A “Lymphocyte MicroRNA Signature” as Predictive Biomarker of Immunotherapy Response and Plasma PD-1/PD-L1 Expression Levels in Patients with Metastatic Renal Cell Carcinoma: Pointing towards Epigenetic Reprogramming

 , , ,
, , , 

Abstract
Simple Summary
Abstract
1. Introduction
2. Results
2.1. Patients Characteristics
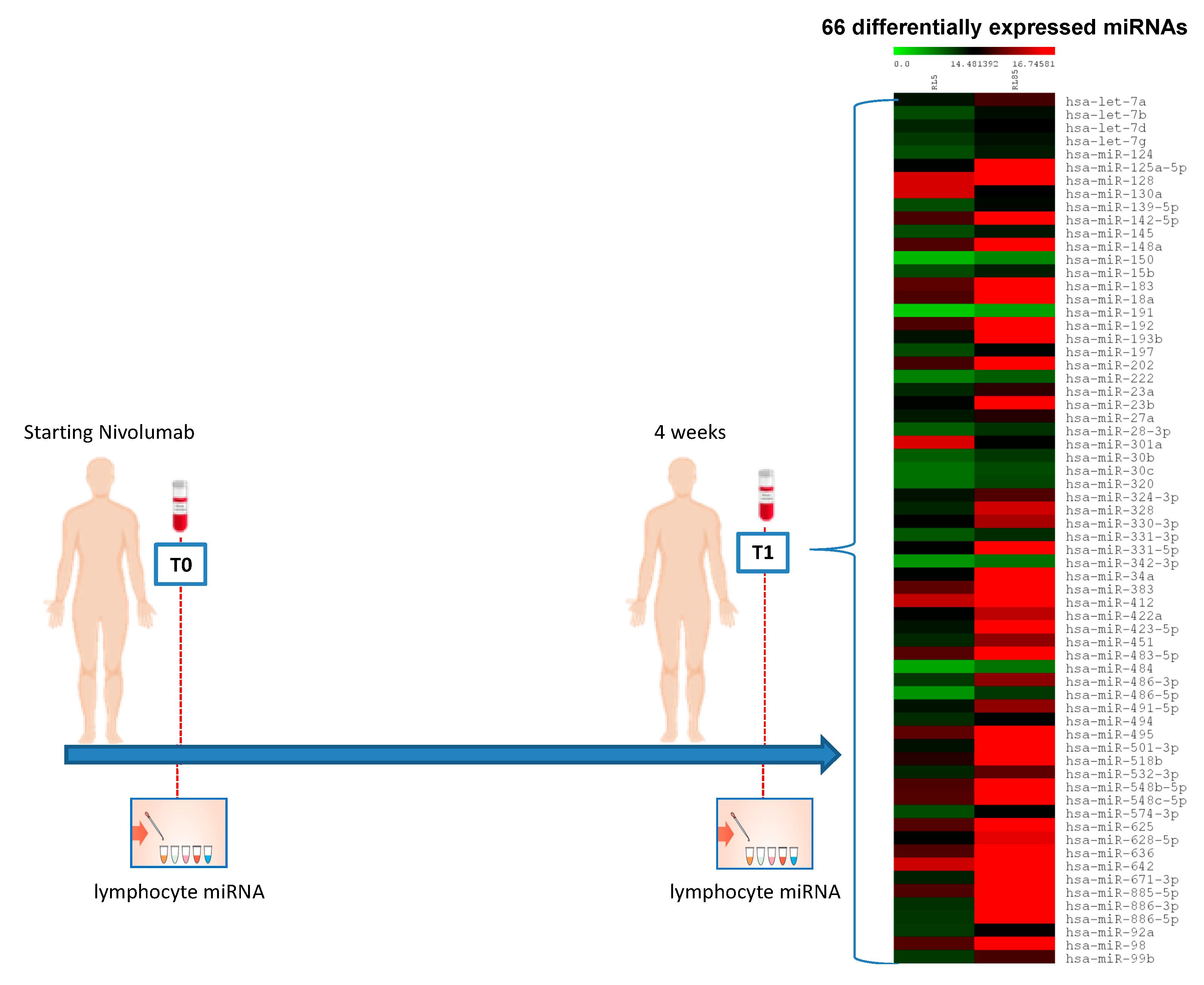
2.2. Expression Profile of Lymphocyte miRNAs as Predictive Biomarkers of Anti-PD-1 Treatment Outcome in Metastatic ccRCC Patients
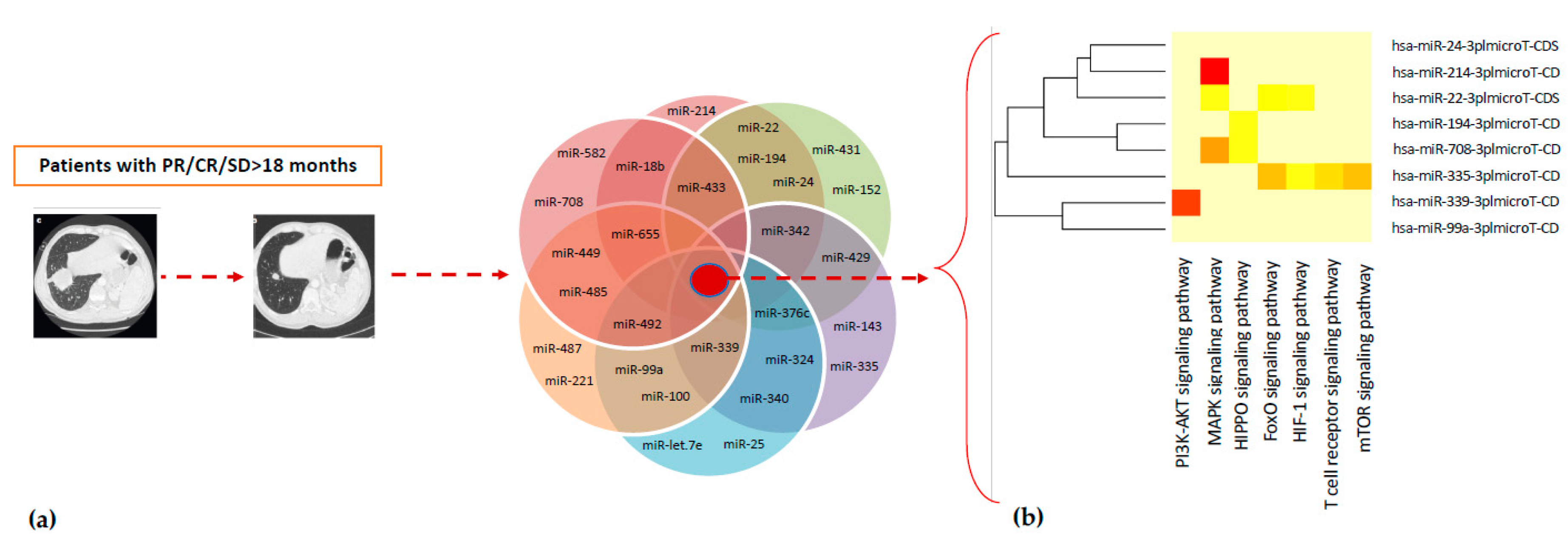
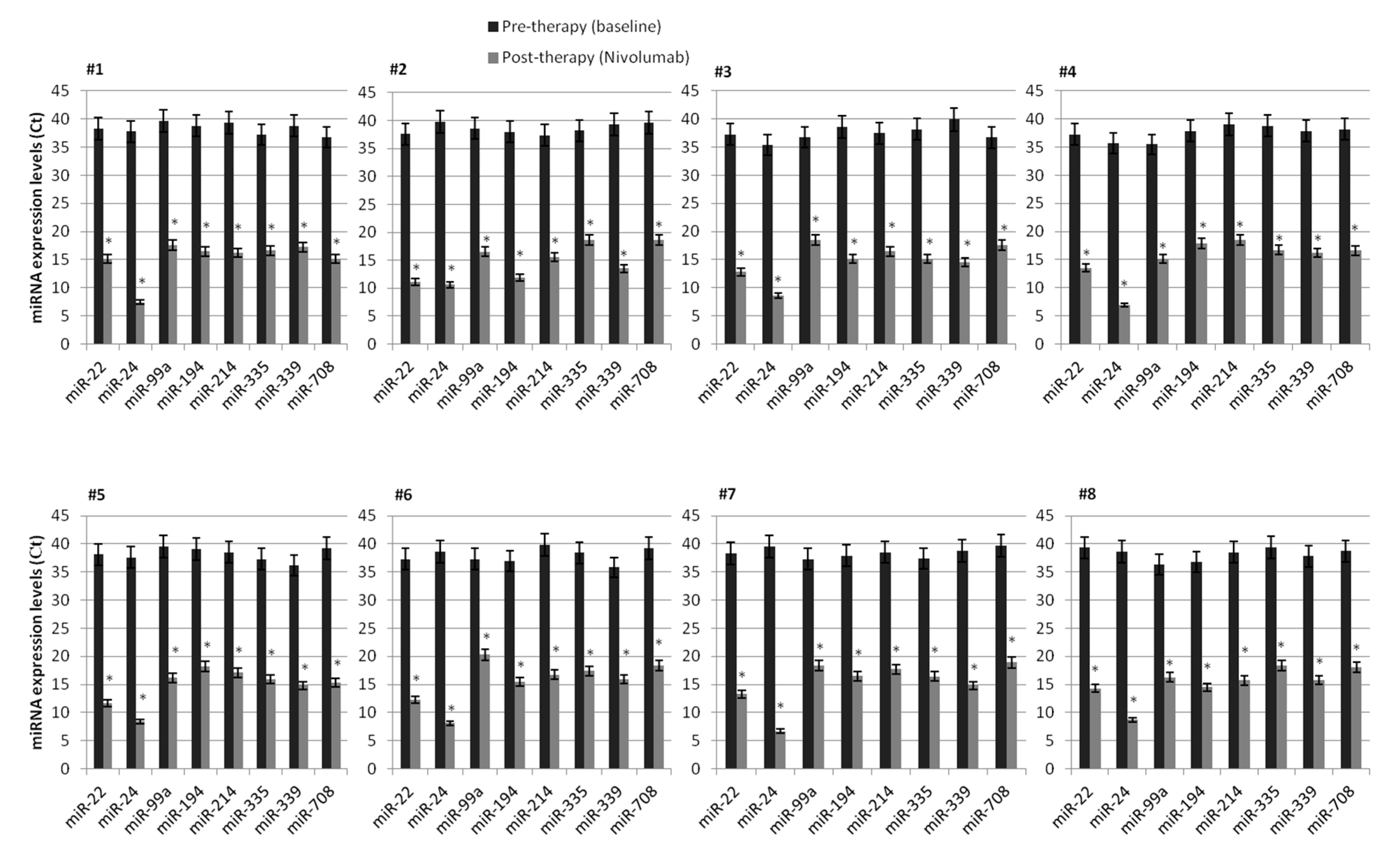
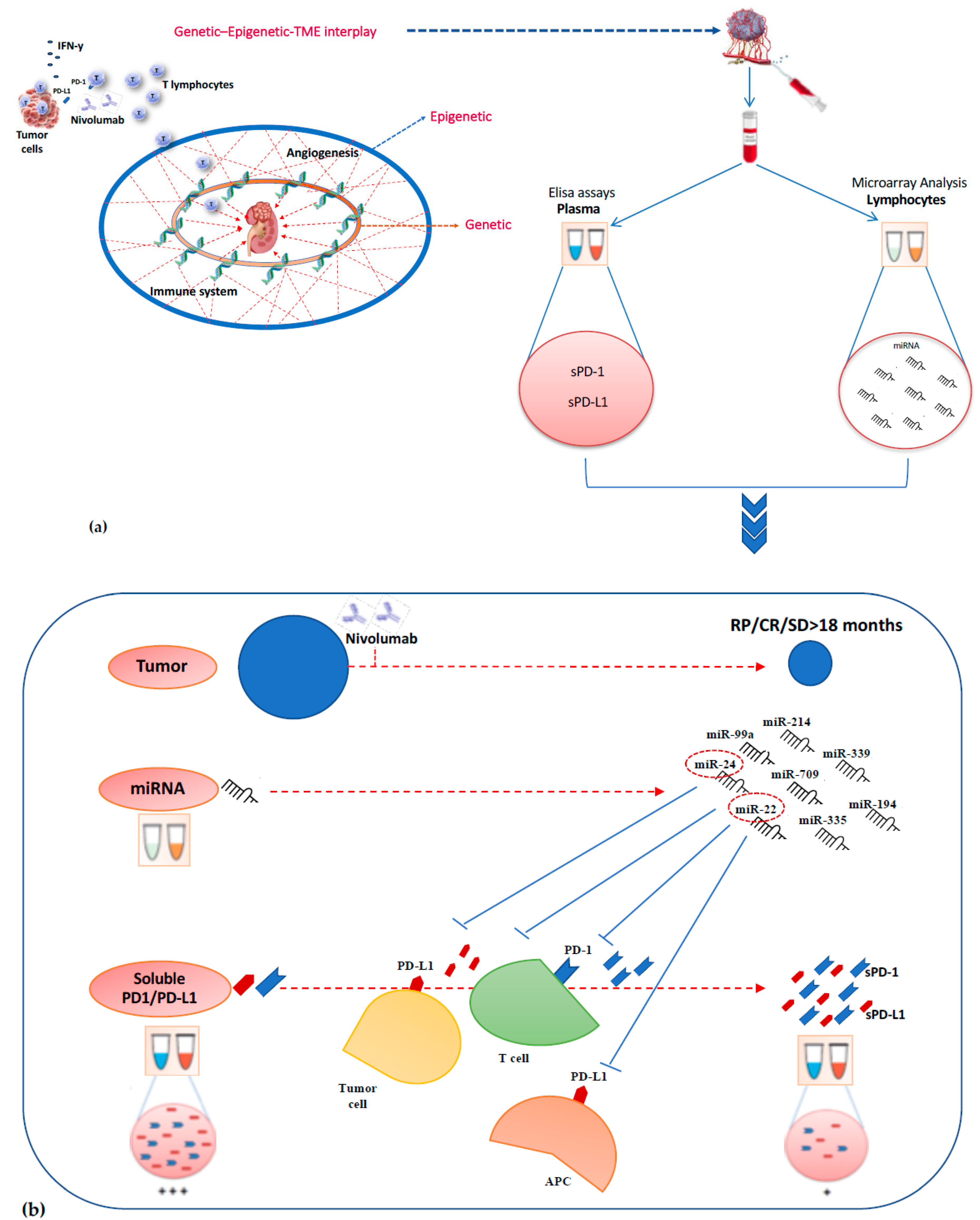
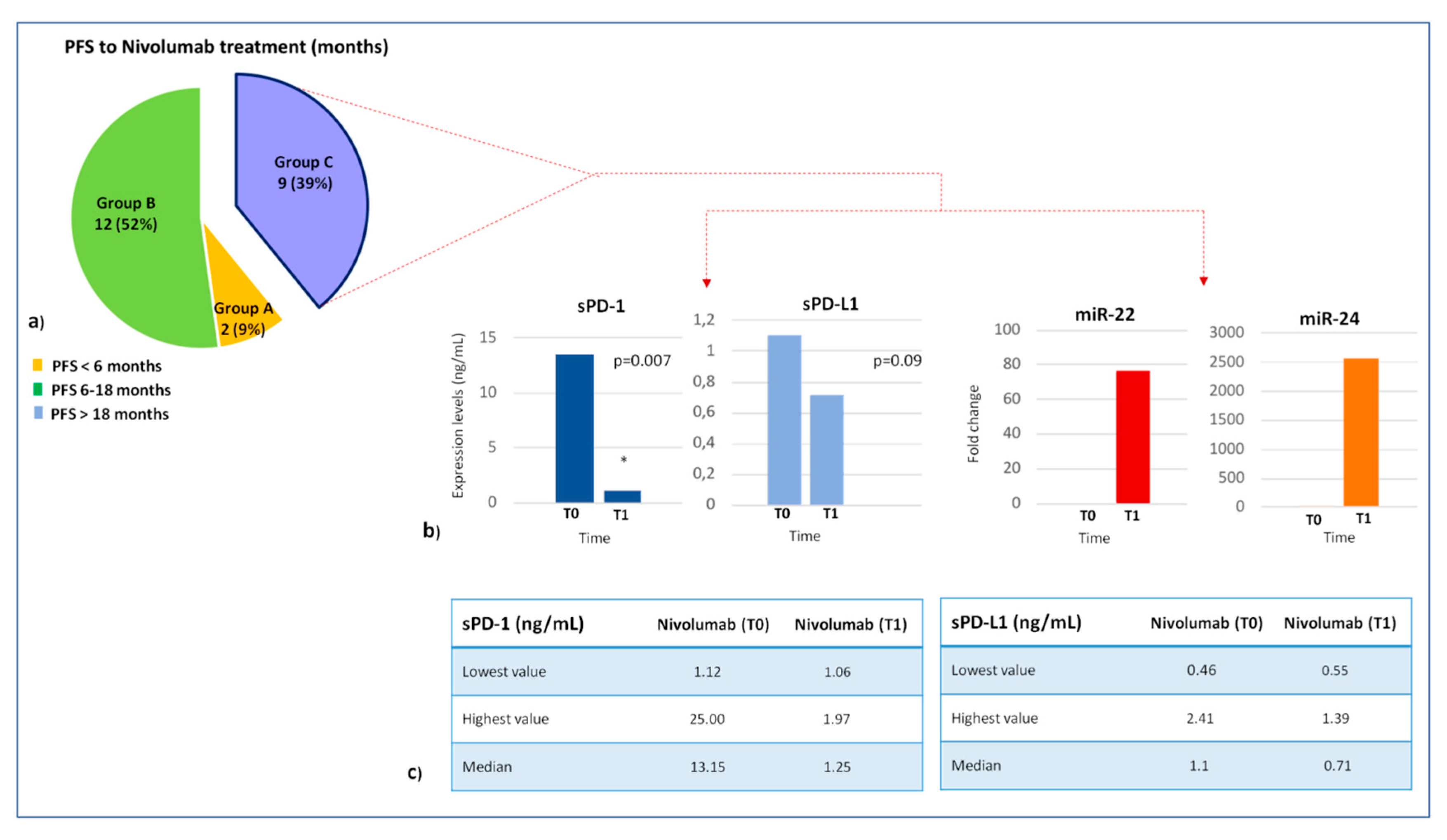
2.3. Association Between Lymphocyte miRNA Expression and Plasma Levels of Soluble PD-1/PD-L1 in Long-Responders Patients
3. Discussion
4. Patients and Methods
4.1. Study Population
4.2. Sample Collection and Lymphocytes Isolation
4.3. miRNA Expression Profile Analysis
4.4. Quantitative Real-Time PCR
4.5. miRNA Data Analysis
4.6. Determination of Soluble PD-L1 and PD-1 Concentrations in Plasma
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Angulo, J.; Shapiro, O. The Changing Therapeutic Landscape of Metastatic Renal Cancer. Cancers 2019, 11, 1227. [Google Scholar] [CrossRef]
- Motzer, R.J.; Escudier, B.; McDermott, D.F.; George, S.; Hammers, H.J.; Srinivas, S.; Tykodi, S.S.; Sosman, J.A.; Procopio, G.; Plimack, E.R.; et al. Nivolumab versus Everolimus in Advanced Renal-Cell Carcinoma. N. Engl. J. Med. 2015, 373, 1803–1813. [Google Scholar] [CrossRef]
- Motzer, R.J.; Tannir, N.M.; McDermott, D.F.; Frontera, O.A.; Melichar, B.; Choueiri, T.K.; Plimack, E.R.; Barthélémy, P.; Porta, C.; George, S.; et al. Nivolumab Plus Ipilimumab versus Sunitinib in Advanced Renal-Cell Carcinoma. N. Engl. J. Med. 2018, 378, 1277–1290. [Google Scholar] [CrossRef]
- Choueiri, T.; Motzer, R.; Rini, B.; Haanen, J.; Campbell, M.; Venugopal, B.; Kollmannsberger, C.; Gravis-Mescam, G.; Uemura, M.; Lee, J.; et al. Updated Efficacy Results from the JAVELIN Renal 101 Trial: First- Line Avelumab Plus Axitinib Versus Sunitinib in Patients With Advanced Renal Cell Carcinoma. Ann. Oncol. 2020, 31, 1030–1039. [Google Scholar] [CrossRef]
- Rini, B.I.; Plimack, E.R.; Stus, V.; Gafanov, R.; Hawkins, R.; Nosov, D.; Pouliot, F.; Alekseev, B.; Soulières, D.; Melichar, B.; et al. Pembrolizumab Plus Axitinib versus Sunitinib for Advanced Renal-Cell Carcinoma. N. Engl. J. Med. 2019, 380, 1116–1127. [Google Scholar] [CrossRef]
