Salmonella Breaks Tumor Immune Tolerance by Downregulating Tumor Programmed Death-Ligand 1 Expression


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
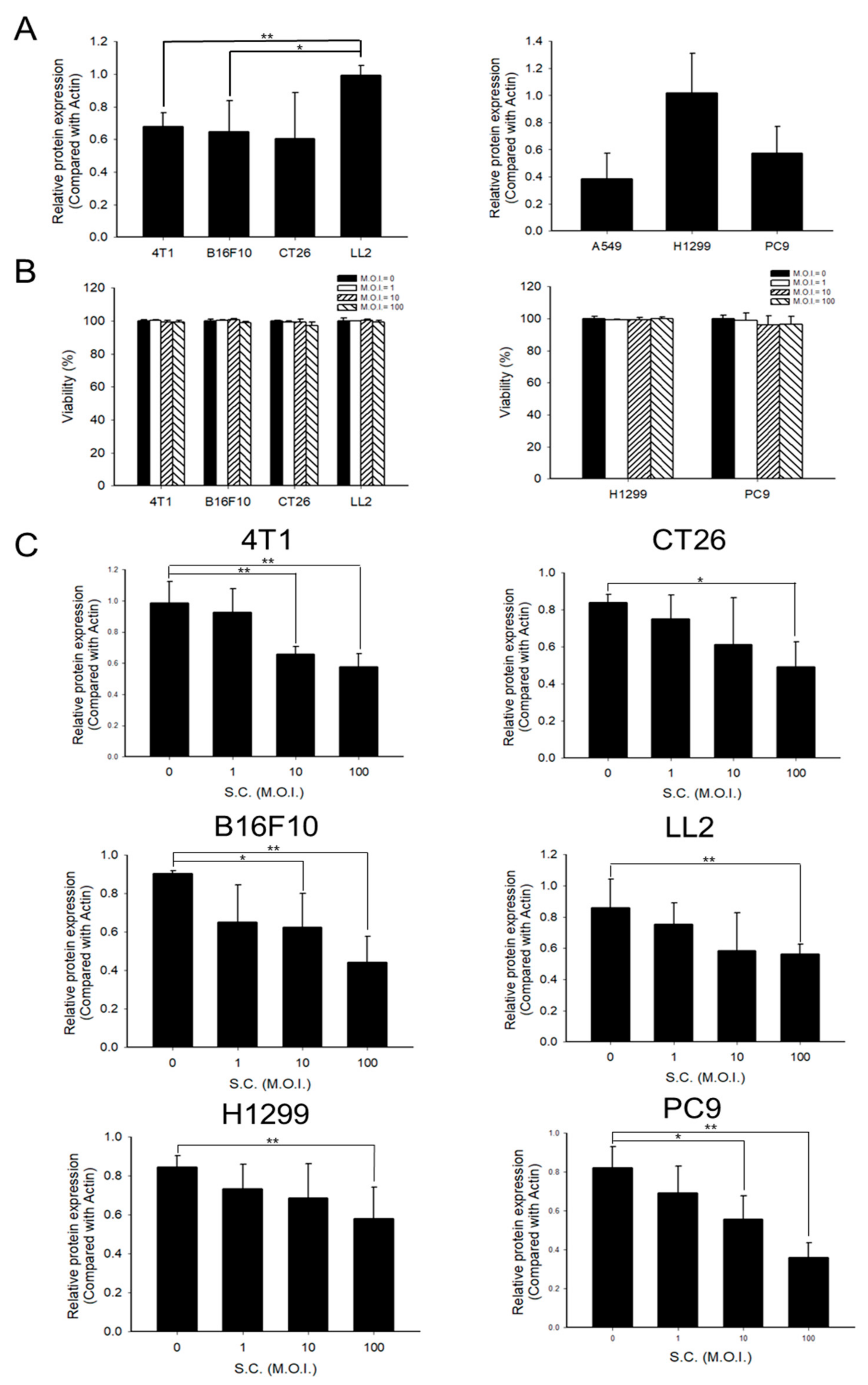
2.1. PD-L1 Was Downregulated by Salmonella
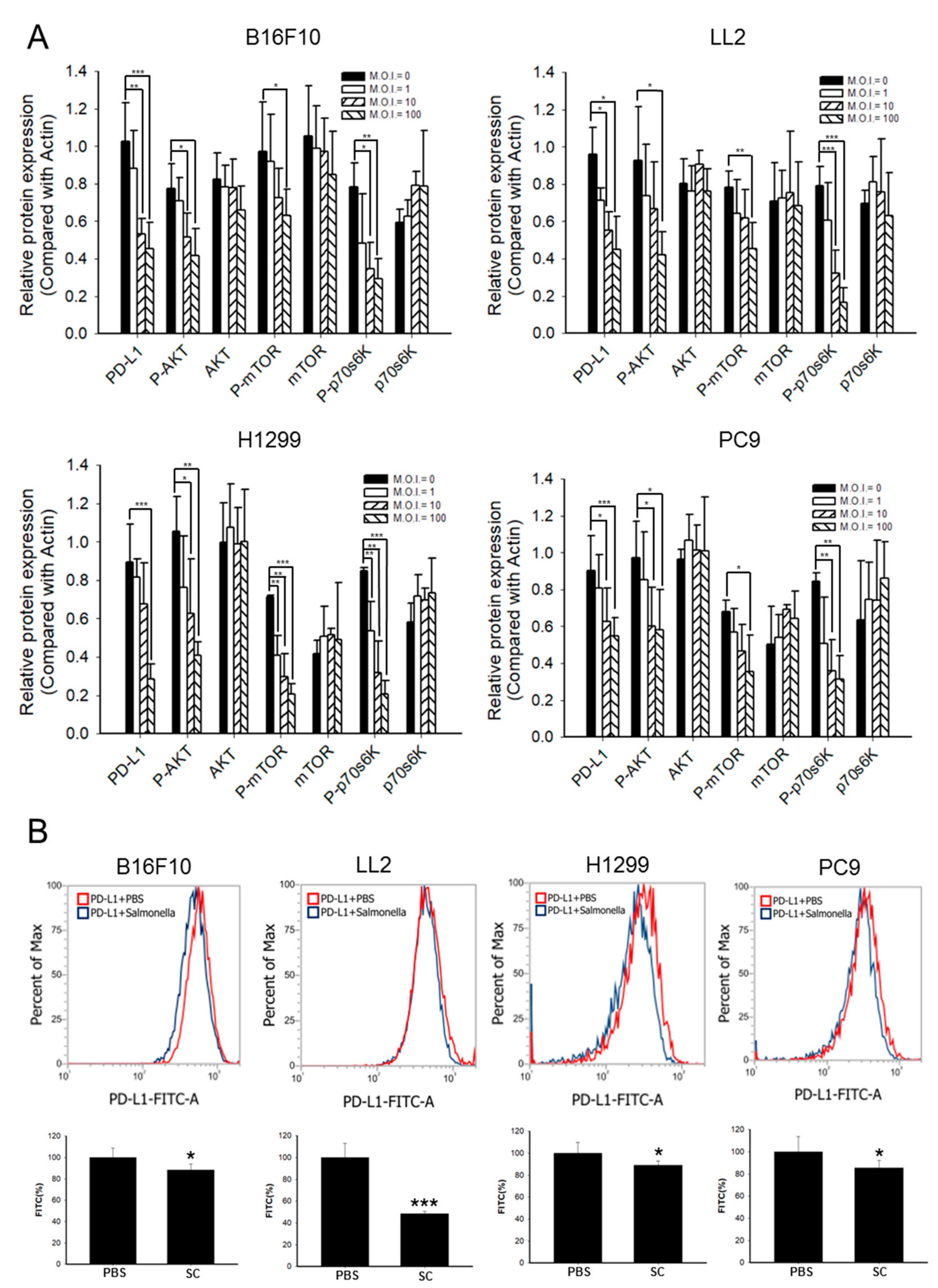
2.2. Phosph-Protein Kinase B (P-AKT)/Phosph-Mammalian Targets of Rapamycin (P-mTOR) Pathway Was Involved in Regulating PD-L1 Expression after Salmonella Treatment
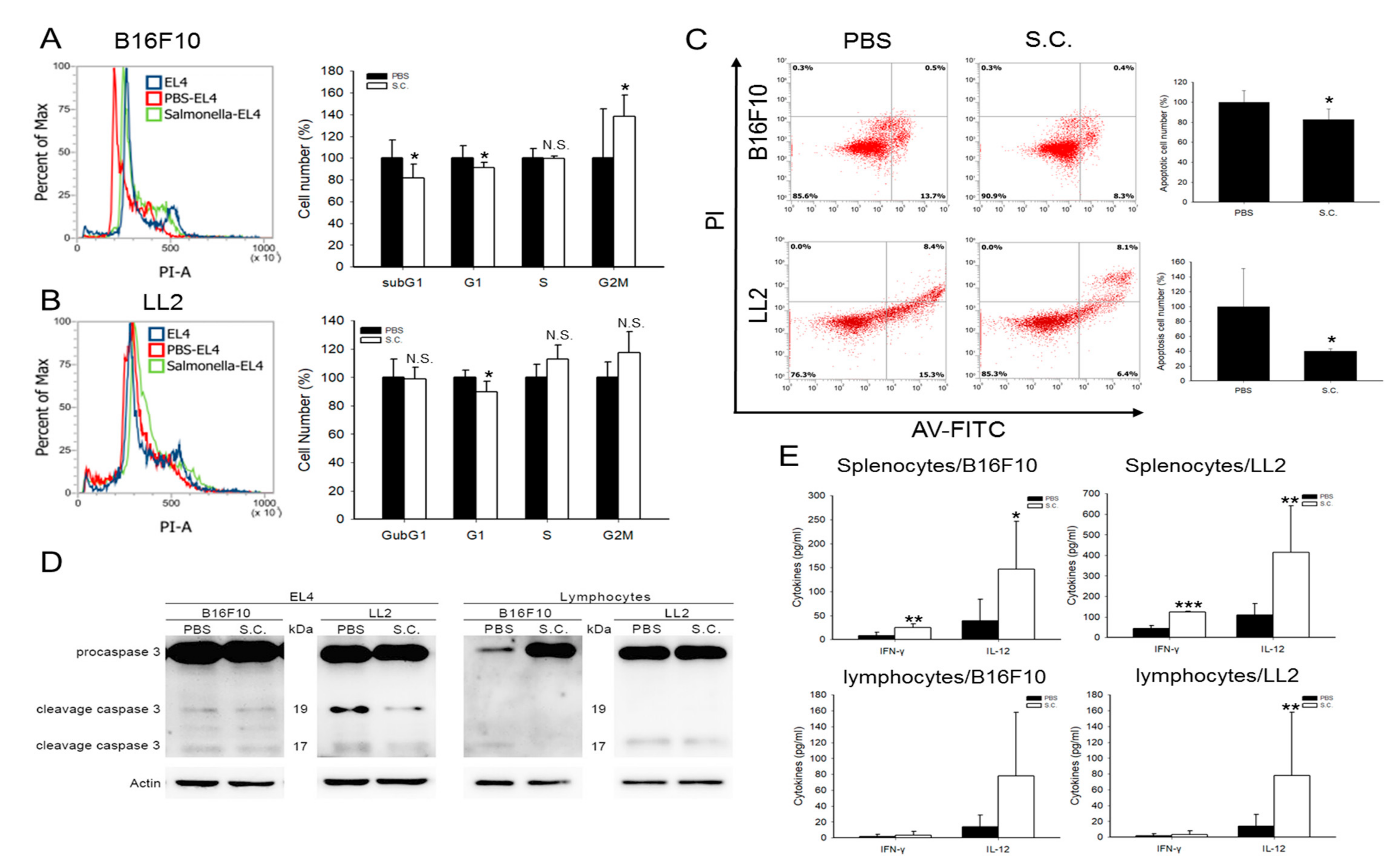
2.3. Salmonella-Treated Cells Influenced the Cell Cycle, Apoptosis, and Function of Immune Cells
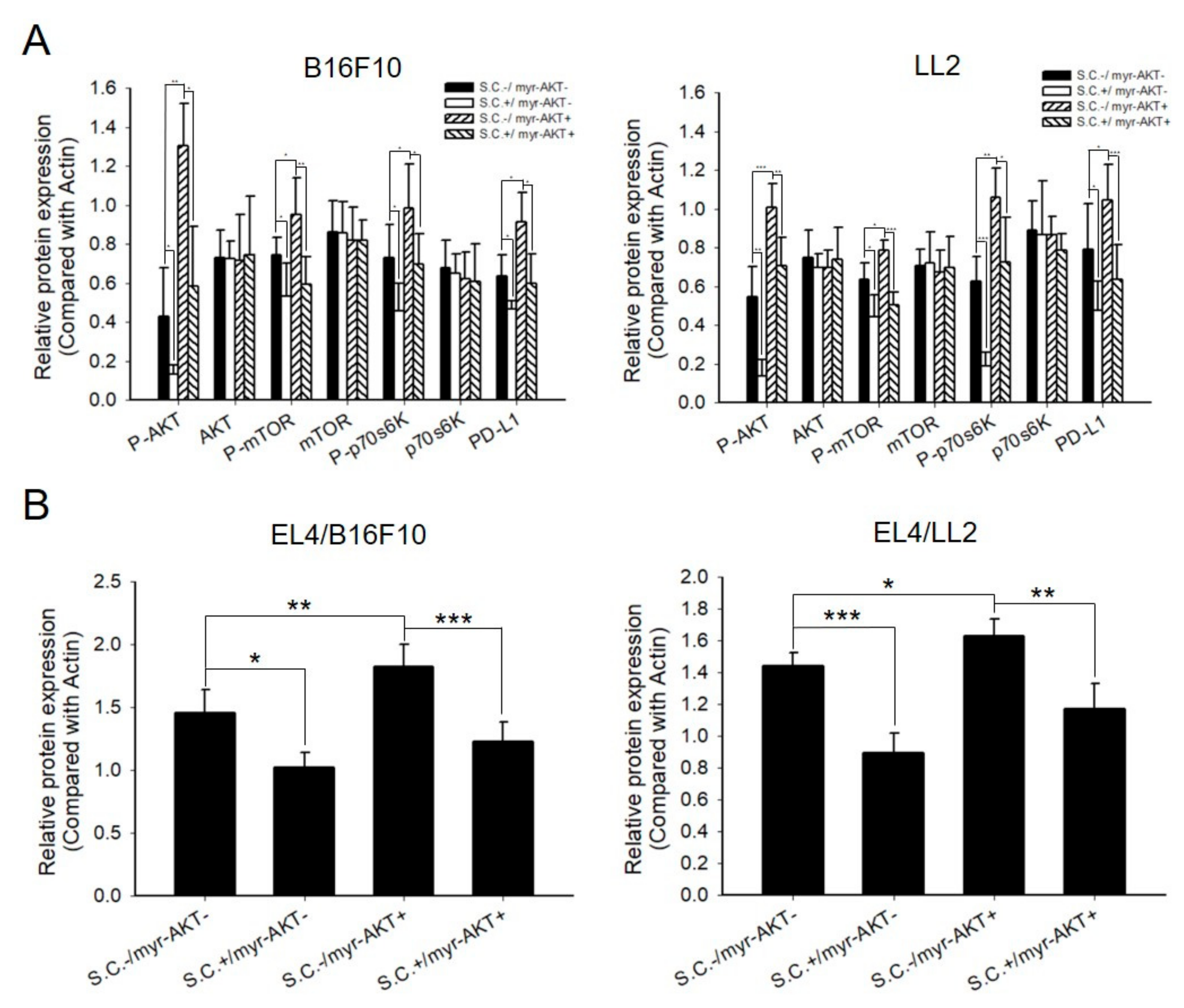
2.4. Salmonella Downregulated PD-L1 and Caspase 3 through AKT Signal Pathway in Tumor and EL4 Cells, Respectively
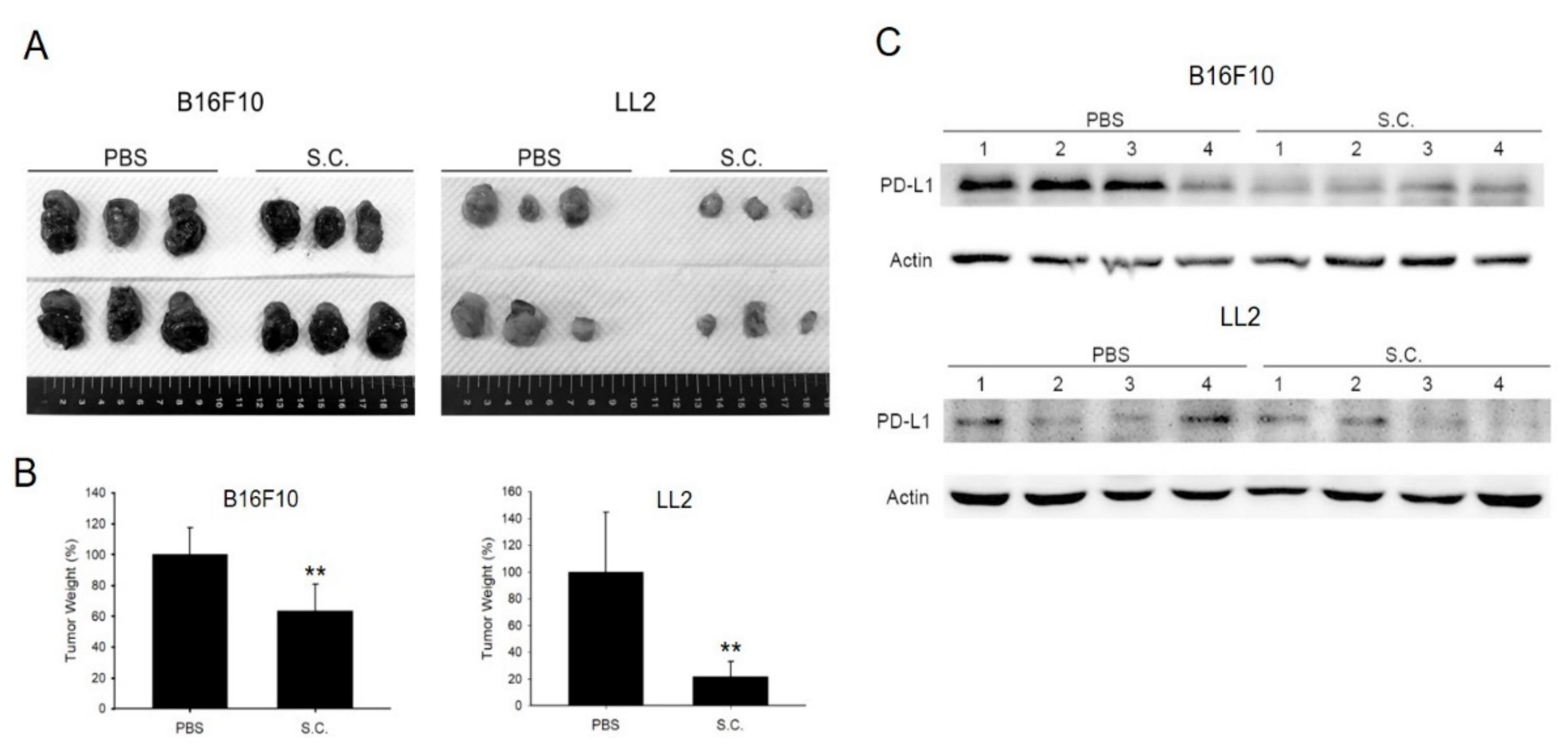
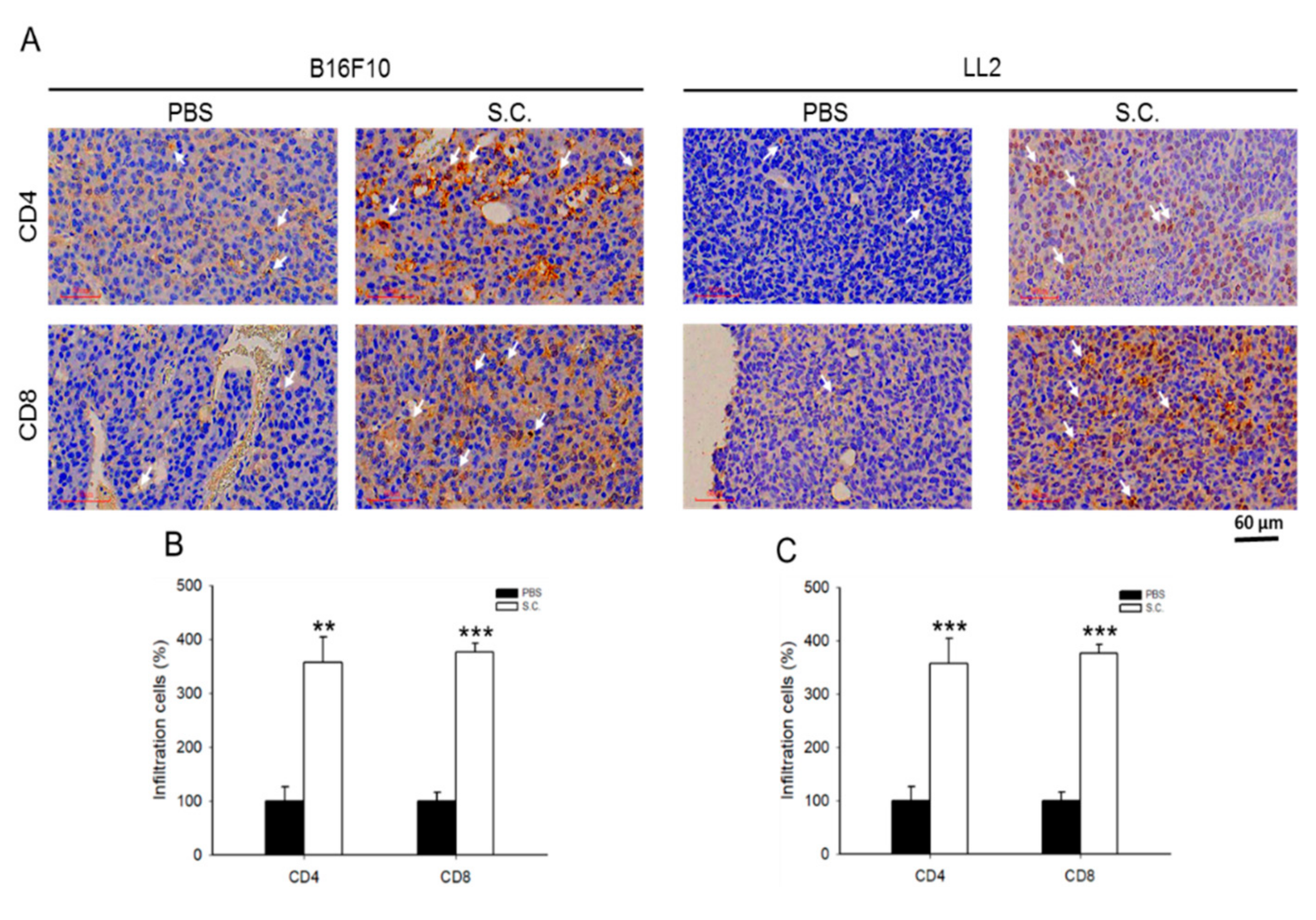
2.5. Salmonella Enhanced T Cell Infiltration and Suppresses Tumor Growth via Downregulation of PD-L1 In Vivo
3. Discussion
4. Materials and Methods
4.1. Bacteria, Cell Lines, Plasmids and Animal
4.2. Western Blot Analysis
4.3. Cell Viability Assay
4.4. Flow Cytometry
4.5. Cytokine Measurement
4.6. Animal Studies
4.7. Immunohistochemical Staining
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fan, L.; Li, Y.; Chen, J.Y.; Zheng, Y.F.; Xu, X.M. Immune Checkpoint Modulators in Cancer Immunotherapy: Recent Advances and Combination Rationales. Cancer Lett. 2019, 456, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Huang, A.C.; Zhang, W.; Zhang, G.; Wu, M.; Xu, W.; Yu, Z.; Yang, J.; Wang, B.; Sun, H.; et al. Exosomal PD-L1 Contributes to Immunosuppression and is Associated with Anti-PD-1 Response. Nature 2018, 560, 382–386. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, R.E.; Cao, X. Co-Stimulatory and Co-Inhibitory Pathways in Cancer Immunotherapy. Adv. Cancer Res. 2019, 143, 145–194. [Google Scholar] [PubMed]
- Coley, W.B. Contribution to the Knowledge of Sarcoma. Ann. Surg. 1891, 14, 199–220. [Google Scholar] [CrossRef] [PubMed]
- Silva, V.L.; Al-Jamal, W.T. Exploiting the cancer niche: Tumor-associated macrophages and hypoxia as promising synergistic targets for nano-based therapy. J. Control. Release 2017, 253, 82–96. [Google Scholar] [CrossRef] [PubMed]
- Garay, P.A.; McAllister, A.K. Novel Roles for Immune Molecules in Neural Development: Implications for Neurodevelopmental Disorders. Front. Synaptic Neurosci. 2010, 2, 136. [Google Scholar] [CrossRef] [PubMed]
- Yaghoubi, A.; Khazaei, M.; Hasanian, S.M.; Avan, A.; Cho, W.C.; Soleimanpour, S. Bacteriotherapy in Breast Cancer. Int. J. Mol. Sci. 2019, 20, 5880. [Google Scholar] [CrossRef]
- Lee, C.H.; Hsieh, J.L.; Wu, C.L.; Hsu, P.Y.; Shiau, A.L. T Cell Augments the Antitumor Activity of Tumor-Targeting Salmonella. Appl. Microbiol. Biotechnol. 2011, 90, 1381–1388. [Google Scholar] [CrossRef]
- Sivan, A.; Corrales, L.; Hubert, N.; Williams, J.B.; Aquino-Michaels, K.; Earley, Z.M.; Benyamin, F.W.; Lei, Y.M.; Jabri, B.; Alegre, M.L.; et al. Commensal Bifidobacterium Promotes Antitumor Immunity and Facilitates Anti-PD-L1 Efficacy. Science 2015, 350, 1084–1089. [Google Scholar] [CrossRef]
