; Tripodi, M.; Mancone, C.; Londei, P.; Benelli, D. eIF6 over-expression increases the motility and invasiveness of cancer cells by modulating the expression of a critical subset of membrane-bound proteins. BMC Cancer 2015, 15, 131. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Peng, F.; Zhong, Y.; Chen, Y.; Tang, M.; Li, D. Rhein suppresses matrix metalloproteinase production by regulating the Rac1/ROS/MAPK/AP-1 pathway in human ovarian carcinoma cells. Int. J. Oncol. 2017, 50, 933–941. [Google Scholar] [CrossRef] [PubMed]
- Tsang, C.M.; Lau, E.P.W.; Di, K.; Cheung, P.Y.; Hau, P.M.; Ching, Y.P.; Wong, Y.C.; Cheung, A.L.M.; Wan, T.S.K.; Tong, Y.; et al. Berberine inhibits Rho GTPases and cell migration at low doses but induces G2 arrest and apoptosis at high doses in human cancer cells. Int. J. Mol. Med. 2009, 24, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Meng, X.; Wu, D.; Qiu, Z.; Luo, H. A Natural Isoquinoline Alkaloid With Antitumor Activity: Studies of the Biological Activities of Berberine. Front. Pharmacol. 2019, 10, 9. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, S. Berberine suppresses growth and metastasis of endometrial cancer cells via miR-101/COX-2. Biomed. Pharmacother. 2018, 103, 1287–1293. [Google Scholar] [CrossRef]
- Cox, A.D.; Der, C.J.; Philips, M.R. Targeting RAS Membrane Association: Back to the Future for Anti-RAS Drug Discovery? Clin. Cancer Res. 2015, 21, 1819–1827. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Kashima, H.; Rahmanto, Y.S.; Banno, K.; Yu, Y.; Matoba, Y.; Watanabe, K.; Iijima, M.; Takeda, T.; Kunitomi, H.; et al. Drug repositioning of mevalonate pathway inhibitors as antitumor agents for ovarian cancer. Oncotarget 2017, 8, 72147–72156. [Google Scholar] [CrossRef]
- Wärnberg, F.; White, D.; Anderson, E.; Knox, F.; Clarke, R.B.; Morris, J.; Bundred, N.J. Effect of a farnesyl transferase inhibitor (R115777) on ductal carcinoma in situ of the breast in a human xenograft model and on breast and ovarian cancer cell growth in vitro and in vivo. Breast Cancer Res. 2006, 8, R21. [Google Scholar] [CrossRef]
- Møller, L.L.V.; Klip, A.; Sylow, L. Rho GTPases—Emerging Regulators of Glucose Homeostasis and Metabolic Health. Cells 2019, 8, 434. [Google Scholar] [CrossRef]
- Tolias, K.F.; Duman, J.G.; Um, K. Control of synapse development and plasticity by Rho GTPase regulatory proteins. Prog. Neurobiol. 2011, 94, 133–148. [Google Scholar] [CrossRef]
- Joseph, J.; Radulovich, N.; Wang, T.; Raghavan, V.; Zhu, C.-Q.; Tsao, M.-S. Rho guanine nucleotide exchange factor ARHGEF10 is a putative tumor suppressor in pancreatic ductal adenocarcinoma. Oncogene 2020, 39, 308–321. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Sui, L.; Zhuang, J.; He, S.; Song, Y.; Ye, Y.; Xia, W. ARHGAP24 regulates cell ability and apoptosis of colorectal cancer cells via the regulation of P53. Oncol. Lett. 2018, 16, 3517–3524. [Google Scholar] [CrossRef] [PubMed]
- Surviladze, Z.; Waller, A.; Strouse, J.J.; Bologa, C.; Ursu, O.; Salas, V.; Parkinson, J.F.; Phillips, G.K.; Romero, E.; Wandinger-Ness, A.; et al. A Potent and Selective Inhibitor of Cdc42 GTPase. In Probe Reports from the NIH Molecular Libraries Program; National Center for Biotechnology Information (US): Bethesda, MD, USA, 2010. [Google Scholar]