- Incorvaia, L.; Fanale, D.; Badalamenti, G.; Barraco, N.; Bono, M.; Corsini, L.R.; Galvano, A.; Gristina, V.; Listì, A.; Vieni, S.; et al. Programmed Death Ligand 1 (PD-L1) as a Predictive Biomarker for Pembrolizumab Therapy in Patients with Advanced Non-Small-Cell Lung Cancer (NSCLC). Adv. Ther. 2019, 36, 2600–2617. [Google Scholar] [CrossRef]
- Joosten, S.C.; Smits, K.M.; Aarts, M.J.; Melotte, V.; Koch, A.; Tjan-Heijnen, V.C.; Van Engeland, M. Epigenetics in Renal Cell Cancer: Mechanisms and Clinical Applications. Nat. Rev. Urol. 2018, 15, 430–451. [Google Scholar] [CrossRef]
- Badalamenti, G.; Fanale, D.; Incorvaia, L.; Barraco, N.; Listì, A.; Maragliano, R.; Vincenzi, B.; Calò, V.; Iovanna, J.L.; Bazan, V.; et al. Role of Tumor-Infiltrating Lymphocytes in Patients With Solid Tumors: Can a Drop Dig a Stone? Cell. Immunol. 2019, 343, 103753. [Google Scholar] [CrossRef]
- Schanza, L.-M.; Seles, M.; Stotz, M.; Fosselteder, J.; Hutterer, G.C.; Pichler, M.; Stiegelbauer, V. MicroRNAs Associated with Von Hippel–Lindau Pathway in Renal Cell Carcinoma: A Comprehensive Review. Int. J. Mol. Sci. 2017, 18, 2495. [Google Scholar] [CrossRef]
- Rolfo, C.; Fanale, D.; Hong, D.; Tsimberidou, A.M.; Piha-Paul, S.; Pauwels, P.; Meerbeeck, J.; Caruso, S.; Bazan, V.; Cicero, G.; et al. Impact of MicroRNAs in Resistance to Chemotherapy and Novel Targeted Agents in Non-Small Cell Lung Cancer. Curr. Pharm. Biotechnol. 2014, 15, 475–485. [Google Scholar] [CrossRef]
- Fanale, D.; Castiglia, M.; Bazan, V.; Russo, A. Involvement of Non-coding RNAs in Chemo- and Radioresistance of Colorectal Cancer. Adv. Exp. Med. Biol. 2016, 937, 207–228. [Google Scholar] [PubMed]
- Bronte, F.; Bronte, G.; Fanale, D.; Caruso, S.; Bronte, E.; Bavetta, M.G.; Fiorentino, E.; Rolfo, C.; Bazan, V.; Di Marco, V.; et al. HepatomiRNoma: The Proposal of a New Network of Targets for Diagnosis, Prognosis and Therapy in Hepatocellular Carcinoma. Crit. Rev. Oncol. 2016, 97, 312–321. [Google Scholar] [CrossRef] [PubMed]
- Arif, K.M.T.; Elliott, E.K.; Haupt, L.M.; Griffiths, L.R. Regulatory Mechanisms of Epigenetic miRNA Relationships in Human Cancer and Potential as Therapeutic Targets. Cancers 2020, 12, 2922. [Google Scholar] [CrossRef] [PubMed]
- Dhawan, A.; Scott, J.G.; Harris, A.L.; Buffa, F.M. Pan-Cancer Characterisation of MicroRNA across Cancer Hallmarks Reveals MicroRNA-Mediated Downregulation of Tumour Suppressors. Nat. Commun. 2018, 9, 1–13. [Google Scholar] [CrossRef]
- Dias, F.; Teixeira, A.L.; Ferreira, M.; Adem, B.; Bastos, N.; Vieira, J.; Fernandes, M.; Sequeira, M.I.; Maurício, J.; Lobo, F.; et al. Plasmatic miR-210, miR-221 and miR-1233 Profile: Potential Liquid Biopsies Candidates for Renal Cell Carcinoma. Oncotarget 2017, 8, 103315–103326. [Google Scholar] [CrossRef]
