Chemerin Is Induced in Non-Alcoholic Fatty Liver Disease and Hepatitis B-Related Hepatocellular Carcinoma


 and
and 
Simple Summary
Abstract
1. Introduction
2. Results
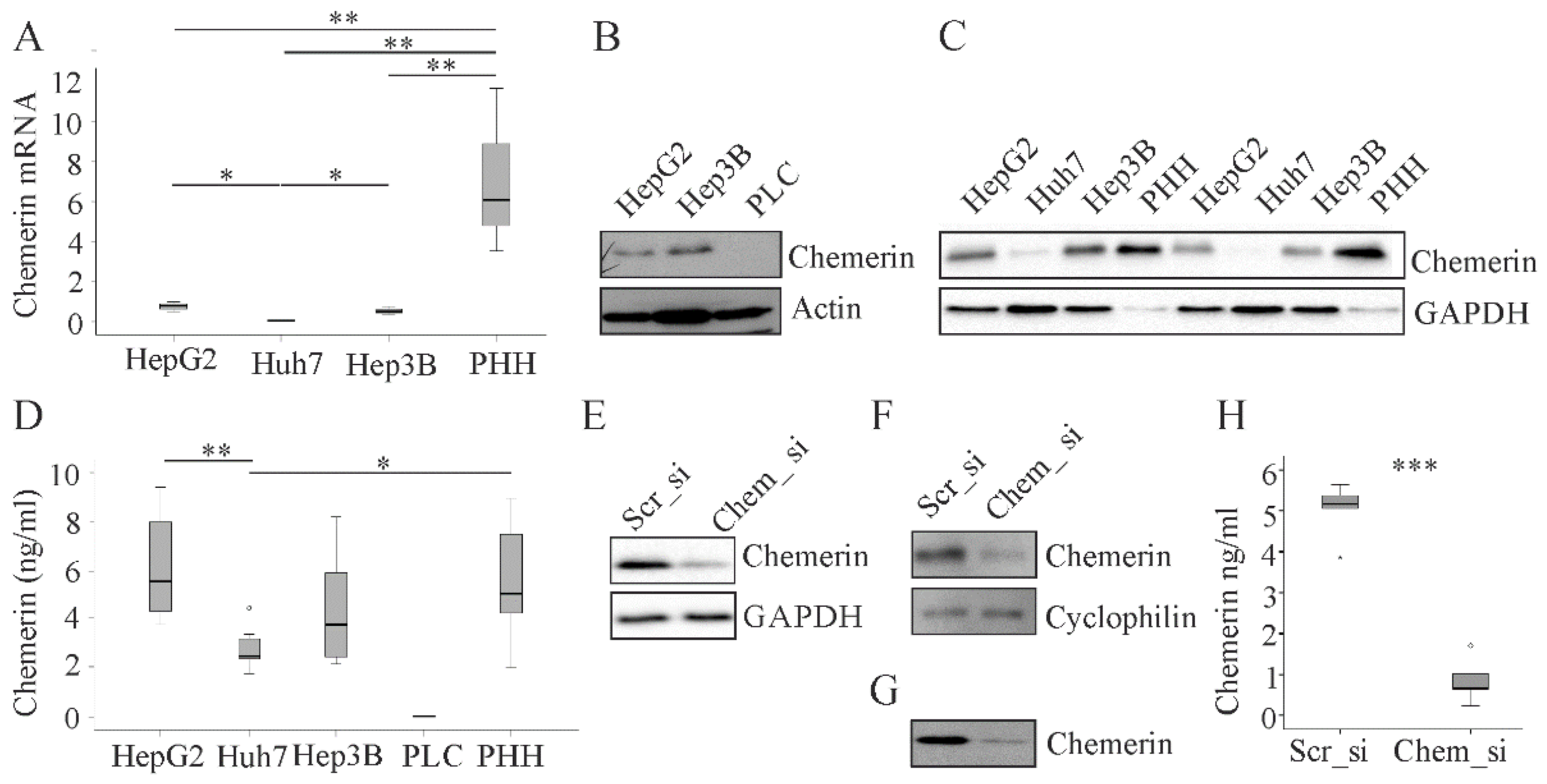
2.1. Chemerin in Primary Human Hepatocytes and Hepatocyte Cell Lines.
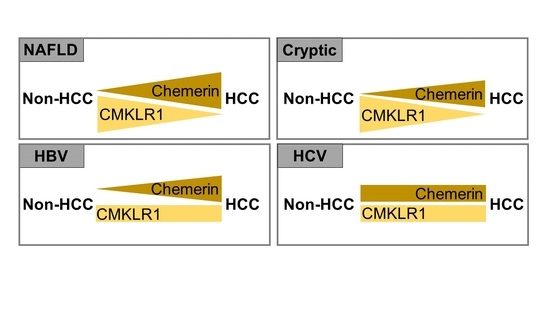
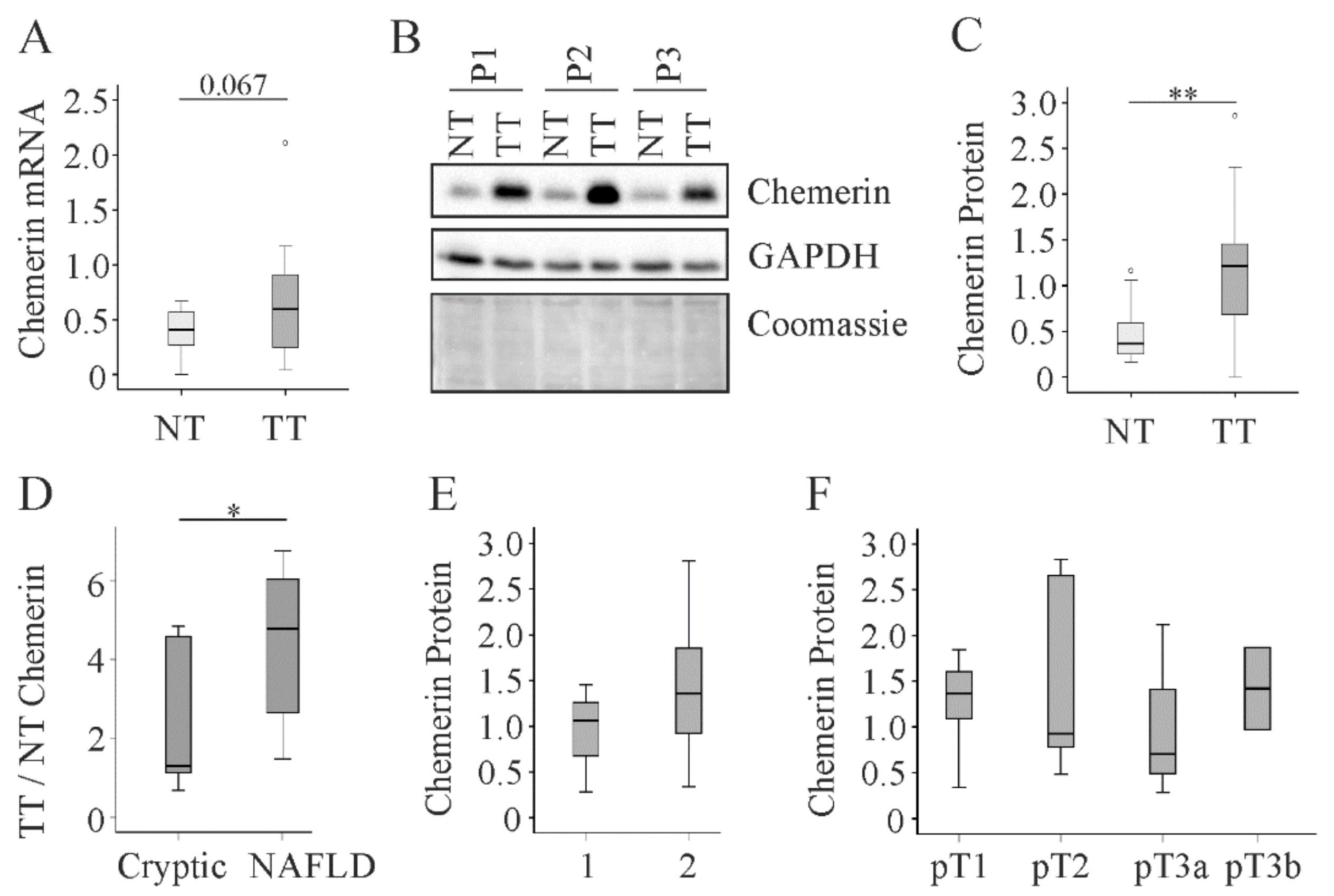
2.2. Chemerin in Human Non-Viral HCC
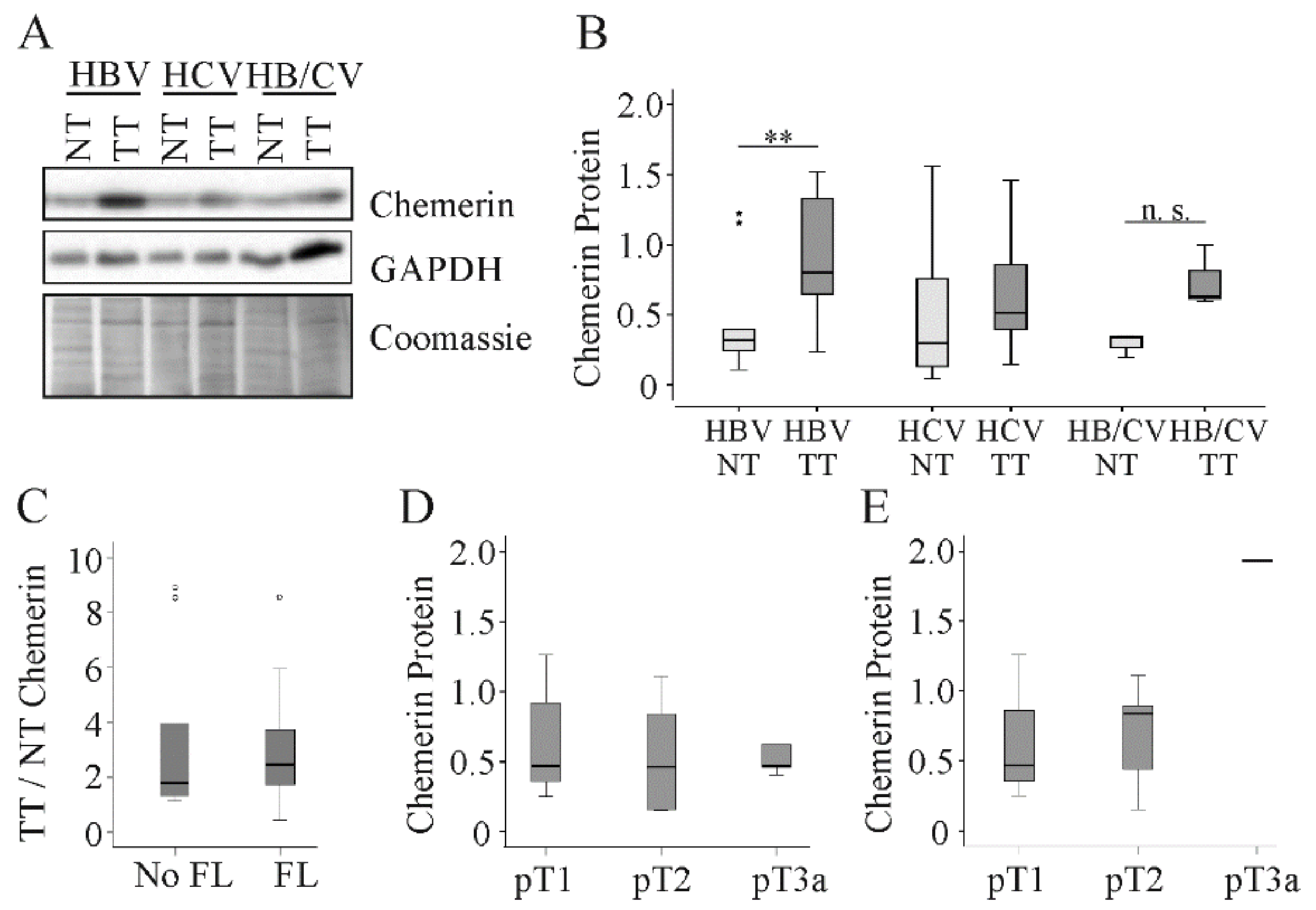
2.3. Chemerin in Human Viral HCC
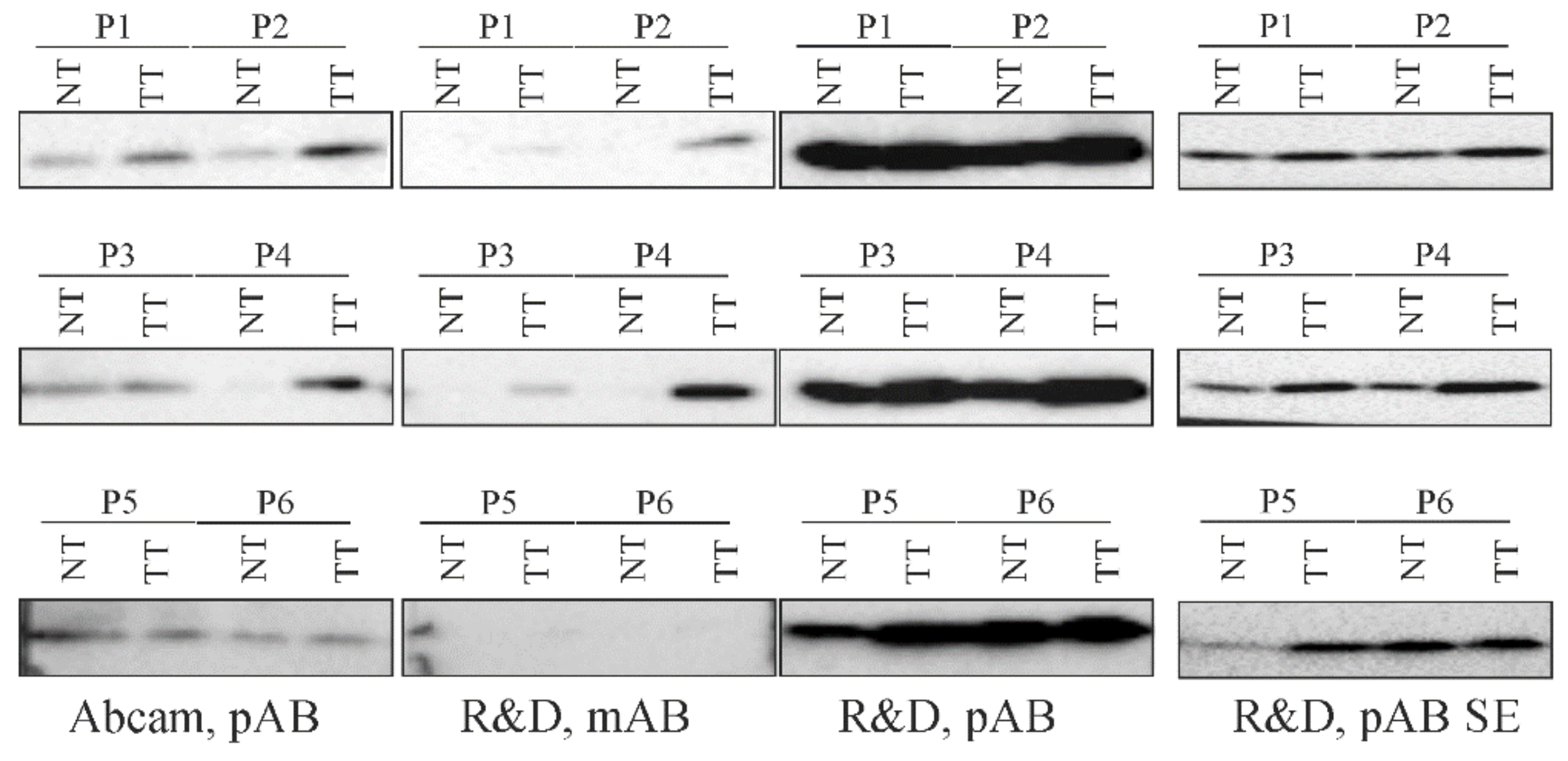
2.4. Detection of Chemerin Isoforms by Different Antibodies.
2.5. Detection of Chemerin in Human HCC Tissues by Different Antibodies
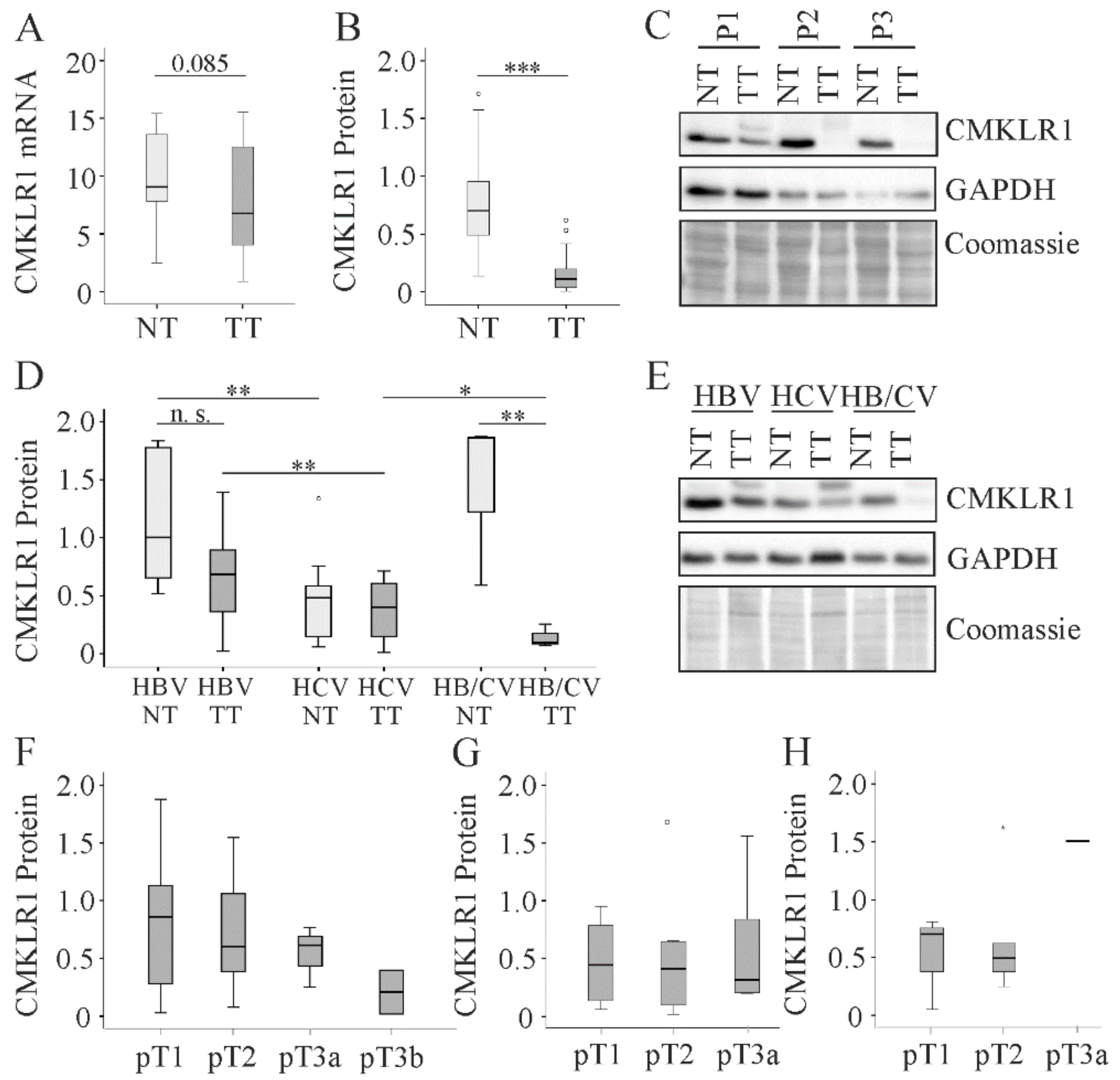
2.6. CMKLR1 in HCC
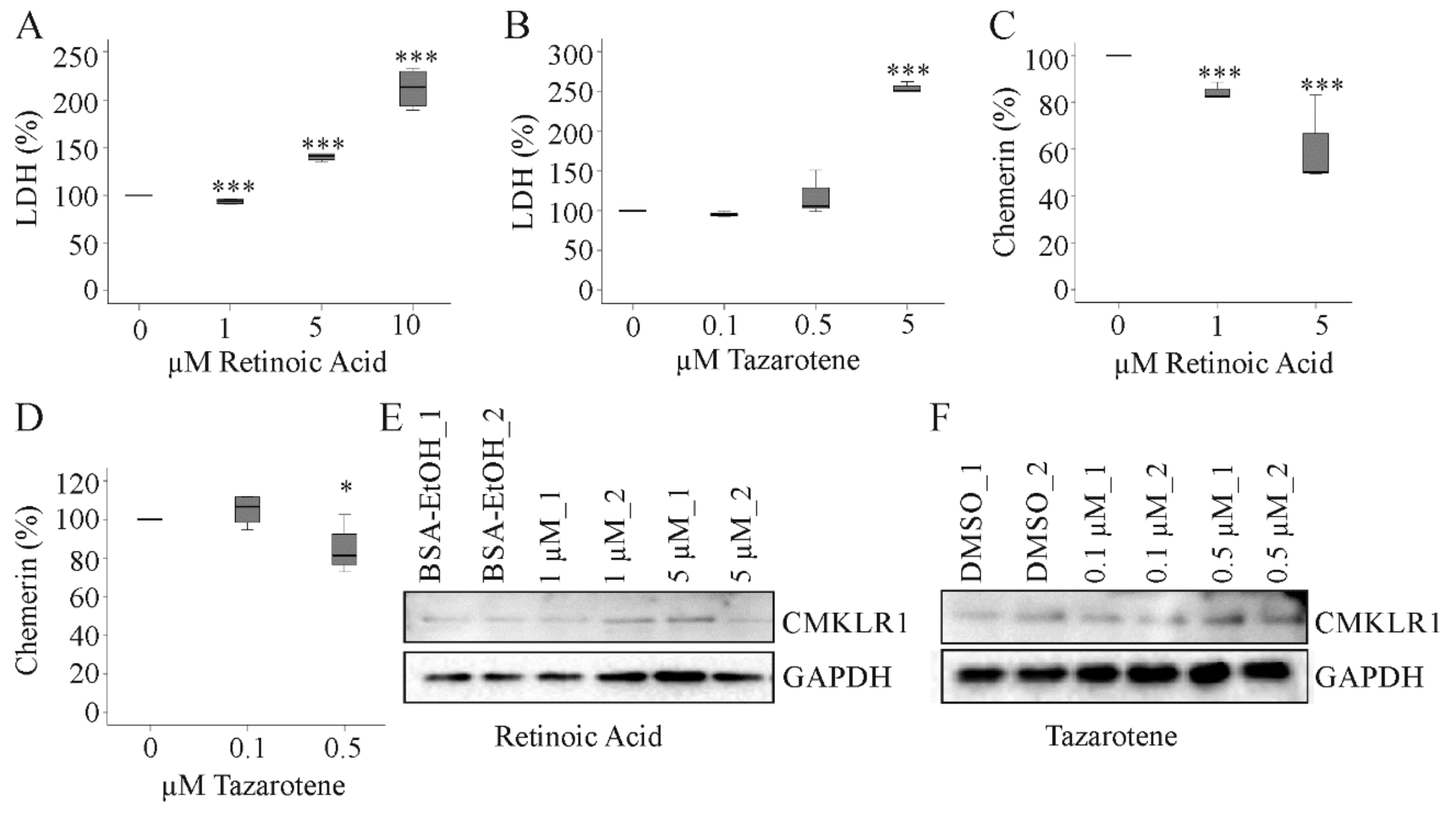
2.7. Effect of Tazarotene and Retinoic Acid on Hepatocyte Chemerin and CMKLR1 Protein
3. Discussion
4. Materials and Methods
4.1. Human HCC Tissues
4.2. Cell Lines and Primary Human Hepatocytes
4.3. Transfection of Cells with siRNA
4.4. Recombinant Expression of Chemerin Isoforms in HepG2 Cells
4.5. Tazarotene and Retinoic Acid
4.6. SDS-PAGE and Immunoblotting
4.7. ELISA
4.8. Real-Time RT-PCR.
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Buechler, C.; Feder, S.; Haberl, E.M.; Aslanidis, C. Chemerin Isoforms and Activity in Obesity. Int. J. Mol. Sci. 2019, 20, 1128. [Google Scholar] [CrossRef] [PubMed]
- Bondue, B.; Wittamer, V.; Parmentier, M. Chemerin and Its Receptors in Leukocyte Trafficking, Inflammation and Metabolism. Cytokine Growth Factor Rev. 2011, 22, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Mariani, F.; Roncucci, L. Chemerin/Chemr23 Axis in Inflammation Onset and Resolution. Inflamm. Res. 2015, 64, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Goralski, K.B.; Jackson, A.E.; McKeown, B.T.; Sinal, C.J. More Than an Adipokine: The Complex Roles of Chemerin Signaling in Cancer. Int. J. Mol. Sci. 2019, 20, 4778. [Google Scholar] [CrossRef] [PubMed]
