Loss of BAP1 Is Associated with Upregulation of the NFkB Pathway and Increased HLA Class I Expression in Uveal Melanoma

 ,
, 
Abstract
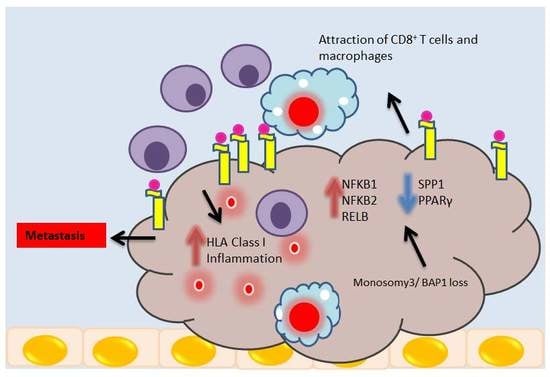
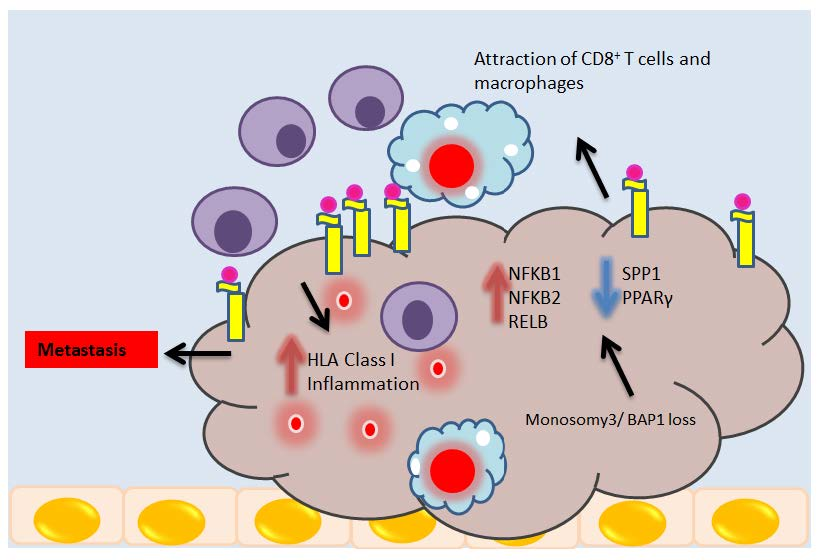
1. Introduction
2. Results
2.1. HLA Expression is Related to NFkB Pathway Molecules
2.2. NFkB Pathway is Associated with HLA Class I Regulatory Factors and the Antigen-Loading Machinery
2.3. Expression of NFkB Pathway Molecules is Related to the Presence of Infiltrating Leukocytes
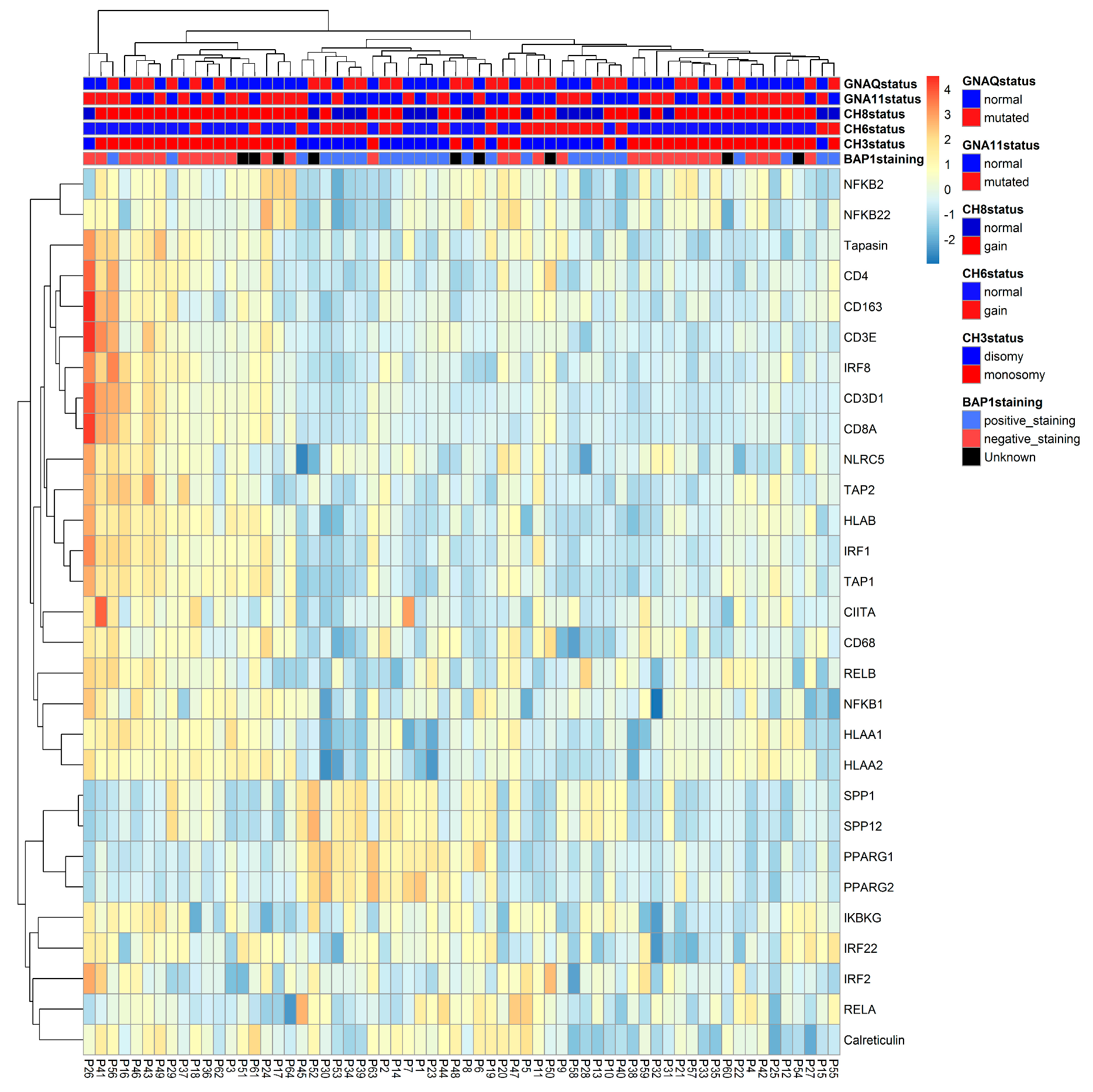
2.4. Confirmation of Associations Found by Heatmap and Survival
2.4.1. Distribution of NFkB Pathway and HLA Class I Gene Expression in Primary UM
2.4.2. Associations between Survival in UM and Chromosome 3, BAP1 and Inflammatory Factors
2.5. NFkB Pathway and Chromosome 3/BAP1 Status
2.6. NFkB Upregulation is Present in Tumor Cells
3. Discussion
4. Materials and Methods
4.1. Study Population
4.2. Immunohistochemistry and Immunofluorescence
4.3. Chromosome Analysis
4.4. Gene Expression
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Kujala, E.; Mäkitie, T.; Kivelä, T. Very long-term prognosis of patients with malignant uveal melanoma. Investig. Ophthalmol. Vis. Sci. 2003, 44, 4651–4659. [Google Scholar] [CrossRef]
- Dogrusöz, M.; Jager, M.J.; Damato, B. Uveal melanoma treatment and prognostication. Asia Pac. J. Ophthalmol. 2017, 6, 186–196. [Google Scholar] [CrossRef]
- De Waard-Siebinga, I.; Hilders, C.G.; Hansen, B.E.; van Delft, J.L.; Jager, M.J. HLA expression and tumor-infiltrating immune cells in uveal melanoma. Graefe’s Arch. Clin. Exp. Ophthalmol. 1996, 234, 34–42. [Google Scholar] [CrossRef]
- Blom, D.J.; Luyten, G.P.; Mooy, C.; Kerkvliet, S.; Zwinderman, A.H.; Jager, M.J. Human leukocyte antigen class I expression. Marker of poor prognosis in uveal melanoma. Investig. Ophthalmol. Vis. Sci. 1997, 38, 1865–1872. [Google Scholar]
- Ericsson, C.; Seregard, S.; Bartolazzi, A.; Levitskaya, E.; Ferrone, S.; Kiessling, R.; Larsson, O. Association of HLA class I and class II antigen expression and mortality in uveal melanoma. Investig. Ophthalmol. Vis. Sci. 2001, 42, 2153–2156. [Google Scholar]
- Jager, M.J.; Hurks, H.M.; Levitskaya, J.; Kiessling, R. HLA expression in uveal melanoma: There is no rule without some exception. Hum. Immunol. 2002, 63, 444–451. [Google Scholar] [CrossRef]
- Robertson, A.G.; Shih, J.; Yau, C.; Gibb, E.A.; Oba, J.; Mungall, K.L.; Hess, J.M.; Uzunangelov, V.; Walter, V.; Danilova, L.; et al. Integrative analysis identifies four molecular and clinical subsets in uveal melanoma. Cancer Cell 2017, 32, 204–220. [Google Scholar] [CrossRef]
