FOXK2 Transcription Factor and Its Emerging Roles in Cancer

 ,
,  and
and
Abstract
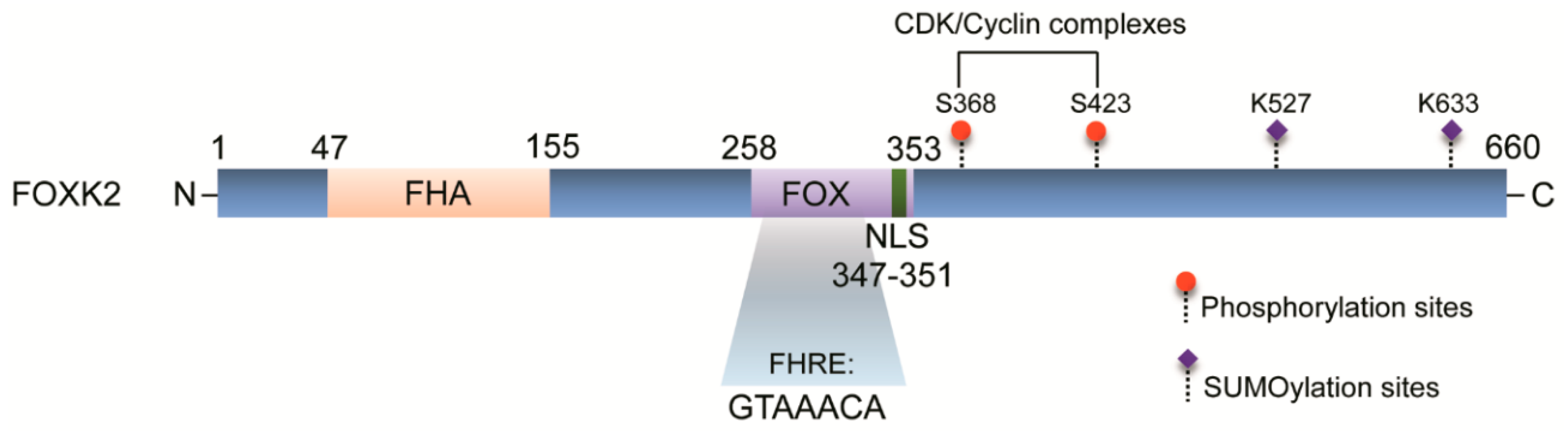
1. FOXK2 Transcription Factor
2. Mechanistic Insights into FOXK2 Transcriptional Activity
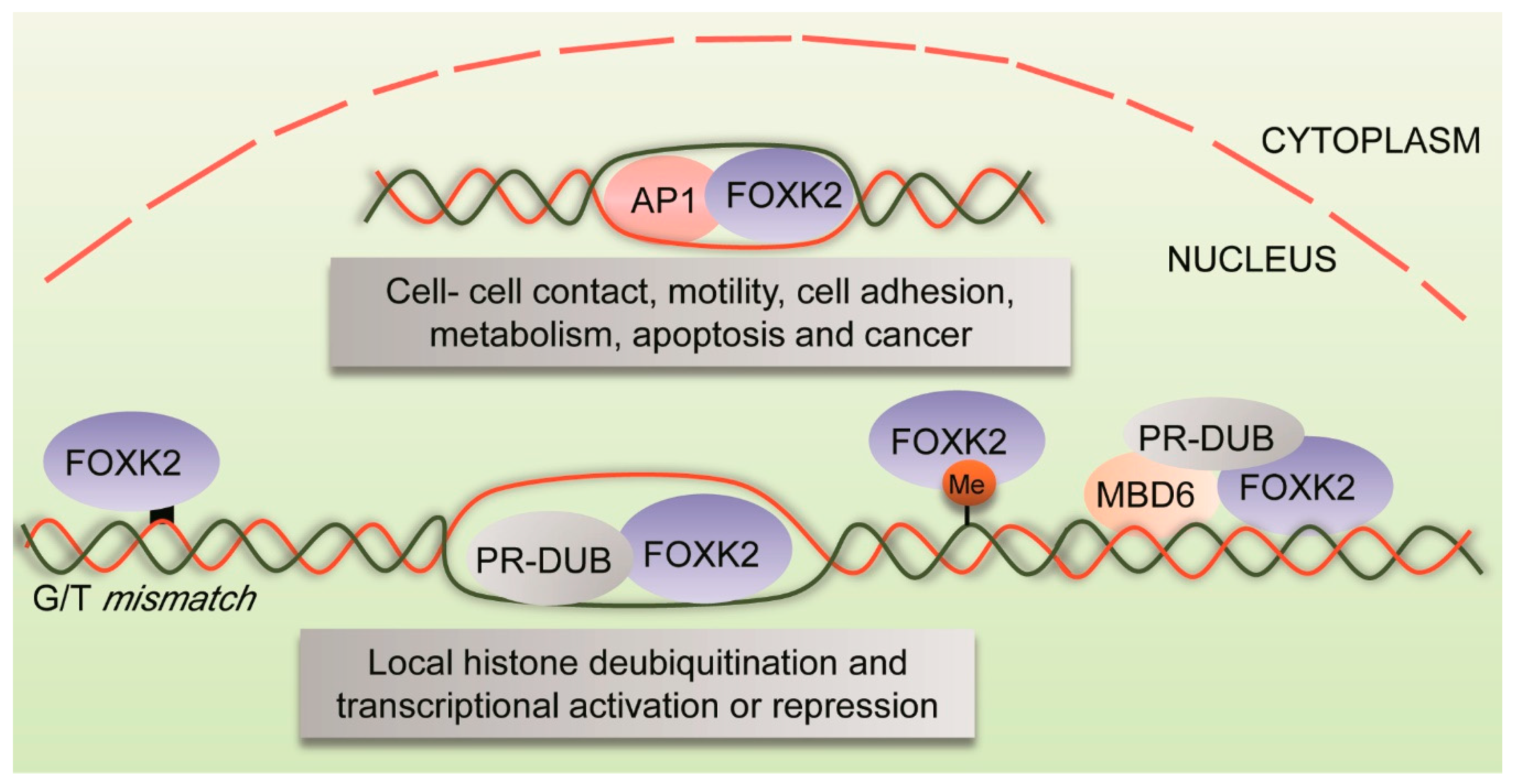
2.1. Chromatin Binding Specificities of FOXK2
2.2. FOXK2 and Histone Ubiquitination
2.3. FOXK2 and Cell Metabolism
2.3.1. FOXK2 and Autophagy
2.3.2. FOXK2 and Aerobic Glycolysis
2.4. FOXK2 and DNA Methylation
3. FOXK2 in DNA Repair and Cell Cycle Control
3.1. FOXK2 and DNA Repair
3.2. FOXK2 and Cell Cycle Regulation
4. Tumour-Specific Roles of FOXK2 as Either an Oncogene or Tumour Suppressor
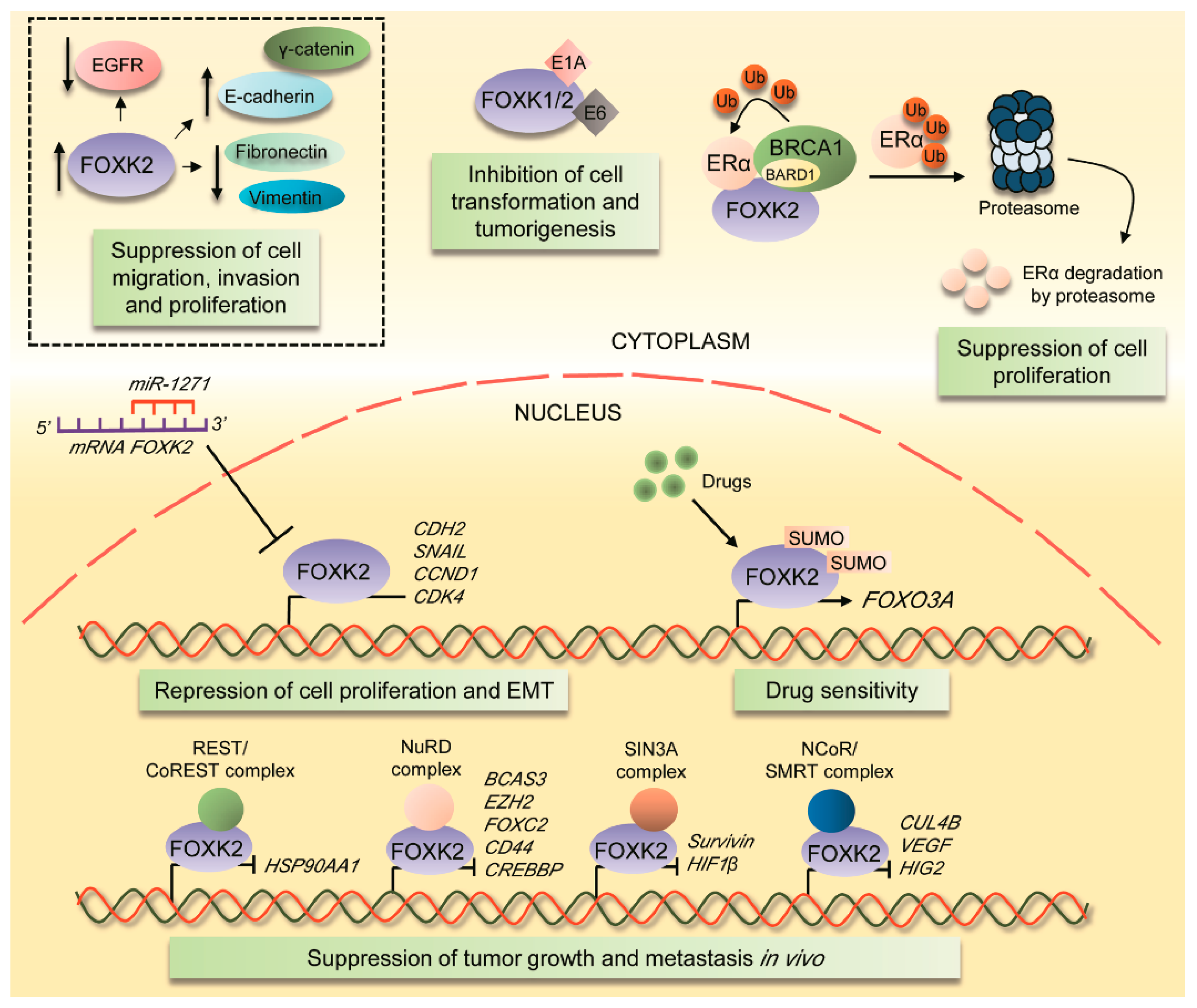
4.1. FOXK2 Role as a Tumour Suppressor
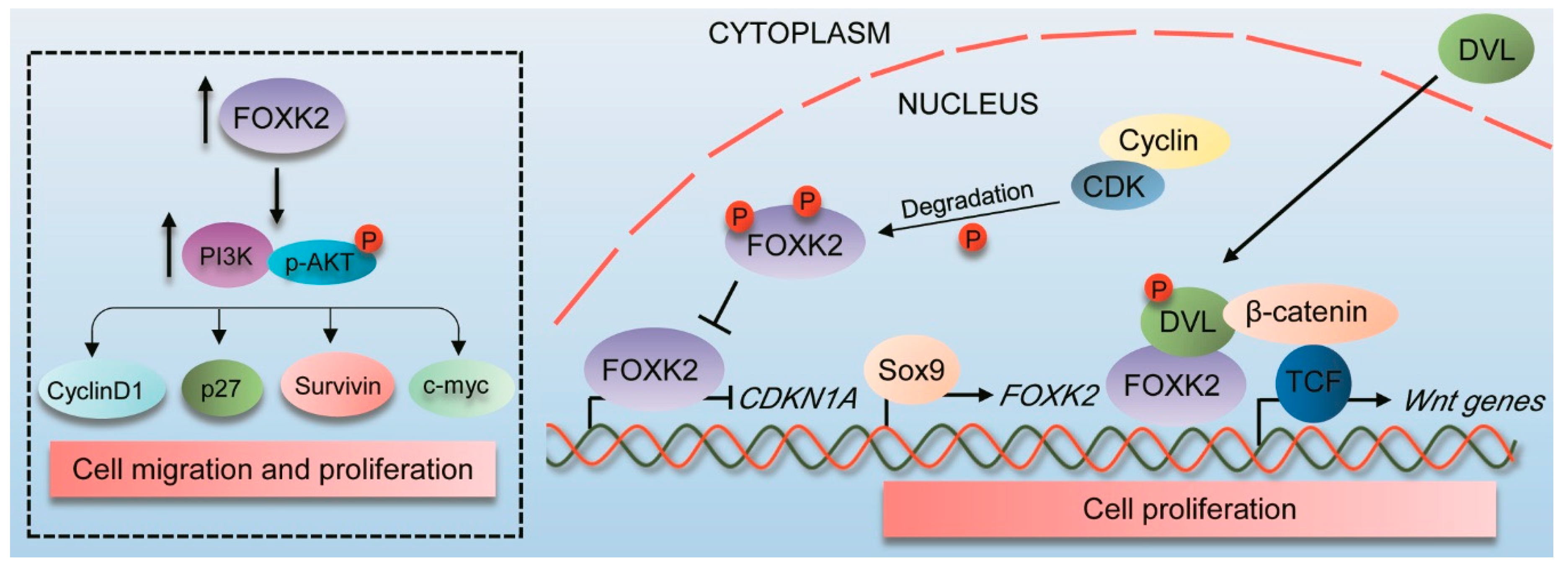
4.2. FOXK2 Role as an Oncogene
5. Prognostic Value of Assessing FOXK2 Expression in Cancer Patients
6. Concluding Remarks and Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AP-1 | activator protein-1 |
| BAP-1 | BRCA1-associated protein 1 |
| BARD-1 | BRCA1-associated RING domain protein 1 |
| BRCA-1 | Breast cancer type 1 susceptibility protein |
| ChIP | chromatin immunoprecipitation |
| EMT | epithelial-mesenchymal transition |
| ERα | estrogen receptor alpha |
| EZH2 | enhancer of zeste 2 polycomb repressive complex 2 subunit |
| FOX | forkhead BOX |
| FOXK1 | forkhead box K1 |
| FOXK2 | forkhead box K2 |
| FOXO3a | forkhead box O3 |
| FHA | forkhead associated domain |
| Gsk3 | glycogen synthase kinase 3 |
| HCC | hepatocellular carcinoma |
| HIF1α | hypoxia-inducing factor 1 alpha |
| HIV | human immunodeficiency virus |
| HDAC | histone deacetylase |
| HPV | human papillomavirus |
| IL-2 | interleukin-2 |
| ILF | interleukin-enhancer binding factor |
| MBD | methyl binding domain protein |
| mTOR | mammalian target of rapamycin |
| NCoR/SMRT | nuclear receptor co-repressor/silencing mediator for retinoid or thyroid-hormone receptors |
| NFAT | nuclear factor of activating cells |
| NLS | nuclear localization signal |
| NNMT | nicotinamide N-methyltransferase |
| NURD | nucleosome remodeling deacetylase |
| PR-DUB | Polycomb repressive deubiquitinase |
| REST | RE1-silencing transcription factor |
| SOX9 | Sex-determining region Y box 9 |
| TCGA | The Cancer Genome Atlas |
References
- Kaestner, K.H.; Knochel, W.; Martinez, D.E. Unified nomenclature for the winged helix/forkhead transcription factors. Genes Dev. 2000, 14, 142–146. [Google Scholar] [PubMed]
- Lam, E.W.; Brosens, J.J.; Gomes, A.R.; Koo, C.Y. Forkhead box proteins: Tuning forks for transcriptional harmony. Nat. Rev. Cancer 2013, 13, 482–495. [Google Scholar] [CrossRef]
- Li, C.; Lai, C.F.; Sigman, D.S.; Gaynor, R.B. Cloning of a cellular factor, interleukin binding factor, that binds to NFAT-like motifs in the human immunodeficiency virus long terminal repeat. Proc. Natl. Acad. Sci. USA 1991, 88, 7739–7743. [Google Scholar] [CrossRef]