- Treeck, O.; Buechler, C.; Ortmann, O. Chemerin and Cancer. Int. J. Mol. Sci. 2019, 20, 3750. [Google Scholar] [CrossRef] [PubMed]
- Pachynski, R.K.; Zabel, B.A.; Kohrt, H.E.; Tejeda, N.M.; Monnier, J.; Swanson, C.D.; Holzer, A.K.; Gentles, A.J.; Sperinde, G.V.; Edalati, A.; et al. The Chemoattractant Chemerin Suppresses Melanoma by Recruiting Natural Killer Cell Antitumor Defenses. J. Exp. Med. 2012, 209, 1427–1435. [Google Scholar] [CrossRef] [PubMed]
- Haberl, E.M.; Pohl, R.; Rein-Fischboeck, L.; Feder, S.; Sinal, C.J.; Bruckmann, A.; Hoering, M.; Krautbauer, S.; Liebisch, G.; Buechler, C. Overexpression of Hepatocyte Chemerin-156 Lowers Tumor Burden in a Murine Model of Diethylnitrosamine-Induced Hepatocellular Carcinoma. Int. J. Mol. Sci. 2019, 21, 252. [Google Scholar] [CrossRef]
- Li, J.J.; Yin, H.K.; Guan, D.X.; Zhao, J.S.; Feng, Y.X.; Deng, Y.Z.; Wang, X.; Li, N.; Wang, X.F.; Cheng, S.Q.; et al. Chemerin Suppresses Hepatocellular Carcinoma Metastasis through Cmklr1-Pten-Akt Axis. Br. J. Cancer 2018, 118, 1337–1348. [Google Scholar] [CrossRef]
- Lin, Y.; Yang, X.; Liu, W.; Li, B.; Yin, W.; Shi, Y.; He, R. Chemerin Has a Protective Role in Hepatocellular Carcinoma by Inhibiting the Expression of Il-6 and Gm-Csf and Mdsc Accumulation. Oncogene 2017, 36, 3599–3608. [Google Scholar] [CrossRef]
- Toulany, J.; Parlee, S.D.; Sinal, C.J.; Slayter, K.; McNeil, S.; Goralski, K.B. Cmklr1 Activation Ex Vivo Does Not Increase Proportionally to Serum Total Chemerin in Obese Humans. Endocr. Connect. 2016, 5, 70–81. [Google Scholar] [CrossRef]