- Maat, W.; Ly, L.V.; Jordanova, E.S.; de Wolff-Rouendaal, D.; Schalij-Delfos, N.E.; Jager, M.J. Monosomy of chromosome 3 and an inflammatory phenotype occur together in uveal melanoma. Investig. Ophthalmol. Vis. Sci. 2008, 49, 505–510. [Google Scholar] [CrossRef]
- Bronkhorst, I.H.; Vu, T.H.; Jordanova, E.S.; Luyten, G.P.; Burg, S.H.; Jager, M.J. Different subsets of tumor-infiltrating lymphocytes correlate with macrophage influx and monosomy 3 in uveal melanoma. Investig. Ophthalmol. Vis. Sci. 2012, 53, 5370–5378. [Google Scholar] [CrossRef]
- Gezgin, G.; Dogrusöz, M.; van Essen, T.H.; Kroes, W.G.; Luyten, G.P.; van der Velden, P.A.; Walter, V.; Verdijk, R.M.; van Hall, T.; van der Burg, S.H.; et al. Genetic evolution of uveal melanoma guides the development of an inflammatory microenvironment. Cancer Immunol. Immunother. 2017, 66, 903–912. [Google Scholar] [CrossRef]
- Prescher, G.; Bornfeld, N.; Becher, R. Nonrandom chromosomal abnormalities in primary uveal melanoma. J. Natl. Cancer Inst. 1990, 82, 1765–1769. [Google Scholar] [CrossRef]
- Horsthemke, B.; Prescher, G.; Becher, R.; Bornfeld, N. Loss of chromosome 3 alleles and multiplication of chromosome 8 alleles in uveal melanoma. Genes Chromosomes Cancer 1992, 4, 217–221. [Google Scholar] [CrossRef]
- Scholes, A.G.; Damato, B.E.; Nunn, J.; Hiscott, P.; Grierson, I.; Field, J.K. Monosomy 3 in uveal melanoma: Correlation with clinical and histologic predictors of survival. Investig. Ophthalmol. Vis. Sci. 2003, 44, 1008–1011. [Google Scholar] [CrossRef]
- Aalto, Y.; Eriksson, L.; Seregard, S.; Larsson, O.; Knuutila, S. Concomitant loss of chromosome 3 and whole arm losses and gains of chromosome 1, 6, or 8 in metastasizing primary uveal melanoma. Investig. Ophthalmol. Vis. Sci. 2001, 42, 313–317. [Google Scholar]
- Cassoux, N.; Rodrigues, M.J.; Plancher, C.; Asselain, B.; Levy-Gabriel, C.; Lumbroso-Le, R.L.; Piperno-Neumann, S.; Dendale, R.; Sastre, X.; Desjardins, L.; et al. Genome-wide profiling is a clinically relevant and affordable prognostic test in posterior uveal melanoma. Br. J. Ophthalmol. 2014, 98, 769–774. [Google Scholar] [CrossRef]
- Versluis, M.; de Lange, M.J.; van Pelt, S.I.; Ruivenkamp, C.A.; Kroes, W.G.; Cao, J.; Jager, M.J.; Luyten, G.P.M.; van der Velden, P.A. Digital PCR validates 8q dosage as prognostic tool in uveal melanoma. PLoS ONE 2015, 10, e0116371. [Google Scholar] [CrossRef]
