BRCA1 Promoter Hypermethylation is Associated with Good Prognosis and Chemosensitivity in Triple-Negative Breast Cancer

 ,
, 

 and
and
Abstract
1. Introduction
2. Results
2.1. Patient and Tumor Characteristics
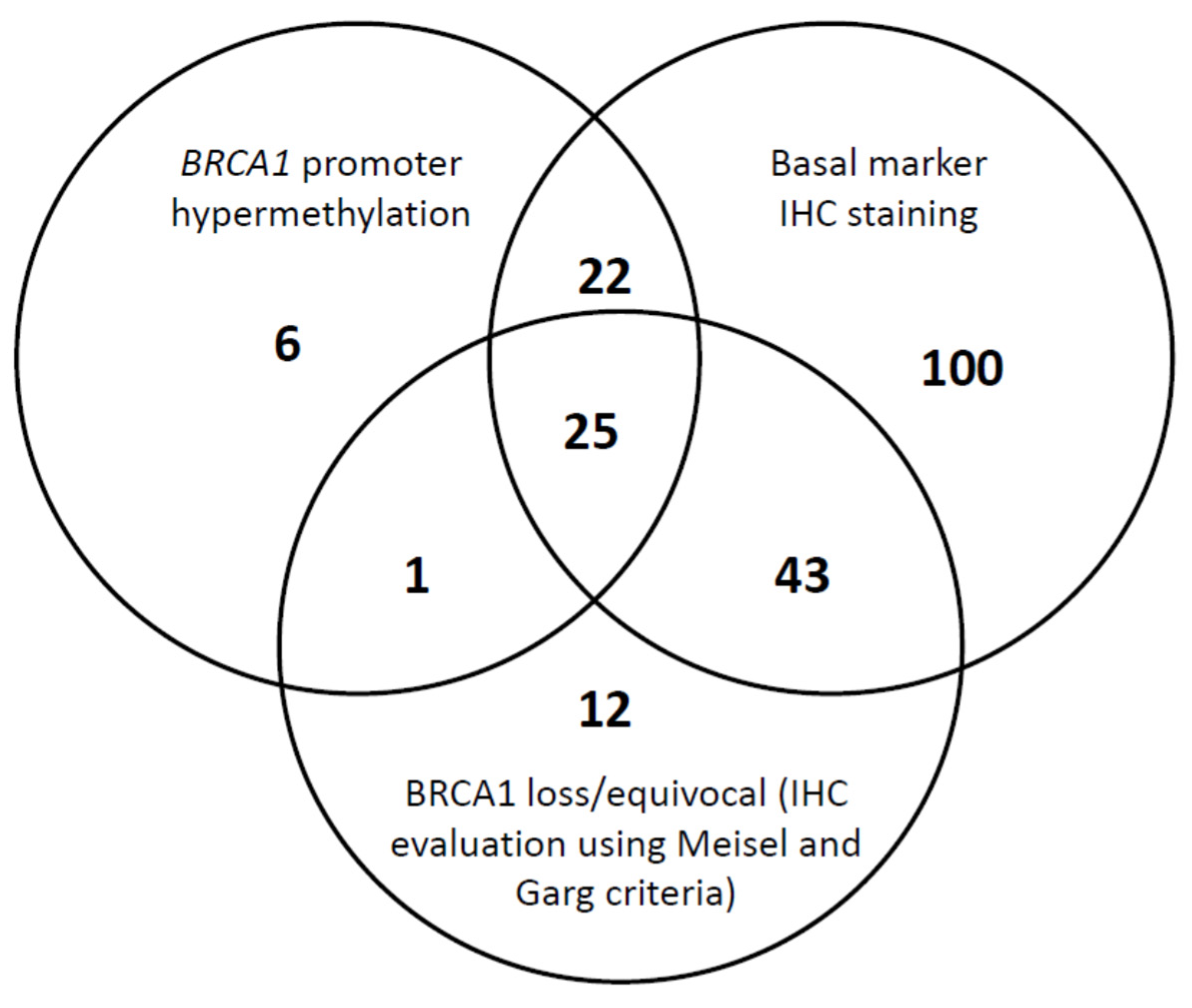
2.2. In Situ BRCA1 IHC Analysis
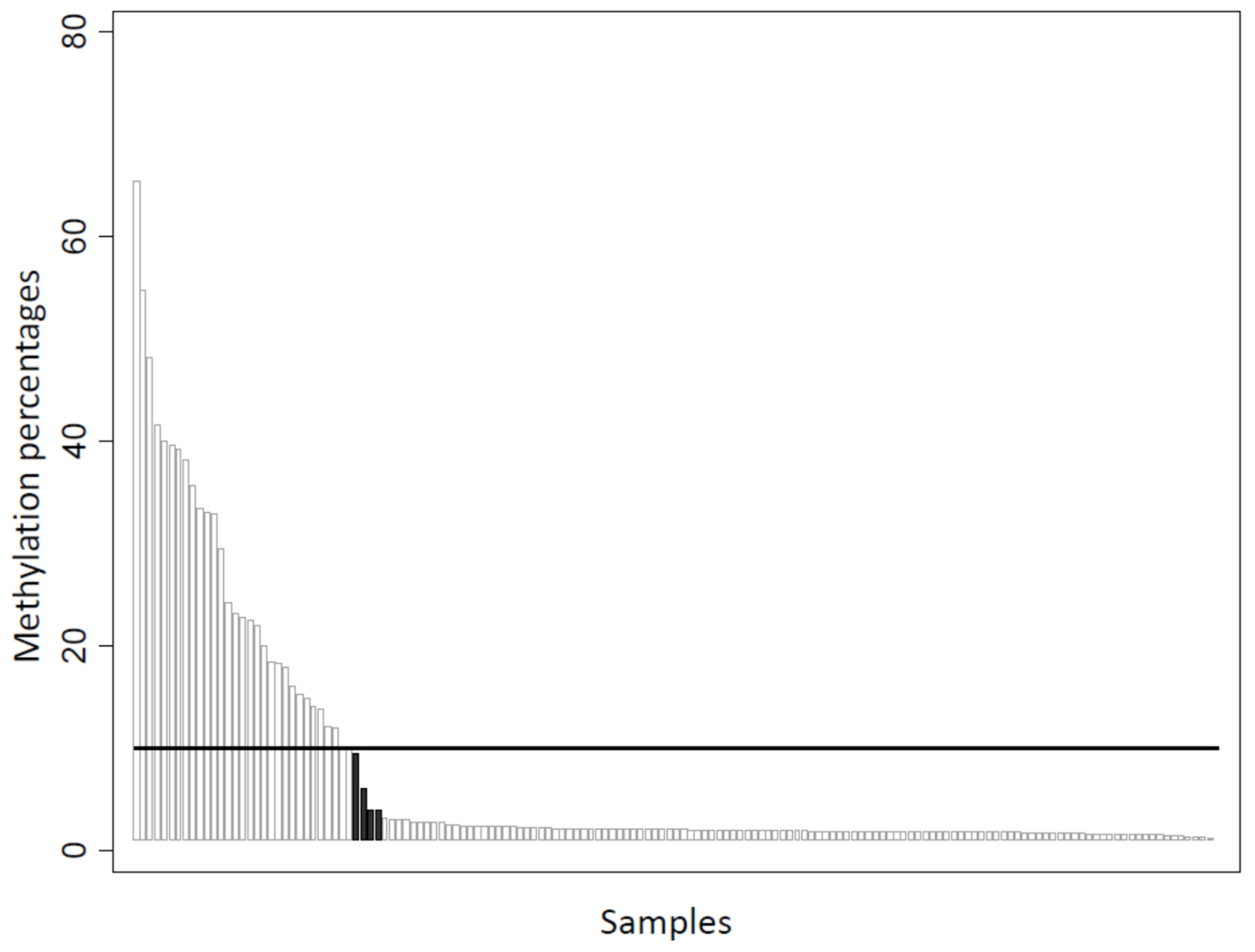
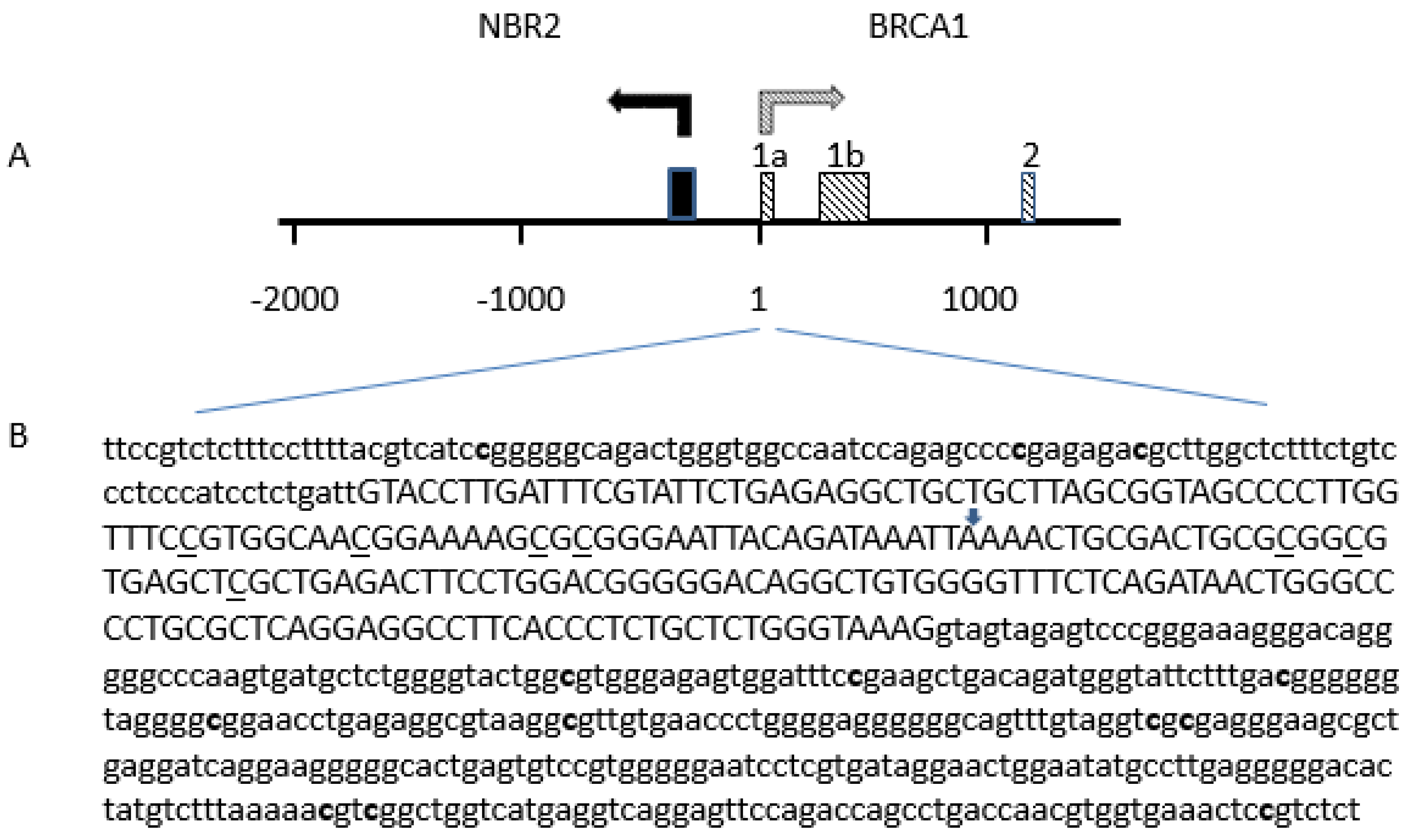
2.3. Detection of BRCA1 Promoter Hypermethylation
2.4. BRCA1 Expression, BRCA1 Promoter Hypermethylation, and Clinicopathological Associations
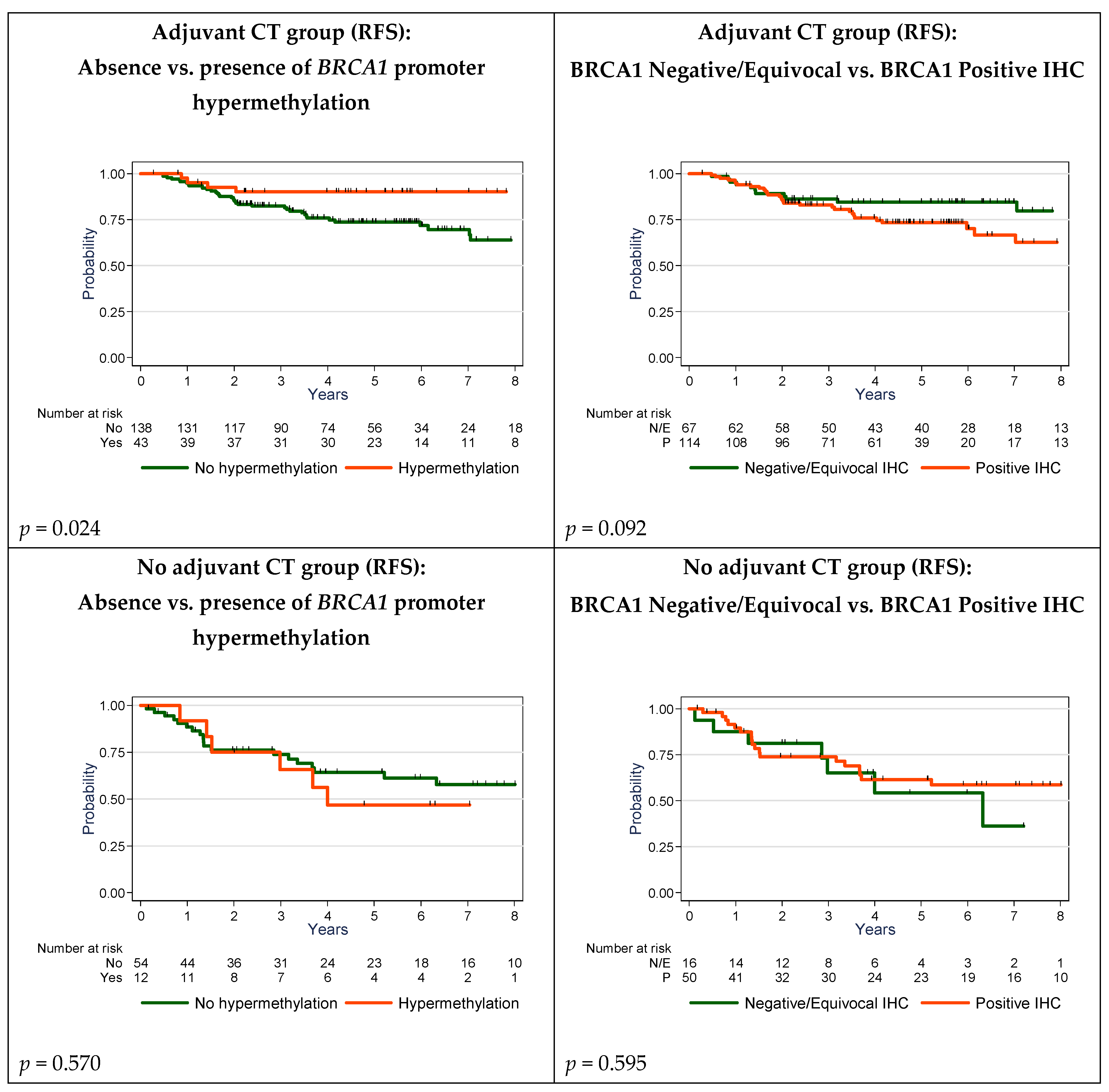
2.5. Survival Analyses
3. Discussion
4. Materials and Methods
4.1. Objectives
4.2. Patients and Tumor Samples
4.3. Tissue Microarray and Immunohistochemistry
4.4. TMA Analysis/Interpretation
4.5. Tissue Processing and DNA Extraction
4.6. BRCA1 Promoter Methylation Status
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BRCA1 | Breast cancer type 1 susceptibility protein |
| CK 5/6 | Cytokeratin 5/6 |
| CT | Chemotherapy |
| DNA | Deoxyribonucleic acid |
| EGFR | Epidermal growth factor receptor |
| ER | Estrogen receptor |
| HER2 | Human epidermal growth factor receptor 2 |
| IHC | Immunohistochemistry |
| MS-PCR | Methylation-specific polymerase chain reaction |
| OS | Overall survival |
| PARP-1 | Poly(ADP-ribose) polymerase 1 |
| PARPi | PARP inhibitor |
| PCR | Polymerase chain reaction |
| PR | Progesterone receptor |
| RFS | Relapse-free survival |
| RNA | Ribonucleic acid |
| TMA | Tissue microarray |
| TNBC | Triple-negative breast cancer |
References
- Dent, R.; Trudeau, M.; Pritchard, K.I.; Hanna, W.M.; Kahn, H.K.; Sawka, C.A.; Lickley, L.A.; Rawlinson, E.; Sun, P.; Narod, S.A. Triple-negative breast cancer: Clinical features and patterns of recurrence. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2007, 13, 4429–4434. [Google Scholar] [CrossRef] [PubMed]
- Elias, A.D. Triple-negative breast cancer: A short review. Am. J. Clin. Oncol. 2010, 33, 637–645. [Google Scholar] [CrossRef] [PubMed]
- Carey, L.A.; Dees, E.C.; Sawyer, L.; Gatti, L.; Moore, D.T.; Collichio, F.; Ollila, D.W.; Sartor, C.I.; Graham, M.L.; Perou, C.M. The triple negative paradox: Primary tumor chemosensitivity of breast cancer subtypes. Clin. Cancer Res. 2007, 13, 2329–2334. [Google Scholar] [CrossRef] [PubMed]
- Perou, C.M.; Sorlie, T.; Eisen, M.B.; van de Rijn, M.; Jeffrey, S.S.; Rees, C.A.; Pollack, J.R.; Ross, D.T.; Johnsen, H.; Akslen, L.A.; et al. Molecular portraits of human breast tumours. Nature 2000, 406, 747–752. [Google Scholar] [CrossRef]
- Sorlie, T.; Perou, C.M.; Tibshirani, R.; Aas, T.; Geisler, S.; Johnsen, H.; Hastie, T.; Eisen, M.B.; van de Rijn, M.; Jeffrey, S.S.; et al. Gene expression patterns of breast carcinomas distinguish tumor subclasses with clinical implications. Proc. Natl. Acad. Sci. USA 2001, 98, 10869–10874. [Google Scholar] [CrossRef]
- Prat, A.; Adamo, B.; Cheang, M.C.; Anders, C.K.; Carey, L.A.; Perou, C.M. Molecular characterization of basal-like and non-basal-like triple-negative breast cancer. Oncologist 2013, 18, 123–133. [Google Scholar] [CrossRef]
- Cheang, M.C.; Voduc, D.; Bajdik, C.; Leung, S.; McKinney, S.; Chia, S.K.; Perou, C.M.; Nielsen, T.O. Basal-like breast cancer defined by five biomarkers has superior prognostic value than triple-negative phenotype. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2008, 14, 1368–1376. [Google Scholar] [CrossRef]
- Lakhani, S.R.; Van De Vijver, M.J.; Jacquemier, J.; Anderson, T.J.; Osin, P.P.; McGuffog, L.; Easton, D.F. The pathology of familial breast cancer: Predictive value of immunohistochemical markers estrogen receptor, progesterone receptor, HER-2, and p53 in patients with mutations in BRCA1 and BRCA2. J. Clin. Oncol. 2002, 20, 2310–2318. [Google Scholar] [CrossRef]
- Stevens, K.N.; Vachon, C.M.; Couch, F.J. Genetic susceptibility to triple-negative breast cancer. Cancer Res. 2013, 73, 2025–2030. [Google Scholar] [CrossRef]