- Chang, S.S.; Eisenberg, D.; Zhao, L.; Adams, C.; Leib, R.; Morser, J.; Leung, L. Chemerin Activation in Human Obesity. Obesity (Silver Spring) 2016, 24, 1522–1529. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Chen, Y.L.; Jiang, L.; Chen, J.K. Reduced Expression of Chemerin Is Associated with a Poor Prognosis and a Lowed Infiltration of Both Dendritic Cells and Natural Killer Cells in Human Hepatocellular Carcinoma. Clin. Lab. 2011, 57, 879–885. [Google Scholar] [PubMed]
- Park, J.W.; Chen, M.; Colombo, M.; Roberts, L.R.; Schwartz, M.; Chen, P.J.; Kudo, M.; Johnson, P.; Wagner, S.; Orsini, L.S.; et al. Global Patterns of Hepatocellular Carcinoma Management from Diagnosis to Death: The Bridge Study. Liver Int. 2015, 35, 2155–2166. [Google Scholar] [CrossRef] [PubMed]
- Fingas, C.D.; Best, J.; Sowa, J.P.; Canbay, A. Epidemiology of Nonalcoholic Steatohepatitis and Hepatocellular Carcinoma. Clin. Liver Dis. (Hoboken) 2016, 8, 119–122. [Google Scholar] [CrossRef]
- Massoud, O.; Charlton, M. Nonalcoholic Fatty Liver Disease/Nonalcoholic Steatohepatitis and Hepatocellular Carcinoma. Clin. Liver Dis. 2018, 22, 201–211. [Google Scholar] [CrossRef]
- Schutte, K.; Schulz, C.; Poranzke, J.; Antweiler, K.; Bornschein, J.; Bretschneider, T.; Arend, J.; Ricke, J.; Malfertheiner, P. Characterization and Prognosis of Patients with Hepatocellular Carcinoma (Hcc) in the Non-Cirrhotic Liver. BMC Gastroenterol. 2014, 14, 117. [Google Scholar] [CrossRef]
- Fan, J.G.; Kim, S.U.; Wong, V.W. New Trends on Obesity and Nafld in Asia. J. Hepatol. 2017, 67, 862–873. [Google Scholar] [CrossRef]
- Buechler, C. Chemerin in Liver Diseases. Diabetol. Metab. Syndr. 2014, 3, 1–6. [Google Scholar]
- Docke, S.; Lock, J.F.; Birkenfeld, A.L.; Hoppe, S.; Lieske, S.; Rieger, A.; Raschzok, N.; Sauer, I.M.; Florian, S.; Osterhoff, M.A.; et al. Elevated Hepatic Chemerin Gene Expression in Progressed Human Non-Alcoholic Fatty Liver Disease. Eur. J. Endocrinol. 2013, 169, 547–557. [Google Scholar] [CrossRef]