- Corsini, L.R.; Bronte, G.; Terrasi, M.; Amodeo, V.; Fanale, D.; Fiorentino, E.; Cicero, G.; Bazan, V.; Russo, A. The Role of MicroRNAs in Cancer: Diagnostic and Prognostic Biomarkers and Targets of Therapies. Expert Opin. Ther. Targets 2012, 16, S103–S109. [Google Scholar] [CrossRef]
- Caruso, S.; Bazan, V.; Rolfo, C.; Insalaco, L.; Fanale, D.; Bronte, G.; Corsini, L.R.; Rizzo, S.; Cicero, G.; Russo, A. MicroRNAs in Colorectal Cancer Stem Cells: New Regulators of Cancer Stemness? Oncogenesis 2012, 1, e32. [Google Scholar] [CrossRef]
- Cortez, M.A.; Anfossi, S.; Ramapriyan, R.; Menon, H.; Atalar, S.C.; Aliru, M.; Welsh, J.; Calin, G.A. Role of miRNAs in Immune Responses and Immunotherapy in Cancer. Genes Chromosom. Cancer 2019, 58, 244–253. [Google Scholar] [CrossRef]
- Tsiakanikas, P.; Giaginis, C.; Kontos, C.K.; Scorilas, A.; Constantinos, G. Clinical Utility of MicroRNAs in Renal Cell Carcinoma: Current Evidence and Future Perspectives. Expert Rev. Mol. Diagn. 2018, 18, 981–991. [Google Scholar] [CrossRef]
- Incorvaia, L.; Bronte, G.; Bazan, V.; Badalamenti, G.; Rizzo, S.; Pantuso, G.; Natoli, C.; Russo, A. Beyond Evidence-Based Data: Scientific Rationale and Tumor Behavior to Drive Sequential and Personalized Therapeutic Strategies for the Treatment of Metastatic Renal Cell Carcinoma. Oncotarget 2016, 7, 21259–21271. [Google Scholar] [CrossRef][Green Version]
- Corsini, L.R.; Fanale, D.; Passiglia, F.; Incorvaia, L.; Gennusa, V.; Bazan, V.; Russo, A. Monoclonal Antibodies for the Treatment of Non-Hematological Tumors: A Safety Review. Expert Opin. Drug Saf. 2018, 17, 1197–1209. [Google Scholar] [CrossRef]
- Linehan, W.M.; Ricketts, C.J. The Cancer Genome Atlas of Renal Cell Carcinoma: Findings and Clinical Implications. Nat. Rev. Urol. 2019, 16, 539–552. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.; Huang, J.; Xiao, H.; Liang, Z. MicroRNA-22 is Downregulated in Clear Cell Renal Cell Carcinoma, and Inhibits Cell Growth, Migration and Invasion by Targeting PTEN. Mol. Med. Rep. 2016, 13, 4800–4806. [Google Scholar] [CrossRef] [PubMed]
- Bar, N.; Dikstein, R. miR-22 Forms a Regulatory Loop in PTEN/AKT Pathway and Modulates Signaling Kinetics. PLoS ONE 2010, 5, e10859. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, N.; Yi, C.; Wang, Y.; Wang, H.; Wang, J. MicroRNA-22 Functions as a Tumor Suppressor by Targeting SIRT1 in Renal Cell Carcinoma. Oncol. Rep. 2015, 35, 559–567. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.; Zhao, H.; Saar, M.; Peehl, D.M.; Brooks, J.D. miR-22 Regulates Invasion, Gene Expression and Predicts Overall Survival in Patients with Clear Cell Renal Cell Carcinoma. Kidney Cancer 2019, 3, 119–132. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Li, Y.; Nie, L.; He, T.; Hu, J.; Liu, J.; Chen, M.; Shi, M.; Jiang, Z.; Gui, Y.; et al. MicroRNA-24-2 is Associated With Cell Proliferation, Invasion, Migration and Apoptosis in Renal Cell Carcinoma. Mol. Med. Rep. 2017, 16, 9157–9164. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Quan, J.; Liu, S.; Dai, K.; Jin, L.; He, T.; Pa, X.; Lai, Y. MicroRNA-23a/24-2/27a as a Potential Diagnostic Biomarker for Cancer: A Systematic Review and Meta-Analysis. Mol. Clin. Oncol. 2017. [Google Scholar] [CrossRef]
- Cui, L.; Zhou, H.; Zhao, H.; Zhou, Y.; Xu, R.; Xu, X.; Zheng, L.; Xue, Z.; Xia, W.; Zhang, B.; et al. MicroRNA-99a Induces G1-Phase Cell Cycle Arrest and Suppresses Tumorigenicity in Renal Cell Carcinoma. BMC Cancer 2012, 12, 546. [Google Scholar] [CrossRef]
- Braga, E.A.; Fridman, M.V.; Loginov, V.I.; Dmitriev, A.A.; Morozov, S.G. Molecular Mechanisms in Clear Cell Renal Cell Carcinoma: Role of miRNAs and Hypermethylated miRNA Genes in Crucial Oncogenic Pathways and Processes. Front. Genet. 2019, 10. [Google Scholar] [CrossRef]
- Osako, Y.; Yoshino, H.; Sakaguchi, T.; Sugita, S.; Yonemori, M.; Nakagawa, M.; Enokida, H. Potential Tumor Suppressive Role of MicroRNA 99a 3p in Sunitinib Resistant Renal Cell Carcinoma Cells Through the Regulation of RRM2. Int. J. Oncol. 2019, 54, 1759–1770. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, R.D.C.; Ivanovic, R.F.; Leite, K.R.M.; Viana, N.I.; Pimenta, R.C.; Pontes-Júnior, J.; Guimarães, V.; Morais, D.R.; Abe, D.K.; Nesrallah, A.J.; et al. Expression of Micro-RNAs and Genes Related to Angiogenesis in ccRCC and Associations With Tumor Characteristics. BMC Urol. 2017, 17, 113. [Google Scholar] [CrossRef] [PubMed]
- Nofech-Mozes, R.; Khella, H.W.Z.; Scorilas, A.; Youssef, L.; Krylov, S.N.; Lianidou, E.; Sidiropoulos, K.G.; Gabril, M.; Evans, A.; Mac-Way, F. MicroRNA-194 is a Marker for Good Prognosis in Clear Cell Renal Cell Carcinoma. Cancer Med. 2016, 5, 656–664. [Google Scholar] [CrossRef] [PubMed]
- Khella, H.; Bakhet, M.; Allo, G.; Jewett, M.; Girgis, A.; Latif, A.; Von Both, I.; Bjarnason, G.; Mac-Way, F. miR-192, miR-194 and miR-215: A Convergent MicroRNA Network Suppressing Tumor Progression in Renal Cell Carcinoma. Carcinogenesis 2013, 34, 2231–2239. [Google Scholar] [CrossRef]
- Jiao, M.; Guo, H.; Chen, Y.; Li, L.; Zhang, L. DARS-AS1 Promotes Clear Cell Renal Cell Carcinoma by Sequestering Mir-194-5p to up-Regulate DARS. Biomed. Pharmacother. 2020, 128, 110323. [Google Scholar] [CrossRef]
