1. Introduction
Monoclonal antibodies (mAbs) are one of the current success stories of modern biotechnology. Hundreds of different diagnostic kits based on mAb technology are available in the market [
1,
2,
3]. The key advantage of mAbs is their high specificity for the relevant disease targets, which limits harm to healthy cells, resulting in fewer adverse effects compared to traditional pharmaceuticals based on small organic molecules. However, early excitement has been tempered by problems associated with the use of mAbs. These large (150 kDa) multimeric proteins contain numerous disulphide bonds and posttranslational modifications such as glycosylation, and require a sophisticated eukaryotic machinery to be produced in an active form [
3]. The fragment crystallizable region (Fc) of mAbs can interact with Fc receptors expressed at the surface of many cell types, which increases their cross-reactivity and retention in the circulation. Moreover, mAbs developed in animals must be humanized prior to use in clinical applications. Consequently, the production of therapeutic mAbs necessitates use of very large cultures of mammalian cells followed by extensive purification under good manufacturing practice (GMP) conditions. Overall, high production costs, regulatory affairs, safety and short shelf life have thus limited broad use of these antibodies.
These limitations have sparked considerable interest in the development of molecular tools that offer the functionality of mAbs, but that can be easily engineered, synthesized, and tailored to match desired characteristics. Aptamers have enormous potential as such a tool [
4,
5]. These small (5–30 kDa), single-stranded DNA and RNA molecules carry the blueprint for their own synthesis in their primary sequence, so they can be synthesized by chemical or enzymatic procedures. They fold into well-defined three-dimensional structures, show high affinity and specificity for their targets, and can be used to inhibit biological functions. Aptamers are often viewed as “chemical antibodies” with applications ranging from target detection to drug discovery and delivery. In many respects, aptamers are superior to antibodies. They can be chemically synthesized in high purity at low cost (thousands of times cheaper than mAbs), and are considered as a synthetic chemical product, but not a biological product. Biological products are produced from or extracted from a biological (living) system and require, in addition to physico-chemical testing, biological testing for full characterization. Lastly, aptamers are low-immunogenic and low-toxic and can be applied as drugs themselves [
5,
6,
7]. Indeed, one RNA aptamer (Macugen™) directed against vascular endothelial growth factor was approved by the Food and Drug Administration (FDA) in 2005 [
8]. An emerging market of aptamers is considered as a rival to antibodies in the biotech industry.
In this article, we selected aptamers ECM-APT-01 and ECM-APT-02 against the epithelial cell adhesion molecule (EpCAM) expressed on primary lung cancer cells isolated from tumors of patients with non-small cell lung cancer (NSCLC) using competitive displacement by monoclonal EpCAM antibody. Moreover, we confirmed aptamers specificity for the EpCAM antibody binding site and its ability to detect EpCAM in clinical samples. EpCAM is a 40 kDa glycoprotein initially identified as a marker for carcinoma, and further studies revealed EpCAM as a prevalent protein also expressed by normal epithelia; however, at a lower level than by carcinoma cells [
9]. EpCAM is expressed in various carcinomas such as those in the colon and rectum, prostate, liver, esophagus, lung, head and neck, pancreas, and breast [
10]. High-level and mostly homogenous EpCAM expression was found on 85% of adenocarcinomas and on 72% of squamous cell carcinomas [
11]. In addition, EpCAM is one of the main biomarkers of circulating tumor cells and microemboli in blood [
12].
3. Discussion
Traditionally, aptamers are raised against pure (recombinant) proteins by Systematic Evolution of Ligands by EXponential Enrichment (SELEX) [
14,
15]. However, it is very likely that at least some of the selected aptamers will not be able to recognize the native proteins in vivo, if (for example) the binding sites are hidden inside a cell, or if the folded structures of recombinant and native proteins differ significantly. The goal of our work was the targeted selection of aptamers to a native cell-surface protein of interest (i.e., a protein that is properly folded, posttranslationally modified, and surrounded by other biomolecules). The selection approach was based on LIgand-Guided Selection (LIGS) of target-specific aptamers [
16]. It is a variation of Systematic Evolution of Ligands by Exponential Enrichment (SELEX) and has been introduced by Zumrut et al. in 2016. It does not require any gene transfection and modification of cells. The cells expressing EpCAM were incubated with a DNA library and then washed several times. After removing unbound DNA, the cells were incubated with the excess of a monoclonal antibody against EpCAM. The antibody binds and replaces anti-EpCAM aptamers. Cells are removed and the unbound DNA is amplified by polymerase chain reaction (PCR). All resulted aptamer pools were sequenced, and the aptamer clones from the largest families and the highest enrichment factor were synthesized chemically. Two unique aptamers EPCAM-APT-01 and EPCAM-APT-02 with nanomolar affinity to EpCAM were chosen for CTC detection and isolation.
There is a number of existing DNA aptamers that have been selected to human recombinant EpCAM protein. For example, Wei Duan et al. developed DNA- and RNA-aptamers for EpCAM in 2011 and 2013, respectively [
17,
18]. As authors mentioned in the second article, RNA is notoriously prone to nuclease degradation, which limits its application in clinical research [
18]. DNA-aptamer SYL3C was selected against recombinant EpCAM protein bound with beads. SYL3C specificity and affinity were estimated using flow cytometry and confocal microscopy using different cell cultures.