- Nirula, A.; Moore, D.J.; Gaynor, R.B. Constitutive binding of the transcription factor interleukin-2 (IL-2) enhancer binding factor to the IL-2 promoter. J. Boil. Chem. 1997, 272, 7736–7745. [Google Scholar] [CrossRef]
- Li, C.; Lusis, A.J.; Sparkes, R.; Nirula, A.; Gaynor, R. Characterization and chromosomal mapping of the gene encoding the cellular DNA binding protein ILF. Genomics 1992, 13, 665–671. [Google Scholar] [CrossRef]
- Hackmann, K.; Stadler, A.; Schallner, J.; Franke, K.; Gerlach, E.M.; Schrock, E.; Rump, A.; Fauth, C.; Tinschert, S.; Oexle, K. Severe intellectual disability, West syndrome, Dandy-Walker malformation, and syndactyly in a patient with partial tetrasomy 17q25.3. Am. J. Med. Genet. Part A 2013, 161, 3144–3149. [Google Scholar] [CrossRef] [PubMed]
- Crujeiras, A.B.; Pissios, P.; Moreno-Navarrete, J.M.; Diaz-Lagares, A.; Sandoval, J.; Gomez, A.; Ricart, W.; Esteller, M.; Casanueva, F.F.; Fernandez-Real, J.M. An Epigenetic Signature in Adipose Tissue Is Linked to Nicotinamide N-Methyltransferase Gene Expression. Mol. Nutr. Food Res. 2018, 62, e1700933. [Google Scholar] [CrossRef]
- Reinhardt, H.C.; Yaffe, M.B. Phospho-Ser/Thr-binding domains: Navigating the cell cycle and DNA damage response. Nat. Rev. Mol. Cell Boil. 2013, 14, 563–580. [Google Scholar] [CrossRef]
- Almawi, A.W.; Matthews, L.A.; Guarne, A. FHA domains: Phosphopeptide binding and beyond. Prog. Biophys. Mol. Boil. 2017, 127, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Katoh, M.; Katoh, M. Identification and characterization of human FOXK1 gene in silico. Int. J. Mol. Med. 2004, 14, 127–132. [Google Scholar] [CrossRef]
- Chen, X.; Ji, Z.; Webber, A.; Sharrocks, A.D. Genome-wide binding studies reveal DNA binding specificity mechanisms and functional interplay amongst Forkhead transcription factors. Nucleic Acids Res. 2016, 44, 1566–1578. [Google Scholar] [CrossRef]
- Ji, Z.; Donaldson, I.J.; Liu, J.; Hayes, A.; Zeef, L.A.; Sharrocks, A.D. The forkhead transcription factor FOXK2 promotes AP-1-mediated transcriptional regulation. Mol. Cell. Boil. 2012, 32, 385–398. [Google Scholar] [CrossRef]
- Hess, J.; Angel, P.; Schorpp-Kistner, M. AP-1 subunits: Quarrel and harmony among siblings. J. Cell Sci. 2004, 117, 5965–5973. [Google Scholar] [CrossRef]
- Ji, Z.; Mohammed, H.; Webber, A.; Ridsdale, J.; Han, N.; Carroll, J.S.; Sharrocks, A.D. The forkhead transcription factor FOXK2 acts as a chromatin targeting factor for the BAP1-containing histone deubiquitinase complex. Nucleic Acids Res. 2014, 42, 6232–6242. [Google Scholar] [CrossRef]
- Bowman, C.J.; Ayer, D.E.; Dynlacht, B.D. Foxk proteins repress the initiation of starvation-induced atrophy and autophagy programs. Nat. Cell Boil. 2014, 16, 1202–1214. [Google Scholar] [CrossRef]
- Baymaz, H.I.; Fournier, A.; Laget, S.; Ji, Z.; Jansen, P.W.; Smits, A.H.; Ferry, L.; Mensinga, A.; Poser, I.; Sharrocks, A.; et al. MBD5 and MBD6 interact with the human PR-DUB complex through their methyl-CpG-binding domain. Proteomics 2014, 14, 2179–2189. [Google Scholar] [CrossRef]
