An NF-kappaB- and IKK-Independent Function of NEMO Prevents Hepatocarcinogenesis by Suppressing Compensatory Liver Regeneration


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
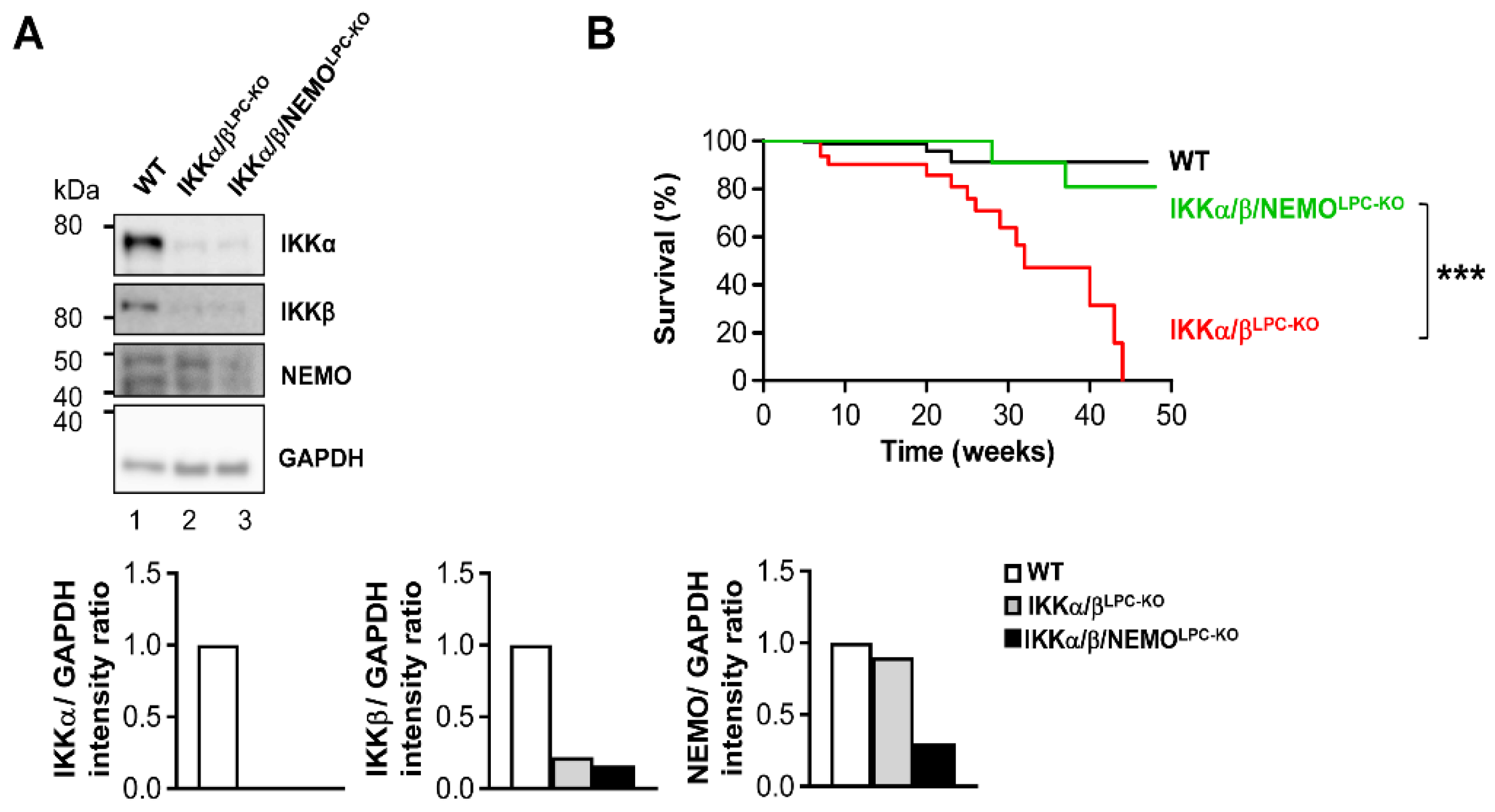
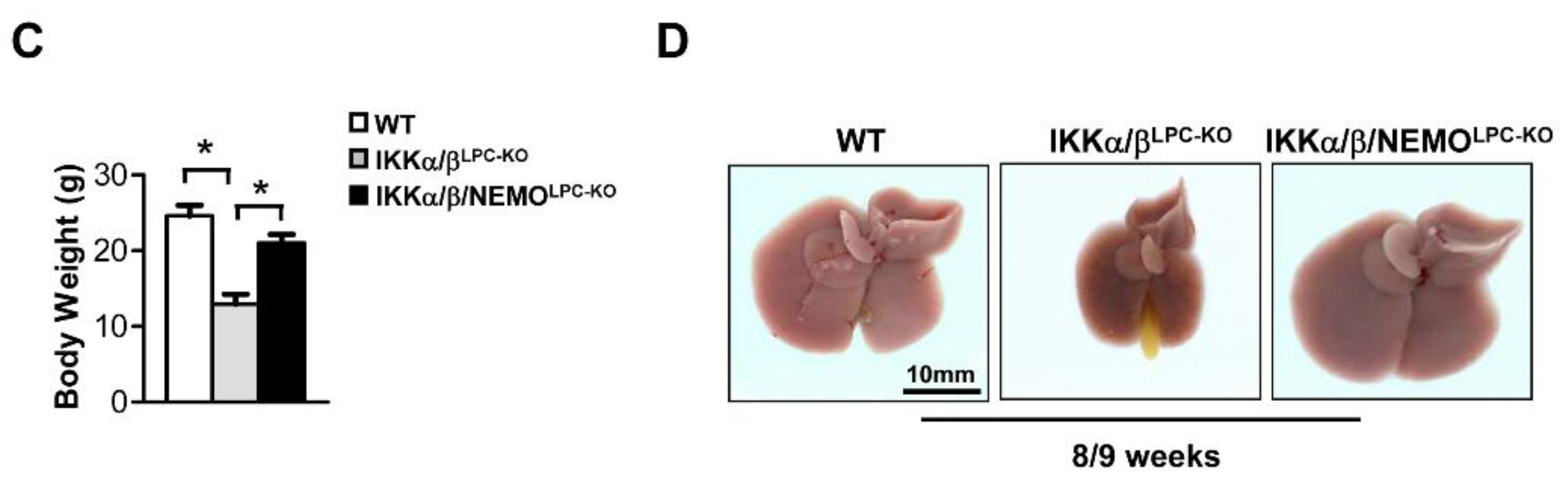
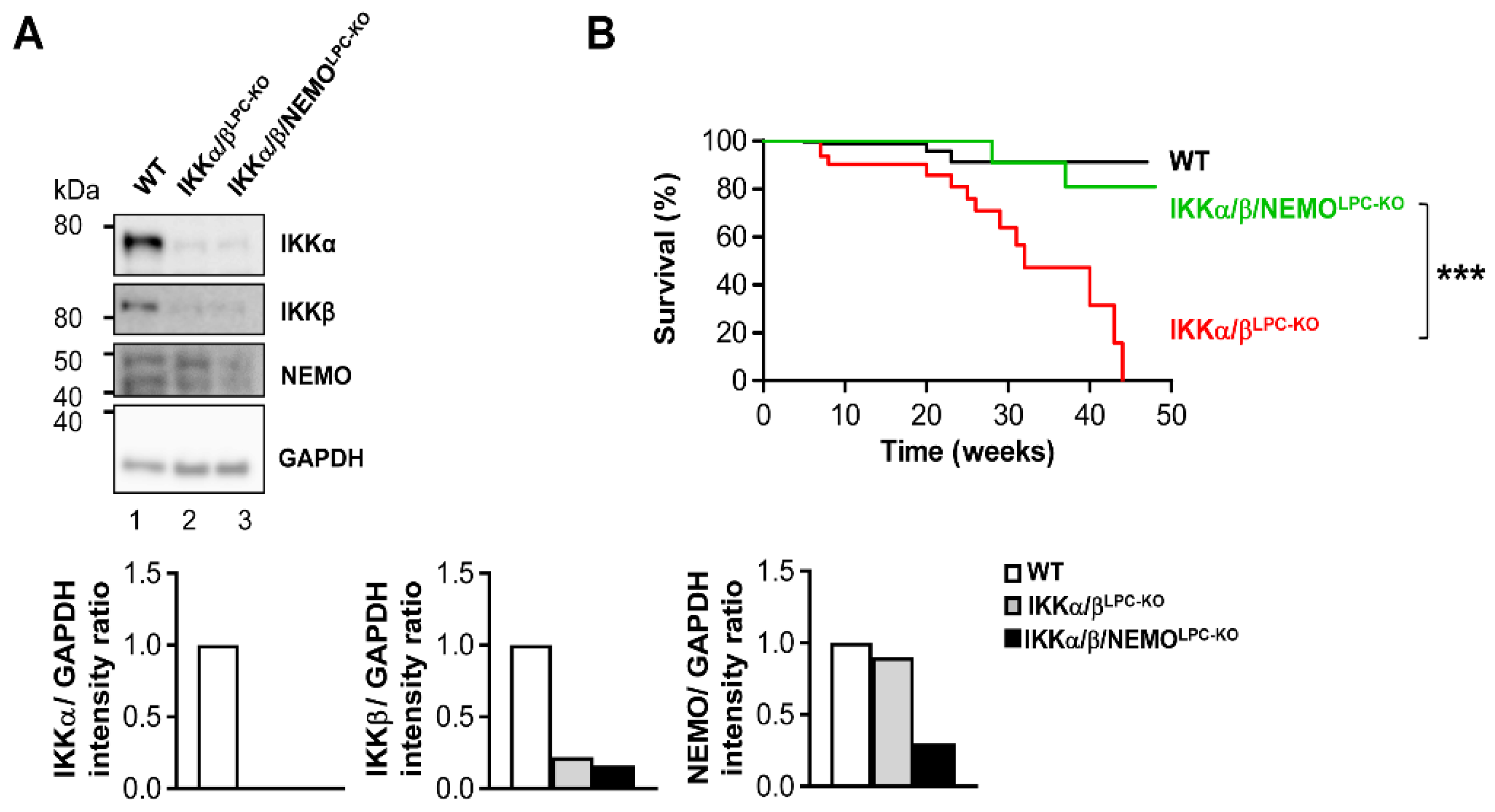
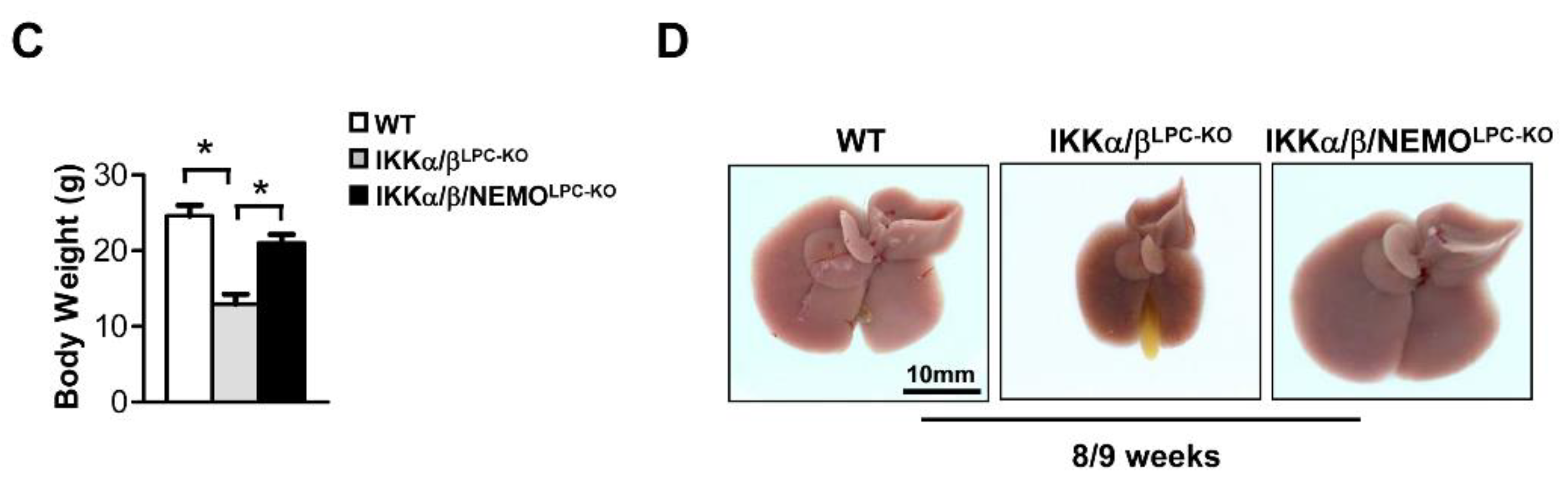
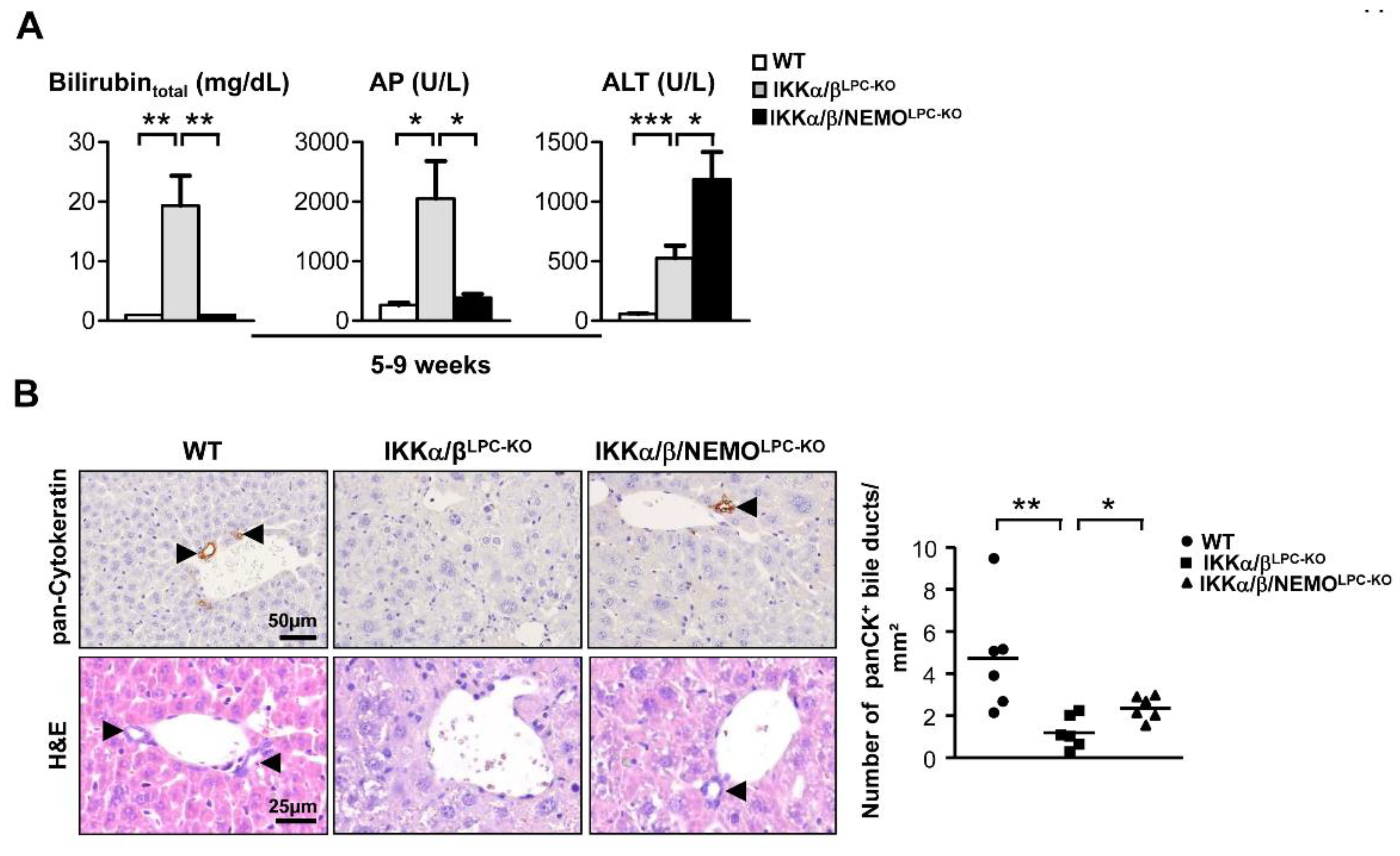
2.1. Additional Deletion of Nemo Rescues IKKα/βLPC-KO Mice from Growth Retardation and Lethal Cholestasis
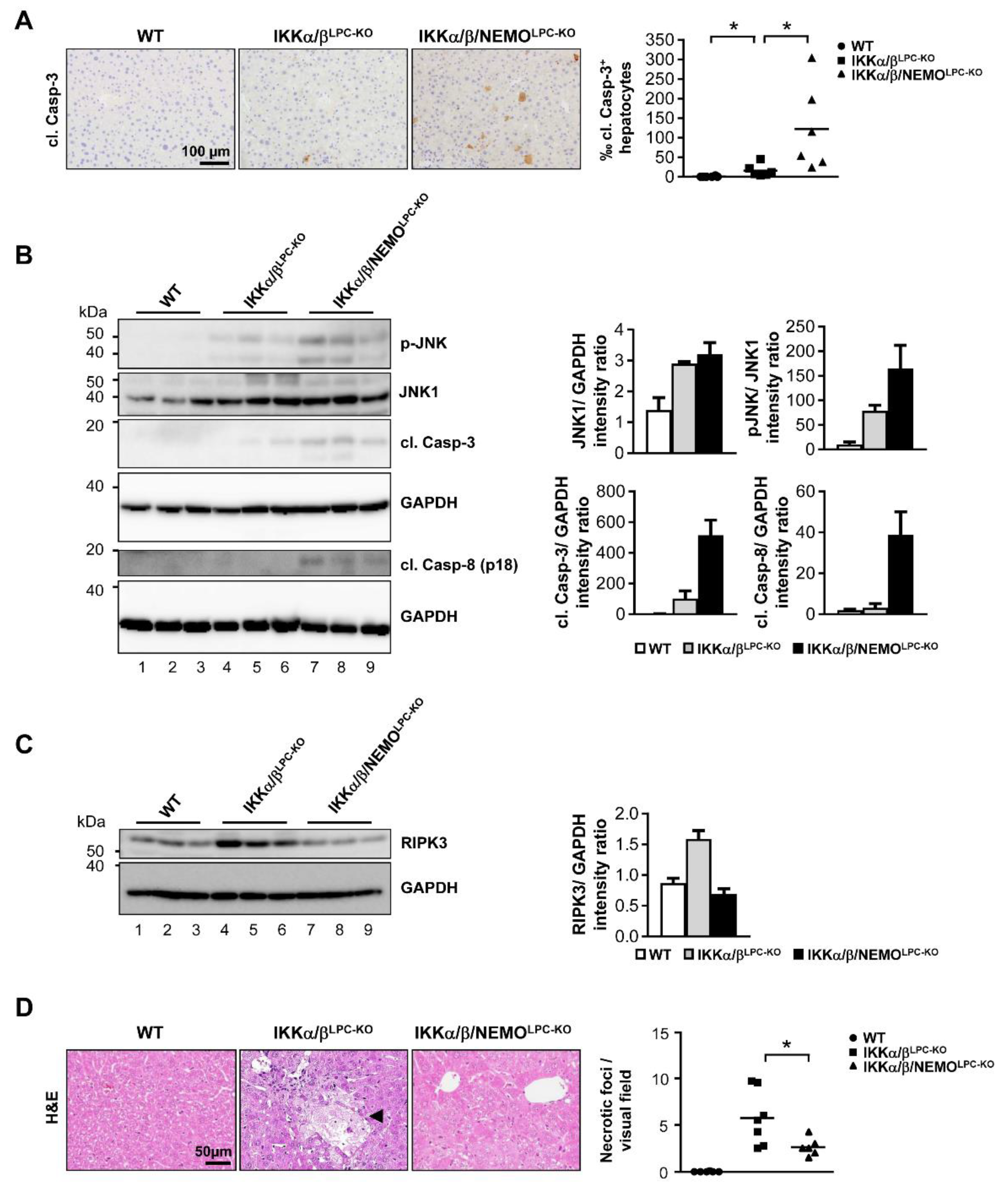
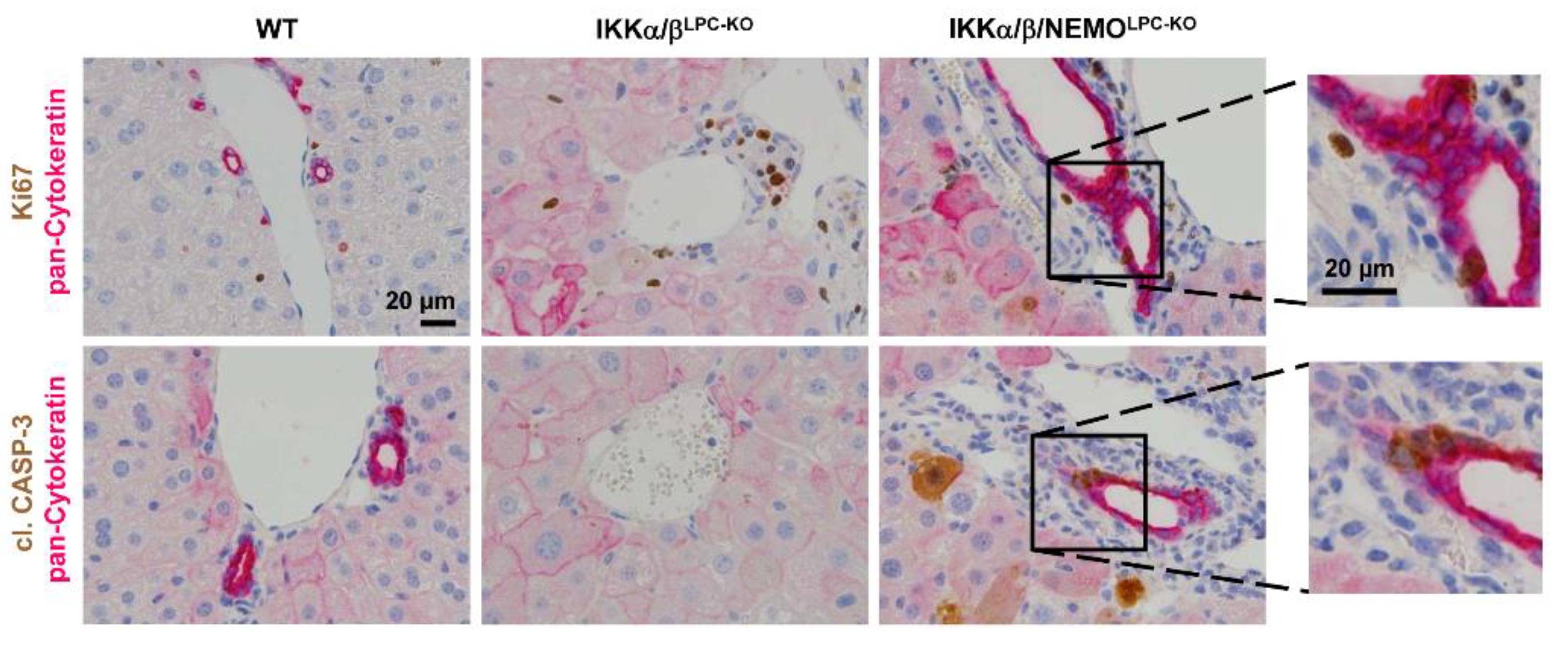
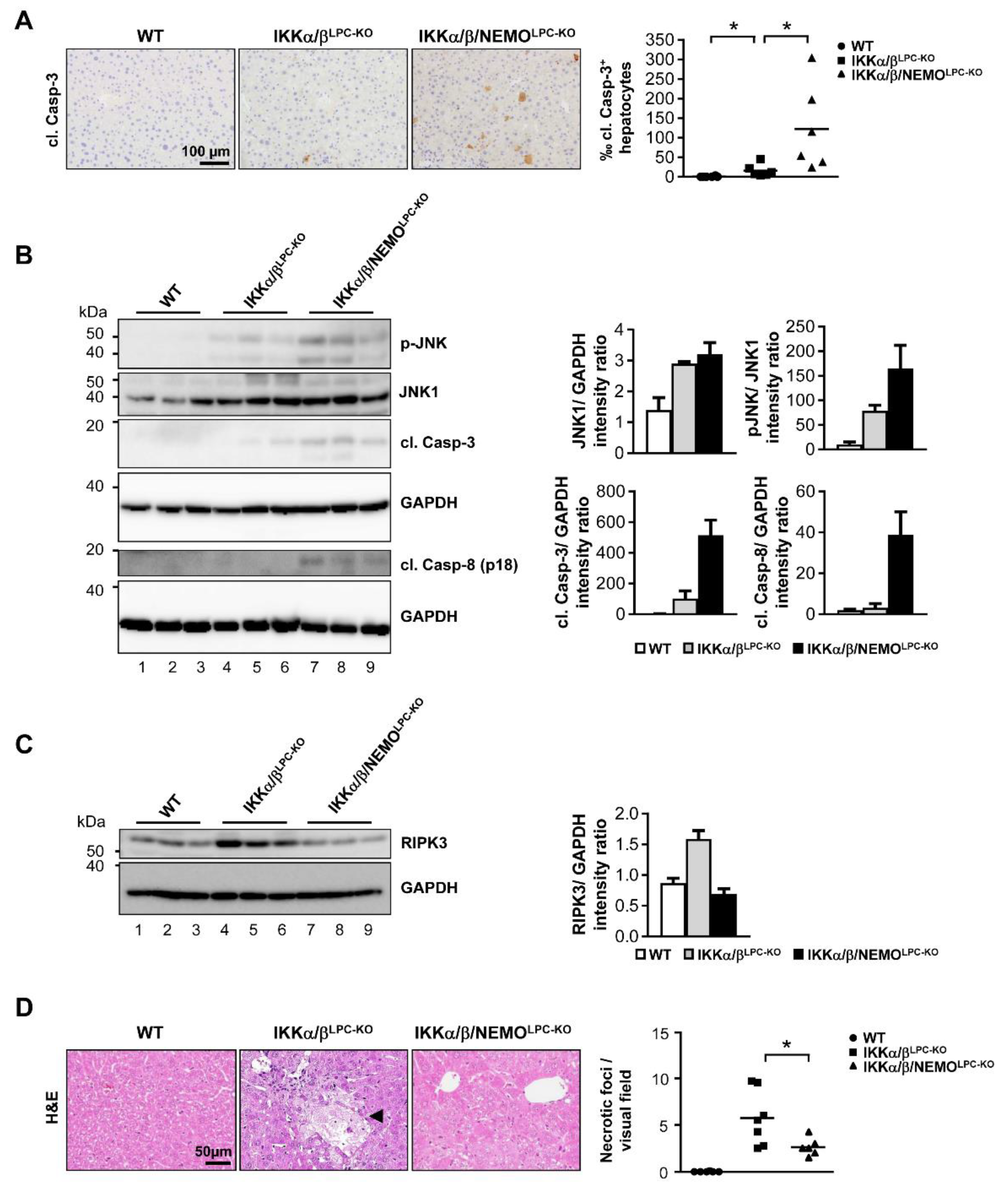
2.2. Free NEMO Molecules Correlate with Increased Necroptosis of Parenchymal Liver Cell, but Low Levels of Apoptosis
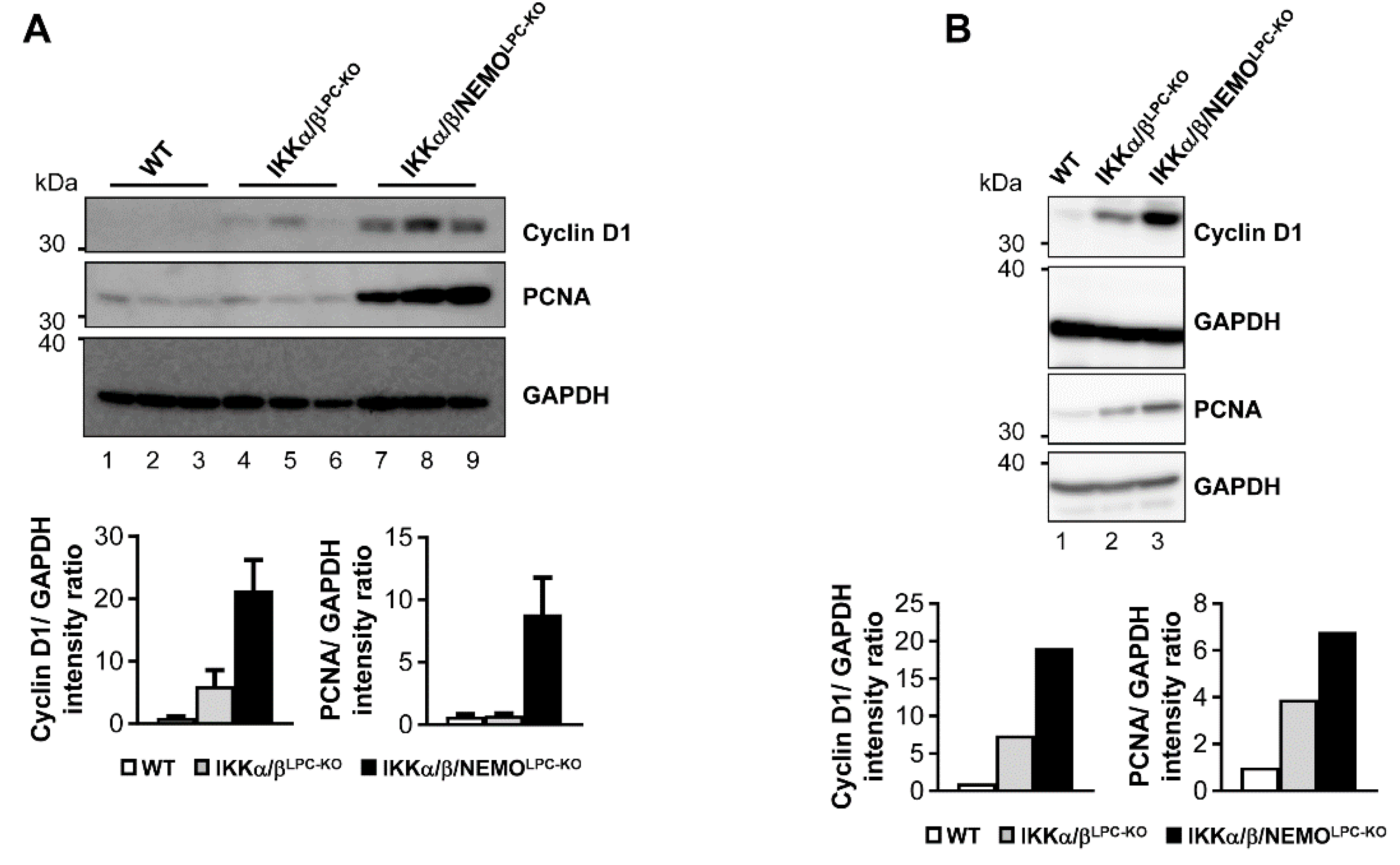
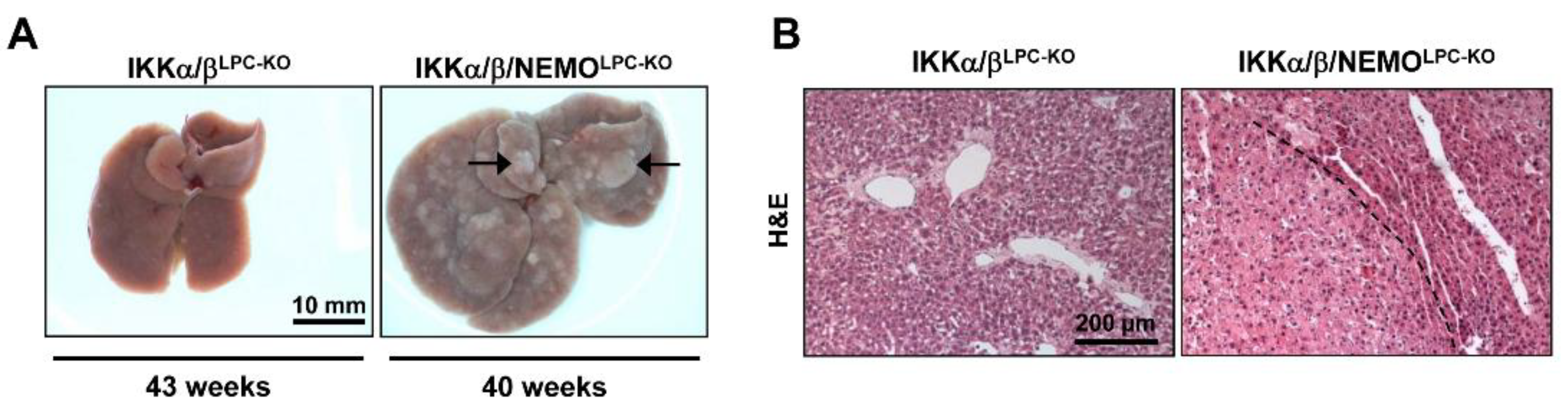
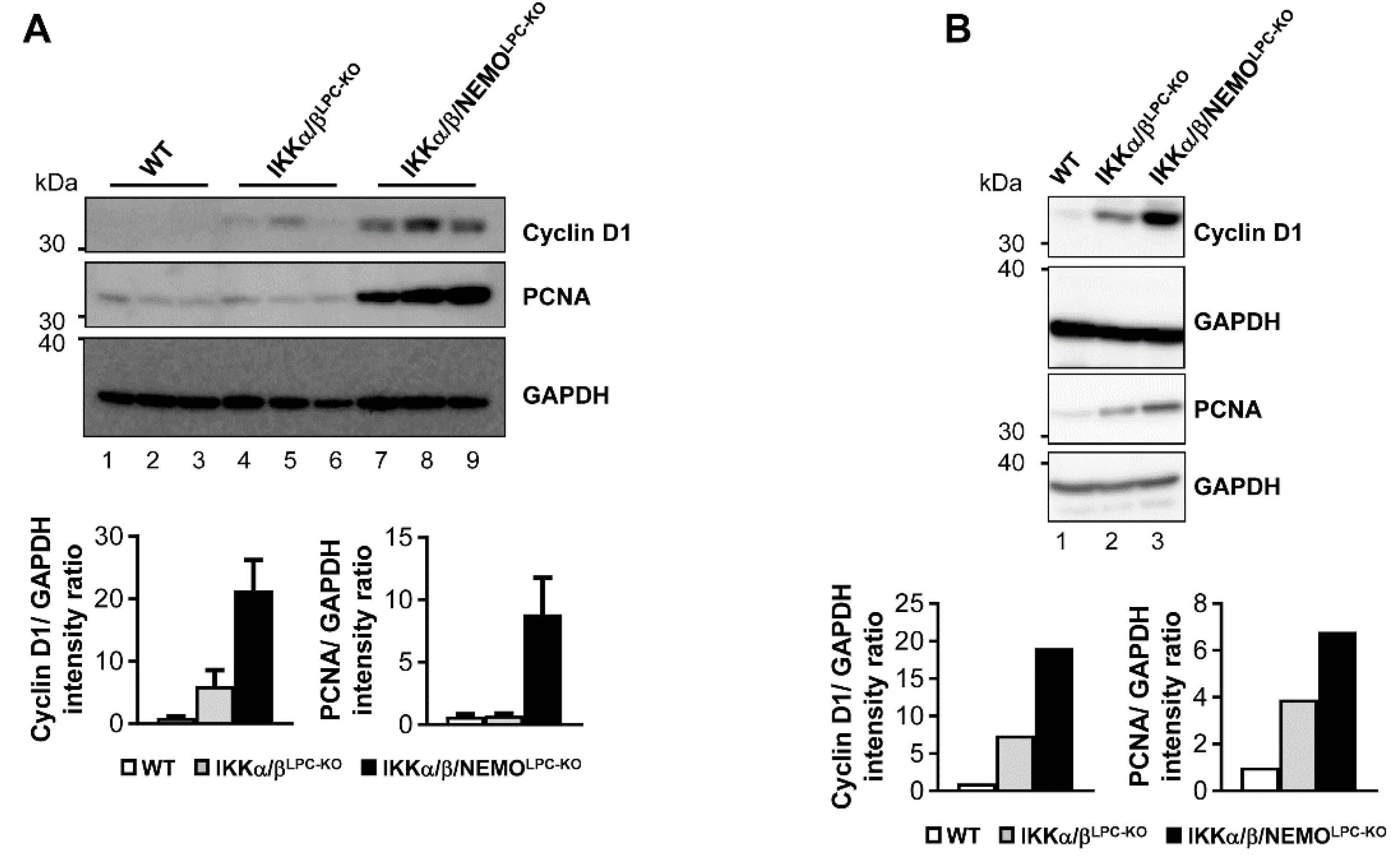
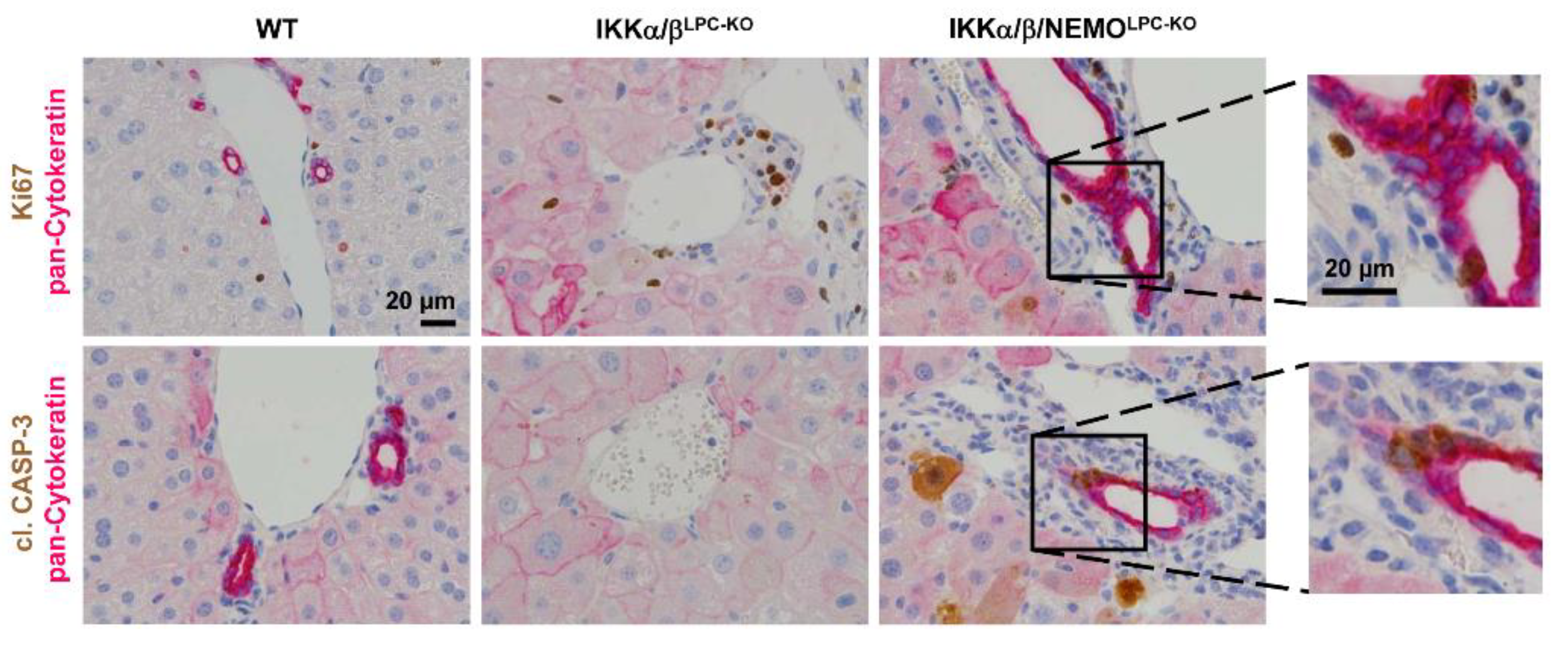
2.3. Additional Deletion of Nemo Rescues IKKα/βLPC-KO Mice from Cholestasis, but Triggers Hepatocarcinogenesis
3. Discussion
4. Methods
4.1. Generation and In Vivo Treatments of Genetically Modified Mouse Models
4.2. Isolation and Stimulation of Primary Hepatocytes
4.3. Western Blot and Immunoblot Analysis
4.4. Electrophoretic Mobility Shift Assay (EMSA)