- Van Gils, W.; Kilic, E.; Brüggenwirth, H.T.; Vaarwater, J.; Verbiest, M.M.; Beverloo, B.; van Til-Berg, M.E.; Paridaens, D.; Luyten, G.P.M.; de Klein, A. Regional deletion and amplification on chromosome 6 in a uveal melanoma case without abnormalities on chromosomes 1p, 3 and 8. Melanoma Res. 2008, 18, 10–15. [Google Scholar] [CrossRef]
- Damato, B.; Dopierala, J.; Klaasen, A.; van Dijk, M.; Sibbring, J.; Coupland, S.E. Multiplex ligation-dependent probe amplification of uveal melanoma: Correlation with metastatic death. Investig. Ophthalmol. Vis. Sci. 2009, 50, 3048–3055. [Google Scholar] [CrossRef]
- Van Raamsdonk, C.D.; Bezrookove, V.; Green Get Bauer, J.; Gaugler, L.; O’brien, J.M.; Simpson, E.M.; Barsh, G.S.; Bastian, B.C. Frequent somatic mutations of GNAQ in uveal melanoma and blue naevi. Nature 2009, 457, 599–602. [Google Scholar] [CrossRef]
- Van Raamsdonk, C.D.; Griewank, K.G.; Crosby, M.B.; Garrido, M.C.; Vemula, S.; Wiesner, T.; Obenauf, A.C.; Wackernagel, W.; Green, G.; Bouvier, N.; et al. Mutations in GNA11 in uveal melanoma. N. Engl. J. Med. 2010, 363, 2191–2199. [Google Scholar] [CrossRef]
- Yu, F.X.; Luo, J.; Mo, J.S.; Liu, G.; Kim, Y.C.; Meng, Z.; Zhao, L.; Peyman, G.; Ouyang, H.; Jiang, W.; et al. Mutant Gq/11 promote uveal melanoma tumorigenesis by activating YAP. Cancer Cell 2014, 25, 822–830. [Google Scholar] [CrossRef]
- Vader, M.J.; Madigan, M.C.; Versluis, M.; Suleiman, H.M.; Gezgin, G.; Gruis, N.A.; Out-Luiting, J.J.; Bergman, W.; Verdijk, R.M.; Jager, M.J.; et al. GNAQ and GNA11 mutations and downstream YAP activation in choroidal nevi. Brit. J. Cancer 2017, 117, 884–887. [Google Scholar] [CrossRef]
- Harbour, J.W.; Onken, M.D.; Roberson, E.D.; Duan, S.; Cao, L.; Worley, L.A.; Council, M.L.; Matatall, K.A.; Helms, C.; Bowcock, A.M. Frequent mutation of BAP1 in metastasizing uveal melanomas. Science 2010, 330, 1410–1413. [Google Scholar] [CrossRef]
- Brantley, M.A.; Harbour, J.W. Inactivation of retinoblastoma protein in uveal melanoma by phosphorylation of sites in the COOH-terminal region. Cancer Res. 2000, 60, 4320–4323. [Google Scholar]
- Babchia, N.; Calipel, A.; Mouriaux, F.; Faussat, A.M.; Mascarelli, F. The PI3K/Akt and mTOR/P70S6K signaling pathways in human uveal melanoma cells: Interaction with B-Raf/ERK. Investig. Ophthalmol. Vis. Sci. 2010, 51, 421–429. [Google Scholar] [CrossRef]
- Coupland, S.E.; Lake, S.L.; Zeschnigk, M.; Damato, B.E. Molecular pathology of uveal melanoma. Eye 2013, 2, 230–242. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef]
- Pires, B.R.B.; Silva, R.C.M.C.; Ferreira, G.M.; Abdelhay, E. Nf-kappab: Two sides of the same coin. Genes 2018, 9, 24. [Google Scholar] [CrossRef]