- Marais, A.; Ji, Z.; Child, E.S.; Krause, E.; Mann, D.J.; Sharrocks, A.D. Cell cycle-dependent regulation of the forkhead transcription factor FOXK2 by CDK.cyclin complexes. J. Boil. Chem. 2010, 285, 35728–35739. [Google Scholar] [CrossRef]
- Komorek, J.; Kuppuswamy, M.; Subramanian, T.; Vijayalingam, S.; Lomonosova, E.; Zhao, L.J.; Mymryk, J.S.; Schmitt, K.; Chinnadurai, G. Adenovirus type 5 E1A and E6 proteins of low-risk cutaneous beta-human papillomaviruses suppress cell transformation through interaction with FOXK1/K2 transcription factors. J. Virol. 2010, 84, 2719–2731. [Google Scholar] [CrossRef]
- Liu, Y.; Ao, X.; Jia, Z.; Bai, X.Y.; Xu, Z.; Hu, G.; Jiang, X.; Chen, M.; Wu, H. FOXK2 transcription factor suppresses ERalpha-positive breast cancer cell growth through down-regulating the stability of ERalpha via mechanism involving BRCA1/BARD1. Sci. Rep. 2015, 5, 8796. [Google Scholar] [CrossRef]
- Shan, L.; Zhou, X.; Liu, X.; Wang, Y.; Su, D.; Hou, Y.; Yu, N.; Yang, C.; Liu, B.; Gao, J.; et al. FOXK2 Elicits Massive Transcription Repression and Suppresses the Hypoxic Response and Breast Cancer Carcinogenesis. Cancer Cell 2016, 30, 708–722. [Google Scholar] [CrossRef]
- Wang, W.; Li, X.; Lee, M.; Jun, S.; Aziz, K.E.; Feng, L.; Tran, M.K.; Li, N.; McCrea, P.D.; Park, J.I.; et al. FOXKs promote Wnt/beta-catenin signaling by translocating DVL into the nucleus. Dev. Cell 2015, 32, 707–718. [Google Scholar] [CrossRef] [PubMed]
- Carbone, M.; Yang, H.; Pass, H.I.; Krausz, T.; Testa, J.R.; Gaudino, G. BAP1 and cancer. Nat. Rev. Cancer 2013, 13, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Chittock, E.C.; Latwiel, S.; Miller, T.C.; Muller, C.W. Molecular architecture of polycomb repressive complexes. Biochem. Soc. Trans. 2017, 45, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Okino, Y.; Machida, Y.; Frankland-Searby, S.; Machida, Y.J. BRCA1-associated protein 1 (BAP1) deubiquitinase antagonizes the ubiquitin-mediated activation of FoxK2 target genes. J. Boil. Chem. 2015, 290, 1580–1591. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Brault, J.J.; Schild, A.; Cao, P.; Sandri, M.; Schiaffino, S.; Lecker, S.H.; Goldberg, A.L. FoxO3 coordinately activates protein degradation by the autophagic/lysosomal and proteasomal pathways in atrophying muscle cells. Cell Metab. 2007, 6, 472–483. [Google Scholar] [CrossRef]
- Shi, X.; Garry, D.J. Sin3 interacts with Foxk1 and regulates myogenic progenitors. Mol. Cell. Biochem. 2012, 366, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Kong, Y.; Rothermel, B.; Garry, D.J.; Bassel-Duby, R.; Williams, R.S. The winged-helix/forkhead protein myocyte nuclear factor beta (MNF-beta) forms a co-repressor complex with mammalian sin3B. Biochem. J. 2000, 345 Pt 2, 335–343. [Google Scholar] [CrossRef]