In some cases, aptamers developed to recombinant proteins do not bind to the same proteins in primary cells and tissues, because these native proteins have posttranslational modifications covering the binding sites of aptamers, and conformations which are not recognizable by the aptamers. In our previous study, DNA aptamers showed good binding ability to postoperative lung adenocarcinoma tissues and had no or low affinity for adenocarcinoma cell cultures [
19].
4. Materials and Methods
4.1. List of Chemicals
Monoclonal antibodies to EpCAM, α-tubulin, a secondary antibody labeled with Alexa Fluor 405 and a secondary antibody labeled with Cy-3 were purchased from Abcam, Cambridge, UK.
4.2. Patient-Derived Tumor Samples
This study was approved by the Local Committee on Ethics of the Krasnoyarsk Regional Clinical Cancer Center named after A.I. Kryzhanovsky No. 8/2011 since 16 March 2011 and Krasnoyarsk State Medical University 37/2012 since 31 January 2012, Krasnoyarsk, Russia. Solid tumors were removed aseptically and immediately immersed in ice-cold RPMI-1640 and transported to the laboratory.
4.3. Aptamer Selection
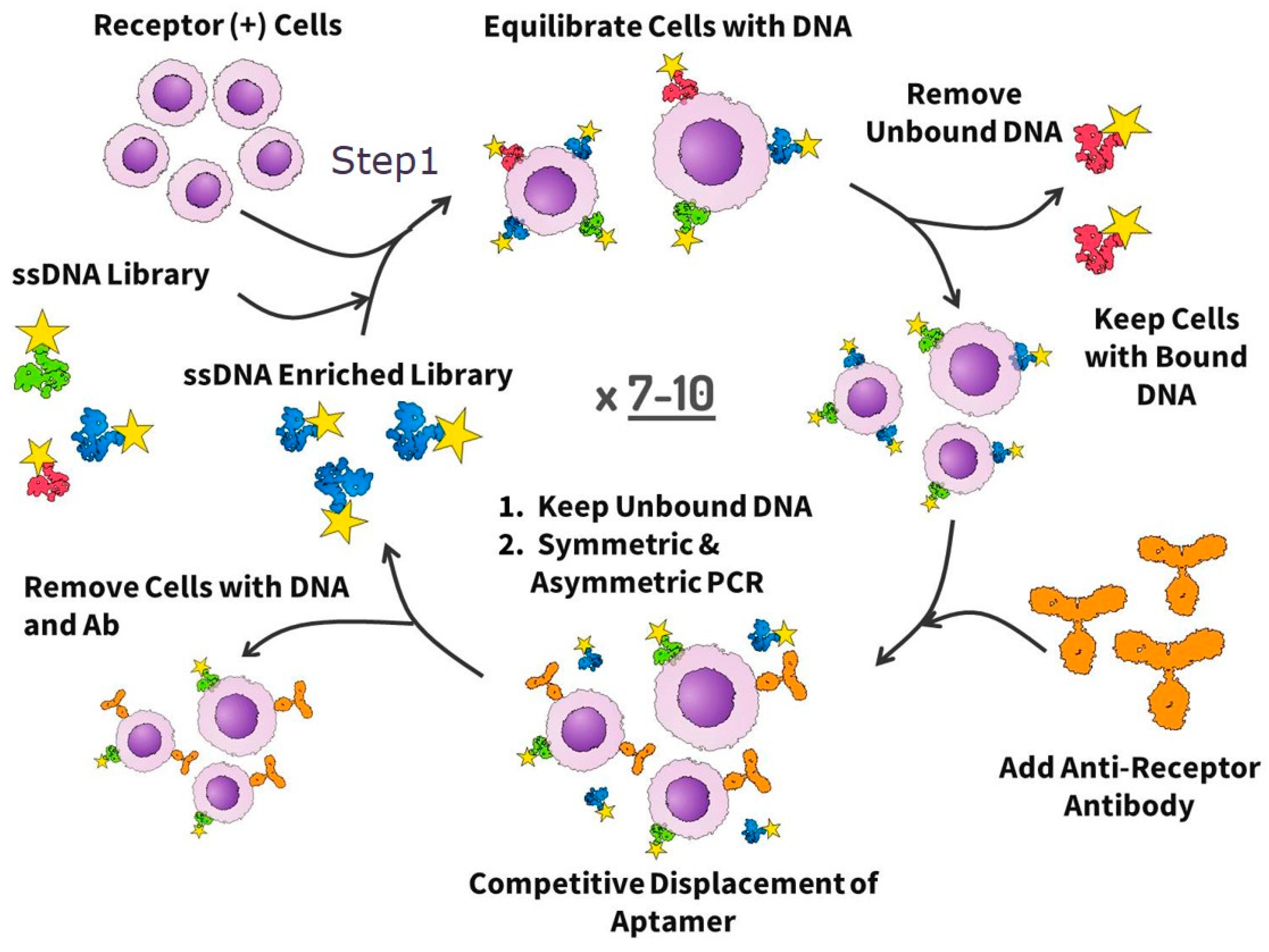
The aptamers were selected from an ssDNA library based on the modified cell-SELEX procedure as shown in
Figure 1. The selection was started with an ssDNA library containing a 40 nt length variable internal region with three stems and loops and flanked on each side by a 20 nt primer. The final form of the 80 nt library is 5′-CTC CTC TGA CTG TAA CCA CGY ZYZ YZY ZNN NNY ZYZ YZY ZYZ NNN YZY ZYZ YZY ZNN NNY G CAT AGG TAG TCC AGA AGC C-3′ where N is a nucleotide mixture that produces the ratio of A/C/G/T as 1:1:1:1, Y is a mixture that produces the ratio of A/C/G/T as 45:5:45:5, and Z is a mixture that produces the ratio of A/C/G/T as 5:45:5:45. Before each round of selection and binding experiment, the ssDNA library and aptamer pools were denatured by heating for 5 min at 95 °C in Dulbecco’s phosphate buffered saline (DPBS, Sigma-Aldrich, St. Louis, MO, USA) and then renatured on ice for 10 min.
