Tumor Suppressor Function of miR-127-3p and miR-376a-3p in Osteosarcoma Cells

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
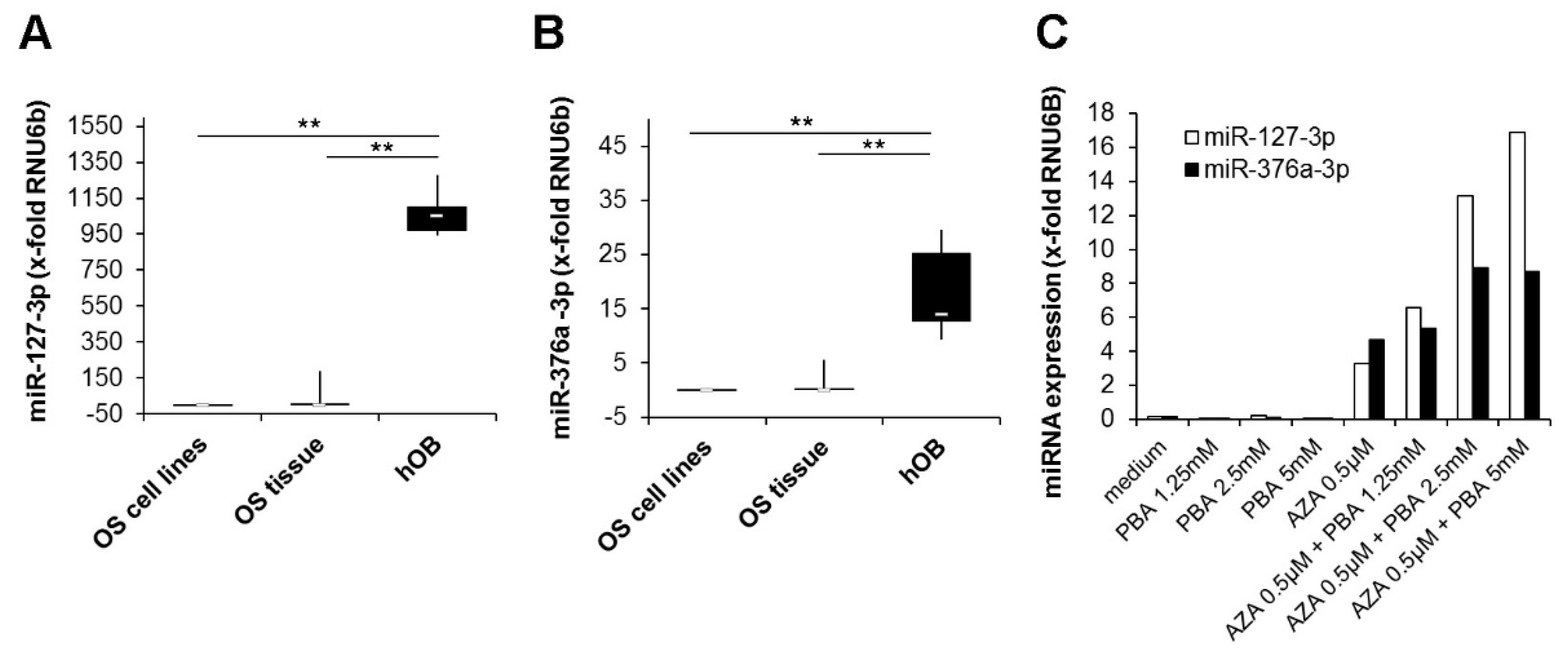
2.1. Silencing of miR-127 and miR-376a in Osteosarcoma Cell Lines and Tissue
2.2. Restoration of miR-127-3p and miR-376a-3p Inhibits Proliferation of Osteosarcoma Cells
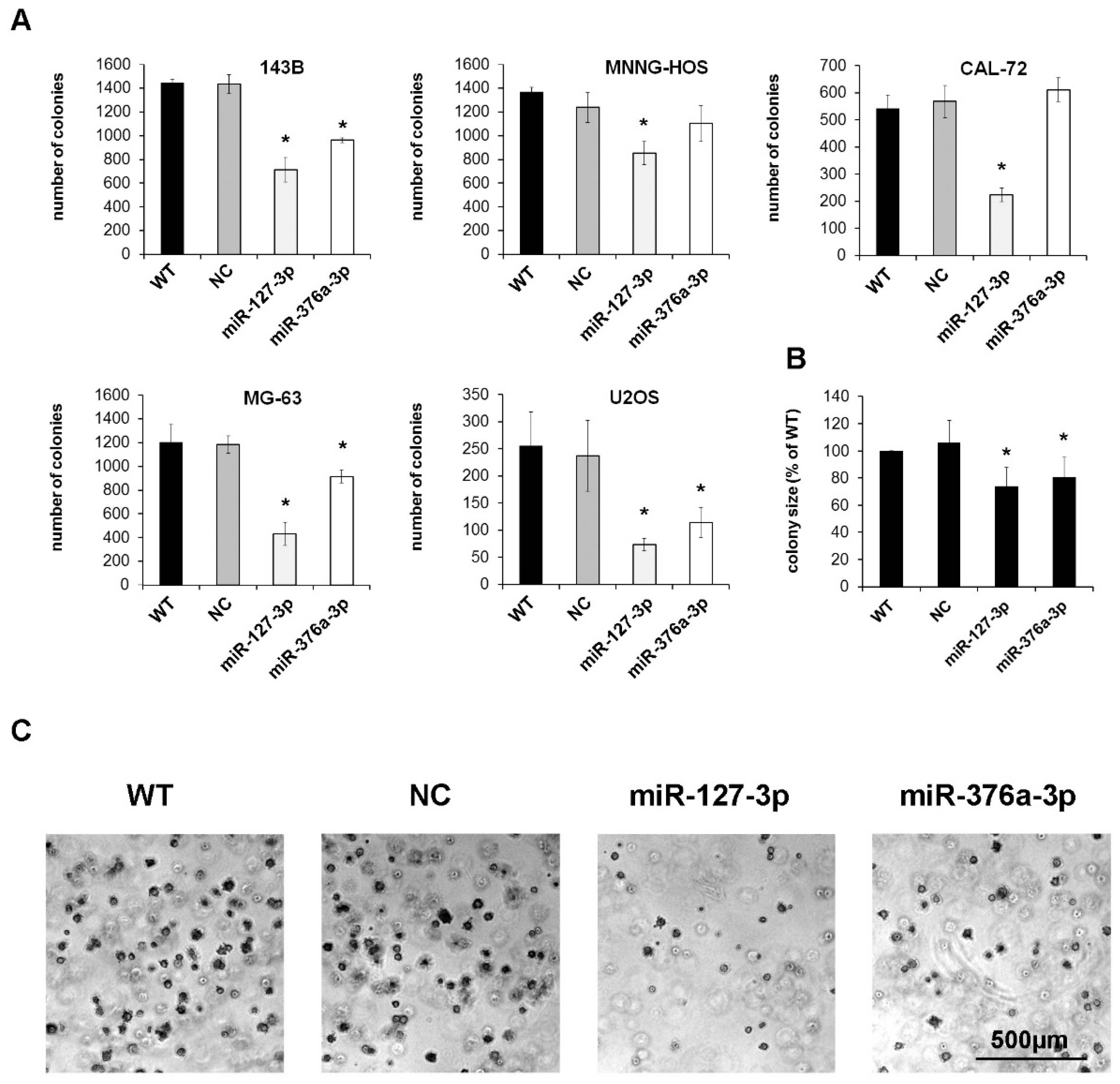
2.3. Mir-127-3p and miR-376a-3p Reduce the Colony Forming Capacity of Osteosarcoma Cells
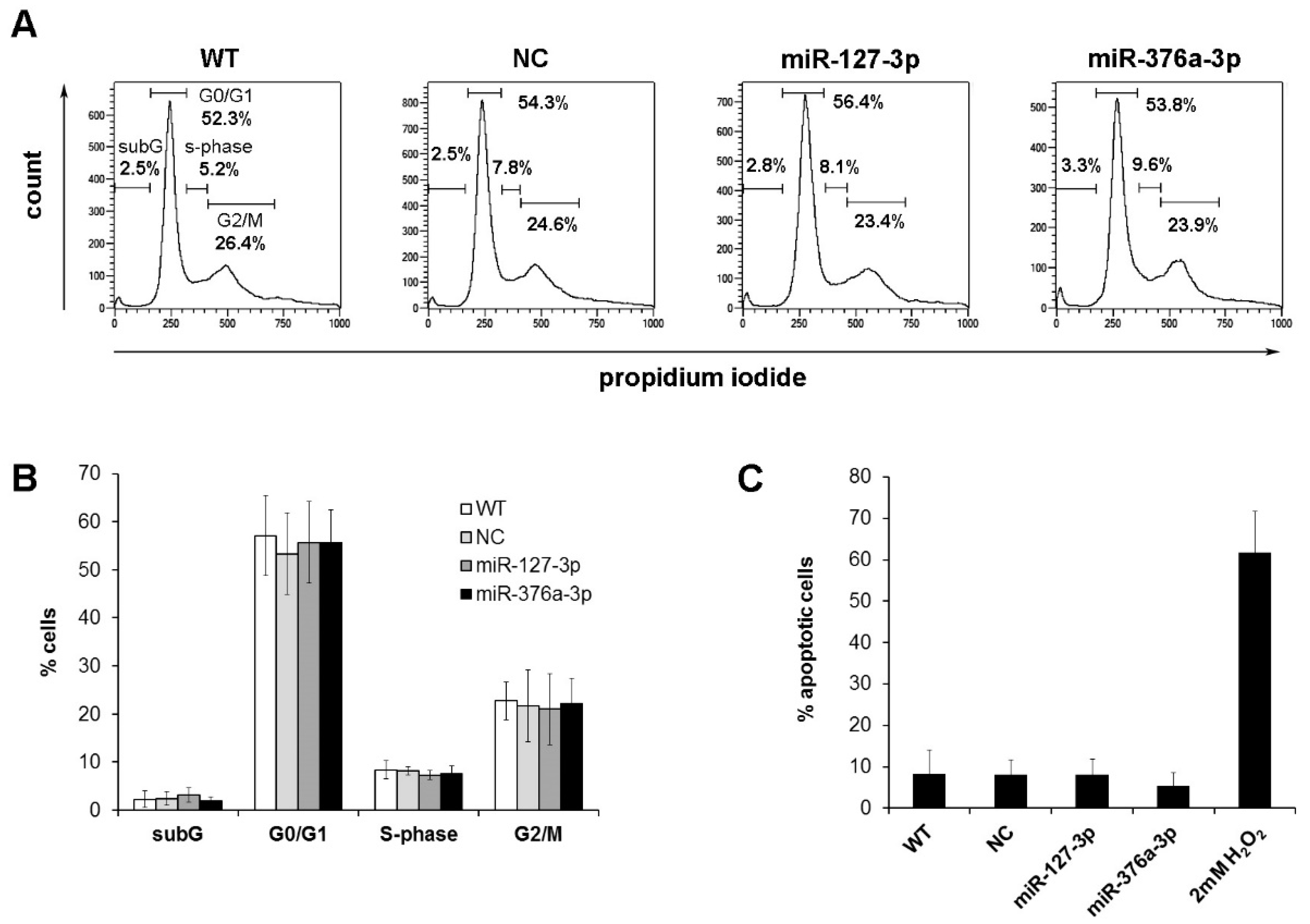
2.4. Cell Cycle Analysis of Osteosarcoma Cells with Restored miRNA Expression
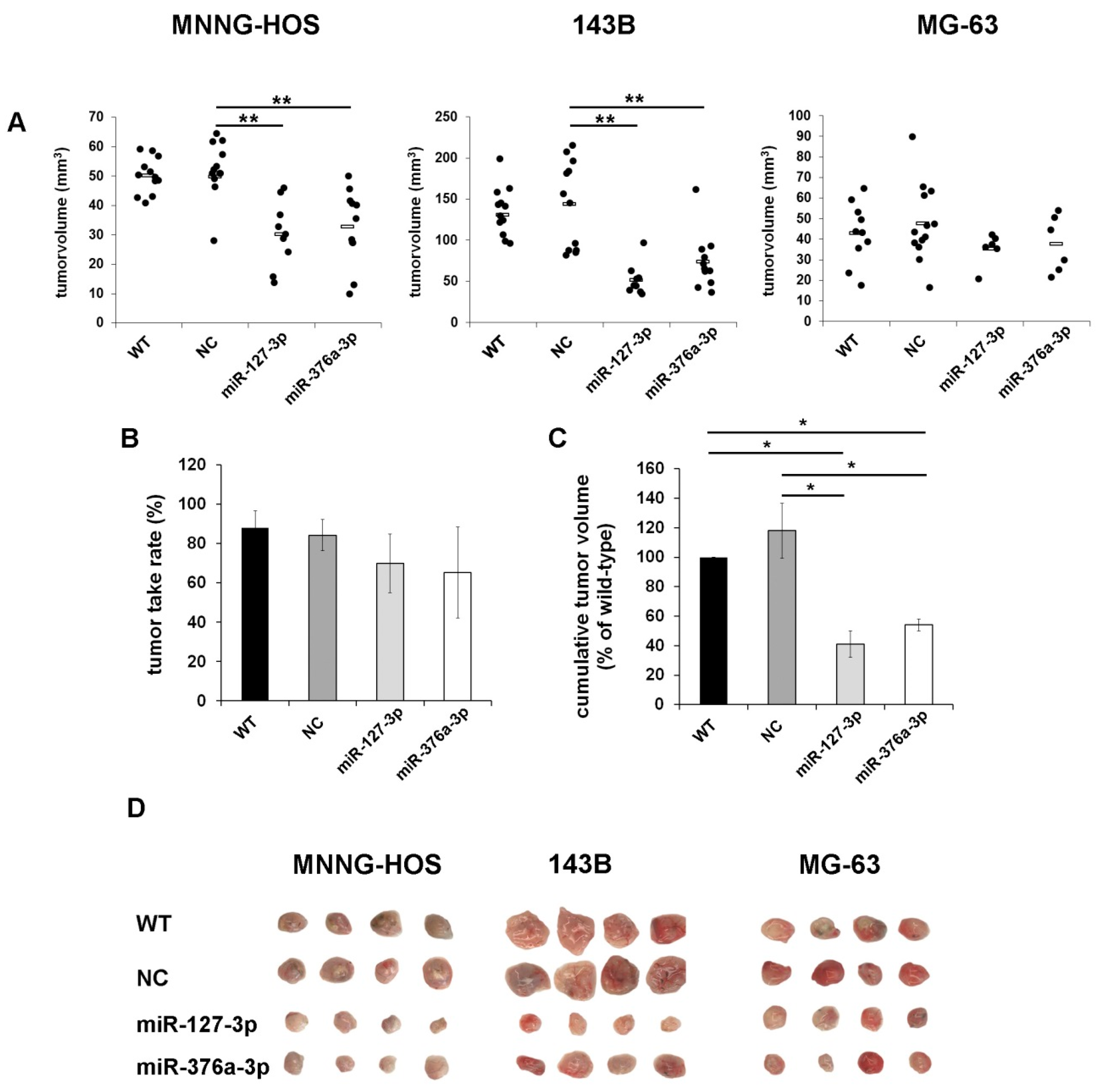
2.5. Restoration of miR-127-3p and miR-376a-3p Inhibits Tumor Growth in Vivo
3. Discussion
4. Materials and Methods
4.1. Tissue Samples and Cell Culture
4.2. Transfection of Osteosarcoma Cell Lines
4.3. RNA Extraction, cDNA Synthesis, and RT-qPCR
4.4. Proliferation Assay
4.5. Colony Formation Assay
4.6. Cell Cycle Analysis
4.7. Apoptosis Detection
4.8. Chicken Chorioallantoic Membrane (CAM) Assay
4.9. Data Analysis and Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Damron, T.A.; Ward, W.G.; Stewart, A. Osteosarcoma, chondrosarcoma, and Ewing’s sarcoma: National Cancer Data Base Report. Clin. Orthop. Relat. Res. 2007, 459, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Mirabello, L.; Troisi, R.J.; Savage, S.A. Osteosarcoma incidence and survival rates from 1973 to 2004: Data from the Surveillance, Epidemiology, and End Results Program. Cancer 2009, 115, 1531–1543. [Google Scholar] [CrossRef] [PubMed]
- Luetke, A.; Meyers, P.A.; Lewis, I.; Juergens, H. Osteosarcoma treatment—Where do we stand? A state of the art review. Cancer Treat. Rev. 2014, 40, 523–532. [Google Scholar] [CrossRef] [PubMed]