- Schrade, K.; Tröger, J.; Eldahshan, A.; Zühlke, K.; Abdul Azeez, K.R.; Elkins, J.M.; Neuenschwander, M.; Oder, A.; Elkewedi, M.; Jaksch, S.; et al. An AKAP-Lbc-RhoA interaction inhibitor promotes the translocation of aquaporin-2 to the plasma membrane of renal collecting duct principal cells. PLoS One 2018, 13. [Google Scholar] [CrossRef] [PubMed]
- Diviani, D.; Raimondi, F.; Del Vescovo, C.D.; Dreyer, E.; Reggi, E.; Osman, H.; Ruggieri, L.; Gonano, C.; Cavin, S.; Box, C.L.; et al. Small-Molecule Protein-Protein Interaction Inhibitor of Oncogenic Rho Signaling. Cell Chem. Biol. 2016, 23, 1135–1146. [Google Scholar] [CrossRef]
- Vives, V.; Cres, G.; Richard, C.; Busson, M.; Ferrandez, Y.; Planson, A.-G.; Zeghouf, M.; Cherfils, J.; Malaval, L.; Blangy, A. Pharmacological inhibition of Dock5 prevents osteolysis by affecting osteoclast podosome organization while preserving bone formation. Nat. Commun. 2015, 6, 6218. [Google Scholar] [CrossRef]
- Bouquier, N.; Vignal, E.; Charrasse, S.; Weill, M.; Schmidt, S.; Léonetti, J.-P.; Blangy, A.; Fort, P. A cell active chemical GEF inhibitor selectively targets the Trio/RhoG/Rac1 signaling pathway. Chem. Biol. 2009, 16, 657–666. [Google Scholar] [CrossRef]
- Kudela, E.; Samec, M.; Kubatka, P.; Nachajova, M.; Laucekova, Z.; Liskova, A.; Dokus, K.; Biringer, K.; Simova, D.; Gabonova, E.; et al. Breast Cancer in Young Women: Status Quo and Advanced Disease Management by a Predictive, Preventive, and Personalized Approach. Cancers 2019, 11, 1791. [Google Scholar] [CrossRef]
- Carlomagno, N.; Incollingo, P.; Tammaro, V.; Peluso, G.; Rupealta, N.; Chiacchio, G.; Sandoval Sotelo, M.L.; Minieri, G.; Pisani, A.; Riccio, E.; et al. Diagnostic, Predictive, Prognostic, and Therapeutic Molecular Biomarkers in Third Millennium: A Breakthrough in Gastric Cancer. Biomed. Res. Int. 2017, 2017. [Google Scholar] [CrossRef]
- Huang, J.; Hu, W.; Sood, A.K. Prognostic Biomarkers in Ovarian Cancer. Cancer Biomark. 2010, 8, 231–251. [Google Scholar] [CrossRef]
- Zhao, Y.; Zong, Z.; Xu, H. RhoC expression level is correlated with the clinicopathological characteristics of ovarian cancer and the expression levels of ROCK-I, VEGF, and MMP9. Gynecol. Oncol. 2010, 116, 563–571. [Google Scholar] [CrossRef]
- Wang, C.; Wang, W.; Liu, Y.; Yong, M.; Yang, Y.; Zhou, H. Rac GTPase activating protein 1 promotes oncogenic progression of epithelial ovarian cancer. Cancer Sci. 2018, 109, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Chen, S.; Li, Y.; Gao, Y.; Huang, S.; Li, H.; Zhu, Y. SMURF1-mediated ubiquitination of ARHGAP26 promotes ovarian cancer cell invasion and migration. Exp. Mol. Med. 2019, 51, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Luo, N.; Guo, J.; Chen, L.; Yang, W.; Qu, X.; Cheng, Z. ARHGAP10, downregulated in ovarian cancer, suppresses tumorigenicity of ovarian cancer cells. Cell Death Dis. 2016, 7, e2157. [Google Scholar] [CrossRef] [PubMed]