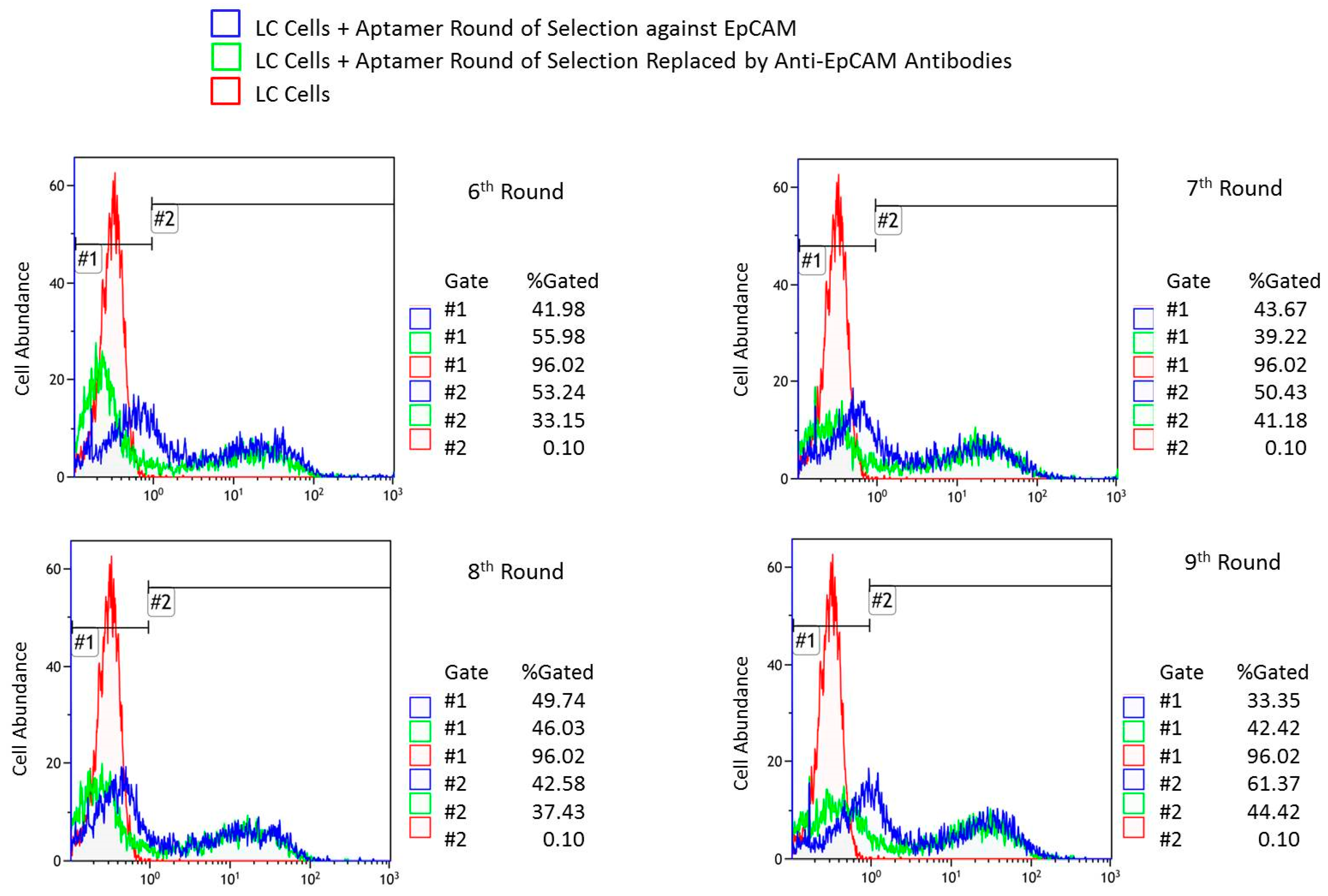
The first five rounds of aptamer development were just positive selections without antibody displacement. The sixth and eighth round included the antibody displacement as follows: (a) incubation of LC tumor tissue with an aptamer pool in DPBS buffer, (b) washing the unbound aptamers with DPBS three times, (c) incubation of the same tumor piece with an anti-EpCAM antibody at the concentration of 2 ng μL−1 for 30 min at 22 °C resulting in aptamer replacement by the antibody and release of the bound oligonucleotides into DPBS buffer, (c) removing of the cells with bound antibodies and oligonucleotides by centrifugation and collecting aptamer candidates in DPBS buffer, and (d) amplification of released aptamers by PCR. The seventh and the ninth selection rounds were without an antibody displacement step. In total, nine rounds of selection were performed.