- Pohl, R.; Haberl, E.M.; Rein-Fischboeck, L.; Zimny, S.; Neumann, M.; Aslanidis, C.; Schacherer, D.; Krautbauer, S.; Eisinger, K.; Weiss, T.S.; et al. Hepatic Chemerin Mrna Expression Is Reduced in Human Nonalcoholic Steatohepatitis. Eur. J. Clin. Investig. 2017, 47, 7–18. [Google Scholar] [CrossRef]
- Haberl, E.M.; Pohl, R.; Rein-Fischboeck, L.; Feder, S.; Sinal, C.J.; Buechler, C. Chemerin in a Mouse Model of Non-Alcoholic Steatohepatitis and Hepatocarcinogenesis. Anticancer Res. 2018, 38, 2649–2657. [Google Scholar]
- Wang, C.C.; Cheng, P.N.; Kao, J.H. Systematic Review: Chronic Viral Hepatitis and Metabolic Derangement. Aliment. Pharmacol Ther. 2020, 51, 216–230. [Google Scholar] [CrossRef]
- Feder, S.; Bruckmann, A.; McMullen, N.; Sinal, C.J.; Buechler, C. Chemerin Isoform-Specific Effects on Hepatocyte Migration and Immune Cell Inflammation. Int. J. Mol. Sci. 2020, 21, 7205. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, T.; Oppenheim, J.J. Chemokine-Like Receptor 1 (Cmklr1) and Chemokine (C-C Motif) Receptor-Like 2 (Ccrl2); Two Multifunctional Receptors with Unusual Properties. Exp. Cell Res. 2011, 317, 674–684. [Google Scholar] [CrossRef] [PubMed]
- Zabel, B.A.; Kwitniewski, M.; Banas, M.; Zabieglo, K.; Murzyn, K.; Cichy, J. Chemerin Regulation and Role in Host Defense. Am. J. Clin. Exp. Immunol. 2014, 3, 1–19. [Google Scholar] [PubMed]
- Shiota, G.; Kanki, K. Retinoids and Their Target Genes in Liver Functions and Diseases. J. Gastroenterol. Hepatol. 2013, 28 (Suppl. S1), 33–37. [Google Scholar] [CrossRef]
- Theodosiou, M.; Laudet, V.; Schubert, M. From Carrot to Clinic: An Overview of the Retinoic Acid Signaling Pathway. Cell Mol. Life Sci. 2010, 67, 1423–1445. [Google Scholar] [CrossRef]