- He, L.; Gomes, A.P.; Wang, X.; Yoon, S.O.; Lee, G.; Nagiec, M.J.; Cho, S.; Chavez, A.; Islam, T.; Yu, Y.; et al. mTORC1 Promotes Metabolic Reprogramming by the Suppression of GSK3-Dependent Foxk1 Phosphorylation. Mol. Cell 2018, 70, 949–960. [Google Scholar] [CrossRef] [PubMed]
- Sukonina, V.; Ma, H.; Zhang, W.; Bartesaghi, S.; Subhash, S.; Heglind, M.; Foyn, H.; Betz, M.J.; Nilsson, D.; Lidell, M.E.; et al. FOXK1 and FOXK2 regulate aerobic glycolysis. Nature 2019, 566, 279–283. [Google Scholar] [CrossRef]
- Spruijt, C.G.; Gnerlich, F.; Smits, A.H.; Pfaffeneder, T.; Jansen, P.W.; Bauer, C.; Munzel, M.; Wagner, M.; Muller, M.; Khan, F.; et al. Dynamic readers for 5-(hydroxy)methylcytosine and its oxidized derivatives. Cell 2013, 152, 1146–1159. [Google Scholar] [CrossRef]
- Iurlaro, M.; Ficz, G.; Oxley, D.; Raiber, E.A.; Bachman, M.; Booth, M.J.; Andrews, S.; Balasubramanian, S.; Reik, W. A screen for hydroxymethylcytosine and formylcytosine binding proteins suggests functions in transcription and chromatin regulation. Genome Boil. 2013, 14, R119. [Google Scholar] [CrossRef] [PubMed]
- Du, Q.; Luu, P.L.; Stirzaker, C.; Clark, S.J. Methyl-CpG-binding domain proteins: Readers of the epigenome. Epigenomics 2015, 7, 1051–1073. [Google Scholar] [CrossRef] [PubMed]
- Fujii, Y.; Nakamura, M. FOXK2 transcription factor is a novel G/T-mismatch DNA binding protein. J. Biochem. 2010, 147, 705–709. [Google Scholar] [CrossRef]
- Liu, J.; Lee, J.B.; Fishel, R. Stochastic Processes and Component Plasticity Governing DNA Mismatch Repair. J. Mol. Boil. 2018, 430, 4456–4468. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Pak, H.; Hammond-Martel, I.; Ghram, M.; Rodrigue, A.; Daou, S.; Barbour, H.; Corbeil, L.; Hebert, J.; Drobetsky, E.; et al. Tumor suppressor and deubiquitinase BAP1 promotes DNA double-strand break repair. Proc. Natl. Acad. Sci. USA 2014, 111, 285–290. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Coic, E.; Lee, K.; Lee, C.S.; Kim, J.A.; Wu, Q.; Haber, J.E. Regulation of budding yeast mating-type switching donor preference by the FHA domain of Fkh1. PLoS Genet. 2012, 8, e1002630. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Spellman, P.T.; Volpe, T.; Brown, P.O.; Botstein, D.; Davis, T.N.; Futcher, B. Two yeast forkhead genes regulate the cell cycle and pseudohyphal growth. Nature 2000, 406, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Pic-Taylor, A.; Darieva, Z.; Morgan, B.A.; Sharrocks, A.D. Regulation of cell cycle-specific gene expression through cyclin-dependent kinase-mediated phosphorylation of the forkhead transcription factor Fkh2p. Mol. Cell. Boil. 2004, 24, 10036–10046. [Google Scholar] [CrossRef] [PubMed]
- Darieva, Z.; Bulmer, R.; Pic-Taylor, A.; Doris, K.S.; Geymonat, M.; Sedgwick, S.G.; Morgan, B.A.; Sharrocks, A.D. Polo kinase controls cell-cycle-dependent transcription by targeting a coactivator protein. Nature 2006, 444, 494–498. [Google Scholar] [CrossRef]