- Leng, R.; Liao, G.; Wang, H.; Kuang, J.; Tang, L. Rac1 expression in epithelial ovarian cancer: effect on cell EMT and clinical outcome. Med. Oncol. 2015, 32, 28. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Cui, X.; Chen, D.; Yang, Y.; Piao, J.; Lin, Z.; Yan, G.; Shen, D. Clinical implication of Tiam1 overexpression in the prognosis of patients with serous ovarian carcinoma. Oncol. Lett. 2016, 12, 3492–3498. [Google Scholar] [CrossRef]
- Wang, W.; Du, H.; Liu, H.; Hu, F.; Liu, G. SMAD specific E3 ubiquitin protein ligase 1 promotes ovarian cancer cell migration and invasion via the activation of the RhoA/ROCK signaling pathway. Oncol. Rep. 2018, 41, 668–676. [Google Scholar] [CrossRef]
- Tanaka, K.; Matsumoto, Y.; Ishikawa, H.; Fukumitsu, N.; Numajiri, H.; Murofushi, K.; Oshiro, Y.; Okumura, T.; Satoh, T.; Sakurai, H. Impact of RhoA overexpression on clinical outcomes in cervical squamous cell carcinoma treated with concurrent chemoradiotherapy. J. Radiat. Res. 2020, 61, 221–230. [Google Scholar] [CrossRef]
- Yang, Y.; Wu, Q.; Li, N.; Che, S.; Jin, T.; Nan, Y.; Lin, Z.; Chen, L. Upregulation of Tiam1 contributes to cervical cancer disease progression and indicates poor survival outcome. Hum. Pathol. 2018, 75, 179–188. [Google Scholar] [CrossRef]
- Orgaz, J.L.; Herraiz, C.; Sanz-Moreno, V. Rho GTPases modulate malignant transformation of tumor cells. Small GTPases 2014, 5, e983867. [Google Scholar] [CrossRef]
- Boesch, M.; Sopper, S.; Marth, C.; Fiegl, H.; Wiedemair, A.; Rössler, J.; Hatina, J.; Wolf, D.; Reimer, D.; Zeimet, A.G. Evaluation of Vav3.1 as prognostic marker in endometrial cancer. J. Cancer Res. Clin. Oncol. 2018, 144, 2067–2076. [Google Scholar] [CrossRef]
- Chen, S.; Chen, X.; Xiu, Y.-L.; Sun, K.-X.; Zhao, Y. Inhibition of Ovarian Epithelial Carcinoma Tumorigenesis and Progression by microRNA 106b Mediated through the RhoC Pathway. PLoS One 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Chen, S.; Xiu, Y.-L.; Sun, K.-X.; Zong, Z.-H.; Zhao, Y. RhoC is a major target of microRNA-93-5P in epithelial ovarian carcinoma tumorigenesis and progression. Mol. Cancer 2015, 14, 31. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Nakayama, I.; Shibazaki, M.; Yashima-Abo, A.; Miura, F.; Sugiyama, T.; Masuda, T.; Maesawa, C. Loss of HOXD10 expression induced by upregulation of miR-10b accelerates the migration and invasion activities of ovarian cancer cells. Int. J. Oncol. 2013, 43, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.H.; Song, J.-Y.; Park, H.; Jeong, J.-Y.; Kwon, A.-Y.; Heo, J.H.; Kang, H.; Kim, G.; An, H.J. miR-145, targeting high-mobility group A2, is a powerful predictor of patient outcome in ovarian carcinoma. Cancer Lett. 2015, 356, 937–945. [Google Scholar] [CrossRef]
- Sang, X.-B.; Zong, Z.-H.; Wang, L.-L.; Wu, D.-D.; Chen, S.; Liu, B.-L.; Zhao, Y. E2F-1 targets miR-519d to regulate the expression of the ras homolog gene family member C. Oncotarget 2017, 8, 14777–14793. [Google Scholar] [CrossRef]
- Wang, Y.; Li, J.; Xu, C.; Zhang, X. MicroRNA-139-5p Inhibits Cell Proliferation and Invasion by Targeting RHO-Associated Coiled-Coil-Containing Protein Kinase 2 in Ovarian Cancer. Oncol. Res. 2018, 26, 411–420. [Google Scholar] [CrossRef]