- Ottaviani, G.; Jaffe, N. The etiology of osteosarcoma. Cancer Treat. Res. 2009, 152, 15–32. [Google Scholar] [CrossRef]
- Ottaviani, G.; Jaffe, N. The epidemiology of osteosarcoma. Cancer Treat. Res. 2009, 152, 3–13. [Google Scholar] [CrossRef]
- Bielack, S.S.; Smeland, S.; Whelan, J.S.; Marina, N.; Jovic, G.; Hook, J.M.; Krailo, M.D.; Gebhardt, M.; Papai, Z.; Meyer, J.; et al. Methotrexate, Doxorubicin, and Cisplatin (MAP) Plus Maintenance Pegylated Interferon Alfa-2b Versus MAP Alone in Patients With Resectable High-Grade Osteosarcoma and Good Histologic Response to Preoperative MAP: First Results of the EURAMOS-1 Good Response Randomized Controlled Trial. J. Clin. Oncol. 2015, 33, 2279–2287. [Google Scholar]
- Chakraborty, C.; Sharma, A.R.; Sharma, G.; Doss, C.G.P.; Lee, S.S. Therapeutic miRNA and siRNA: Moving from Bench to Clinic as Next Generation Medicine. Mol. Ther. Nucleic Acids 2017, 8, 132–143. [Google Scholar] [CrossRef]
- Garzon, R.; Calin, G.A.; Croce, C.M. MicroRNAs in Cancer. Annu. Rev. Med. 2009, 60, 167–179. [Google Scholar] [CrossRef]
- Chan, J.A.; Krichevsky, A.M.; Kosik, K.S. MicroRNA-21 is an antiapoptotic factor in human glioblastoma cells. Cancer Res. 2005, 65, 6029–6033. [Google Scholar] [CrossRef]
- Michael, M.Z.; O’ Connor, S.M.; Holst Pellekaan, N.G.; Young, G.P.; James, R.J. Reduced accumulation of specific microRNAs in colorectal neoplasia. Mol. Cancer Res. 2003, 1, 882–891. [Google Scholar]
- Takamizawa, J.; Konishi, H.; Yanagisawa, K.; Tomida, S.; Osada, H.; Endoh, H.; Harano, T.; Yatabe, Y.; Nagino, M.; Nimura, Y.; et al. Reduced expression of the let-7 microRNAs in human lung cancers in association with shortened postoperative survival. Cancer Res. 2004, 64, 3753–3756. [Google Scholar] [CrossRef] [PubMed]