- Yumioka, T.; Osaki, M.; Sasaki, R.; Yamaguchi, N.; Onuma, K.; Iwamoto, H.; Morizane, S.; Honda, M.; Takenaka, A.; Okada, F. Lysosome-Associated Membrane Protein 2 (LAMP-2) Expression Induced by Mir-194-5p Downregulation Contributes to Sunitinib Resistance in Human Renal Cell Carcinoma Cells. Oncol. Lett. 2017. [Google Scholar] [CrossRef]
- Ran, L.; Liang, J.; Deng, X.; Wu, J. miRNAs in Prediction of Prognosis in Clear Cell Renal Cell Carcinoma. BioMed Res. Int. 2017, 2017, 1–6. [Google Scholar] [CrossRef]
- Das, F.; Dey, N.; Bera, A.; Kasinath, B.S.; Ghosh-Choudhury, N.; Choudhury, G.G. MicroRNA-214 Reduces Insulin-like Growth Factor-1 (IGF-1) Receptor Expression and Downstream mTORC1 Signaling in Renal Carcinoma Cells. J. Biol. Chem. 2016, 291, 14662–14676. [Google Scholar] [CrossRef]
- Xu, H.; Wu, S.; Shen, X.; Shi, Z.; Wu, D.; Yuan, Y.; Jiang, W.; Wang, Q.; Ke, Q.; Mao, Q.; et al. Methylation-Mediated miR-214 Regulates Proliferation and Drug Sensitivity of Renal Cell Carcinoma Cells Through Targeting LIVIN. J. Cell. Mol. Med. 2020, 24, 6410–6425. [Google Scholar] [CrossRef]
- Wang, K.; Chen, X.; Zhan, Y.; Jiang, W.; Liu, X.; Wang, X.; Wu, B. miR-335 Inhibits the Proliferation and Invasion of Clear Cell Renal Cell Carcinoma Cells Through Direct Suppression of BCL-W. Tumor Biol. 2015, 36, 6875–6882. [Google Scholar] [CrossRef]
- Liu, H.; Yan, L.; Tang, K.; Ye, T.; Sun, X.; Yang, X.; Xu, H.; Yuan, P. MP16-13 Setd2-Mediated Loss of Mir-339 Promotes Tumor PD-L1 Expression and Attenuates Antitumor Immunity in Renal Cell Carcinoma. J. Urol. 2019, 201. [Google Scholar] [CrossRef]
- Saini, S.; Yamamura, S.; Majid, S.; Shahryari, V.; Hirata, H.; Tanaka, Y.; Dahiya, R. MicroRNA-708 Induces Apoptosis and Suppresses Tumorigenicity in Renal Cancer Cells. Cancer Res. 2011, 71, 6208–6219. [Google Scholar] [CrossRef] [PubMed]
- Mihnea, D.; Baoqing, C.; Xiao, F.; George, A.C.; Dragomir, M.; Chen, B.; Fu, X.; Calin, G.A. Key Questions About the Checkpoint Blockade-Are MicroRNAs an Answer? Cancer Biol. Med. 2018, 15, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Gibbons, D.L.; Goswami, S.; Cortez, M.A.A.; Ahn, Y.-H.; Byers, L.A.; Zhang, X.; Yi, X.; Dwyer, D.W.; Lin, W.; et al. Metastasis is Regulated via MicroRNA-200/ZEB1 Axis Control of Tumour Cell PD-L1 Expression and Intratumoral Immunosuppression. Nat. Commun. 2014, 5, 1–12. [Google Scholar] [CrossRef]
- Raimondi, A.; Randon, G.; Sepe, P.; Claps, M.; Verzoni, E.; De Braud, F.; Procopio, G. The Evaluation of Response to Immunotherapy in Metastatic Renal Cell Carcinoma: Open Challenges in the Clinical Practice. Int. J. Mol. Sci. 2019, 20, 4263. [Google Scholar] [CrossRef]