- Mei, J.; Huang, Y.; Hao, L.; Liu, Y.; Yan, T.; Qiu, T.; Xu, R.; Xu, B.; Xiao, Z.; Jiang, X.; et al. DAAM1-mediated migration and invasion of ovarian cancer cells are suppressed by miR-208a-5p. Pathol. Res. Pract. 2019, 215, 152452. [Google Scholar] [CrossRef]
- Liu, B.-L.; Sun, K.-X.; Zong, Z.-H.; Chen, S.; Zhao, Y. MicroRNA-372 inhibits endometrial carcinoma development by targeting the expression of the Ras homolog gene family member C (RhoC). Oncotarget 2015, 7, 6649–6664. [Google Scholar] [CrossRef]
- Howe, E.N.; Cochrane, D.R.; Richer, J.K. Targets of miR-200c mediate suppression of cell motility and anoikis resistance. Breast Cancer Res. 2011, 13, R45. [Google Scholar] [CrossRef]
- Cheng, Y.-X.; Chen, G.-T.; Chen, C.; Zhang, Q.-F.; Pan, F.; Hu, M.; Li, B.-S. MicroRNA-200b inhibits epithelial-mesenchymal transition and migration of cervical cancer cells by directly targeting RhoE. Mol. Med. Rep. 2016, 13, 3139–3146. [Google Scholar] [CrossRef]
- Dong, J.; Wang, M.; Ni, D.; Zhang, L.; Wang, W.; Cui, X.; Fu, S.; Yao, S. MicroRNA-217 functions as a tumor suppressor in cervical cancer cells through targeting Rho-associated protein kinase 1. Oncol. Lett. 2018, 16, 5535–5542. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Wang, L.; Pan, X.; Yang, C. lncRNA OIP5-AS1 targets ROCK1 to promote cell proliferation and inhibit cell apoptosis through a mechanism involving miR-143-3p in cervical cancer. Braz. J. Med. Biol. Res. 2020, 53, e8883. [Google Scholar] [CrossRef]
- Agostini, A.; Brunetti, M.; Davidson, B.; Tropé, C.G.; Eriksson, A.G.Z.; Heim, S.; Panagopoulos, I.; Micci, F. The microRNA miR-192/215 family is upregulated in mucinous ovarian carcinomas. Sci. Rep. 2018, 8, 11069. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Chen, X. Identification of potential biomarkers in cervical cancer with combined public mRNA and miRNA expression microarray data analysis. Oncol. Lett. 2018, 16, 5200–5208. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zheng, H.-C.; Chen, S.; Gou, W.-F.; Xiao, L.-J.; Niu, Z.-F. The role of RhoC in ovarian epithelial carcinoma: A marker for carcinogenesis, progression, prognosis, and target therapy. Gynecol. Oncol. 2013, 130, 570–578. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Wu, Z.; Li, J.; Yang, X.; Wang, Y.; Yu, Y.; Ye, J.; Xu, C.; Qin, W.; Zhang, Z. MCAM is a novel metastasis marker and regulates spreading, apoptosis and invasion of ovarian cancer cells. Tumor Biol. 2012, 33, 1619–1628. [Google Scholar] [CrossRef]
- Wu, N.; Ren, D.; Li, S.; Ma, W.; Hu, S.; Jin, Y.; Xiao, S. RCC2 over-expression in tumor cells alters apoptosis and drug sensitivity by regulating Rac1 activation. BMC Cancer 2018, 18, 67. [Google Scholar] [CrossRef]
- Golubnitschaja, O.; Debald, M.; Yeghiazaryan, K.; Kuhn, W.; Pešta, M.; Costigliola, V.; Grech, G. Breast cancer epidemic in the early twenty-first century: evaluation of risk factors, cumulative questionnaires and recommendations for preventive measures. Tumor Biol. 2016, 37, 12941–12957. [Google Scholar] [CrossRef]