- Raza, U.; Zhang, J.D.; Sahin, O. MicroRNAs: Master regulators of drug resistance, stemness, and metastasis. J. Mol. Med. 2014, 92, 321–336. [Google Scholar] [CrossRef] [PubMed]
- Ji, F.; Zhang, H.; Wang, Y.; Li, M.; Xu, W.; Kang, Y.; Wang, Z.; Wang, Z.; Cheng, P.; Tong, D.; et al. MicroRNA-133a, downregulated in osteosarcoma, suppresses proliferation and promotes apoptosis by targeting Bcl-xL and Mcl-1. Bone 2013, 56, 220–226. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Cai, L.; Li, H. miR185 regulates the growth of osteosarcoma cells via targeting Hexokinase 2. Mol. Med. Rep. 2019. [Google Scholar] [CrossRef]
- Pei, Y.; Yao, Q.; Li, Y.; Zhang, X.; Xie, B. microRNA-211 regulates cell proliferation, apoptosis and migration/invasion in human osteosarcoma via targeting EZRIN. Cell. Mol. Biol. Lett. 2019, 24, 48. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.; Li, Z.; Yu, Z.; Yan, F.; Liu, Y.; Lu, X.; Yang, W. MicroRNA-33b suppresses migration and invasion by targeting c-Myc in osteosarcoma cells. PLoS ONE 2014, 9, e115300. [Google Scholar] [CrossRef]
- Chu, Y.; Hu, X.; Wang, G.; Wang, Z.; Wang, Y. Downregulation of miR-136 promotes the progression of osteosarcoma and is associated with the prognosis of patients with osteosarcoma. Oncol. Lett. 2019, 17, 5210–5218. [Google Scholar] [CrossRef]
- Xu, K.; Xiong, W.; Zhao, S.; Wang, B. MicroRNA-106b serves as a prognostic biomarker and is associated with cell proliferation, migration, and invasion in osteosarcoma. Oncol. Lett. 2019, 18, 3342–3348. [Google Scholar] [CrossRef]