- Song, Y.; Barry, W.T.; Seah, D.S.; Tung, N.M.; Garber, J.E.; Lin, N.U. Patterns of recurrence and metastasis in BRCA1/BRCA2-associated breast cancers. Cancer 2020, 126, 271–280. [Google Scholar] [CrossRef]
- Turner, N.; Tutt, A.; Ashworth, A. Hallmarks of ‘BRCAness’ in sporadic cancers. Nat. Rev. Cancer 2004, 4, 814–819. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Palma, J.; Kinders, R.; Shi, Y.; Donawho, C.; Ellis, P.A.; Rodriguez, L.E.; Colon-Lopez, M.; Saltarelli, M.; LeBlond, D.; et al. An enzyme-linked immunosorbent poly (ADP-ribose) polymerase biomarker assay for clinical trials of PARP inhibitors. Anal. Biochem. 2008, 381, 240–247. [Google Scholar] [CrossRef] [PubMed]
- Helleday, T.; Petermann, E.; Lundin, C.; Hodgson, B.; Sharma, R.A. DNA repair pathways as targets for cancer therapy. Nat. Rev. Cancer 2008, 8, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Tutt, A.; Robson, M.; Garber, J.E.; Domchek, S.M.; Audeh, M.W.; Weitzel, J.N.; Friedlander, M.; Arun, B.; Loman, N.; Schmutzler, R.K.; et al. Oral poly(ADP-ribose) polymerase inhibitor olaparib in patients with BRCA1 or BRCA2 mutations and advanced breast cancer: A proof-of-concept trial. Lancet 2010, 376, 235–244. [Google Scholar] [CrossRef]
- Robson, M.; Im, S.A.; Senkus, E.; Xu, B.; Domchek, S.M.; Masuda, N.; Delaloge, S.; Li, W.; Tung, N.; Armstrong, A.; et al. Olaparib for Metastatic Breast Cancer in Patients with a Germline BRCA Mutation. N. Engl. J. Med. 2017, 377, 523–533. [Google Scholar] [CrossRef]
- Litton, J.K.; Rugo, H.S.; Ettl, J.; Hurvitz, S.A.; Goncalves, A.; Lee, K.H.; Fehrenbacher, L.; Yerushalmi, R.; Mina, L.A.; Martin, M.; et al. Talazoparib in Patients with Advanced Breast Cancer and a Germline BRCA Mutation. N. Engl. J. Med. 2018, 379, 753–763. [Google Scholar] [CrossRef]
- Rakha, E.A.; El-Sheikh, S.E.; Kandil, M.A.; El-Sayed, M.E.; Green, A.R.; Ellis, I.O. Expression of BRCA1 protein in breast cancer and its prognostic significance. Hum. Pathol. 2008, 39, 857–865. [Google Scholar] [CrossRef]
- Bal, A.; Verma, S.; Joshi, K.; Singla, A.; Thakur, R.; Arora, S.; Singh, G. BRCA1-methylated sporadic breast cancers are BRCA-like in showing a basal phenotype and absence of ER expression. Virchows Arch. 2012, 461, 305–312. [Google Scholar] [CrossRef]
- Birgisdottir, V.; Stefansson, O.A.; Bodvarsdottir, S.K.; Hilmarsdottir, H.; Jonasson, J.G.; Eyfjord, J.E. Epigenetic silencing and deletion of the BRCA1 gene in sporadic breast cancer. Breast Cancer Res. BCR 2006, 8, R38. [Google Scholar] [CrossRef]
- Lips, E.H.; Mulder, L.; Hannemann, J.; Laddach, N.; Vrancken Peeters, M.T.; van de Vijver, M.J.; Wesseling, J.; Nederlof, P.M.; Rodenhuis, S. Indicators of homologous recombination deficiency in breast cancer and association with response to neoadjuvant chemotherapy. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. ESMO 2011, 22, 870–876. [Google Scholar] [CrossRef]
- Mirza, S.; Sharma, G.; Prasad, C.P.; Parshad, R.; Srivastava, A.; Gupta, S.D.; Ralhan, R. Promoter hypermethylation of TMS1, BRCA1, ERalpha and PRB in serum and tumor DNA of invasive ductal breast carcinoma patients. Life Sci. 2007, 81, 280–287. [Google Scholar] [CrossRef] [PubMed]
- Wei, M.; Grushko, T.A.; Dignam, J.; Hagos, F.; Nanda, R.; Sveen, L.; Xu, J.; Fackenthal, J.; Tretiakova, M.; Das, S.; et al. BRCA1 promoter methylation in sporadic breast cancer is associated with reduced BRCA1 copy number and chromosome 17 aneusomy. Cancer Res. 2005, 65, 10692–10699. [Google Scholar] [CrossRef] [PubMed]
- Lips, E.H.; Mulder, L.; Oonk, A.; van der Kolk, L.E.; Hogervorst, F.B.; Imholz, A.L.; Wesseling, J.; Rodenhuis, S.; Nederlof, P.M. Triple-negative breast cancer: BRCAness and concordance of clinical features with BRCA1-mutation carriers. Br. J. Cancer 2013, 108, 2172–2177. [Google Scholar] [CrossRef] [PubMed]