- Cohen, J. A Power Primer. Psychol. Bull. 1992, 112, 155–159. [Google Scholar] [CrossRef]
- Kukla, M.; Adamek, B.; Waluga, M.; Zalewska-Ziob, M.; Kasperczyk, J.; Gabriel, A.; Mazur, W.; Sobala-Szczygiel, B.; Buldak, R.J.; Zajecki, W.; et al. Hepatic Chemerin and Chemokine-Like Receptor 1 Expression in Patients with Chronic Hepatitis C. Biomed. Res. Int. 2014, 2014, 517820. [Google Scholar] [CrossRef]
- Wanninger, J.; Bauer, S.; Eisinger, K.; Weiss, T.S.; Walter, R.; Hellerbrand, C.; Schaffler, A.; Higuchi, A.; Walsh, K.; Buechler, C. Adiponectin Upregulates Hepatocyte Cmklr1 Which Is Reduced in Human Fatty Liver. Mol. Cell Endocrinol. 2012, 349, 248–254. [Google Scholar] [CrossRef]
- Yao, S.; Johnson, C.; Hu, Q.; Yan, L.; Liu, B.; Ambrosone, C.B.; Wang, J.; Liu, S. Differences in Somatic Mutation Landscape of Hepatocellular Carcinoma in Asian American and European American Populations. Oncotarget 2016, 7, 40491–40499. [Google Scholar] [CrossRef] [PubMed]
- Kwiecien, K.; Brzoza, P.; Bak, M.; Majewski, P.; Skulimowska, I.; Bednarczyk, K.; Cichy, J.; Kwitniewski, M. The methylation status of the chemerin promoter region located from − 252 to + 258 bp regulates constitutive but not acute-phase cytokine-inducible chemerin expression levels. Sci Rep 2020, 10, 13702. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, L.J.; Chavan, N.R.; DeHoff, L.B.; Preston, J.D.; Maddox, H.F.; O’Brien, J.M.; Armstrong, D.A.; Marsit, C.J.; Pearson, K.J. Smoking During Pregnancy Increases Chemerin Expression in Neonatal Tissue. Exp. Physiol. 2019, 104, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Krautbauer, S.; Wanninger, J.; Eisinger, K.; Hader, Y.; Beck, M.; Kopp, A.; Schmid, A.; Weiss, T.S.; Dorn, C.; Buechler, C. Chemerin Is Highly Expressed in Hepatocytes and Is Induced in Non-Alcoholic Steatohepatitis Liver. Exp. Mol. Pathol. 2013, 95, 199–205. [Google Scholar] [CrossRef]