- Wang, H.; Zhao, F.; Cai, S.; Pu, Y. MiR-193a regulates chemoresistance of human osteosarcoma cells via repression of IRS2. J. Bone Oncol. 2019, 17, 100241. [Google Scholar] [CrossRef]
- Yu, W.C.; Chen, H.H.; Qu, Y.Y.; Xu, C.W.; Yang, C.; Liu, Y. MicroRNA-221 promotes cisplatin resistance in osteosarcoma cells by targeting PPP2R2A. Biosci. Rep. 2019, 39. [Google Scholar] [CrossRef]
- Huang, C.; Wang, Q.; Ma, S.; Sun, Y.; Vadamootoo, A.S.; Jin, C. A four serum-miRNA panel serves as a potential diagnostic biomarker of osteosarcoma. Int. J. Clin. Oncol. 2019, 24, 976–982. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Yan, P.; Wang, J.; Zhang, Y.; Zhang, M.; Wang, Z.; Fu, Q.; Liang, W. Clinical significance of tumor miR-21, miR-221, miR-143, and miR-106a as biomarkers in patients with osteosarcoma. Int. J. Biol. Markers 2019, 34, 184–193. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Gao, D.Y.; Huang, L. In vivo delivery of miRNAs for cancer therapy: Challenges and strategies. Adv. Drug Deliv. Rev. 2015, 81, 128–141. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Rana, T.M. Therapeutic targeting of microRNAs: Current status and future challenges. Nat. Rev. Drug Discov. 2014, 13, 622–638. [Google Scholar] [CrossRef] [PubMed]
- Fellenberg, J.; Saehr, H.; Lehner, B.; Depeweg, D. A microRNA signature differentiates between giant cell tumor derived neoplastic stromal cells and mesenchymal stem cells. Cancer Lett. 2012, 321, 162–168. [Google Scholar] [CrossRef]
- Gao, X.; Wang, X.; Cai, K.; Wang, W.; Ju, Q.; Yang, X.; Wang, H.; Wu, H. MicroRNA-127 is a tumor suppressor in human esophageal squamous cell carcinoma through the regulation of oncogene FMNL3. Eur. J. Pharmacol. 2016, 791, 603–610. [Google Scholar] [CrossRef]