- Birkbak, N.J.; Wang, Z.C.; Kim, J.Y.; Eklund, A.C.; Li, Q.; Tian, R.; Bowman-Colin, C.; Li, Y.; Greene-Colozzi, A.; Iglehart, J.D.; et al. Telomeric allelic imbalance indicates defective DNA repair and sensitivity to DNA-damaging agents. Cancer Discov. 2012, 2, 366–375. [Google Scholar] [CrossRef]
- Jacot, W.; Thezenas, S.; Senal, R.; Viglianti, C.; Laberenne, A.C.; Lopez-Crapez, E.; Bibeau, F.; Bleuse, J.P.; Romieu, G.; Lamy, P.J. BRCA1 promoter hypermethylation, 53BP1 protein expression and PARP-1 activity as biomarkers of DNA repair deficit in breast cancer. BMC Cancer 2013, 13, 523. [Google Scholar] [CrossRef]
- Zhang, L.; Long, X. Association of BRCA1 promoter methylation with sporadic breast cancers: Evidence from 40 studies. Sci. Rep. 2015, 5, 17869. [Google Scholar] [CrossRef]
- Yamashita, N.; Tokunaga, E.; Kitao, H.; Hitchins, M.; Inoue, Y.; Tanaka, K.; Hisamatsu, Y.; Taketani, K.; Akiyoshi, S.; Okada, S.; et al. Epigenetic Inactivation of BRCA1 Through Promoter Hypermethylation and Its Clinical Importance in Triple-Negative Breast Cancer. Clin. Breast Cancer 2015, 15, 498–504. [Google Scholar] [CrossRef]
- Zhu, X.; Shan, L.; Wang, F.; Wang, J.; Wang, F.; Shen, G.; Liu, X.; Wang, B.; Yuan, Y.; Ying, J.; et al. Hypermethylation of BRCA1 gene: Implication for prognostic biomarker and therapeutic target in sporadic primary triple-negative breast cancer. Breast Cancer Res. Treat. 2015, 150, 479–486. [Google Scholar] [CrossRef]
- Cai, F.F.; Chen, S.; Wang, M.H.; Lin, X.Y.; Zhang, L.; Zhang, J.X.; Wang, L.X.; Yang, J.; Ding, J.H.; Pan, X.; et al. Pyrosequencing quantified methylation level of BRCA1 promoter as prognostic factor for survival in breast cancer patient. Oncotarget 2016, 7, 27499–27510. [Google Scholar] [CrossRef]
- Sun, T.; Ruscito, I.; Dimitrova, D.; Chekerov, R.; Kulbe, H.; Baron, U.; Blanchard, V.; Panici, P.B.; Darb-Esfahani, S.; Sehouli, J.; et al. Genetic Versus Epigenetic BRCA1 Silencing Pathways: Clinical Effects in Primary Ovarian Cancer Patients: A Study of the Tumor Bank Ovarian Cancer Consortium. Int. J. Gynecol. Cancer 2017. [Google Scholar] [CrossRef]
- Cancer Genome Atlas Research, N. Integrated genomic analyses of ovarian carcinoma. Nature 2011, 474, 609–615. [Google Scholar] [CrossRef] [PubMed]
- Esteller, M.; Silva, J.M.; Dominguez, G.; Bonilla, F.; Matias-Guiu, X.; Lerma, E.; Bussaglia, E.; Prat, J.; Harkes, I.C.; Repasky, E.A.; et al. Promoter hypermethylation and BRCA1 inactivation in sporadic breast and ovarian tumors. J. Natl. Cancer Inst. 2000, 92, 564–569. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Khan, S.; Sun, Y.; Hess, K.; Shmulevich, I.; Sood, A.K.; Zhang, W. Association of BRCA1 and BRCA2 mutations with survival, chemotherapy sensitivity, and gene mutator phenotype in patients with ovarian cancer. JAMA 2011, 306, 1557–1565. [Google Scholar] [CrossRef] [PubMed]
- Bol, G.M.; Suijkerbuijk, K.P.; Bart, J.; Vooijs, M.; van der Wall, E.; van Diest, P.J. Methylation profiles of hereditary and sporadic ovarian cancer. Histopathology 2010, 57, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Evans, D.G.R.; van Veen, E.M.; Byers, H.J.; Wallace, A.J.; Ellingford, J.M.; Beaman, G.; Santoyo-Lopez, J.; Aitman, T.J.; Eccles, D.M.; Lalloo, F.I.; et al. A Dominantly Inherited 5’ UTR Variant Causing Methylation-Associated Silencing of BRCA1 as a Cause of Breast and Ovarian Cancer. Am. J. Hum. Genet. 2018, 103, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Collins, N.; Wooster, R.; Stratton, M.R. Absence of methylation of CpG dinucleotides within the promoter of the breast cancer susceptibility gene BRCA2 in normal tissues and in breast and ovarian cancers. Br. J. Cancer 1997, 76, 1150–1156. [Google Scholar] [CrossRef] [PubMed]
- Hughes-Davies, L.; Huntsman, D.; Ruas, M.; Fuks, F.; Bye, J.; Chin, S.F.; Milner, J.; Brown, L.A.; Hsu, F.; Gilks, B.; et al. EMSY links the BRCA2 pathway to sporadic breast and ovarian cancer. Cell 2003, 115, 523–535. [Google Scholar] [CrossRef]