4.5. Histological and Immunohistochemical Stainings
4.6. Serum Analysis
4.7. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgements
Conflicts of Interest
References
- Villanueva, A.; Luedde, T. The transition from inflammation to cancer in the liver. Clin. Liver Dis. 2016, 8, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Gerbes, A.; Zoulim, F.; Tilg, H.; Dufour, J.F.; Bruix, J.; Paradis, V.; Salem, R.; Peck-Radosavljevic, M.; Galle, P.R.; Greten, T.F.; et al. Gut roundtable meeting paper: Selected recent advances in hepatocellular carcinoma. Gut 2018, 67, 380–388. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.S.; Merle, P.; Granito, A.; Huang, Y.H.; Bodoky, G.; Pracht, M.; Yokosuka, O.; Rosmorduc, O.; Gerolami, R.; Caparello, C.; et al. Outcomes of sequential treatment with sorafenib followed by regorafenib for hcc: Additional analyses from the phase iii resorce trial. J. Hepatol. 2018, 69, 353–358. [Google Scholar] [CrossRef] [PubMed]
- Hacker, H.; Karin, M. Regulation and function of ikk and ikk-related kinases. Sci. STKE Signal Transduct. Knowl. Environ. 2006, 2006, re13. [Google Scholar] [CrossRef] [PubMed]
- Ben-Neriah, Y.; Karin, M. Inflammation meets cancer, with nf-kappab as the matchmaker. Nat. Immunol. 2011, 12, 715–723. [Google Scholar] [CrossRef] [PubMed]