- Nagpal, S.; Patel, S.; Jacobe, H.; DiSepio, D.; Ghosn, C.; Malhotra, M.; Teng, M.; Duvic, M.; Chandraratna, R.A. Tazarotene-Induced Gene 2 (Tig2), a Novel Retinoid-Responsive Gene in Skin. J. Invest. Dermatol. 1997, 109, 91–95. [Google Scholar] [CrossRef]
- Zheng, Y.; Luo, S.; Wang, G.; Peng, Z.; Zeng, W.; Tan, S.; Xi, Y.; Fan, J. Downregulation of Tazarotene Induced Gene-2 (Tig2) in Skin Squamous Cell Carcinoma. Eur. J. Dermatol. 2008, 18, 638–641. [Google Scholar]
- Krautbauer, S.; Meier, E.M.; Rein-Fischboeck, L.; Pohl, R.; Weiss, T.S.; Sigruener, A.; Aslanidis, C.; Liebisch, G.; Buechler, C. Ceramide and Polyunsaturated Phospholipids Are Strongly Reduced in Human Hepatocellular Carcinoma. Biochim. Biophys. Acta 2016, 1861, 1767–1774. [Google Scholar] [CrossRef]
- Kee, K.M.; Wang, J.H.; Lin, C.Y.; Wang, C.C.; Cheng, Y.F.; Lu, S.N. Validation of the 7th Edition Tnm Staging System for Hepatocellular Carcinoma: An Analysis of 8,828 Patients in a Single Medical Center. Dig. Dis. Sci. 2013, 58, 2721–2728. [Google Scholar] [CrossRef]
- Wanninger, J.; Liebisch, G.; Eisinger, K.; Neumeier, M.; Aslanidis, C.; Voggenreiter, L.; Pohl, R.; Weiss, T.S.; Krautbauer, S.; Buechler, C. Adiponectin Isoforms Differentially Affect Gene Expression and the Lipidome of Primary Human Hepatocytes. Metabolites 2014, 4, 394–407. [Google Scholar] [CrossRef]
- Weiss, T.S.; Dayoub, R. Thy-1 (Cd90)-Positive Hepatic Progenitor Cells, Hepatoctyes, and Non-Parenchymal Liver Cells Isolated from Human Livers. Method. Mol. Biol. 2017, 1506, 75–89. [Google Scholar]