- Gelmon, K.A.; Tischkowitz, M.; Mackay, H.; Swenerton, K.; Robidoux, A.; Tonkin, K.; Hirte, H.; Huntsman, D.; Clemons, M.; Gilks, B.; et al. Olaparib in patients with recurrent high-grade serous or poorly differentiated ovarian carcinoma or triple-negative breast cancer: A phase 2, multicentre, open-label, non-randomised study. Lancet Oncol. 2011, 12, 852–861. [Google Scholar] [CrossRef]
- Drew, Y.; Mulligan, E.A.; Vong, W.T.; Thomas, H.D.; Kahn, S.; Kyle, S.; Mukhopadhyay, A.; Los, G.; Hostomsky, Z.; Plummer, E.R.; et al. Therapeutic potential of poly(ADP-ribose) polymerase inhibitor AG014699 in human cancers with mutated or methylated BRCA1 or BRCA2. J. Natl. Cancer Inst. 2011, 103, 334–346. [Google Scholar] [CrossRef]
- Veeck, J.; Ropero, S.; Setien, F.; Gonzalez-Suarez, E.; Osorio, A.; Benitez, J.; Herman, J.G.; Esteller, M. BRCA1 CpG island hypermethylation predicts sensitivity to poly(adenosine diphosphate)-ribose polymerase inhibitors. J. Clin. Oncol. 2010, 28, e563–e564. [Google Scholar] [CrossRef]
- Milner, R.; Wombwell, H.; Eckersley, S.; Barnes, D.; Warwicker, J.; Van Dorp, E.; Rowlinson, R.; Dearden, S.; Hughes, G.; Harbron, C.; et al. Validation of the BRCA1 antibody MS110 and the utility of BRCA1 as a patient selection biomarker in immunohistochemical analysis of breast and ovarian tumours. Virchows Arch. 2013, 462, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Al-Mulla, F.; Abdulrahman, M.; Varadharaj, G.; Akhter, N.; Anim, J.T. BRCA1 gene expression in breast cancer: A correlative study between real-time RT-PCR and immunohistochemistry. J. Histochem. Cytochem. 2005, 53, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Meisel, J.L.; Hyman, D.M.; Garg, K.; Zhou, Q.; Dao, F.; Bisogna, M.; Gao, J.; Schultz, N.D.; Grisham, R.N.; Phillips, M.; et al. The performance of BRCA1 immunohistochemistry for detecting germline, somatic, and epigenetic BRCA1 loss in high-grade serous ovarian cancer. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. ESMO 2014, 25, 2372–2378. [Google Scholar] [CrossRef] [PubMed]
- Korlimarla, A.; Prabhu, J.S.; Remacle, J.; Rajarajan, S.; Raja, U.; Ce, A.; Srinath, B.S.; Manjunath, S.; Ks, G.; Correa, M.; et al. Identification of BRCA1 Deficiency Using Multi-Analyte Estimation of BRCA1 and Its Repressors in FFPE Tumor Samples from Patients with Triple Negative Breast Cancer. PLoS ONE 2016, 11, e0153113. [Google Scholar] [CrossRef] [PubMed]
- Guo, T.; Ren, Y.; Wang, B.; Huang, Y.; Jia, S.; Tang, W.; Luo, Y. Promoter methylation of BRCA1 is associated with estrogen, progesterone and human epidermal growth factor receptor-negative tumors and the prognosis of breast cancer: A meta-analysis. Mol. Clin. Oncol. 2015, 3, 1353–1360. [Google Scholar] [CrossRef] [PubMed]
- Schmid, P.; Chui, S.Y.; Emens, L.A. Atezolizumab and Nab-Paclitaxel in Advanced Triple-Negative Breast Cancer. Reply. N. Engl. J. Med. 2019, 380, 987–988. [Google Scholar] [CrossRef]
- Yarchoan, M.; Hopkins, A.; Jaffee, E.M. Tumor Mutational Burden and Response Rate to PD-1 Inhibition. N. Engl. J. Med. 2017, 377, 2500–2501. [Google Scholar] [CrossRef]
- Mittendorf, E.A.; Philips, A.V.; Meric-Bernstam, F.; Qiao, N.; Wu, Y.; Harrington, S.; Su, X.; Wang, Y.; Gonzalez-Angulo, A.M.; Akcakanat, A.; et al. PD-L1 Expression in Triple-Negative Breast Cancer. Cancer Immunol. Res. 2014, 2, 361–370. [Google Scholar] [CrossRef]
- Garg, K.; Levine, D.A.; Olvera, N.; Dao, F.; Bisogna, M.; Secord, A.A.; Berchuck, A.; Cerami, E.; Schultz, N.; Soslow, R.A. BRCA1 immunohistochemistry in a molecularly characterized cohort of ovarian high-grade serous carcinomas. Am. J. Surg. Pathol. 2013, 37, 138–146. [Google Scholar] [CrossRef]
- Cho, E.Y.; Chang, M.H.; Choi, Y.L.; Lee, J.E.; Nam, S.J.; Yang, J.H.; Park, Y.H.; Ahn, J.S.; Im, Y.H. Potential candidate biomarkers for heterogeneity in triple-negative breast cancer (TNBC). Cancer Chemother. Pharm. 2011, 68, 753–761. [Google Scholar] [CrossRef]