- Garry, D.J.; Meeson, A.; Elterman, J.; Zhao, Y.; Yang, P.; Bassel-Duby, R.; Williams, R.S. Myogenic stem cell function is impaired in mice lacking the forkhead/winged helix protein MNF. Proc. Natl. Acad. Sci. USA 2000, 97, 5416–5421. [Google Scholar] [CrossRef]
- Petrie, M.A.; Kimball, A.L.; McHenry, C.L.; Suneja, M.; Yen, C.L.; Sharma, A.; Shields, R.K. Distinct Skeletal Muscle Gene Regulation from Active Contraction, Passive Vibration, and Whole Body Heat Stress in Humans. PLoS ONE 2016, 11, e0160594. [Google Scholar] [CrossRef] [PubMed]
- van der Heide, L.P.; Wijchers, P.J.; von Oerthel, L.; Burbach, J.P.; Hoekman, M.F.; Smidt, M.P. FoxK2 is required for cellular proliferation and survival. J. Cell. Physiol. 2015, 230, 1013–1023. [Google Scholar] [CrossRef] [PubMed]
- Nestal de Moraes, G.; Khongkow, P.; Gong, C.; Yao, S.; Gomes, A.R.; Ji, Z.; Kandola, N.; Delbue, D.; Man, E.P.; Khoo, U.S.; et al. Forkhead box K2 modulates epirubicin and paclitaxel sensitivity through FOXO3a in breast cancer. Oncogenesis 2015, 4, e167. [Google Scholar] [CrossRef]
- Chen, S.; Jiang, S.; Hu, F.; Xu, Y.; Wang, T.; Mei, Q. Foxk2 inhibits non-small cell lung cancer epithelial-mesenchymal transition and proliferation through the repression of different key target genes. Oncol. Rep. 2017, 37, 2335–2347. [Google Scholar] [CrossRef]
- Zhang, F.; Ma, X.; Li, H.; Zhang, Y.; Li, X.; Chen, L.; Guo, G.; Gao, Y.; Gu, L.; Xie, Y.; et al. FOXK2 suppresses the malignant phenotype and induces apoptosis through inhibition of EGFR in clear-cell renal cell carcinoma. Int. J. Cancer 2018, 142, 2543–2557. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wei, X.; Niu, W.; Wang, D.; Wang, B.; Zhuang, H. Downregulation of FOXK2 is associated with poor prognosis in patients with gastric cancer. Mol. Med. Rep. 2018, 18, 4356–4364. [Google Scholar] [CrossRef]
- Wang, B.; Zhang, X.; Wang, W.; Zhu, Z.; Tang, F.; Wang, D.; Liu, X.; Zhuang, H.; Yan, X. Forkhead box K2 inhibits the proliferation, migration, and invasion of human glioma cells and predicts a favorable prognosis. OncoTargets Ther. 2018, 11, 1067–1075. [Google Scholar] [CrossRef]
- Qian, Y.; Xia, S.; Feng, Z. Sox9 mediated transcriptional activation of FOXK2 is critical for colorectal cancer cells proliferation. Biochem. Biophys. Res. Commun. 2017, 483, 475–481. [Google Scholar] [CrossRef]
- Lin, M.F.; Yang, Y.F.; Peng, Z.P.; Zhang, M.F.; Liang, J.Y.; Chen, W.; Liu, X.H.; Zheng, Y.L. FOXK2, regulted by miR-1271-5p, promotes cell growth and indicates unfavorable prognosis in hepatocellular carcinoma. Int. J. Biochem. Cell Boil. 2017, 88, 155–161. [Google Scholar] [CrossRef]
- Jia, Z.; Wan, F.; Zhu, Y.; Shi, G.; Zhang, H.; Dai, B.; Ye, D. Forkhead-box series expression network is associated with outcome of clear-cell renal cell carcinoma. Oncol. Lett. 2018, 15, 8669–8680. [Google Scholar] [CrossRef]