- George, S.; Motzer, R.J.; Hammers, H.J.; Redman, B.G.; Kuzel, T.M.; Tykodi, S.S.; Plimack, E.R.; Jiang, J.; Waxman, I.M.; Rini, B.I. Safety and Efficacy of Nivolumab in Patients With Metastatic Renal Cell Carcinoma Treated Beyond Progression. JAMA Oncol. 2016, 2, 1179–1186. [Google Scholar] [CrossRef]
- Incorvaia, L.; Fanale, D.; Badalamenti, G.; Porta, C.; Olive, D.; De Luca, I.; Brando, C.; Rizzo, M.; Messina, C.; Rediti, M.; et al. Baseline Plasma Levels of Soluble PD-1, PD-L1, and BTN3A1 Predict Response to Nivolumab Treatment in Patients With Metastatic Renal Cell Carcinoma: A Step Toward a Biomarker for Therapeutic Decisions. OncoImmunology 2020, 9, 1832348. [Google Scholar] [CrossRef]
- Di Fiore, R.; Fanale, D.; Drago-Ferrante, R.; Chiaradonna, F.; Giuliano, M.; De Blasio, A.; Amodeo, V.; Corsini, L.R.; Bazan, V.; Tesoriere, G.; et al. Genetic and Molecular Characterization of the Human Osteosarcoma 3AB-OS Cancer Stem Cell Line: A Possible Model for Studying Osteosarcoma Origin and Stemness. J. Cell. Physiol. 2013, 228, 1189–1201. [Google Scholar] [CrossRef]
- Rizzo, S.; Cangemi, A.; Galvano, A.; Fanale, D.; Buscemi, S.; Ciaccio, M.; Russo, A.; Castorina, S.; Bazan, V. Analysis of miRNA Expression Profile Induced by Short Term Starvation in Breast Cancer Cells Treated With Doxorubicin. Oncotarget 2017, 8, 71924–71932. [Google Scholar] [CrossRef]
- Fanale, D.; Amodeo, V.; Bazan, V.; Insalaco, L.; Incorvaia, L.; Barraco, N.; Castiglia, M.; Rizzo, S.; Santini, D.; Giordano, A.; et al. Can the microRNA Expression Profile Help to Identify Novel Targets for Zoledronic Acid in Breast Cancer? Oncotarget 2016, 7, 29321–29332. [Google Scholar] [CrossRef]
- Cabibi, D.; Caruso, S.; Bazan, V.; Castiglia, M.; Bronte, G.; Ingrao, S.; Fanale, D.; Cangemi, A.; Calò, V.; Listi, A.; et al. Analysis of Tissue and Circulating MicroRNA Expression During Metaplastic Transformation of the Esophagus. Oncotarget 2016, 7, 47821–47830. [Google Scholar] [CrossRef] [PubMed]
- Vlachos, I.S.; Kostoulas, N.; Vergoulis, T.; Georgakilas, G.; Reczko, M.; Maragkakis, M.; Paraskevopoulou, M.D.; Prionidis, K.; Dalamagas, T.; Hatzigeorgiou, A.G. DIANA miRPath v.2.0: Investigating the Combinatorial Effect of MicroRNAs in Pathways. Nucleic Acids Res. 2012, 40, W498–W504. [Google Scholar] [CrossRef] [PubMed]
- Fromm, B.; Billipp, T.; Peck, L.E.; Johansen, M.; Tarver, J.E.; King, B.L.; Newcomb, J.M.; Sempere, L.F.; Flatmark, K.; Hovig, E.; et al. A Uniform System for the Annotation of Vertebrate microRNA Genes and the Evolution of the Human MicroRNAome. Annu. Rev. Genet. 2015, 49, 213–242. [Google Scholar] [CrossRef]