For each round of selection, tumors were separated from necrotic tissues, washed with DPBS and cut into small pieces with a scalpel on a Petri dish with a small amount of DPBS. For the first round of selection, tumor tissue minced into small pieces was incubated with 100 μL of DPBS containing 1 μM ssDNA library for 30 min at 25 °C with gentle shaking. Then, the sample was centrifuged at 4000× g for 5 min, supernatant containing unbound aptamers was removed and the pellet was rinsed three times with DPBS. Aptamers bound with tissue were released by denaturation in 10 mM Tris-HCl buffer containing 10 mM ethylenediamine tetraacetic acid (EDTA), pH 7.4 (TE, Sigma-Aldrich, St. Louis, MO, USA) at 95 °C for 10 min followed by centrifugation at 13,400× g for 15 min. Next, the supernatant was collected and the aptamers were amplified using symmetric and asymmetric PCR.
For the symmetric PCR, 5 μL of the aptamer pool in 10 mM TE-buffer was mixed with 45 μL of symmetric PCR MasterMix, containing the following: 1× PCR buffer B, 1× Enhancer 1, 1 mM MgCl2, 0.025 U μL−1 KAPA2G HotStart Robust polymerase (KAPABiosystems, Wilmington, MA, USA), 220 μM dNTPs, 300 nM forward primer (5′-CTC CTC TGA CTG TAA CCA CG-3′), and 300 nM reverse primer (5′-GGC TTC TGG ACT ACC TAT GC-3′) (Integrated DNA Technologies, Coralville, IA, USA). Amplification was performed using the following PCR program: preheat for 2 min at 95 °C, 15 cycles of 30 s at 95 °C, 15 s at 56.3 °C, and 15 s at 72 °C. Afterward, asymmetric PCR was performed where 5 μL of the symmetric PCR product was mixed with 45 μL of the asymmetric PCR Master Mix containing the following: 1× PCR buffer B, 1× Enhancer 1, 1 mM MgCl2, 0.025 U μL−1 KAPA2G HotStart Robust polymerase, 220 μM dNTPs, 1 μM FAM-forward primer (5′-FAM-CTC CTC TGA CTG TAA CCA CG-3′), and 50 nM reverse primer (5′-GGC TTC TGG ACT ACC TAT GC-3′). Amplification was performed using the following PCR program: preheat for 2 min at 95 °C, 15 cycles of 30 s at 95 °C, 15 s at 56.3 °C, and 15 s at 72 °C. The PCR product was washed by 30 kDa cutoff filters and the concentration of the ssDNA was measured by NanoDrop (Thermo Scientific, Wilmington, DE, USA). Fluorescence of ssDNA labeled with FAM was analyzed in the gel-documenting system GBOX/EF2-E (Syngene, Frederick, MD, USA). Evolved aptamer pools were stored at −20 °C.
The sixth and the eighth rounds of aptamer selection were done as described below. The asymmetric PCR product obtained from the previous round was incubated with lung cancer tissue for 30 min with shaking at 25 °C. Thereafter, the sample was incubated with monoclonal antibodies to EpCAM (2 ng μL−1) for 30 min with shaking at 25 °C. As a result of this process, aptamers bound to the cell membrane receptors were replaced by antibodies and released into DPBS. The sample was centrifuged at 4000× g for 5 min and the supernatant was collected. The aptamers were then amplified using symmetric and asymmetric PCR and washed by cutoff filters. The seventh and the ninth rounds were positive selections and similar to the first five rounds.