- Zhao, T.; Wei, T.; Guo, J.; Wang, Y.; Shi, X.; Guo, S.; Jia, X.; Jia, H.; Feng, Z. PD-1-siRNA Delivered by Attenuated Salmonella Enhances the Antimelanoma Effect of Pimozide. Cell Death Dis. 2019, 10, 164. [Google Scholar] [CrossRef]
- Pangilinan, C.R.; Lee, C.H. Salmonella-Based Targeted Cancer Therapy: Updates on A Promising and Innovative Tumor Immunotherapeutic Strategy. Biomedicines 2019, 7, 36. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Wu, C.L.; Shiau, A.L. Toll-like Receptor 4 Mediates an Antitumor Host Response Induced by Salmonella choleraesuis. Clin. Cancer Res. 2008, 14, 1905–1912. [Google Scholar] [CrossRef] [PubMed]
- Kuan, Y.D.; Lee, C.H. Salmonella Overcomes Tumor Immune Tolerance by Inhibition of Tumor Indoleamine 2, 3-Dioxygenase 1 Expression. Oncotarget 2016, 7, 374–385. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.P.; Kurzrock, R. PD-L1 Expression as a Predictive Biomarker in Cancer Immunotherapy. Mol. Cancer Ther. 2015, 14, 847–856. [Google Scholar] [CrossRef] [PubMed]
- Ingram, J.P.; Tursi, S.; Zhang, T.; Guo, W.; Yin, C.; Wynosky-Dolfi, A.M.; van der Heijden, J.; Cai, K.Q.; Yamamoto, M.; Finlay, B.B.; et al. A Nonpyroptotic IFN-γ-Triggered Cell Death Mechanism in Nonphagocytic Cells Promotes Salmonella Clearance In Vivo. J. Immunol. 2018, 200, 3626–3634. [Google Scholar] [CrossRef]
- Fresno Vara, J.A.; Casado, E.; de Castro, J.; Cejas, P.; Belda-Iniesta, C.; González-Barón, M. PI3K/Akt Signalling Pathway and Cancer. Cancer Treat. Rev. 2004, 30, 193–204. [Google Scholar] [CrossRef]