- Kozomara, A.; Griffiths-Jones, S. miRBase: Integrating MicroRNA Annotation and Deep-Sequencing Data. Nucleic Acids Res. 2011, 39, D152–D157. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S.; Sato, Y.; Furumichi, M.; Tanabe, M. KEGG for Integration and Interpretation of Large-Scale Molecular Data Sets. Nucleic Acids Res. 2012, 40, D109–D114. [Google Scholar] [CrossRef]
- Bian, B.; Fanale, D.; Dusetti, N.; Roque, J.; Pastor, S.; Chretien, A.-S.; Incorvaia, L.; Russo, A.; Olive, D.; Iovanna, J. Prognostic Significance of Circulating PD-1, PD-L1, pan-BTN3As, BTN3A1 and BTLA in Patients With Pancreatic Adenocarcinoma. OncoImmunology 2019, 8, e1561120. [Google Scholar] [CrossRef] [PubMed]
- Incorvaia, L.; Badalamenti, G.; Rinaldi, G.; Iovanna, J.L.; Olive, D.; Swayden, M.; Terruso, L.; Vincenzi, B.; Fulfaro, F.; Bazan, V.; et al. Can the Plasma PD-1 Levels Predict the Presence and Efficiency of Tumor-Infiltrating Lymphocytes in Patients With Metastatic Melanoma? Ther. Adv. Med. Oncol. 2019, 11, 175883591984887. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Baseline Characteristics | |
|---|---|
| Tot (No.) | 23 |
| Median Age (range)—years | 62 (36–71) |
| Sex, n. (%) | |
| Male | 19 (82.6%) |
| Female | 4 (17.4%) |
| Histological classification | |
| Clear cell | 23 (100%) |
| Other | 0 (0%) |
| Prior nephrectomy | |
| Yes | 7 (30.4%) |
| No | 16 (69.6%) |
| No. of evaluable disease sites, n (%) | |
| ≤2 | 5 (19.1%) |
| ≥3 | 18 (80.9%) |
| Site of metastasis | |
| Lung only | 7 (30.4%) |
| Lung + others | 16 (69.6) |
| Site of metastasis, individual | |
| Lung | 11 (47.8%) |
| Lymph node | 12 (52.2%) |
| Liver | 6 (26.1%) |
| Bone | 5 (21.7%) |
| Pancreas | 1 (4.3%) |
| SNC | 4 (17.4%) |
| IMDC Prognostic Risk Group, n (%) | |
| Favorable | 6 (26.1%) |
| Intermediate | 17 (73.9%) |
| Poor | 0 (0%) |
| Best response to nivolumab treatment | |
| Complete Response (CR) | 1 (4.3%) |
| Partial Response (PR) | 9 (39.2%) |
| Stable Disease (SD) | 11 (47.8%) |
| Progressive Disease (PD) | 2 (8.7%) |
| Median duration of response (range) -months | 15 (3−29) |
| microRNAs | |||
|---|---|---|---|
| miR-99a | miR-708 | miR-655 | miR-582-3p |
| miR-492 | miR-487a | miR-485-3p | miR-449a |
| miR-433 | miR-431 | miR-429 | miR-376c |
| miR-342-5p | miR-340 | miR-339-5p | miR-335 |
| miR-324-5p | miR-25 | miR-24 | miR-22 |
| miR-221 | miR-214 | miR-194 | miR-18b |
| miR-152 | miR-143 | miR-100 | miR-let-7e |