- Liu, X.; Meng, Z.; Xing, Y.; Zhong, Q.; Zhang, X.; Qu, J. MiR-127 inhibits ovarian cancer migration and invasion by up-regulating ITGA6. Minerva Med. 2019. [Google Scholar] [CrossRef]
- Wang, L.; Wang, X.; Jiang, X. miR-127 suppresses gastric cancer cell migration and invasion via targeting Wnt7a. Oncol. Lett. 2019, 17, 3219–3226. [Google Scholar] [CrossRef]
- Wang, Y.; Cong, W.; Wu, G.; Ju, X.; Li, Z.; Duan, X.; Wang, X.; Gao, H. MiR-376a suppresses the proliferation and invasion of non-small-cell lung cancer by targeting c-Myc. Cell Biol. Int. 2018, 42, 25–33. [Google Scholar] [CrossRef]
- Yao, S.; Liu, Y.; Yao, Z.; Zhao, Y.; Wang, H.; Xu, Y.; Zhang, J.; Li, J.; Yang, S. MicroRNA-376a regulates cell proliferation and apoptosis by targeting forkhead box protein P2 in lymphoma. Oncol. Lett. 2018, 16, 3169–3176. [Google Scholar] [CrossRef]
- Yu, Y.; Liu, L.; Ma, R.; Gong, H.; Xu, P.; Wang, C. MicroRNA-127 is aberrantly downregulated and acted as a functional tumor suppressor in human pancreatic cancer. Tumour Biol. 2016, 37, 14249–14257. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Chen, Y.; Wang, H.; Zheng, X.; Li, C.; Han, Z. miR-376a inhibits breast cancer cell progression by targeting neuropilin-1 NR. Onco Targets Ther. 2018, 11, 5293–5302. [Google Scholar] [CrossRef] [PubMed]
- Kunz, P.; Schenker, A.; Sahr, H.; Lehner, B.; Fellenberg, J. Optimization of the chicken chorioallantoic membrane assay as reliable in vivo model for the analysis of osteosarcoma. PLoS ONE 2019, 14, e0215312. [Google Scholar] [CrossRef] [PubMed]
- Moro, M.; Bertolini, G.; Caserini, R.; Borzi, C.; Boeri, M.; Fabbri, A.; Leone, G.; Gasparini, P.; Galeone, C.; Pelosi, G.; et al. Establishment of patient derived xenografts as functional testing of lung cancer aggressiveness. Sci. Rep. 2017, 7, 6689. [Google Scholar] [CrossRef] [PubMed]