- Karin, M. NF-kB as a critical link between inflammation and cancer. Cold Spring Harb. Perspect. Biol. 2009, 1, a000141. [Google Scholar] [CrossRef]
- McNulty, S.; Tohidian, N.B.; Meyskens, F.L., Jr. RelA, p50 and inhibitor of kappa B alpha are elevated in human metastatic melanoma cells and respond aberrantly to ultraviolet light B. Pigment Cell Res. 2001, 14, 456–465. [Google Scholar] [CrossRef]
- Ueda, Y.; Richmond, A. NF-κB activation in melanoma. Pigment Cell Res. 2006, 19, 112–124. [Google Scholar] [CrossRef]
- Meir, T.; Dror, R.; Yu, X.; Qian, J.; Simon, I.; Pe’er, J.; Chowers, I. Molecular characteristics of liver metastases from uveal melanoma. Investig. Ophthalmol. Vis. Sci. 2007, 48, 4890–4896. [Google Scholar] [CrossRef]
- Dror, R.; Lederman, M.; Umezawa, K.; Barak, V.; Pe’er, J.; Chowers, I. Characterizing the Involvement of the Nuclear Factor-kappa B (NFκB) Transcription Factor in Uveal Melanoma. Investig. Ophthalmol. Vis. Sci. 2010, 51, 1811–1816. [Google Scholar] [CrossRef]
- Singh, M.K.; Singh, L.; Pushker, N.; Saini, N.; Meel, R.; Chosdol, K.; Bakhshi, S.; Sen, S.; Venkatesh, P.; Chawla, B.; et al. Identification of canonical NFκB (C-NFκB) pathway in uveal melanoma and their relation with patient outcome. Clin. Exp. Metastasis 2019, 36, 271–790. [Google Scholar] [CrossRef]
- Van Essen, T.H.; van Pelt, S.I.; Bronkhorst, I.H.; Versluis, M.; Némati, F.; Laurent, C.; Luyten, G.P.M.; van Hall, T.; van den Elsen, P.J.; van der Velden, P.A.; et al. Upregulation of HLA expression in primary uveal melanoma by infiltrating leukocytes. PLoS ONE 2016, 11, e0164292. [Google Scholar] [CrossRef]
- Stam, N.J.; Vroom, T.M.; Peters, P.J.; Pastoors, E.B.; Ploegh, H.L. HLA-A-and HLA-B-specific monoclonal antibodies reactive with free heavy chains in western blots, in formalin-fixed, paraffin-embedded tissue sections and in cryo-immuno-electron microscopy. Int. Immunol. 1990, 2, 113–125. [Google Scholar] [CrossRef]
- Krishnakumar, S.; Abhyankar, D.; Sundaram, A.L.; Pushparaj, V.; Shanmugam, M.P.; Biswas, J. Major histocompatibility antigens and antigen-processing molecules in uveal melanoma. Clin. Cancer Res. 2003, 9, 4159–4164. [Google Scholar]
- Bronkhorst, I.H.; Ly, L.V.; Jordanova, E.S.; Vrolijk, J.; Versluis, M.; Luyten, G.P.M.; Jager, M.J. Detection of M2-macrophages in uveal melanoma and relation with survival. Investig. Ophthalmol. Vis. Sci. 2011, 52, 643–650. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S. NF-κB signaling in inflammation. Signal Transduct. Target Ther. 2017, 2, 17023–17029. [Google Scholar] [CrossRef]