- Wanninger, J.; Neumeier, M.; Weigert, J.; Bauer, S.; Weiss, T.S.; Schaffler, A.; Krempl, C.; Bleyl, C.; Aslanidis, C.; Scholmerich, J.; et al. A Signaling Pathways. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 297, G611–G618. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. Nih Image to Imagej: 25 Years of Image Analysis. Nat. Method. 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Neumann, M.; Meier, E.M.; Rein-Fischboeck, L.; Krautbauer, S.; Eisinger, K.; Aslanidis, C.; Pohl, R.; Weiss, T.S.; Buechler, C. Chemokine-Like Receptor 1 mRNA Weakly Correlates with Non-Alcoholic Steatohepatitis Score in Male but Not Female Individuals. Int. J. Mol. Sci. 2016, 17, 1335. [Google Scholar] [CrossRef] [PubMed]
- Faul, F.; Erdfelder, E.; Lang, A.G.; Buchner, A. G*Power 3: A Flexible Statistical Power Analysis Program for the Social, Behavioral, and Biomedical Sciences. Behav. Res. Method. 2007, 39, 175–191. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patients’ Characteristics | HBV | HCV | HBV/HCV | Non-Viral |
|---|---|---|---|---|
| Number | 10 | 11 | 3 | 21 |
| Sex (male/female) | 8/2 | 8/3 | 2/1 | 21/0 |
| Type 2 diabetes | 2 | 3 | 0 | 11 |
| Age (years) | 60 (35–78) | 54 (48–71) | 61 (54–76) | 63 (47–84) |
| BMI (kg/m²) | 24.5 (18.7–29.4) | 25.2 (18.8–28.7) | 26.2 (22.4–30.0) | 27.7 (19.7–44.6) |
| AST (U/l) | 40 (19–103)**, %%% | 75 (39–151)**, && | 95 (78–200)%%% | 31.5 (14–145)20&& |
| ALT (U/l) | 40 (24–23)% | 60 (27–145) | 134 (66–167)% | 42 (23–378)20 |
| Bilirubin (mg/dl) | 0.7 (0.3–1.6) | 0.7 (0.4–3.7) | 0.5 (0.5–0.8) | 0.6 (0.2–2.5)20 |
| Steatosis Grade 0/1/2/3/nd | 7/1/2/0$ | 6/2/1/1/1 | 2/0/0/1/0 | 11/3/3/4$ |
| Fibrosis Stage 0/1/2/3/4 | 2/2/0/3/3*,$ | 0/0/0/3/8*,&&& | 0/0/1/1/1 | 8/10/1/2$, &&& |
| Grading G1/G2/nd | 1/4/5 | 0/8/3 | 0/1/2 | 3/14/4 |
| TNM Stage I/II/IIIa/ nd | 3/6/1/0 | 4/3/4/0 | 1/0/1/1 | 9/6/5/1 |
| Output of Power Calculation | Cryptic | NAFLD | HBV | HCV | HBV/HCV |