- Byrski, T.; Dent, R.; Blecharz, P.; Foszczynska-Kloda, M.; Gronwald, J.; Huzarski, T.; Cybulski, C.; Marczyk, E.; Chrzan, R.; Eisen, A.; et al. Results of a phase II open-label, non-randomized trial of cisplatin chemotherapy in patients with BRCA1-positive metastatic breast cancer. Breast Cancer Res. BCR 2012, 14, R110. [Google Scholar] [CrossRef] [PubMed]
- Perez-Valles, A.; Martorell-Cebollada, M.; Nogueira-Vazquez, E.; Garcia-Garcia, J.A.; Fuster-Diana, E. The usefulness of antibodies to the BRCA1 protein in detecting the mutated BRCA1 gene. An immunohistochemical study. J. Clin. Pathol. 2001, 54, 476–480. [Google Scholar] [CrossRef] [PubMed]
- Vorrius, T.R.; Snyder, K.; Pica-Mendez, A.; Tan, C.; Laterza, O.; Toniatti, C.; Carpenter, C.; Lee, H.; Tanaka, W.; Zhang, Z.-Q. Immunohistochemical Detection of BRCA-1 and BRCA-2 Expression in Human Breast and Ovarian Tumors. J. Histotechnol. 2009, 32, 202–203. [Google Scholar] [CrossRef]
- Xu, C.F.; Chambers, J.A.; Solomon, E. Complex regulation of the BRCA1 gene. J. Biol. Chem. 1997, 272, 20994–20997. [Google Scholar] [CrossRef] [PubMed]
- Rice, J.C.; Ozcelik, H.; Maxeiner, P.; Andrulis, I.; Futscher, B.W. Methylation of the BRCA1 promoter is associated with decreased BRCA1 mRNA levels in clinical breast cancer specimens. Carcinogenesis 2000, 21, 1761–1765. [Google Scholar] [CrossRef]
- Bianco, T.; Chenevix-Trench, G.; Walsh, D.C.; Cooper, J.E.; Dobrovic, A. Tumour-specific distribution of BRCA1 promoter region methylation supports a pathogenetic role in breast and ovarian cancer. Carcinogenesis 2000, 21, 147–151. [Google Scholar] [CrossRef]
- Kondrashova, O.; Topp, M.; Nesic, K.; Lieschke, E.; Ho, G.Y.; Harrell, M.I.; Zapparoli, G.V.; Hadley, A.; Holian, R.; Boehm, E.; et al. Methylation of all BRCA1 copies predicts response to the PARP inhibitor rucaparib in ovarian carcinoma. Nat. Commun. 2018, 9, 3970. [Google Scholar] [CrossRef]
- Ignatov, T.; Poehlmann, A.; Ignatov, A.; Schinlauer, A.; Costa, S.D.; Roessner, A.; Kalinski, T.; Bischoff, J. BRCA1 promoter methylation is a marker of better response to anthracycline-based therapy in sporadic TNBC. Breast Cancer Res. Treat. 2013, 141, 205–212. [Google Scholar] [CrossRef]
- Stefansson, O.A.; Villanueva, A.; Vidal, A.; Marti, L.; Esteller, M. BRCA1 epigenetic inactivation predicts sensitivity to platinum-based chemotherapy in breast and ovarian cancer. Epigenetics 2012, 7, 1225–1229. [Google Scholar] [CrossRef]
- Xu, Y.; Diao, L.; Chen, Y.; Liu, Y.; Wang, C.; Ouyang, T.; Li, J.; Wang, T.; Fan, Z.; Fan, T.; et al. Promoter methylation of BRCA1 in triple-negative breast cancer predicts sensitivity to adjuvant chemotherapy. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. ESMO 2013, 24, 1498–1505. [Google Scholar] [CrossRef]
- Tutt, A.; Tovey, H.; Cheang, M.C.U.; Kernaghan, S.; Kilburn, L.; Gazinska, P.; Owen, J.; Abraham, J.; Barrett, S.; Barrett-Lee, P.; et al. Carboplatin in BRCA1/2-mutated and triple-negative breast cancer BRCAness subgroups: The TNT Trial. Nat. Med. 2018, 24, 628–637. [Google Scholar] [CrossRef] [PubMed]
- Baker, L.A.; Holliday, H.; Swarbrick, A. ID4 controls luminal lineage commitment in normal mammary epithelium and inhibits BRCA1 function in basal-like breast cancer. Endocr. Relat. Cancer 2016, 23, R381–R392. [Google Scholar] [CrossRef] [PubMed]
- Crippa, E.; Lusa, L.; De Cecco, L.; Marchesi, E.; Calin, G.A.; Radice, P.; Manoukian, S.; Peissel, B.; Daidone, M.G.; Gariboldi, M.; et al. miR-342 regulates BRCA1 expression through modulation of ID4 in breast cancer. PLoS ONE 2014, 9, e87039. [Google Scholar] [CrossRef] [PubMed]
- Roldan, G.; Delgado, L.; Muse, I.M. Tumoral expression of BRCA1, estrogen receptor alpha and ID4 protein in patients with sporadic breast cancer. Cancer Biol. 2006, 5, 505–510. [Google Scholar] [CrossRef]