- Kadkol, S.S.; Lin, A.Y.; Barak, V.; Kalickman, I.; Leach, L.; Valyi-Nagy, K.; Majumdar, D.; Setty, S.; Maniotis, A.J.; Folberg, R.; et al. Osteopontin expression and serum levels in metastatic uveal melanoma: A pilot study. Investig. Ophthalmol. Vis. Sci. 2006, 47, 802–806. [Google Scholar] [CrossRef]
- Simões, C.C.; Call, M.K.; Corrêa, Z.M.; Spaulding, A.G.; Augsburger, J.J. Clinical and histopathological features and immunoreactivity of human choroidal and ciliary melanomas as prognostic factors for metastasis and death. Graefe’s Arch. Clin. Exp. Ophthalmol. 2011, 249, 1795–1803. [Google Scholar] [CrossRef]
- Onken, M.D.; Worley, L.A.; Ehlers, J.P.; Harbour, J.W. Gene expression profiling in uveal melanoma reveals two molecular classes and predicts metastatic death. Cancer Res. 2004, 64, 7205–7209. [Google Scholar] [CrossRef]
- Velho, T.R.; Kapiteijn, E.; Jager, M.J. New therapeutic agents in uveal melanoma. Anti-Cancer Res. 2012, 32, 2591–2598. [Google Scholar] [CrossRef]
- Fang, Y.; Fu, D.; Shen, X.Z. The potential role of ubiquitin c-terminal hydrolases in oncogenesis. Biochem. Biophys. Acta Rev. Cancer 2010, 1806, 1–6. [Google Scholar] [CrossRef]
- Karim, R.; Tummers, B.; Meyers, C.; Biryukov, J.L.; Alam, S.; Backendorf, C.; Jha, V.; Offringa, R.; van Ommen, G.J.; Melief, C.J.; et al. Human papillomavirus (HPV) upregulates the cellular deubiquitinase UCHL1 to suppress the keratinocyte’s innate immune response. PLoS Pathog. 2013, 9, e1003384. [Google Scholar] [CrossRef]
- Tummers, B.; Goedemans, R.; Pelascini, L.P.; Jordanova, E.S.; Van Esch, E.M.; Meyers, C.; Melief, C.J.; Boer, J.M.; van der Burg, S.H. The interferon-related developmental regulator 1 is used by human papillomavirus to suppress NFκB activation. Nat. Commun. 2015, 6, 6537–6549. [Google Scholar] [CrossRef]
- Koopmans, A.E.; Verdijk, R.M.; Brouwer, R.W.; van Den Bosch, T.P.; van Den Berg, M.M.; Vaarwater, J.; Kockx, C.E.; Paridaens, D.; Naus, N.C.; Nellist, M.; et al. Clinical significance of immunohistochemistry for detection of BAP1 mutations in uveal melanoma. Modern Pathol. 2014, 27, 1321–1330. [Google Scholar] [CrossRef]
- Van Essen, T.H.; van Pelt, S.I.; Versluis, M.; Bronkhorst, I.H.; van Duinen, S.G.; Marinkovic, M.; Kroes, W.G.; Ruivenkamp, C.A.; Shukla, S.; De Klein, A.; et al. Prognostic parameters in uveal melanoma and their association with BAP1 expression. Br. J. Ophthalmol. 2014, 98, 1738–1743. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Markers | HLA-A Probe 1 | HLA-A Probe 2 | HLA-A Probe 3 | HLA-B | ||||
|---|---|---|---|---|---|---|---|---|
| r | p | r | p | r | p | r | p | |