- Harvey, R.D. Immunologic and Clinical Effects of Targeting PD-1 in Lung Cancer. Clin. Pharmacol. Ther. 2014, 96, 214–223. [Google Scholar] [CrossRef]
- Oestreich, K.J.; Yoon, H.; Ahmed, R.; Boss, J.M. NFATc1 Regulates PD-1 Expression upon T Cell Activation. J. Immunol. 2008, 181, 4832–4839. [Google Scholar] [CrossRef]
- Lee, S.R.; Lo, E.H. Interactions between p38 Mitogen-Activated Protein Kinase and Caspase-3 in Cerebral Endothelial Cell Death after Hypoxia-Reoxygenation. Stroke 2003, 34, 2704–2709. [Google Scholar] [CrossRef]
- David, P.; Megger, D.A.; Kaiser, T.; Werner, T.; Liu, J.; Chen, L.; Sitek, B.; Dittmer, U.; Zelinskyy, G. The PD-1/PD-L1 Pathway Affects the Expansion and Function of Cytotoxic CD8+ T Cells During an Acute Retroviral Infection. Front. Immunol. 2019, 10, 54. [Google Scholar] [CrossRef]
- Lemiale, V.; Meert, A.P.; Vincent, F.; Darmon, M.; Bauer, P.R.; Van de Louw, A.; Azoulay, E.; Groupe de Recherche en Reanimation Respiratoire du patient d’Onco-Hématologie (Grrr-OH). Severe toxicity from checkpoint protein inhibitors: What intensive care physicians need to know? Ann. Intensive Care 2019, 9, 25. [Google Scholar] [CrossRef] [PubMed]
- Spranger, S.; Sivan, A.; Corrales, L.; Gajewski, T.F. Tumor and Host Factors Controlling Antitumor Immunity and Efficacy of Cancer Immunotherapy. Adv. Immunol. 2016, 130, 75–93. [Google Scholar] [PubMed]