4.4. Flow Cytometric Binding Analysis of Aptamers
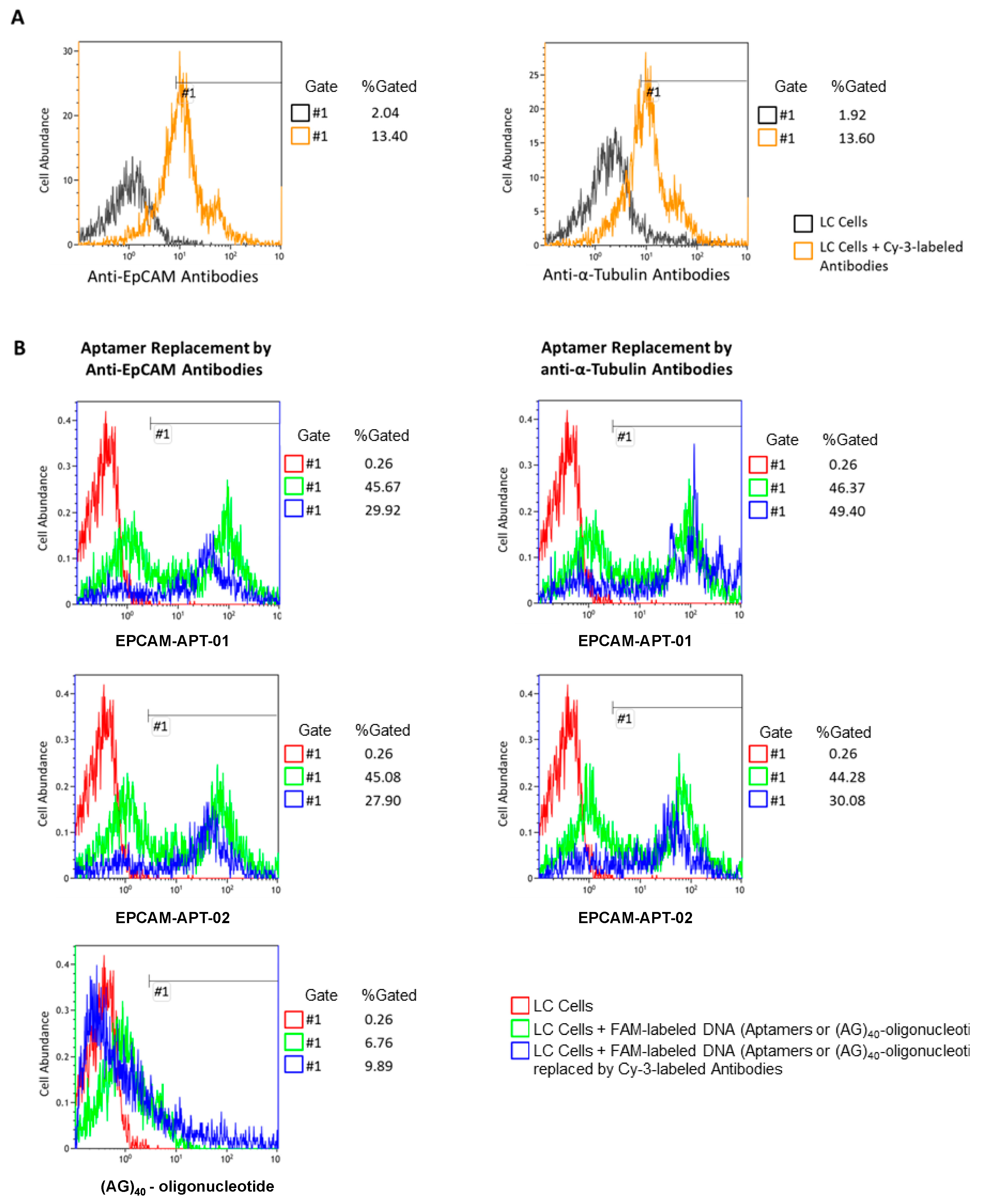
The affinity and specificity of the evolved aptamers was defined by flow cytometry using FC-500 Flow Cytometer (Beckman Coulter Inc., Porterville, CA, USA). Lung cancer material was washed with Dulbecco’s Phosphate Buffered Saline (DPBS) and minced into small pieces and then pipetted gently with DPBS to remove cell clusters and obtain a homogeneous solution. Cell suspension was filtered through 70 μm filters; obtained cells were centrifuged at 3000× g for 5 min and washed three times with DPBS. Next, cells were pre-incubated with masking DNA (1 ng μL−1 of salmon sperm DNA) for 30 min and then with 50 nM of FAM-labeled aptamers from each pool of a selection round or synthetic aptamer sequences for 30 min at 25 °C with shaking. Each sample contained 3 × 105 cells. LC cells pre-incubated with 1 ng μL−1 masking DNA and 50 nM FAM-labeled (AG)40-oligonucleotide were used as a control. The measurements were carried out using flow cytometry. The samples were then incubated with 2 ng μL−1 of Cy-3 labeled anti-EpCAM monoclonal antibody for 30 min with shaking at 25 °C followed by flow cytometry.