| NFkB1 | 0.438 | 0.001 | 0.454 | 0.001 | 0.358 | 0.04 | 0.433 | 0.001 |
| NFkB2, pr1 | 0.417 | 0.01 | 0.403 | 0.01 | 0.258 | 0.40 | 0.395 | 0.01 |
| NFkB2, pr2 | 0.239 | 0.57 | 0.285 | 0.22 | 0.171 | 1 | 0.254 | 0.43 |
| RELA | −0.055 | 1 | −0.270 | 0.31 | −0.280 | 0.25 | −0.147 | 1 |
| RELB | 0.343 | 0.05 | 0.327 | 0.08 | 0.429 | 0.001 | 0.419 | 0.01 |
| SPP1, pr1 | −0.285 | 0.23 | −0.303 | 0.15 | −0.241 | 0.55 | −0.325 | 0.09 |
| SPP1, pr2 | −0.311 | 0.12 | −0.335 | 0.07 | −0.263 | 0.36 | −0.346 | 0.05 |
| PPARγ, pr1 | −0.325 | 0.09 | −0.355 | 0.04 | −0.363 | 0.03 | −0.327 | 0.08 |
| PPARγ, pr2 | −0.294 | 0.18 | −0.338 | 0.06 | −0.327 | 0.08 | −0.265 | 0.35 |
| IkBkG | 0.188 | 1 | 0.039 | 1 | 0.250 | 0.47 | 0.235 | 0.62 |
| Markers | CIITA | NLRC5 | IRF1 | IRF2 | IRF2, pr2 | IRF8 | TAP1 | TAP2 | Tapasin | Calreticulin | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| r | p | r | p | r | p | r | p | r | p | r | p | r | p | r | p | r | p | r | p | |
| NFkB1 | 0.212 | 0.93 | 0.091 | 1 | 0.346 | 0.05 | 0.123 | 1 | 0.248 | 0.48 | 0.236 | 0.61 | 0.396 | 0.01 | 0.086 | 1 | 0.240 | 0.56 | 0.342 | 0.06 |
| NFkB2, pr1 | 0.261 | 0.37 | 0.205 | 1 | 0.327 | 0.08 | 0.143 | 1 | 0.081 | 1 | 0.242 | 0.54 | 0.409 | 0.01 | 0.052 | 1 | 0.208 | 0.99 | 0.393 | 0.01 |
| NFkB2, pr2 | 0.290 | 0.20 | 0.191 | 1 | 0.238 | 0.59 | 0.215 | 0.89 | 0.194 | 1 | 0.219 | 0.82 | 0.320 | 0.10 | 0.055 | 1 | 0.326 | 0.09 | 0.344 | 0.05 |
| RELA | −0.249 | 0.48 | −0.122 | 1 | −0.180 | 1 | 0.145 | 1 | −0.010 | 1 | −0.175 | 1 | −0.206 | 1 | 0.118 | 1 | −0.011 | 1 | 0.281 | 0.24 |
| RELB | 0.106 | 1 | 0.262 | 0.37 | 0.449 | 0.001 | 0.122 | 1 | 0.082 | 1 | 0.439 | 0.001 | 0.440 | 0.001 | 0.444 | 0.001 | 0.267 | 0.33 | 0.134 | 1 |
| SPP1, pr1 | −0.255 | 0.42 | −0.175 | 1 | −0.394 | 0.01 | −0.119 | 1 | −0.081 | 1 | −0.305 | 0.14 | −0.439 | 0.001 | −0.258 | 0.40 | −0.102 | 1 | −0.081 | 1 |
| SPP1, pr2 | −0.252 | 0.45 | −0.174 | 1 | −0.406 | 0.01 | −0.134 | 1 | −0.067 | 1 | −0.294 | 0.18 | −0.462 | 0.001 | −0.260 | 0.38 | −0.107 | 1 | −0.089 | 1 |
| PPARγ, pr1 | −0.172 | 1 | −0.229 | 0.68 | −0.372 | 0.02 | −0.062 | 1 | 0.058 | 1 | −0.394 | 0.01 | −0.443 | 0.001 | −0.299 | 0.17 | −0.259 | 0.39 | 0.147 | 1 |
| PPARγ, pr2 | −0.152 | 1 | −0.218 | 0.83 | −0.346 | 0.05 | −0.082 | 1 | −0.020 | 1 | −0.345 | 0.05 | −0.384 | 0.02 | −0.263 | 0.36 | −0.215 | 0.88 | 0.138 | 1 |
| IkBkG | 0.022 | 1 | 0.215 | 0.87 | 0.167 | 1 | 0.035 | 1 | 0.266 | 0.34 | 0.279 | 0.25 | 0.084 | 1 | 0.298 | 0.17 | 0.209 | 0.98 | 0.067 | 1 |
| Markers | CD3E | CD3D1 | CD4 | CD8A | CD68 | CD163 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| r | p | r | p | r | p | r | p | r | p | r | p | |