- Gajewski, T.F.; Woo, S.R.; Zha, Y.; Spaapen, R.; Zheng, Y.; Corrales, L.; Spranger, S. Cancer Immunotherapy Strategies Based on Overcoming Barriers within the Tumor Microenvironment. Curr. Opin. Immunol. 2013, 25, 268–276. [Google Scholar] [CrossRef] [PubMed]
- Alsaab, H.O.; Sau, S.; Alzhrani, R.; Tatiparti, K.; Bhise, K.; Kashaw, S.K.; Iyer, A.K. PD-1 and PD-L1 Checkpoint Signaling Inhibition for Cancer Immunotherapy: Mechanism, Combinations, and Clinical Outcome. Front. Pharmacol. 2017, 8, 561. [Google Scholar] [CrossRef]
- Hendriks, D.; He, Y.; Koopmans, I.; Wiersma, V.R.; van Ginkel, R.J.; Samplonius, D.F.; Helfrich, W.; Bremer, E. Programmed Death Ligand 1 (PD-L1)-Targeted TRAIL Combines PD-L1-Mediated Checkpoint Inhibition with TRAIL-Mediated Apoptosis Induction. Oncoimmunology 2016, 5, e1202390. [Google Scholar] [CrossRef]
- Rozali, E.N.; Hato, S.V.; Robinson, B.W.; Lake, R.A.; Lesterhuis, W.J. Programmed Death Ligand 2 in Cancer-Induced Immune Suppression. Clin. Dev. Immunol. 2012, 2012, 656340. [Google Scholar] [CrossRef]
- Chang, W.W.; Lee, C.H. Salmonella as an Innovative Therapeutic Antitumor Agent. Int. J. Mol. Sci. 2014, 15, 14546–14554. [Google Scholar] [CrossRef]
- Tu, D.G.; Chang, W.W.; Lin, S.T.; Kuo, C.Y.; Tsao, Y.T.; Lee, C.H. Salmonella Inhibits Tumor Angiogenesis by Downregulation of Vascular Endothelial Growth Factor. Oncotarget 2016, 7, 37513–37523. [Google Scholar] [CrossRef]
- Loke, P.; Allison, J.P. PD-L1 and PD-L2 are Differentially Regulated by Th1 and Th2 Cells. Proc. Natl. Acad. Sci. USA 2003, 100, 5336–5341. [Google Scholar] [CrossRef]