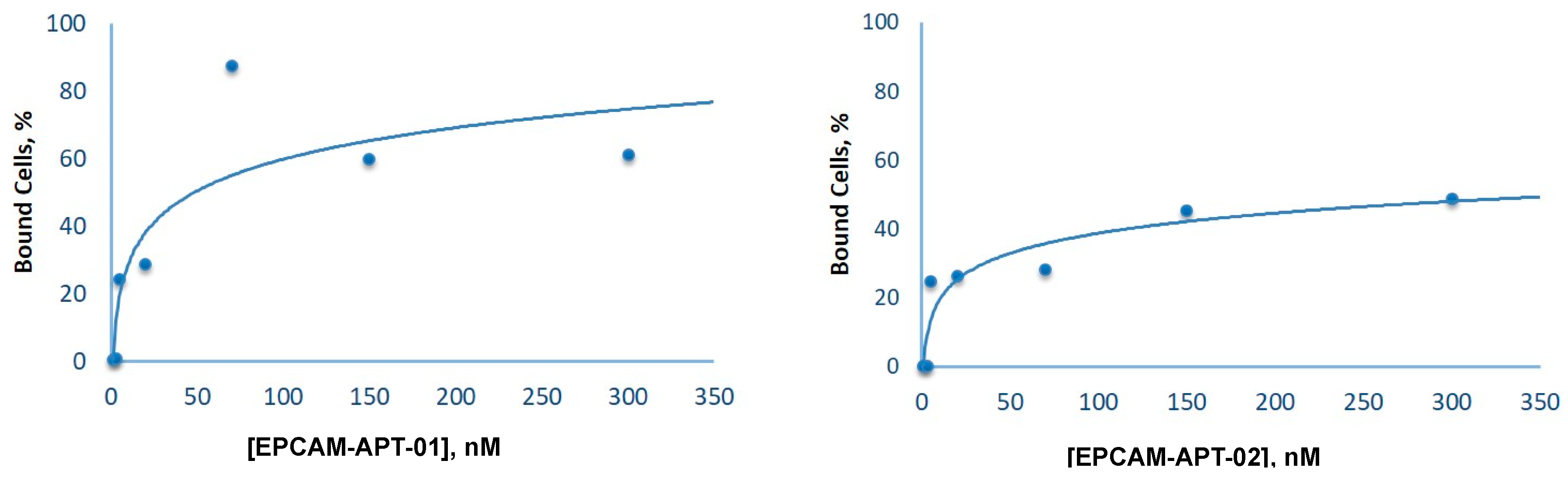
To evaluate the dissociation constants of EPCAM-APT-01 and EPCAM-APT-02, LC cells were incubated with 1 nМ, 3 nМ, 5 nМ, 20 nМ, 70 nМ, 150 nМ, and 300 nМ of FAM-labeled aptamers. The measurements were carried out using flow cytometry. The data were analyzed with Kaluza® 1.2 Software (Beckman Coulter Inc., Indianapolis, IN, USA). The dissociation constants were determined from the plots as half of the concentrations observed at maximum binding.
4.5. NGS Sample Preparation Procedure
To barcode the pools for NGS, the pools were first prepared by adding an 8 nt barcode to the 5′ end of the aptamers using symmetric PCR. Approximately 10 ng in 5 µL volumes of each pool was used for a PCR reaction in order to amplify and barcode single-stranded (ss) DNA molecules using a forward primer (5′-BBB BBB BBC TCC TCT GAC TGT AAC CACG-3′) containing an 8-base barcode (B) at the 5′ end and a reverse primer (5’-GGC TTC TGG ACT ACC TAT GC-3′). Symmetric amplification was performed by preparing five 50 µL PCR reactions using the following: 7% DMSO, 1.5 mM MgCl2, 0.2 mM dNTP (Promega Corporation, Madison, WI, USA), 0.4 µM barcoded forward primer, 0.4 µM reverse primer, and 0.02 U µL−1 of Phire II polymerase. PCR was performed in a Mastercycler pro S thermal cycler (Eppendorf, Hamburg, Germany). The settings for the thermal cycler were as follows: melting at 94 °C for 30 s, annealing at 58 °C for 15 s and extending at 72 °C for 15 s. After PCR amplification, the five 50 µL aliquots for each pool were combined and then separated and visualized by gel electrophoresis (2% agarose, 1× Tris-Acetate EDTA buffer). The band corresponding to 88 bp DNA was cut-out, then sliced into small pieces, weighed and placed in a new 50 mL Eppendorf tube with 1 mL of water per 1 mg of gel and left to elute for 24 h. The buffer was then collected and the DNA was concentrated using 3 kDa centrifugal filters (Pall Corporation, Show Low, AZ, USA). All the pools were combined and sequenced at Eurofins Genomics (Louisville, KY, USA).
4.6. Mathematical Analysis of Aptamer DNA Sequences
The central 40-nucleotides random region of the 80 nt aptamer sequences was analyzed. The 20-nucleotides forward and reverse primers were dropped in the analysis to simplify the procedure. The main challenge of the mathematical analysis was to choose unique aptamers forming the best families of sequences with good enrichment. To solve this problem, the following algorithm was used.
For each aptamer from the best round of aptamer selection against EpCAM the total amount of copies of the similar sequences where and where was calculated.
Aptamers with the greatest values of and with various values of were chosen (i.e., aptamers forming the largest families of sequences).
From the aptamers that were chosen in paragraph 2, sequences with the best enrichment were chosen ( in most rounds of selection).
The detailed description of the mathematical analyses is presented in
Appendix A.