| NFkB1 | 0.398 | 0.01 | 0.298 | 0.17 | 0.227 | 0.71 | 0.365 | 0.03 | 0.172 | 1 | 0.009 | 1 |
| NFkB2, pr1 | 0.378 | 0.02 | 0.213 | 0.91 | 0.195 | 1 | 0.258 | 0.39 | 0.375 | 0.02 | −0.086 | 1 |
| NFkB2, pr2 | 0.361 | 0.03 | 0.242 | 0.54 | 0.278 | 0.26 | 0.267 | 0.33 | 0.411 | 0.01 | 0.040 | 1 |
| RELA | 0.110 | 1 | −0.095 | 1 | −0.119 | 1 | −0.125 | 1 | −0.062 | 1 | −0.065 | 1 |
| RELB | 0.363 | 0.03 | 0.365 | 0.03 | 0.308 | 0.13 | 0.327 | 0.08 | 0.149 | 1 | 0.317 | 0.11 |
| SPP1, pr1 | −0.295 | 0.18 | −0.247 | 0.49 | −0.337 | 0.06 | −0.280 | 0.25 | −0.442 | 0.001 | −0.265 | 0.35 |
| SPP1, pr2 | −0.297 | 0.17 | −0.251 | 0.46 | −0.330 | 0.08 | −0.281 | 0.25 | −0.447 | 0.001 | −0.243 | 0.53 |
| PPARγ, pr1 | −0.174 | 1 | −0.327 | 0.08 | −0.475 | 0.001 | −0.256 | 0.41 | −0.221 | 0.79 | −0.399 | 0.01 |
| PPARγ, pr2 | −0.195 | 1 | −0.311 | 0.12 | −0.460 | 0.001 | −0.276 | 0.27 | −0.209 | 0.97 | −0.353 | 0.04 |
| IkBkG | 0.184 | 1 | 0.196 | 1 | 0.233 | 0.64 | 0.188 | 1 | −0.009 | 1 | 0.354 | 0.04 |
| Markers | HLA-A Probe 1 | HLA-A Probe 2 | HLA-A Probe 3 | HLA-B | ||||
|---|---|---|---|---|---|---|---|---|
| r | p | r | p | r | p | r | p | |
| NFkB1 | 0.292 | 0.06 | 0.255 | 0.10 | 0.139 | 0.37 | 0.269 | 0.08 |
| NFkB2, pr1 | 0.408 | 0.007 | 0.431 | 0.004 | 0.251 | 0.11 | 0.463 | 0.002 |
| NFkB2, pr2 | 0.212 | 0.17 | 0.257 | 0.10 | 0.113 | 0.47 | 0.281 | 0.07 |
| RELA | 0.095 | 0.55 | −0.067 | 0.67 | −0.153 | 0.33 | −0.012 | 0.94 |
| RELB | 0.097 | 0.54 | 0.188 | 0.23 | 0.199 | 0.20 | 0.248 | 0.11 |
| SPP1, pr1 | −0.238 | 0.12 | −0.372 | 0.01 | −0.196 | 0.21 | −0.286 | 0.06 |
| SPP1, pr2 | −0.289 | 0.06 | −0.425 | 0.01 | −0.245 | 0.11 | −0.331 | 0.03 |
| PPARγ, pr1 | −0.354 | 0.02 | −0.431 | 0.004 | −0.445 | 0.003 | −0.320 | 0.04 |
| PPARγ, pr2 | −0.341 | 0.03 | −0.403 | 0.007 | −0.421 | 0.005 | −0.228 | 0.14 |
| IkBkG | 0.001 | 1,0 | −0.023 | 0.89 | 0.188 | 0.23 | 0.061 | 0.70 |
| Markers | All Tumors (n = 55) | Exclusion of High CD8 Tumors (n = 37) | ||||
|---|---|---|---|---|---|---|
| BAP1 + (Mean n = 25) | BAP1 − (Mean n = 30) | p | BAP1 + (Mean n = 22) | BAP1 − (Mean n = 15) | p | |
| NFKB1 | 9.19 | 9.42 | 0.018 | 9.16 | 9.26 | 0.37 |
| NFKB2-pr1 | 6.98 | 7.19 | 0.001 | 6.98 | 7.16 | 0.014 |
| NFKB2-pr2 | 6.94 | 7.14 | 0.003 | 6.94 | 7.15 | 0.01 |
| RELA | 7.13 | 7.05 | 0.12 | 7.14 | 7.09 | 0.47 |
| RELB | 6.68 | 6.73 | 0.23 | 6.66 | 6.70 | 0.48 |
| SPP1-pr1 | 10.97 | 8.56 | <0.001 | 10.91 | 8.38 | 0.001 |
| SPP1-pr2 | 10.41 | 7.97 | <0.001 | 10.34 | 7.84 | <0.001 |
| PPARγ-pr1 | 8.15 | 7.25 | <0.001 | 8.22 | 7.15 | 0.001 |