- Saleh, R.; Taha, R.Z.; Sasidharan Nair, V.; Alajez, N.M.; Elkord, E. PD-L1 Blockade by Atezolizumab Downregulates Signaling Pathways Associated with Tumor Growth, Metastasis, and Hypoxia in Human Triple Negative Breast Cancer. Cancers (Basel) 2019, 11, 1050. [Google Scholar] [CrossRef]
- Ostrand-Rosenberg, S.; Horn, L.A.; Haile, S.T. The Programmed Death-1 Immune-Suppressive Pathway: Barrier to Antitumor Immunity. J. Immunol. 2014, 193, 3835–3841. [Google Scholar] [CrossRef] [PubMed]
- Zerdes, I.; Wallerius, M.; Sifakis, E.G.; Wallmann, T.; Betts, S.; Bartish, M.; Tsesmetzis, N.; Tobin, N.P.; Coucoravas, C.; Bergh, J.; et al. STAT3 Activity Promotes Programmed-Death Ligand 1 Expression and Suppresses Immune Responses in Breast Cancer. Cancers (Basel) 2019, 11, 1479. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Ding, D.; Yan, Y.; Li, H.; Wang, B.; Ma, L.; Ye, Z.; Ma, T.; Wu, Q.; Rodrigues, D.N.; et al. Phosphorylated RB Promotes Cancer Immunity by Inhibiting NF-κB Activation and PD-L1 Expression. Mol. Cell. 2019, 73, 22–35. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Uddin, M.M.; Padmanabhan, S.; Zhu, Y.; Bu, P.; Vancura, A.; Vancurova, I. The Proto-Oncogene Bcl3 Induces Immune Checkpoint PD-L1 Expression, Mediating Proliferation of Ovarian Cancer Cells. J. Biol. Chem. 2018, 293, 15483–15496. [Google Scholar] [CrossRef] [PubMed]