- Thike, A.A.; Tan, P.H.; Ikeda, M.; Iqbal, J. Increased ID4 expression, accompanied by mutant p53 accumulation and loss of BRCA1/2 proteins in triple-negative breast cancer, adversely affects survival. Histopathology 2016, 68, 702–712. [Google Scholar] [CrossRef]
- Wen, Y.H.; Ho, A.; Patil, S.; Akram, M.; Catalano, J.; Eaton, A.; Norton, L.; Benezra, R.; Brogi, E. Id4 protein is highly expressed in triple-negative breast carcinomas: Possible implications for BRCA1 downregulation. Breast Cancer Res. Treat. 2012, 135, 93–102. [Google Scholar] [CrossRef]
- Garcia, A.I.; Buisson, M.; Bertrand, P.; Rimokh, R.; Rouleau, E.; Lopez, B.S.; Lidereau, R.; Mikaelian, I.; Mazoyer, S. Down-regulation of BRCA1 expression by miR-146a and miR-146b-5p in triple negative sporadic breast cancers. EMBO Mol. Med. 2011, 3, 279–290. [Google Scholar] [CrossRef]
- Matamala, N.; Vargas, M.T.; Gonzalez-Campora, R.; Arias, J.I.; Menendez, P.; Andres-Leon, E.; Yanowsky, K.; Llaneza-Folgueras, A.; Minambres, R.; Martinez-Delgado, B.; et al. MicroRNA deregulation in triple negative breast cancer reveals a role of miR-498 in regulating BRCA1 expression. Oncotarget 2016, 7, 20068–20079. [Google Scholar] [CrossRef]
- Tan, X.; Peng, J.; Fu, Y.; An, S.; Rezaei, K.; Tabbara, S.; Teal, C.B.; Man, Y.G.; Brem, R.F.; Fu, S.W. miR-638 mediated regulation of BRCA1 affects DNA repair and sensitivity to UV and cisplatin in triple-negative breast cancer. Breast Cancer Res. BCR 2014, 16, 435. [Google Scholar] [CrossRef]
- Boukerroucha, M.; Josse, C.; ElGuendi, S.; Boujemla, B.; Freres, P.; Maree, R.; Wenric, S.; Segers, K.; Collignon, J.; Jerusalem, G.; et al. Evaluation of BRCA1-related molecular features and microRNAs as prognostic factors for triple negative breast cancers. BMC Cancer 2015, 15, 755. [Google Scholar] [CrossRef]
- Sobral-Leite, M.; Van de Vijver, K.; Michaut, M.; van der Linden, R.; Hooijer, G.K.J.; Horlings, H.M.; Severson, T.M.; Mulligan, A.M.; Weerasooriya, N.; Sanders, J.; et al. Assessment of PD-L1 expression across breast cancer molecular subtypes, in relation to mutation rate, BRCA1-like status, tumor-infiltrating immune cells and survival. Oncoimmunology 2018, 7, e1509820. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Zhao, L.; Lang, J. The BRCA1 Methylation and PD-L1 Expression in Sporadic Ovarian Cancer. Int. J. Gynecol. Cancer 2018, 28, 1514–1519. [Google Scholar] [CrossRef] [PubMed]
- Lamy, P.J.; Fina, F.; Bascoul-Mollevi, C.; Laberenne, A.C.; Martin, P.M.; Ouafik, L.; Jacot, W. Quantification and clinical relevance of gene amplification at chromosome 17q12-q21 in human epidermal growth factor receptor 2-amplified breast cancers. Breast Cancer Res. BCR 2011, 13, R15. [Google Scholar] [CrossRef] [PubMed]
- Lamy, P.J.; Nanni, I.; Fina, F.; Bibeau, F.; Romain, S.; Dussert, C.; Penault Llorca, F.; Grenier, J.; Ouafik, L.H.; Martin, P.M. Reliability and discriminant validity of HER2 gene quantification and chromosome 17 aneusomy analysis by real-time PCR in primary breast cancer. Int. J. Biol. Markers 2006, 21, 20–29. [Google Scholar] [CrossRef]
- Goldhirsch, A.; Glick, J.H.; Gelber, R.D.; Coates, A.S.; Thurlimann, B.; Senn, H.J. Meeting highlights: International expert consensus on the primary therapy of early breast cancer 2005. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. ESMO 2005, 16, 1569–1583. [Google Scholar] [CrossRef]
- Jacot, W.; Gutowski, M.; Azria, D.; Romieu, G. Adjuvant early breast cancer systemic therapies according to daily used technologies. Crit. Rev. Oncol. Hematol. 2011. [Google Scholar] [CrossRef]
- Guiu, S.; Mollevi, C.; Charon-Barra, C.; Boissiere, F.; Crapez, E.; Chartron, E.; Lamy, P.J.; Gutowski, M.; Bourgier, C.; Romieu, G.; et al. Prognostic value of androgen receptor and FOXA1 co-expression in non-metastatic triple negative breast cancer and correlation with other biomarkers. Br. J. Cancer 2018, 119, 76–79. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| n = 248 | % | ||