- Luedde, T.; Heinrichsdorff, J.; De Lorenzi, R.; De Vos, R.; Roskams, T.; Pasparakis, M. Ikk1 and ikk2 cooperate to maintain bile duct integrity in the liver. Proc. Natl. Acad. Sci. USA 2008, 105, 9733–9738. [Google Scholar] [CrossRef] [PubMed]
- Luedde, T.; Beraza, N.; Kotsikoris, V.; van Loo, G.; Nenci, A.; De Vos, R.; Roskams, T.; Trautwein, C.; Pasparakis, M. Deletion of nemo/ikkgamma in liver parenchymal cells causes steatohepatitis and hepatocellular carcinoma. Cancer Cell 2007, 11, 119–132. [Google Scholar] [CrossRef] [PubMed]
- Luedde, T.; Assmus, U.; Wustefeld, T.; Meyer zu Vilsendorf, A.; Roskams, T.; Schmidt-Supprian, M.; Rajewsky, K.; Brenner, D.A.; Manns, M.P.; Pasparakis, M.; et al. Deletion of ikk2 in hepatocytes does not sensitize these cells to tnf-induced apoptosis but protects from ischemia/reperfusion injury. J. Clin. Investig. 2005, 115, 849–859. [Google Scholar] [CrossRef] [PubMed]
- Koppe, C.; Verheugd, P.; Gautheron, J.; Reisinger, F.; Kreggenwinkel, K.; Roderburg, C.; Quagliata, L.; Terracciano, L.; Gassler, N.; Tolba, R.H.; et al. Ikkalpha/beta control biliary homeostasis and hepatocarcinogenesis in mice by phosphorylating the cell-death mediator ripk1. Hepatology 2016, 64, 1217–1231. [Google Scholar] [CrossRef] [PubMed]