- Janssens, J.P.; Schuster, K.; Voss, A. Preventive, predictive, and personalized medicine for effective and affordable cancer care. EPMA J. 2018, 9, 113–123. [Google Scholar] [CrossRef]
- Li, H.; Peyrollier, K.; Kilic, G.; Brakebusch, C. Rho GTPases and cancer. Biofactors 2014, 40, 226–235. [Google Scholar] [CrossRef]
- Polivka, J.; Altun, I.; Golubnitschaja, O. Pregnancy-associated breast cancer: the risky status quo and new concepts of predictive medicine. EPMA J. 2018, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Lisanti, M.P.; Reeves, K.; Peiris-Pagès, M.; Chadwick, A.L.; Sanchez-Alvarez, R.; Howell, A.; Martinez-Outschoorn, U.E.; Sotgia, F. JNK1 stress signaling is hyper-activated in high breast density and the tumor stroma: Connecting fibrosis, inflammation, and stemness for cancer prevention. Cell Cycle 2014, 13, 580–599. [Google Scholar] [CrossRef] [PubMed]
- Golubnitschaja, O.; Yeghiazaryan, K.; Abraham, J.-A.; Schild, H.H.; Costigliola, V.; Debald, M.; Kuhn, W. Breast cancer risk assessment: a non-invasive multiparametric approach to stratify patients by MMP-9 serum activity and RhoA expression patterns in circulating leucocytes. Amino Acids 2017, 49, 273–281. [Google Scholar] [CrossRef]
- Fröhlich, H.; Patjoshi, S.; Yeghiazaryan, K.; Kehrer, C.; Kuhn, W.; Golubnitschaja, O. Premenopausal breast cancer: potential clinical utility of a multi-omics based machine learning approach for patient stratification. EPMA J. 2018, 9, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Zhan, X. The crucial role of multiomic approach in cancer research and clinically relevant outcomes. EPMA J. 2018, 9, 77–102. [Google Scholar] [CrossRef] [PubMed]
- Gerner, C.; Costigliola, V.; Golubnitschaja, O. Multiomic patterns in body fluids: Technological challenge with a great potential to implement the advances paradigm of 3P medicine. Mass Spectrom. Rev. 2019. [Google Scholar] [CrossRef]
- Golubnitschaja, O.; Filep, N.; Yeghiazaryan, K.; Blom, H.J.; Hofmann-Apitius, M.; Kuhn, W. Multi-omic approach decodes paradoxes of the triple-negative breast cancer: lessons for predictive, preventive and personalised medicine. Amino Acids 2018, 50, 383–395. [Google Scholar] [CrossRef]
- Bubnov, R.; Polivka, J.; Zubor, P.; Konieczka, K.; Golubnitschaja, O. “Pre-metastatic niches” in breast cancer: are they created by or prior to the tumour onset? “Flammer Syndrome” relevance to address the question. EPMA J. 2017, 8, 141–157. [Google Scholar] [CrossRef]
- Qian, S.; Golubnitschaja, O.; Zhan, X. Chronic inflammation: key player and biomarker-set to predict and prevent cancer development and progression based on individualized patient profiles. EPMA J. 2019, 10, 365–381. [Google Scholar] [CrossRef]
- Maturo, M.G.; Soligo, M.; Gibson, G.; Manni, L.; Nardini, C. The greater inflammatory pathway-high clinical potential by innovative predictive, preventive, and personalized medical approach. EPMA J. 2020, 11, 1–16. [Google Scholar] [CrossRef]
- Abedi, F.; Hayes, A.W.; Reiter, R.; Karimi, G. Acute lung injury: The therapeutic role of Rho kinase inhibitors. Pharmacol. Res. 2020, 155, 104736. [Google Scholar] [CrossRef]