- Herr, I.; Sahr, H.; Zhao, Z.; Yin, L.; Omlor, G.; Lehner, B.; Fellenberg, J. MiR-127 and miR-376a act as tumor suppressors by in vivo targeting of COA1 and PDIA6 in giant cell tumor of bone. Cancer Lett. 2017, 409, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Tang, L.; Wu, H.; Wang, K.; Gu, D. MiR-127-3p inhibits cell growth and invasiveness by targeting ITGA6 in human osteosarcoma. IUBMB Life 2018, 70, 411–419. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Cheng, L.; Chen, J.; Lu, W.; Wang, P. miR-376a inhibits the proliferation and invasion of osteosarcoma by targeting FBXO11. Hum. Cell 2019, 32, 390–396. [Google Scholar] [CrossRef]
- Zhang, J.; Hou, W.; Chai, M.; Zhao, H.; Jia, J.; Sun, X.; Zhao, B.; Wang, R. MicroRNA-127-3p inhibits proliferation and invasion by targeting SETD8 in human osteosarcoma cells. Biochem. Biophys. Res. Commun. 2016, 469, 1006–1011. [Google Scholar] [CrossRef]
- McMurray, E.N.; Schmidt, J.V. Identification of imprinting regulators at the Meg3 differentially methylated region. Genomics 2012, 100, 184–194. [Google Scholar] [CrossRef]
- Lehner, B.; Kunz, P.; Saehr, H.; Fellenberg, J. Epigenetic silencing of genes and microRNAs within the imprinted Dlk1-Dio3 region at human chromosome 14.32 in giant cell tumor of bone. BMC Cancer 2014, 14, 495. [Google Scholar] [CrossRef]
- Zheng, Y.; Chen, H.; Yin, M.; Ye, X.; Chen, G.; Zhou, X.; Yin, L.; Zhang, C.; Ding, B. MiR-376a and histone deacetylation 9 form a regulatory circuitry in hepatocellular carcinoma. Cell. Physiol. Biochem. 2015, 35, 729–739. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Jiang, H.; Hsu, H.T.; Qian, L.; Fu, Q.; Shen, M.; Chen, S.; Yang, T. MicroRNA-127 inhibits cell proliferation via targeting Kif3b in pancreatic beta cells. Aging (Albany NY USA) 2019, 11, 1342–1355. [Google Scholar] [CrossRef]