- Bettermann, K.; Vucur, M.; Haybaeck, J.; Koppe, C.; Janssen, J.; Heymann, F.; Weber, A.; Weiskirchen, R.; Liedtke, C.; Gassler, N.; et al. Tak1 suppresses a nemo-dependent but nf-kappab-independent pathway to liver cancer. Cancer Cell 2010, 17, 481–496. [Google Scholar] [CrossRef] [PubMed]
- Kondylis, V.; Polykratis, A.; Ehlken, H.; Ochoa-Callejero, L.; Straub, B.K.; Krishna-Subramanian, S.; Van, T.M.; Curth, H.M.; Heise, N.; Weih, F.; et al. Nemo prevents steatohepatitis and hepatocellular carcinoma by inhibiting ripk1 kinase activity-mediated hepatocyte apoptosis. Cancer Cell 2015, 28, 582–598. [Google Scholar] [CrossRef] [PubMed]
- Schwabe, R.F.; Luedde, T. Apoptosis and necroptosis in the liver: A matter of life and death. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 738–752. [Google Scholar] [CrossRef] [PubMed]
- Vucur, M.; Reisinger, F.; Gautheron, J.; Janssen, J.; Roderburg, C.; Cardenas, D.V.; Kreggenwinkel, K.; Koppe, C.; Hammerich, L.; Hakem, R.; et al. Rip3 inhibits inflammatory hepatocarcinogenesis but promotes cholestasis by controlling caspase-8- and jnk-dependent compensatory cell proliferation. Cell Rep. 2013, 4, 776–790. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, X.; Sun, T.; Jiang, J.; Li, Y.; Chen, M.; Wei, Z.; Jiang, W.; Zhou, L. Knockdown of golgi phosphoprotein 2 inhibits hepatocellular carcinoma cell proliferation and motility. Oncotarget 2016, 7, 21404–21415. [Google Scholar] [CrossRef] [PubMed]
- Dondelinger, Y.; Jouan-Lanhouet, S.; Divert, T.; Theatre, E.; Bertin, J.; Gough, P.J.; Giansanti, P.; Heck, A.J.; Dejardin, E.; Vandenabeele, P.; et al. Nf-kappab-independent role of ikkalpha/ikkbeta in preventing ripk1 kinase-dependent apoptotic and necroptotic cell death during tnf signaling. Mol. Cell 2015, 60, 63–76. [Google Scholar] [CrossRef]
- Courtois, G. The nf-kappab signaling pathway in human genetic diseases. Cell Mol. Life Sci. 2005, 62, 1682–1691. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Nishikomori, R.; Heike, T. Diagnosis and treatment in anhidrotic ectodermal dysplasia with immunodeficiency. Allergol. Int. 2012, 61, 207–217. [Google Scholar] [CrossRef]
- Aigelsreiter, A.; Haybaeck, J.; Schauer, S.; Kiesslich, T.; Bettermann, K.; Griessbacher, A.; Stojakovic, T.; Bauernhofer, T.; Samonigg, H.; Kornprat, P.; et al. Nemo expression in human hepatocellular carcinoma and its association with clinical outcome. Hum. Pathol. 2012, 43, 1012–1019. [Google Scholar] [CrossRef]