- Nestal de Moraes, G.; Ji, Z.; Fan, L.Y.; Yao, S.; Zona, S.; Sharrocks, A.D.; Lam, E.W. SUMOylation modulates FOXK2-mediated paclitaxel sensitivity in breast cancer cells. Oncogenesis 2018, 7, 29. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Wang, H.; Li, Q.; Qian, Z.; Shen, C. Forkhead box protein k1 recruits TET1 to act as a tumor suppressor and is associated with MRI detection. Jpn. J. Clin. Oncol. 2016, 46, 209–221. [Google Scholar] [CrossRef] [PubMed]
- Park, S.L.; Patel, Y.M.; Loo, L.W.M.; Mullen, D.J.; Offringa, I.A.; Maunakea, A.; Stram, D.O.; Siegmund, K.; Murphy, S.E.; Tiirikainen, M.; et al. Association of internal smoking dose with blood DNA methylation in three racial/ethnic populations. Clin. Epigenet. 2018, 10, 110. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Wang, J.; Tang, W.; Zhan, X.; Li, Y.; Peng, Y.; Huang, X.; Bai, Y.; Zhao, J.; Li, A.; et al. FOXK1 interaction with FHL2 promotes proliferation, invasion and metastasis in colorectal cancer. Oncogenesis 2016, 5, e271. [Google Scholar] [CrossRef] [PubMed]
- Myatt, S.S.; Lam, E.W. The emerging roles of forkhead box (Fox) proteins in cancer. Nat. Rev. Cancer 2007, 7, 847–859. [Google Scholar] [CrossRef]
- Onorati, A.V.; Dyczynski, M.; Ojha, R.; Amaravadi, R.K. Targeting autophagy in cancer. Cancer 2018, 124, 3307–3318. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Interaction Partners | Technique | Interaction Effect | Models | References |
|---|---|---|---|---|
| FOS and JUN | IP | FOXK2 recruits components of AP-1 complex to chromatin and participates in the transcription of its target genes | U2OS and 293T cell lines | [12] |
| SIN3A and BAP1-containing PR-DUB complexes | RIME, co-IP and PLA assay | FOXK2 recruits BAP-1 to chromatin, promoting local histone deubiquitination and leading to activation and repression of target genes | U2OS and HEK293T cell lines | [14] |
| SIN3A | TAP-MS and co-IP | FOXK2 interacts with SIN3A and represses genes involved in the initiation of starvation-induced atrophy and autophagy programs | C2C12 myoblasts and IMR90 normal fibroblasts | [15] |
| SIN3/HDAC, NCoR, PR-DUB and NSL complexes | GFP pull down prior to MS | FOXK2/PR-DUB share target genes with MBD6, suggesting an association between FOXK2 and sites of DNA damage | 293T and HeLa cell lines | [16] |
| CDK-Cyclin complexes | GST pull-down assay | FOXK2 is regulated by cell cycle regulatory machinery and degraded upon CDK-mediated phosphorylation | HEK293 and U2OS cell lines | [17] |
| E1A and E6 viral proteins | TAP-MS and co-IP | FOXK2 suppresses cell proliferation, oncogenic transformation and tumorigenesis in vivo | KB, A549, 293 and HeLa cell lines; tumours in athymic mice | [18] |
| ERα and BARD1 | Co-IP, mammalian two hybrid system and GST pulldown assay | FOXK2 acts as a scaffold protein for ERα and BARD1, leading to a negative regulation of ERα and target genes and suppressing ERα-positive breast cancer proliferation | HEK 293T, MCF-7, Bcap-37, MDA-MB-231 and T47D cell lines | [19] |