- Bi, L.; Yang, Q.; Yuan, J.; Miao, Q.; Duan, L.; Li, F.; Wang, S. MicroRNA-127-3p acts as a tumor suppressor in epithelial ovarian cancer by regulating the BAG5 gene. Oncol. Rep. 2016, 36, 2563–2570. [Google Scholar] [CrossRef] [PubMed]
- Brooks, D.L.; Schwab, L.P.; Krutilina, R.; Parke, D.N.; Sethuraman, A.; Hoogewijs, D.; Schorg, A.; Gotwald, L.; Fan, M.; Wenger, R.H.; et al. ITGA6 is directly regulated by hypoxia-inducible factors and enriches for cancer stem cell activity and invasion in metastatic breast cancer models. Mol. Cancer 2016, 15, 26. [Google Scholar] [CrossRef] [PubMed]
- Milite, C.; Feoli, A.; Viviano, M.; Rescigno, D.; Cianciulli, A.; Balzano, A.L.; Mai, A.; Castellano, S.; Sbardella, G. The emerging role of lysine methyltransferase SETD8 in human diseases. Clin. Epigenetics 2016, 8, 102. [Google Scholar] [CrossRef] [PubMed]
- Bruchmann, A.; Roller, C.; Walther, T.V.; Schafer, G.; Lehmusvaara, S.; Visakorpi, T.; Klocker, H.; Cato, A.C.; Maddalo, D. Bcl-2 associated athanogene 5 (Bag5) is overexpressed in prostate cancer and inhibits ER-stress induced apoptosis. BMC Cancer 2013, 13, 96. [Google Scholar] [CrossRef]
- Yue, X.; Zhao, Y.; Huang, G.; Li, J.; Zhu, J.; Feng, Z.; Hu, W. A novel mutant p53 binding partner BAG5 stabilizes mutant p53 and promotes mutant p53 GOFs in tumorigenesis. Cell Discov. 2016, 2, 16039. [Google Scholar] [CrossRef]