| Pathway | miRNAs | No. of Gene Targets |
|---|---|---|
| PI3K-Akt signaling | 26 | 183 |
| MAPK signaling | 24 | 130 |
| T-cell receptor signaling | 24 | 55 |
| Hippo signaling | 22 | 87 |
| FOXO signaling | 22 | 75 |
| HIF-1 signaling | 22 | 56 |
| mTOR signaling | 20 | 35 |
| miRNAs | Mean Ct Value | Gene Targets |
|---|---|---|
| miR-22 | 12.86 | AKT3 |
| miR-24 | 7.80 | MAPK1 |
| miR-99a | 17.63 | GSK3B, GIMAP1, TMEM71, TIAM1, PDE4D, TRABD2A, SCGN, CUX1, CD93, EIF1, HIPK1, FNTA, LRRC28 |
| miR-194 | 16.74 | STAT6, PCDHA4, IRF1, NOS1 |
| miR-214 | 16.81 | PIK3CB, PAK3, PAK6 |
| miR-335 | 17.82 | MET, SOS2, PIK3CB, PAK2/3, TGF-A, AKT3, ETS1, PIK3R1, PIK3CG, PIK3CA, CREBBP, EGLN1 |
| miR-339 | 15.77 | PIK3CB, SOS1, PAK6, VEGFA |
| miR-708 | 17.81 | SOS2, CRK, CDC42 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Incorvaia, L.; Fanale, D.; Badalamenti, G.; Brando, C.; Bono, M.; De Luca, I.; Algeri, L.; Bonasera, A.; Corsini, L.R.; Scurria, S.; et al. A “Lymphocyte MicroRNA Signature” as Predictive Biomarker of Immunotherapy Response and Plasma PD-1/PD-L1 Expression Levels in Patients with Metastatic Renal Cell Carcinoma: Pointing towards Epigenetic Reprogramming. Cancers 2020, 12, 3396. https://doi.org/10.3390/cancers12113396
Incorvaia L, Fanale D, Badalamenti G, Brando C, Bono M, De Luca I, Algeri L, Bonasera A, Corsini LR, Scurria S, et al. A “Lymphocyte MicroRNA Signature” as Predictive Biomarker of Immunotherapy Response and Plasma PD-1/PD-L1 Expression Levels in Patients with Metastatic Renal Cell Carcinoma: Pointing towards Epigenetic Reprogramming. Cancers. 2020; 12(11):3396. https://doi.org/10.3390/cancers12113396
Chicago/Turabian StyleIncorvaia, Lorena, Daniele Fanale, Giuseppe Badalamenti, Chiara Brando, Marco Bono, Ida De Luca, Laura Algeri, Annalisa Bonasera, Lidia Rita Corsini, Salvatore Scurria, and et al. 2020. "A “Lymphocyte MicroRNA Signature” as Predictive Biomarker of Immunotherapy Response and Plasma PD-1/PD-L1 Expression Levels in Patients with Metastatic Renal Cell Carcinoma: Pointing towards Epigenetic Reprogramming" Cancers 12, no. 11: 3396. https://doi.org/10.3390/cancers12113396
APA StyleIncorvaia, L., Fanale, D., Badalamenti, G., Brando, C., Bono, M., De Luca, I., Algeri, L., Bonasera, A., Corsini, L. R., Scurria, S., Iovanna, J. L., Russo, A., & Bazan, V. (2020). A “Lymphocyte MicroRNA Signature” as Predictive Biomarker of Immunotherapy Response and Plasma PD-1/PD-L1 Expression Levels in Patients with Metastatic Renal Cell Carcinoma: Pointing towards Epigenetic Reprogramming. Cancers, 12(11), 3396. https://doi.org/10.3390/cancers12113396