- Toso, J.F.; Gill, V.J.; Hwu, P.; Marincola, F.M.; Restifo, N.P.; Schwartzentruber, D.J.; Sherry, R.M.; Topalian, S.L.; Yang, J.C.; Stock, F. Phase I Study of the Intravenous Administration of Attenuated Salmonella Typhimurium to Patients with Metastatic Melanoma. J. Clin. Oncol. 2002, 20, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Niethammer, A.G.; Lubenau, H.; Mikus, G.; Knebel, P.; Hohmann, N.; Leowardi, C.; Beckhove, P.; Akhisaroglu, M.; Ge, Y.; Springer, M.; et al. Double-Blind, Placebo-Controlled First in Human Study to Investigate an Oral Vaccine Aimed to Elicit an Immune Reaction Against the VEGF-Receptor 2 in Patients with Stage IV and Locally Advanced Pancreatic Cancer. BMC Cancer 2012, 12, 361. [Google Scholar] [CrossRef]
- Yano, S.; Takehara, K.L.; Zhao, M.; Tan, Y.; Han, Q.; Li, S.; Bouvet, M.; Fujiwara, T.; Hoffman, R.M. Tumor-Specific Cell-Cycle Decoy by Salmonella Typhimurium A1-R Combined with Tumor-Selective Cell-Cycle Trap by Methioninase Overcome Tumor Intrinsic Chemoresistance as Visualized by FUCCI Imaging. Cell Cycle 2016, 15, 1715–1723. [Google Scholar] [CrossRef]
- Murakami, T.; Hiroshima, Y.; Miyake, K.; Kiyuna, T.; Endo, I.; Zhao, M.; Hoffman, R.M. Efficacy of Tumor-Targeting Salmonella typhimurium A1-R against Malignancies in Patient-Derived Orthotopic Xenograft (PDOX) Murine Models. Cells 2019, 8, 599. [Google Scholar] [CrossRef]
- Liang, K.; Liu, Q.; Li, P.; Luo, H.; Wang, H.; Kong, Q. Genetically engineered Salmonella Typhimurium: Recent advances in cancer therapy. Cancer Lett. 2019, 448, 168–181. [Google Scholar] [CrossRef]