| PPARγ-pr2 | 7.00 | 6.51 | 0.001 | 7.03 | 6.46 | 0.003 |
| IkBkG | 7.94 | 7.85 | 0.085 | 7.93 | 7.78 | 0.006 |
| Characteristics | Number of Cases or Mean | % of Cases or SD | |
|---|---|---|---|
| Gender | Male | 33 | 51% |
| Female | 31 | 49% | |
| Age at enucleation (in years) | 61 | ±15.7 | |
| Cell type | Spindle | 22 | 33% |
| Mixed/epithelioid | 41 | 62% | |
| Ciliary body involvement | No | 40 | 61% |
| Yes | 24 | 36% | |
| Chromosome 3 status | Disomy | 24 | 46% |
| Monosomy | 40 | 61% | |
| BAP1 Staining | Positive | 25 | 46% |
| Negative | 30 | 55% | |
| Unknown | 9 | 16% | |
| GNA11 mutation | No | 31 | 48% |
| Yes | 33 | 52% | |
| GNAQ mutation | No | 36 | 56% |
| Yes | 28 | 44% | |
| Chromosome 6p gain | No | 43 | 65% |
| Yes | 21 | 32% | |
| Chromosome 8q gain | No | 19 | 29% |
| Yes | 45 | 68% | |
| Metastasis | No | 27 | 42% |
| Yes | 37 | 58% | |
| Follow-up time (in months) | 77 | ±55 | |
| Vital status | Dead due to UM metastasis | 37 | 58% |
| Dead due to other causes | 6 | 10% | |
| Cause of death unknown | 3 | 5% | |
| Alive at last follow-up date | 18 | 28 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Souri, Z.; Wierenga, A.P.A.; van Weeghel, C.; van der Velden, P.A.; Kroes, W.G.M.; Luyten, G.P.M.; van der Burg, S.H.; Jochemsen, A.G.; Jager, M.J. Loss of BAP1 Is Associated with Upregulation of the NFkB Pathway and Increased HLA Class I Expression in Uveal Melanoma. Cancers 2019, 11, 1102. https://doi.org/10.3390/cancers11081102
Souri Z, Wierenga APA, van Weeghel C, van der Velden PA, Kroes WGM, Luyten GPM, van der Burg SH, Jochemsen AG, Jager MJ. Loss of BAP1 Is Associated with Upregulation of the NFkB Pathway and Increased HLA Class I Expression in Uveal Melanoma. Cancers. 2019; 11(8):1102. https://doi.org/10.3390/cancers11081102
Chicago/Turabian StyleSouri, Zahra, Annemijn P. A. Wierenga, Christiaan van Weeghel, Pieter A. van der Velden, Wilma G. M. Kroes, Gregorius P. M. Luyten, Sjoerd H. van der Burg, Aart G. Jochemsen, and Martine J. Jager. 2019. "Loss of BAP1 Is Associated with Upregulation of the NFkB Pathway and Increased HLA Class I Expression in Uveal Melanoma" Cancers 11, no. 8: 1102. https://doi.org/10.3390/cancers11081102
APA StyleSouri, Z., Wierenga, A. P. A., van Weeghel, C., van der Velden, P. A., Kroes, W. G. M., Luyten, G. P. M., van der Burg, S. H., Jochemsen, A. G., & Jager, M. J. (2019). Loss of BAP1 Is Associated with Upregulation of the NFkB Pathway and Increased HLA Class I Expression in Uveal Melanoma. Cancers, 11(8), 1102. https://doi.org/10.3390/cancers11081102