| NCoR/SMRT, SIN3A, NuRD and REST/CoREST complexes | TAP-MS, co-IP and GST pull down | FOXK2 suppresses proliferation and invasion in models of breast cancer in vitro and in vivo and regulates several signaling pathways, including hypoxia | MCF-7 and HEK293T cell lines; xenograft tumours in SCID mice; breast carcinoma patients samples | [20] |
| DVL | TAP-MS and co-IP | FOXK2 interacts with DVL and promotes its nuclear translocation, which then activates Wnt signaling contributing to colorectal tumorigenesis | HEK293T, HeLa, HT29, DLD-1 cell lines; xenograft tumours; transgenic mice; colorectal patients samples | [21] |
| Human Cancer | Genes Regulated | Proteins Encoded | Technique | Effect on Transcription | Function | Oncogene or Tumour Suppressor | Models | References |
|---|---|---|---|---|---|---|---|---|
| Osteosarcoma | CDKN1B, MCM3, CDC14A, KDM3A, KLF9, PDE7A and CAPN2 | p27, MCM3, CDC14A, KDM3A, KLF9, PDE7A and CAPN2 | ChIP | Activation and repression | Regulation of gene networks involved in cell adhesion and motility, control of transcription and metabolism, apoptosis and cancer | N/A | Cell lines | [12] |
| Osteosarcoma | CDKN1A | p21 | Luciferase assay | Repression | Implicated in cell cycle regulatory machinery, CDK phosphorylation of FOXK2 impacts cell viability | Oncogene | Cell lines | [17] |
| Breast cancer | FOXO3A | FOXO3a | ChIP, knockdown and overexpression | Activation | Mediates cytotoxic effects of paclitaxel and epirubicin | Tumour suppressor | Cell lines; invasive carcinoma patients samples | [43] |
| Breast cancer | SURVIVIN, BCAS3, CUL4B, EZH2, FOXC2, HIFβ, CD44, VEGF, CREBBP, HIG2 and HSP90AA1 | Survivin, BCAS3, CUL4B, EZH2, FOXC2, HIFβ, CD44, VEGF, CREBBP, HIG2 and HSP90AA1 | ChIP-seq, qChIP and knockdown | Repression | Inhibits cell proliferation, migration and invasion in vitro and suppresses tumour growth and metastasis in vivo, mainly, by repressing hypoxia pathway | Tumour suppressor | Cell lines; xenograft tumours in SCID mice; breast carcinoma patients samples | [20] |
| Non-small cell lung cancer | CDH2, SNAIL, CCND1 and CDK4 | N-cadherin, SNAIL, Cyclin D1 and CDK4 | ChIP-seq, qChIP, luciferase assays, knockdown and overexpression | Repression | Inhibits cell proliferation in part through induction of G1 arrest and suppresses EMT | Tumour suppressor | Cell lines; lung adenocarcinoma patients samples | [44] |
| Human Cancer | Number of Patients | Technique | % FOXK2 Overexpression | Subcellular Localization | Association with Clinical-Biological Parameters | Prognosis | Oncogene or Tumour Suppressor | Reference |
|---|---|---|---|---|---|---|---|---|
| Breast cancer | 53 samples | IHC | 47.2% | N/A | Negative association with ERα expression | N/A | Tumour suppressor | [19] |
| Breast cancer | 86 samples | IHC | 48.8% | Nuclear | Association with FOXO3a expression in samples from patients receiving chemotherapy; Association with ERα and tumour stage | Poor | Tumour suppressor | [43] |