- Schneider, A.T.; Gautheron, J.; Tacke, F.; Vucur, M.; Luedde, T. Receptor interacting protein kinase-1 (ripk1) in hepatocytes does not mediate murine acetaminophen toxicity. Hepatology 2015. [Google Scholar] [CrossRef]
- Schneider, A.T.; Gautheron, J.; Feoktistova, M.; Roderburg, C.; Loosen, S.H.; Roy, S.; Benz, F.; Schemmer, P.; Buchler, M.W.; Nachbur, U.; et al. Ripk1 suppresses a traf2-dependent pathway to liver cancer. Cancer Cell 2017, 31, 94–109. [Google Scholar] [CrossRef]
- Vucur, M.; Schneider, A.T.; Gautheron, J.; Luedde, T. The enigma of ripk1 in the liver: More than just a kinase. Mol. Cell. Oncol. 2017, 4, e1304191. [Google Scholar] [CrossRef] [PubMed]
- Lafont, E.; Draber, P.; Rieser, E.; Reichert, M.; Kupka, S.; de Miguel, D.; Draberova, H.; von Massenhausen, A.; Bhamra, A.; Henderson, S.; et al. Tbk1 and ikkepsilon prevent tnf-induced cell death by ripk1 phosphorylation. Nat. Cell Biol. 2018, 20, 1389–1399. [Google Scholar] [CrossRef] [PubMed]
- Pescatore, A.; Esposito, E.; Draber, P.; Walczak, H.; Ursini, M.V. Nemo regulates a cell death switch in tnf signaling by inhibiting recruitment of ripk3 to the cell death-inducing complex ii. Cell Death Dis. 2016, 7, e2346. [Google Scholar] [CrossRef] [PubMed]