- Fellenberg, J.; Sahr, H.; Kunz, P.; Zhao, Z.; Liu, L.; Tichy, D.; Herr, I. Restoration of miR-127-3p and miR-376a-3p counteracts the neoplastic phenotype of giant cell tumor of bone derived stromal cells by targeting COA1, GLE1 and PDIA6. Cancer Lett. 2016, 371, 134–141. [Google Scholar] [CrossRef]
- Jimenez-Hernandez, L.E.; Vazquez-Santillan, K.; Castro-Oropeza, R.; Martinez-Ruiz, G.; Munoz-Galindo, L.; Gonzalez-Torres, C.; Cortes-Gonzalez, C.C.; Victoria-Acosta, G.; Melendez-Zajgla, J.; Maldonado, V. NRP1-positive lung cancer cells possess tumor-initiating properties. Oncol. Rep. 2018, 39, 349–357. [Google Scholar] [CrossRef]
- Kaiser, B.K.; Yim, D.; Chow, I.T.; Gonzalez, S.; Dai, Z.; Mann, H.H.; Strong, R.K.; Groh, V.; Spies, T. Disulphide-isomerase-enabled shedding of tumour-associated NKG2D ligands. Nature 2007, 447, 482–486. [Google Scholar] [CrossRef]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fellenberg, J.; Lehner, B.; Saehr, H.; Schenker, A.; Kunz, P. Tumor Suppressor Function of miR-127-3p and miR-376a-3p in Osteosarcoma Cells. Cancers 2019, 11, 2019. https://doi.org/10.3390/cancers11122019
Fellenberg J, Lehner B, Saehr H, Schenker A, Kunz P. Tumor Suppressor Function of miR-127-3p and miR-376a-3p in Osteosarcoma Cells. Cancers. 2019; 11(12):2019. https://doi.org/10.3390/cancers11122019
Chicago/Turabian StyleFellenberg, Joerg, Burkhard Lehner, Heiner Saehr, Astrid Schenker, and Pierre Kunz. 2019. "Tumor Suppressor Function of miR-127-3p and miR-376a-3p in Osteosarcoma Cells" Cancers 11, no. 12: 2019. https://doi.org/10.3390/cancers11122019
APA StyleFellenberg, J., Lehner, B., Saehr, H., Schenker, A., & Kunz, P. (2019). Tumor Suppressor Function of miR-127-3p and miR-376a-3p in Osteosarcoma Cells. Cancers, 11(12), 2019. https://doi.org/10.3390/cancers11122019