| Breast cancer | 140/25 samples | IHC/RT-PCR | N/A | N/A | FOXK2 mRNA and protein levels are downregulated in breast carcinomas compared to adjacent tissues; Negative correlation with EZH2 and HIF1β levels, histological grade and lymph node positivity | Favorable | Tumour suppressor | [20] |
| Clear-cell renal cell carcinoma | 42 samples | IHC/qRT-PCR/Western blotting | 50% | Nuclear | FOXK2 mRNA and protein levels are downregulated compared to adjacent non-tumour renal tissues | Favorable | Tumour suppressor | [45] |
| Gastric Cancer | 150 samples | IHC | 48.6% | N/A | FOXK2 mRNA levels downregulated in gastric cancer, compared to non-tumour tissues; FOXK2 protein expression associated with tumour differentiation | Favorable | Tumour suppressor | [46] |
| Glioma | 151/46 samples | IHC/PCR | 39.7% | N/A | FOXK2 expression downregulated in high-grade compared to low-grade gliomas and negatively associated with KI67 staining and tumour grade | Favorable | Tumour suppressor | [47] |
| Non-small cell lung cancer | 50 samples | qRT-PCR | N/A | N/A | FOXK2 mRNA levels are downregulated in lung cancer, compared to non-tumour tissues | Favorable | Tumour suppressor | [44] |
| Colon cancer | 200 samples | IHC | 45.5% | Nuclear | FOXK2 is overexpressed compared to normal colon and associated with DVL nuclear expression | N/A | Oncogene | [21] |
| Colorectal Cancer | 145 samples | IHC | 48.9% | N/A | FOXK2 is overexpressed compared to normal tissues | Poor | Oncogene | [48] |
| Hepatocellular carcinoma | 505/32/12 samples | IHC/qRT-PCR/Western blotting | 54.1% | Nuclear | High FOXK2 expression is positively associated with tumour size, TNM stage and vascular invasion; FOXK2 mRNA levels are negatively associated with miR1271 | Poor | Oncogene | [49] |
| Clear-cell renal cell carcinoma | 525 samples | *TCGA | 49.7% | N/A | FOXK2 gene expression as an independent prognostic factor and associated with tumour grade and gender | Poor | Oncogene | [50] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nestal de Moraes, G.; Carneiro, L.d.T.; Maia, R.C.; Lam, E.W.-F.; Sharrocks, A.D. FOXK2 Transcription Factor and Its Emerging Roles in Cancer. Cancers 2019, 11, 393. https://doi.org/10.3390/cancers11030393
Nestal de Moraes G, Carneiro LdT, Maia RC, Lam EW-F, Sharrocks AD. FOXK2 Transcription Factor and Its Emerging Roles in Cancer. Cancers. 2019; 11(3):393. https://doi.org/10.3390/cancers11030393
Chicago/Turabian StyleNestal de Moraes, Gabriela, Luciana da Torre Carneiro, Raquel Ciuvalschi Maia, Eric Wing-Fai Lam, and Andrew David Sharrocks. 2019. "FOXK2 Transcription Factor and Its Emerging Roles in Cancer" Cancers 11, no. 3: 393. https://doi.org/10.3390/cancers11030393
APA StyleNestal de Moraes, G., Carneiro, L. d. T., Maia, R. C., Lam, E. W.-F., & Sharrocks, A. D. (2019). FOXK2 Transcription Factor and Its Emerging Roles in Cancer. Cancers, 11(3), 393. https://doi.org/10.3390/cancers11030393