- Irrinki, K.M.; Mallilankaraman, K.; Thapa, R.J.; Chandramoorthy, H.C.; Smith, F.J.; Jog, N.R.; Gandhirajan, R.K.; Kelsen, S.G.; Houser, S.R.; May, M.J.; et al. Requirement of fadd, nemo, and bax/bak for aberrant mitochondrial function in tumor necrosis factor alpha-induced necrosis. Mol. Cell. Biol. 2011, 31, 3745–3758. [Google Scholar] [CrossRef] [PubMed]
- Luedde, T.; Schwabe, R.F. Nf-kappab in the liver--linking injury, fibrosis and hepatocellular carcinoma. Nat. Rev. Gastroenterol. Hepatol. 2011, 8, 108–118. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Xia, X.; Zhu, F.; Park, E.; Carbajal, S.; Kiguchi, K.; DiGiovanni, J.; Fischer, S.M.; Hu, Y. Ikkalpha is required to maintain skin homeostasis and prevent skin cancer. Cancer Cell 2008, 14, 212–225. [Google Scholar] [CrossRef] [PubMed]
- Maeda, S.; Kamata, H.; Luo, J.L.; Leffert, H.; Karin, M. Ikkbeta couples hepatocyte death to cytokine-driven compensatory proliferation that promotes chemical hepatocarcinogenesis. Cell 2005, 121, 977–990. [Google Scholar] [CrossRef] [PubMed]
- Kellendonk, C.; Opherk, C.; Anlag, K.; Schutz, G.; Tronche, F. Hepatocyte-specific expression of cre recombinase. Genesis 2000, 26, 151–153. [Google Scholar] [CrossRef]