- Avishai, E.; Yeghiazaryan, K.; Golubnitschaja, O. Impaired wound healing: facts and hypotheses for multi-professional considerations in predictive, preventive and personalised medicine. EPMA J. 2017, 8, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Porazinski, S.; Parkin, A.; Pajic, M. Rho-ROCK Signaling in Normal Physiology and as a Key Player in Shaping the Tumor Microenvironment. Adv. Exp. Med. Biol. 2020, 1223, 99–127. [Google Scholar] [CrossRef] [PubMed]
- Dankel, S.N.; Røst, T.H.; Kulyté, A.; Fandalyuk, Z.; Skurk, T.; Hauner, H.; Sagen, J.V.; Rydén, M.; Arner, P.; Mellgren, G. The Rho GTPase RND3 regulates adipocyte lipolysis. Metab. Clin. Exp. 2019, 101, 153999. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Wang, X.-Q. Regulation of mTORC1 by Small GTPases in Response to Nutrients. J. Nutr. 2020, 150, 1004–1011. [Google Scholar] [CrossRef] [PubMed]
- Kapinova, A.; Kubatka, P.; Golubnitschaja, O.; Kello, M.; Zubor, P.; Solar, P.; Pec, M. Dietary phytochemicals in breast cancer research: anticancer effects and potential utility for effective chemoprevention. Environ. Health. Prev. Med. 2018, 23, 36. [Google Scholar] [CrossRef] [PubMed]
- Uramova, S.; Kubatka, P.; Dankova, Z.; Kapinova, A.; Zolakova, B.; Samec, M.; Zubor, P.; Zulli, A.; Valentova, V.; Kwon, T.K.; et al. Plant natural modulators in breast cancer prevention: status quo and future perspectives reinforced by predictive, preventive, and personalized medical approach. EPMA J. 2018, 9, 403–419. [Google Scholar] [CrossRef]
- Tsakiroglou, P.; VandenAkker, N.E.; Del Bo’, C.; Riso, P.; Klimis-Zacas, D. Role of Berry Anthocyanins and Phenolic Acids on Cell Migration and Angiogenesis: An Updated Overview. Nutrients 2019, 11, 1075. [Google Scholar] [CrossRef]
- Wang, J.; Liu, Y.; Zhao, J.; Zhang, W.; Pang, X. Saponins extracted from by-product of Asparagus officinalis L. suppress tumour cell migration and invasion through targeting Rho GTPase signalling pathway. J. Sci. Food Agric. 2013, 93, 1492–1498. [Google Scholar] [CrossRef]
- Yu, G.; Wang, Z.; Zeng, S.; Liu, S.; Zhu, C.; Xu, R.; Liu, R.-E. Paeoniflorin Inhibits Hepatocyte Growth Factor- (HGF-) Induced Migration and Invasion and Actin Rearrangement via Suppression of c-Met-Mediated RhoA/ROCK Signaling in Glioblastoma. Biomed. Res. Int. 2019, 2019, 9053295. [Google Scholar] [CrossRef]
- Kunin, A.; Polivka, J.; Moiseeva, N.; Golubnitschaja, O. “Dry mouth” and “Flammer” syndromes—neglected risks in adolescents and new concepts by predictive, preventive and personalised approach. EPMA J. 2018, 9, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Goncharenko, V.; Bubnov, R.; Polivka, J.; Zubor, P.; Biringer, K.; Bielik, T.; Kuhn, W.; Golubnitschaja, O. Vaginal dryness: individualised patient profiles, risks and mitigating measures. EPMA J. 2019, 10, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Seifirad, S.; Haghpanah, V. Inappropriate modeling of chronic and complex disorders: How to reconsider the approach in the context of predictive, preventive and personalized medicine, and translational medicine. EPMA J. 2019, 10, 195–209. [Google Scholar] [CrossRef] [PubMed]