|---|---|---|---|
| Age (years) | |||
| Median (range) | 57.8 | 28.5–98.6 | |
| Tumor Size | |||
| T1 | 111 | 44.8 | |
| T2 | 117 | 47.2 | |
| T3/T4 | 20 | 8.0 | |
| Nodal Status | |||
| N- | 162 | 65.3 | |
| N+ | 86 | 34.7 | |
| Tumor Grade | |||
| 1–2 | 59 | 24.2 | |
| 3 | 185 | 75.8 | |
| Missing | 4 | ||
| Histology | |||
| Ductal | 204 | 82.9 | |
| Lobular | 12 | 4.9 | |
| Others | 30 | 12.2 | |
| Missing | 2 | ||
| Basal-like Phetotype | |||
| Nul | 55 | 22.5 | |
| Basal-like | 190 | 77.5 | |
| Missing | 3 | ||
| BRCA1 IHC Expression | |||
| BRCA1-/ | 70 | 28.2 | |
| BRCA1+ | 165 | 66.5 | |
| Equivocal | 13 | 5.2 | |
| BRCA1 Promoter Hypermethylation | |||
| No | 193 | 77.8 | |
| Yes | 55 | 22.2 | |
| PD-L1 Expression (tumor cells) | |||
| < 1% | 100 | 43.5 | |
| ≥ 1% | 130 | 56.5 | |
| Missing | 18 | ||
| PD-L1 Expression (Immune cells) | |||
| ≤ 50% | 179 | 78.5 | |
| > 50% | 49 | 21.5 | |
| Missing | 20 | ||
| Adjuvant Chemotherapy | |||
| No | 66 | 26.7 | |
| Yes | 181 | 73.3 | |
| Missing | 1 | ||
| Relapse | |||
| No | 182 | 73.4 | |
| Yes | 66 | 26.6 | |
| Status | |||
| Alive | 175 | 70.6 | |
| Dead | 73 | 29.4 | |
| BRCA1 Promoter Methylation Status | |||||
|---|---|---|---|---|---|
| No | Yes | p | |||
| n = 193 | % | n = 55 | % | ||
| BRCA1 Methylation (MassARRAY, n = 153) | <0.001 | ||||
| Hypermethylated | 0 | 0 | 31 | 88.6 | |
| Unmethylated | 118 | 100 | 4 | 11.4 | |
| BRCA1 IHC Expression | <0.001 | ||||
| BRCA1- | 52 | 26.9 | 18 | 32.7 | |
| BRCA1+ | 137 | 71.0 | 28 | 50.9 | |
| Equivocal | 4 | 2.1 | 9 | 16.4 | |
| 0.005 | |||||
| BRCA1-/ Equivocal | 56 | 29.0 | 27 | 49.1 | |
| BRCA1+ | 137 | 71.0 | 28 | 50.9 | |
| Basal-Like Phetotype | 0.058 | ||||
| Nul | 48 | 25.1 | 7 | 13.0 | |
| Basal-like | 143 | 74.9 | 47 | 87.0 | |
| PD-L1 Expression (Tumor Cells) | 0.030 | ||||
| <1% | 85 | 47.2 | 15 | 30.0 | |
| ≥1% | 95 | 52.8 | 35 | 70.0 | |
| PD-L1 Expression (Immune Cells) | 0.011 | ||||
| 0/1/2 | 147 | 82.1 | 32 | 65.3 | |
| 3 | 32 | 17.9 | 17 | 34.7 | |
| (n = 248) | Relapse-Free Survival (RFS) | Oveall Survival (OS) | |||||
|---|---|---|---|---|---|---|---|
| N | Events | 5-Year RFS | Hazard Ratio [95% CI] | Events | 5-Year OS | Hazard Ratio [95% CI] | |
| Age (Years) | |||||||
| <55 | 108 | 23 | 79.6 | 1 | 22 | 87.4 | 1 |
| ≥55 | 140 | 43 | 68.0 | 1.52 [0.91; 2.52] | 51 | 75.4 | 2.00 [1.21; 3.30] |
| p = 0.103 | p = 0.005 | ||||||
| Tumor Size | |||||||
| pT1 | 111 | 16 | 85.3 | 1 | 17 | 90.6 | 1 |
| pT2 | 117 | 39 | 66.9 | 2.46 [1.38; 4.41] | 43 | 75.7 | 2.60 [1.48; 4.56] |
| pT3/pT4 | 20 | 11 | 44.9 | 4.88 [2.26; 10.5] | 13 | 54.5 | 6.02 [2.91; 12.4] |
| p < 0.001 | p < 0.001 | ||||||
| Nodal Status | |||||||
| pN- | 162 | 24 | 85.8 | 1 | 35 | 88.9 | 1 |
| pN+ | 86 | 42 | 50.9 | 3.94 [2.39; 6.52] | 38 | 65.2 | 2.45 [1.55; 3.88] |
| p < 0.001 | p < 0.001 | ||||||
| Histological Grade (SBR) | |||||||
| 1–2 | 59 | 17 | 76.5 | 1 | 20 | 84.3 | 1 |
| 3 | 185 | 49 | 71.4 | 1.05 [0.60; 1.82] | 52 | 78.9 | 1.05 [0.62; 1.76] |
| p = 0.875 | p = 0.863 | ||||||
| Histology | |||||||
| Ductal | 204 | 56 | 71.9 | 1 | 65 | 77.8 | 1 |
| Lobular | 12 | 6 | 54.7 | 1.94 [0.83; 4.50] | 4 | 83.3 | 0.99 [0.36; 2.73] |
| Other | 30 | 3 | 89.2 | 0.30 [0.09; 0.97] | 4 | 96.7 | 0.33 [0.12; 0.92] |
| p = 0.021 | p = 0.083 | ||||||
| Adjuvant Chemotherapy | |||||||
| No | 66 | 25 | 60.5 | 1 | 36 | 62.9 | 1 |
| Yes | 181 | 41 | 77.7 | 0.53 [0.32; 0.87] | 37 | 86.9 | 0.34 [0.22; 0.54] |
| p = 0.011 | p < 0.001 | ||||||
| Basal-Like Phenotype | |||||||
| Nul | 55 | 16 | 71.9 | 1 | 18 | 78.8 | 1 |
| Basal-like | 190 | 48 | 73.5 | 0.89 [0.50; 1.57] | 54 | 81.2 | 0.91 [0.53; 1.55] |
| p = 0.690 | p = 0.723 | ||||||
| BRCA1 IHC Expression | |||||||
| BRCA1−/Equivocal | 83 | 18 | 79.2 | 1 | 21 | 82.8 | 1 |
| BRCA1+ | 165 | 48 | 69.7 | 1.47 [0.85; 2.53] | 52 | 79.4 | 1.39 [0.84; 2.31] |
| p = 0.162 | p = 0.199 | ||||||