4.7. Histological Tissue Staining
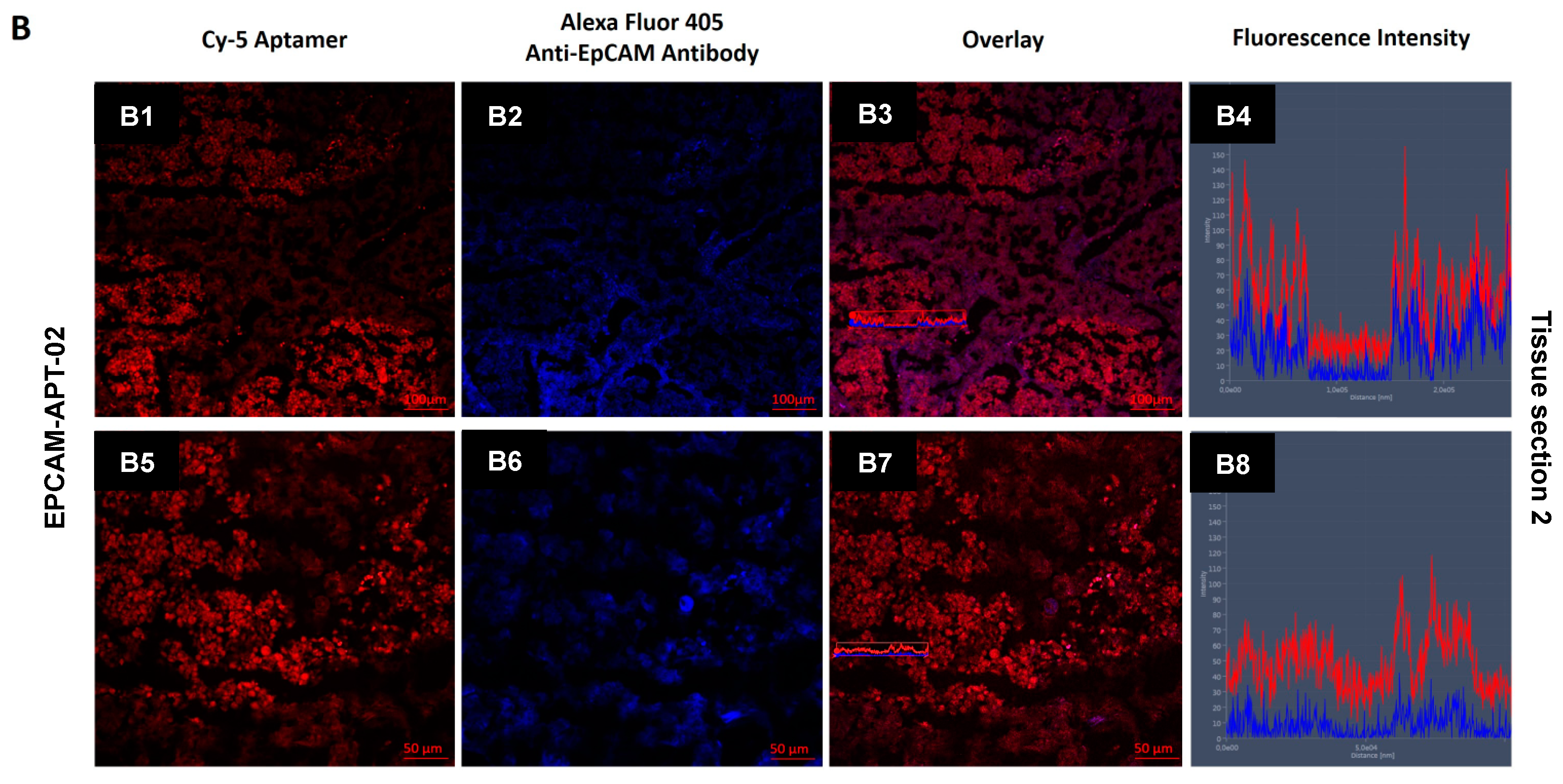
Tissue pieces were frozen in liquid nitrogen and sliced into 5 µm sections by Microm HM525 Cryostat and placed on poly-lysine coated glass slides. First, nonspecific binding of the antibodies was blocked by incubation of the sections with 10% of Bovine Serum Albumin (Sigma-Aldrich) for 30 min, followed by incubation with primary anti-EpCAM antibody (2 ng μL−1) in a humidified atmosphere for 1 h and then with a secondary antibody labeled with Alexa Fluor 405 (2 ng μL−1) in a humidified atmosphere for 1 h and then washed three times with DPBS. Nonspecific binding of the aptamers was blocked with yeast RNA (1 ng µL−1) (Sigma-Aldrich) and then incubated with 50 nM of Cy-5-labeled aptamers for 1 h in a humidified atmosphere and washed with DPBS. Bio Mount mounting medium (Bio-Optica, Milano, Italy) was used to fix the sections.