|---|---|---|---|---|---|
| Chemerin | |||||
| Noncentrality parameter δ | 2.22 | 4.61 | 3.55 | 1.44 | 1.20 |
| Critical t | 2.24 | 2.28 | 2.28 | 2.24 | 4.62 |
| Df | 9.50 | 8.54 | 8.55 | 9.50 | 1.89 |
| Power (1-β err prob) | 0.51 | 0.98 | 0.88 | 0.25 | 0.11 |
| CMKLR1 | |||||
| Noncentrality parameter δ | 4.25 | 6.33 | 1.11 | 0.60 | 3.97 |
| Critical t | 2.28 | 2.24 | 2.28 | 2.24 | 4.62 |
| Df | 8.55 | 9.50 | 8.55 | 9.50 | 1.86 |
| Power (1-β err prob) | 0.96 | 0.99 | 0.17 | 0.08 | 0.52 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haberl, E.M.; Feder, S.; Pohl, R.; Rein-Fischboeck, L.; Dürholz, K.; Eichelberger, L.; Wanninger, J.; Weiss, T.S.; Buechler, C. Chemerin Is Induced in Non-Alcoholic Fatty Liver Disease and Hepatitis B-Related Hepatocellular Carcinoma. Cancers 2020, 12, 2967. https://doi.org/10.3390/cancers12102967
Haberl EM, Feder S, Pohl R, Rein-Fischboeck L, Dürholz K, Eichelberger L, Wanninger J, Weiss TS, Buechler C. Chemerin Is Induced in Non-Alcoholic Fatty Liver Disease and Hepatitis B-Related Hepatocellular Carcinoma. Cancers. 2020; 12(10):2967. https://doi.org/10.3390/cancers12102967
Chicago/Turabian StyleHaberl, Elisabeth M., Susanne Feder, Rebekka Pohl, Lisa Rein-Fischboeck, Kerstin Dürholz, Laura Eichelberger, Josef Wanninger, Thomas S. Weiss, and Christa Buechler. 2020. "Chemerin Is Induced in Non-Alcoholic Fatty Liver Disease and Hepatitis B-Related Hepatocellular Carcinoma" Cancers 12, no. 10: 2967. https://doi.org/10.3390/cancers12102967
APA StyleHaberl, E. M., Feder, S., Pohl, R., Rein-Fischboeck, L., Dürholz, K., Eichelberger, L., Wanninger, J., Weiss, T. S., & Buechler, C. (2020). Chemerin Is Induced in Non-Alcoholic Fatty Liver Disease and Hepatitis B-Related Hepatocellular Carcinoma. Cancers, 12(10), 2967. https://doi.org/10.3390/cancers12102967