- Mi, Z.; Feng, Z.C.; Li, C.; Yang, X.; Ma, M.T.; Rong, P.F. Salmonella-Mediated Cancer Therapy: An Innovative Therapeutic Strategy. J. Cancer 2019, 10, 4765–4776. [Google Scholar] [CrossRef]
- Shiau, A.L.; Shen, Y.T.; Hsieh, J.L.; Wu, C.L.; Lee, C.H. Scutellaria Barbata Inhibits Angiogenesis through Downregulation of HIF-1 α in Lung Tumor. Environ. Toxicol. 2014, 29, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.K.; Chiang, W.C.; Lai, C.H.; Lee, C.H. Salmonella-Mediated Cytolethal Distending Toxin Transfer Inhibits Tumor Growth. Hum. Gene Ther. 2018, 29, 1327–1335. [Google Scholar] [CrossRef] [PubMed]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, M.-C.; Pangilinan, C.R.; Lee, C.-H. Salmonella Breaks Tumor Immune Tolerance by Downregulating Tumor Programmed Death-Ligand 1 Expression. Cancers 2020, 12, 57. https://doi.org/10.3390/cancers12010057
Chen M-C, Pangilinan CR, Lee C-H. Salmonella Breaks Tumor Immune Tolerance by Downregulating Tumor Programmed Death-Ligand 1 Expression. Cancers. 2020; 12(1):57. https://doi.org/10.3390/cancers12010057
Chicago/Turabian StyleChen, Man-Chin, Christian Ronquillo Pangilinan, and Che-Hsin Lee. 2020. "Salmonella Breaks Tumor Immune Tolerance by Downregulating Tumor Programmed Death-Ligand 1 Expression" Cancers 12, no. 1: 57. https://doi.org/10.3390/cancers12010057
APA StyleChen, M.-C., Pangilinan, C. R., & Lee, C.-H. (2020). Salmonella Breaks Tumor Immune Tolerance by Downregulating Tumor Programmed Death-Ligand 1 Expression. Cancers, 12(1), 57. https://doi.org/10.3390/cancers12010057