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koppe, C.; Reisinger, F.; Wehr, K.; Vucur, M.; Trautwein, C.; Tacke, F.; Heikenwalder, M.; Luedde, T. An NF-kappaB- and IKK-Independent Function of NEMO Prevents Hepatocarcinogenesis by Suppressing Compensatory Liver Regeneration. Cancers 2019, 11, 999. https://doi.org/10.3390/cancers11070999
Koppe C, Reisinger F, Wehr K, Vucur M, Trautwein C, Tacke F, Heikenwalder M, Luedde T. An NF-kappaB- and IKK-Independent Function of NEMO Prevents Hepatocarcinogenesis by Suppressing Compensatory Liver Regeneration. Cancers. 2019; 11(7):999. https://doi.org/10.3390/cancers11070999
Chicago/Turabian StyleKoppe, Christiane, Florian Reisinger, Karina Wehr, Mihael Vucur, Christian Trautwein, Frank Tacke, Mathias Heikenwalder, and Tom Luedde. 2019. "An NF-kappaB- and IKK-Independent Function of NEMO Prevents Hepatocarcinogenesis by Suppressing Compensatory Liver Regeneration" Cancers 11, no. 7: 999. https://doi.org/10.3390/cancers11070999
APA StyleKoppe, C., Reisinger, F., Wehr, K., Vucur, M., Trautwein, C., Tacke, F., Heikenwalder, M., & Luedde, T. (2019). An NF-kappaB- and IKK-Independent Function of NEMO Prevents Hepatocarcinogenesis by Suppressing Compensatory Liver Regeneration. Cancers, 11(7), 999. https://doi.org/10.3390/cancers11070999