4.8. Isolation of CTCs from Human Blood
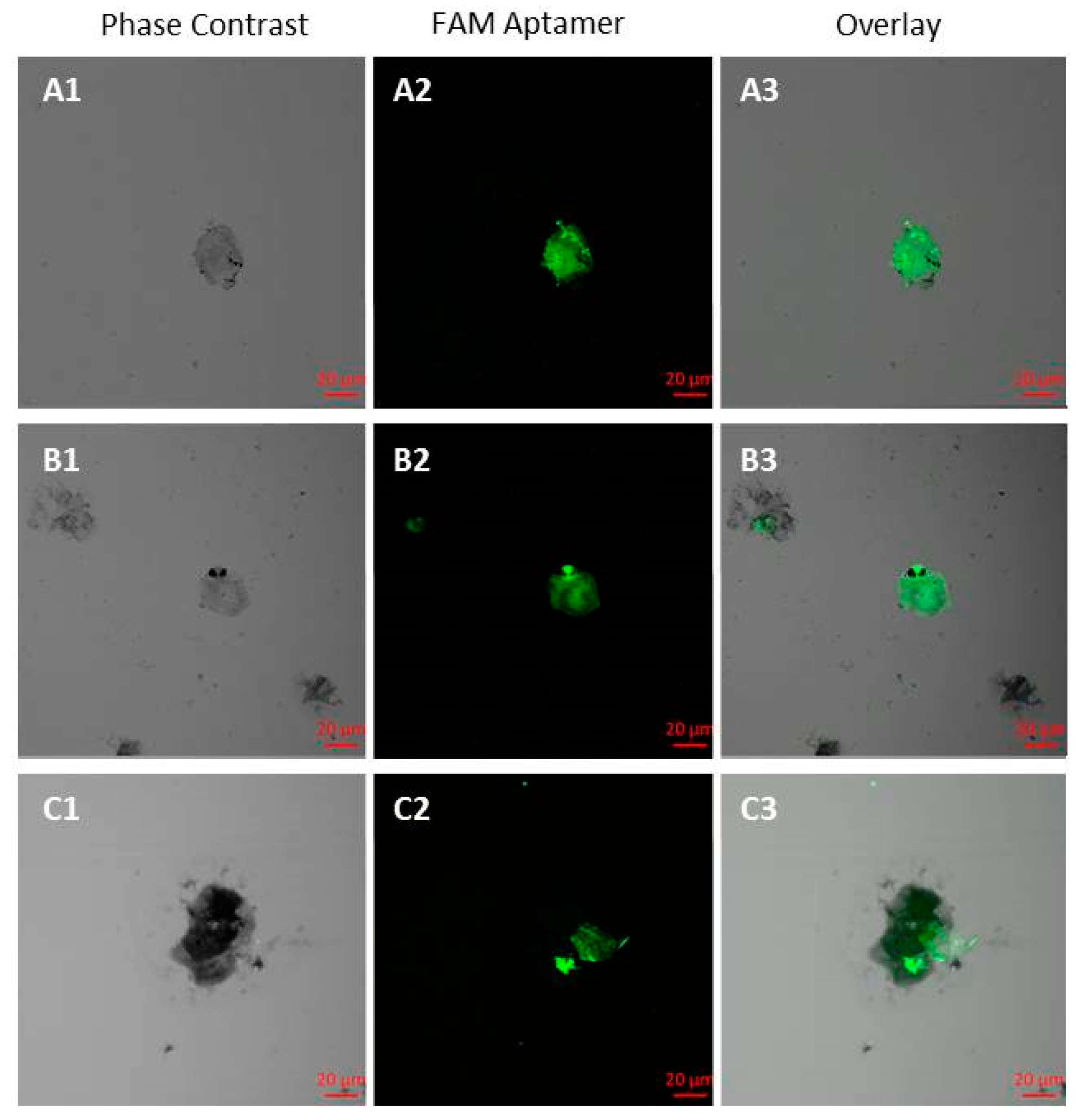
Isolation of CTCs was performed from 3.5 mL of patients’ blood 1–1.5 h after collecting into Becton, Dickinson Vacutainer Heparin Tubes (Becton, Dickinson and Company, Franklin Lakes, NJ, USA). The blood was centrifuged (1500× g for 10 min) to remove the plasma. The cell pellet was transferred into a 15 mL centrifuge tube using a bovine serum albumin (BSA) treated tip. Red blood cells (RBCs) were lysed with hypotonic NH4Cl solution in a Vacutainer tube. Cells were rinsed with 2 mL of 0.42% NH4Cl with heparin, pipetted up and down 5 times and poured into the 15 mL tube with 8 mL of 0.42% NH4Cl with heparin, incubated for 10 min on a shaker and centrifuged at 3500× g for 5 min. The remaining cell pellet was resuspended in 100 μL DPBS and incubated with masking yeast RNA (0.1 mg mL−1) for 30 min at room temperature to reduce nonspecific binding. The sample was centrifuged at 3500× g for 5 min, the supernatant was removed and incubated for 30 min with 100 µg of streptavidin-coated paramagnetic beads (Promega Corporation) functionalized with 100 nM biotinylated aptamers against tumor cells. The cells bound with the magnetic particles via the aptamers were separated on a magnetic stand, and resuspended in 100 μL of calcium and magnesium free DPBS buffer and concentrated by a magnet. The pellet containing mostly CTCs was stained with FAM-labeled aptamers at a final concentration of 50 nM for 30 min. Cell smears were prepared for quantification of CTCs in blood of patients. After staining, to prepare the smears, the cell pellet was spread evenly on a glass slide and then fixed in methanol for 5 min, followed by staining with Romanowsky-Giemsa dye. The counting of circulating tumor cells was done on a fluorescent microscope Axiostar plus (Carl Zeiss Group, Oberkochen, Germany).

 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}