- Duarte, A.A.; Mohsin, S.; Golubnitschaja, O. Diabetes care in figures: current pitfalls and future scenario. EPMA J. 2018, 9, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Bubnov, R.; Babenko, L.; Lazarenko, L.; Kryvtsova, M.; Shcherbakov, O.; Zholobak, N.; Golubnitschaja, O.; Spivak, M. Can tailored nanoceria act as a prebiotic? Report on improved lipid profile and gut microbiota in obese mice. EPMA J. 2019, 10, 317–335. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Therapeutics | Cancer Types | Target | Reference |
|---|---|---|---|
| Y-27632 | Cervical CaSki cells | ↓ ROCK-1, ↑ RhoC | [170] |
| Cervical SiHa cells | ↓ ROCK-2 | [171] | |
| Ovarian CAOV-3, PA-1 cells | ↓ LPA | [173] | |
| Endometrial HEC-1A cells | Reversed effect on LPA and calpeptin-induced cell proliferation | [174] | |
| C3 + Y-27632 | Ovarian OVCAR3, SKOV-3, CAOV-3 cells | ↓ leptin-induced uPA | [172] |
| Fasudil | Ovarian SKOV-3ip1 cells and xenografts | ↓ intracellular cytoskeletal rearrangement, ↓ tyrosine phosphorylation of paxillin, ↓ serine phosphorylation of myosin light chain | [176] |
| Ovarian A2780 cells | Modulation of HIF-1α | [166] | |
| NSC23766 | Cervical C33A, SiHa cells | ↓ Rac1 | [133] |
| Rhein | Ovarian SKOV-3-PM4 | Modulation of MMPs and Rac1/ROS/MAPK/AP-1 | [183] |
| R-ketorolac | Ovarian SKOV-3ip and primary patients derived ovarian cancer cells | Similar effect as NSC23766 and CID2950007/ML141 | [71] |
| Racemic ketorolac | Ovarian cancer patients | ↓ Rac1, ↓ Cdc42 | [70] |
| R-naproxen | Ovarian OvCa429, OvCa433, cervical HeLa T4 + cells | ↓ Rac1, ↓ Cdc42 | [180] |
| ML-141 | Ovarian A2780 cells | ↓ Cdc42 | [182] |
| Curcumin | Ovarian SKOV-3 cells | Activation of RhoA/Rho-kinase signaling | [177] |
| Resveratrol | Ovarian OVCAR-3 cells | ↓ ARH-I, ↓ LC3-positive autophagic vacuoles formation | [178] |
| Type of Cancer | Type of Study | Prognostic Markers | Effect | References |
|---|---|---|---|---|
| clinical trial (n = 42) | ↑ RhoA, RhoC | higher tumor stages, metastasis spreading | [201] | |
| clinical trial (n = 117) | ↓ RacGAP1 | lower risk of recurrence, better overall survival | [203] | |
| clinical trial (n = 85) | ↓ ARHGAP26 | lower overall survival, poor prognosis | [204] | |
| clinical trial (n = 75) | ↓ ARHGAP10 | lower overall survival | [205] | |
| clinical trial (n = 150) | ↑ Rac1 | poor prognosis, risk of recurrence | [206] | |
| clinical trial (n = 330) | ↑ Tiam1 | association with clinical stage, histological grade, lower overall survival | [207] | |
| in vitro (OVCAR3) clinical study (n = 80) | ↑ SMURF1 | lower overall survival | [208] | |
| in vivo (BALB/C nude mice) | ↑ miR-106b | suppression of tumor development and progression | [213] | |
| OC | in vitro (OVCAR3, SKOV3/DDP, HO8910-PM) in vivo (BALB/C nude mice) | ↑ miR-93-5p | suppression of tumorigenesis and cancer progression | [214] |
| in vitro (JHOC-5, JHOC-7, JHOC8, JHOS-2, JHOS-3, JHOS-4, JHOM-1, OVCAR3) | ↑ miR-10b | acquisition of metastatic phenotypes | [215] | |
| clinical trial (n = 74) | ↓ miR-145 | poor prognosis | [216] | |
| in vitro (SKOV3, HO8910, ES-2, CAOVR3, OVCAR3, A2780 and A2780PTX) | ↑ miR-519d | reversion of oncogenic effect of E2F1 | [217] | |