| BRCA1 Promoter Hypermethylation | |||||||
| No | 193 | 56 | 71.0 | 1 | 60 | 79.4 | 1 |
| Yes | 55 | 10 | 79.9 | 0.58 [0.30; 1.14] | 13 | 84.5 | 0.74 [0.40; 1.34] |
| p = 0.112 | p = 0.318 | ||||||
| PD-L1 expression (tumor cells) | |||||||
| <1% | 100 | 32 | 69.4 | 1 | 34 | 81.4 | 1 |
| ≥1% | 130 | 30 | 74.4 | 0.74 [0.45; 1.22] | 35 | 79.5 | 0.89 [0.56; 1.44] |
| p = 0.242 | p = 0.640 | ||||||
| PD-L1 expression (Immune cells) | |||||||
| 0/1/2 | 179 | 48 | 73.4 | 1 | 57 | 79.8 | 1 |
| 3 | 49 | 12 | 71.4 | 0.88 [0.47; 1.66] | 10 | 85.9 | 0.66 [0.34; 1.30] |
| p = 0.703 | p = 0.227 | ||||||
| A. Multivariate analysis: Overall population (n = 248) | ||||
| Relapse-Free Survival | Overall Survival | |||
|---|---|---|---|---|
| Hazard Ratio [95% CI] | p-Value | Hazard Ratio [95% CI] | p-Value | |
| Tumor Size | 0.003 | |||
| T1 | 1 | |||
| T2 | 2.00 [1.12; 3.59] | |||
| T3/T4 | 4.21 [1.81; 9.81] | |||
| Nodal Status | <0.001 | 0.003 | ||
| N- | 1 | 1 | ||
| N+ | 4.99 [2.94; 8.49] | 2.30 [1.35; 3.92] | ||
| Histology | 0.043 | 0.005 | ||
| Ductal | 1 | 1 | ||
| Lobular | 1.11 [0.47; 2.62] | 0.45 [0.15; 1.32] | ||
| Other | 0.30 [0.09; 0.95] | 0.27 [0.09; 0.76] | ||
| Adjuvant Chemotherapy | <0.001 | <0.001 | ||
| No | 1 | 1 | ||
| Yes | 0.39 [0.23; 0.65] | 0.30 [0.18; 0.48] | ||
| B. Multivariate Analysis: Adjuvant Chemotherapy = Yes | ||||
| (n = 181) | Relapse-Free Survival | Overall Survival | ||
| Hazard Ratio [95% CI] | p-Value | Hazard Ratio [95% CI] | p-Value | |
| Tumor size | 0.033 | |||
| T1 | 1 | |||
| T2 | 2.74 [1.22; 6.16] | |||
| T3/T4 | 2.32 [0.65; 8.24] | |||
| Nodal Status | <0.001 | 0.032 | ||
| N− | 1 | 1 | ||
| N+ | 3.50 [1.84; 6.69] | 2.12 [1.06; 4.23] | ||
| BRCA1 Promoter Hypermethylation | 0.021 | 0.052 | ||
| No | 1 | 1 | ||
| Yes | 0.35 [0.12; 0.98] | 0.39 [0.14; 1.13] | ||
| C. Multivariate Analysis: Adjuvant Chemotherapy = No | ||||
| (n = 66) | Relapse-Free Survival | Overall Survival | ||
| Hazard Ratio [95% CI] | p-Value | Hazard Ratio [95% CI] | p-Value | |
| Tumor Size | 0.017 | |||
| T1 | 1 | |||
| T2 | 1.35 [0.58; 3.14] | |||
| T3/T4 | 5.18 [1.61; 16.7] | |||
| Nodal Status | <0.001 | 0.019 | ||
| N− | 1 | 1 | ||
| N+ | 6.81 [2.99; 15.5] | 2.84 [1.20; 6.70] | ||
| Histology | 0.020 | |||
| Ductal | 1 | |||
| Lobular | 0.31 [0.07; 1.39] | |||
| Other | 0.22 [0.05; 0.96] | |||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jacot, W.; Lopez-Crapez, E.; Mollevi, C.; Boissière-Michot, F.; Simony-Lafontaine, J.; Ho-Pun-Cheung, A.; Chartron, E.; Theillet, C.; Lemoine, A.; Saffroy, R.; et al. BRCA1 Promoter Hypermethylation is Associated with Good Prognosis and Chemosensitivity in Triple-Negative Breast Cancer. Cancers 2020, 12, 828. https://doi.org/10.3390/cancers12040828
Jacot W, Lopez-Crapez E, Mollevi C, Boissière-Michot F, Simony-Lafontaine J, Ho-Pun-Cheung A, Chartron E, Theillet C, Lemoine A, Saffroy R, et al. BRCA1 Promoter Hypermethylation is Associated with Good Prognosis and Chemosensitivity in Triple-Negative Breast Cancer. Cancers. 2020; 12(4):828. https://doi.org/10.3390/cancers12040828
Chicago/Turabian StyleJacot, William, Evelyne Lopez-Crapez, Caroline Mollevi, Florence Boissière-Michot, Joelle Simony-Lafontaine, Alexandre Ho-Pun-Cheung, Elodie Chartron, Charles Theillet, Antoinette Lemoine, Raphael Saffroy, and et al. 2020. "BRCA1 Promoter Hypermethylation is Associated with Good Prognosis and Chemosensitivity in Triple-Negative Breast Cancer" Cancers 12, no. 4: 828. https://doi.org/10.3390/cancers12040828
APA StyleJacot, W., Lopez-Crapez, E., Mollevi, C., Boissière-Michot, F., Simony-Lafontaine, J., Ho-Pun-Cheung, A., Chartron, E., Theillet, C., Lemoine, A., Saffroy, R., Lamy, P.-J., & Guiu, S. (2020). BRCA1 Promoter Hypermethylation is Associated with Good Prognosis and Chemosensitivity in Triple-Negative Breast Cancer. Cancers, 12(4), 828. https://doi.org/10.3390/cancers12040828