| in vitro (OVCAR3) | ↑ miR-208a-5p | suppression of cell migration and invasion | [219] | |
| clinical trial (n = 46) | ↓ miR-139-5p | correlation with FIGO stage, lymph node metastasis, poor overall survival | [218] | |
| CC | clinical trial (n = 195) | ↑ Cdc42 | association with clinical stage of tumors | [134] |
| clinical trial (n = 49) | ↑ RhoA | prediction of distant metastasis after chemotherapy | [209] | |
| clinical trial (n = 80) | ↑ RhoA | vascular invasion and metastasis | [132] | |
| clinical trial (n = 298) | ↑ Tiam1 | advanced clinical stage, metastasis spreading, HPV infection, poor overall survival | [210] | |
| in vitro (SiHa, HeLa) | ↑ miR-217 | reduction colony formation, invasion, increased apoptosis | [223] | |
| in vitro (HeLa) | ↑ miR-200b | inhibition of EMT | [222] | |
| in vitro (C33) | ↑ miR-143-3p | positive regulation of cancer progression | ||
| EC | clinical trial (n = 364) | ↑ MIIP | advanced clinical stage, lymph node metastasis | [94] |
| clinical trial (n = 265) | ↑ Vav3 | no significant with clinic pathological features and prognostic outcomes for patients | [212] | |
| in vitro (HEC-1B) in vivo (BALB/C nude mice) | ↑ miR-372 | suppression of tumorigenesis | [220] | |
| in vitro (HEC50, AN3CA) | ↑ miR-200c | inhibition of cell motility and anoikis resistance | [221] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zubor, P.; Dankova, Z.; Kolkova, Z.; Holubekova, V.; Brany, D.; Mersakova, S.; Samec, M.; Liskova, A.; Koklesova, L.; Kubatka, P.; et al. Rho GTPases in Gynecologic Cancers: In-Depth Analysis toward the Paradigm Change from Reactive to Predictive, Preventive, and Personalized Medical Approach Benefiting the Patient and Healthcare. Cancers 2020, 12, 1292. https://doi.org/10.3390/cancers12051292
Zubor P, Dankova Z, Kolkova Z, Holubekova V, Brany D, Mersakova S, Samec M, Liskova A, Koklesova L, Kubatka P, et al. Rho GTPases in Gynecologic Cancers: In-Depth Analysis toward the Paradigm Change from Reactive to Predictive, Preventive, and Personalized Medical Approach Benefiting the Patient and Healthcare. Cancers. 2020; 12(5):1292. https://doi.org/10.3390/cancers12051292
Chicago/Turabian StyleZubor, Pavol, Zuzana Dankova, Zuzana Kolkova, Veronika Holubekova, Dusan Brany, Sandra Mersakova, Marek Samec, Alena Liskova, Lenka Koklesova, Peter Kubatka, and et al. 2020. "Rho GTPases in Gynecologic Cancers: In-Depth Analysis toward the Paradigm Change from Reactive to Predictive, Preventive, and Personalized Medical Approach Benefiting the Patient and Healthcare" Cancers 12, no. 5: 1292. https://doi.org/10.3390/cancers12051292
APA StyleZubor, P., Dankova, Z., Kolkova, Z., Holubekova, V., Brany, D., Mersakova, S., Samec, M., Liskova, A., Koklesova, L., Kubatka, P., Bujnak, J., Kajo, K., Mlyncek, M., Giordano, F. A., & Golubnitschaja, O. (2020). Rho GTPases in Gynecologic Cancers: In-Depth Analysis toward the Paradigm Change from Reactive to Predictive, Preventive, and Personalized Medical Approach Benefiting the Patient and Healthcare. Cancers, 12(5), 1292. https://doi.org/10.3390/cancers12051292