Migration and Invasion Enhancer 1 Is an NF-ĸB-Inducing Gene Enhancing the Cell Proliferation and Invasion Ability of Human Prostate Carcinoma Cells In Vitro and In Vivo

 ,
,
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Expression of MIEN1 in Prostate Carcinoma Cells and Prostate Tissues
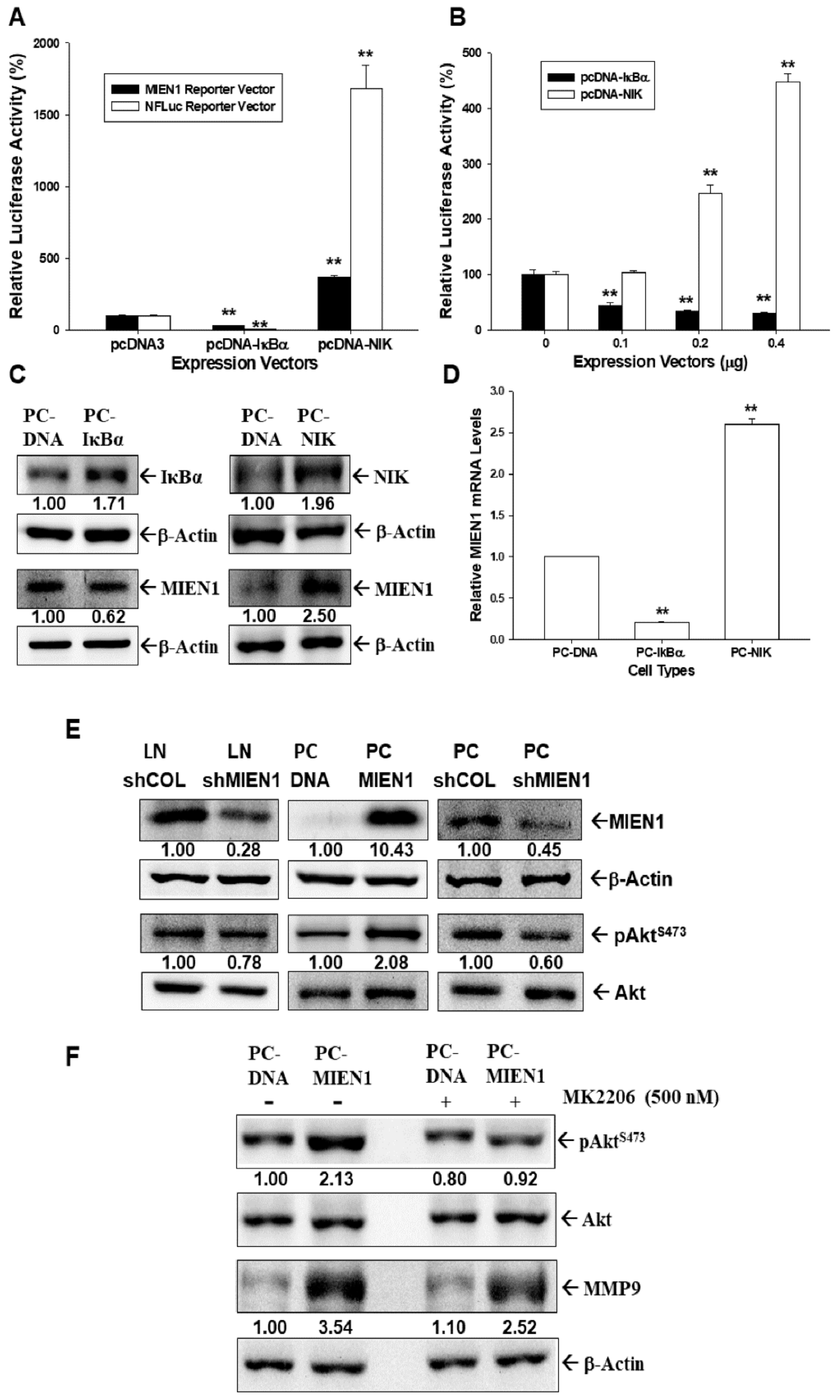
2.2. MIEN1 Is the Downstream Gene of NF-ĸB Signaling and Induces Akt Phosphorylation in Prostate Carcinoma Cells
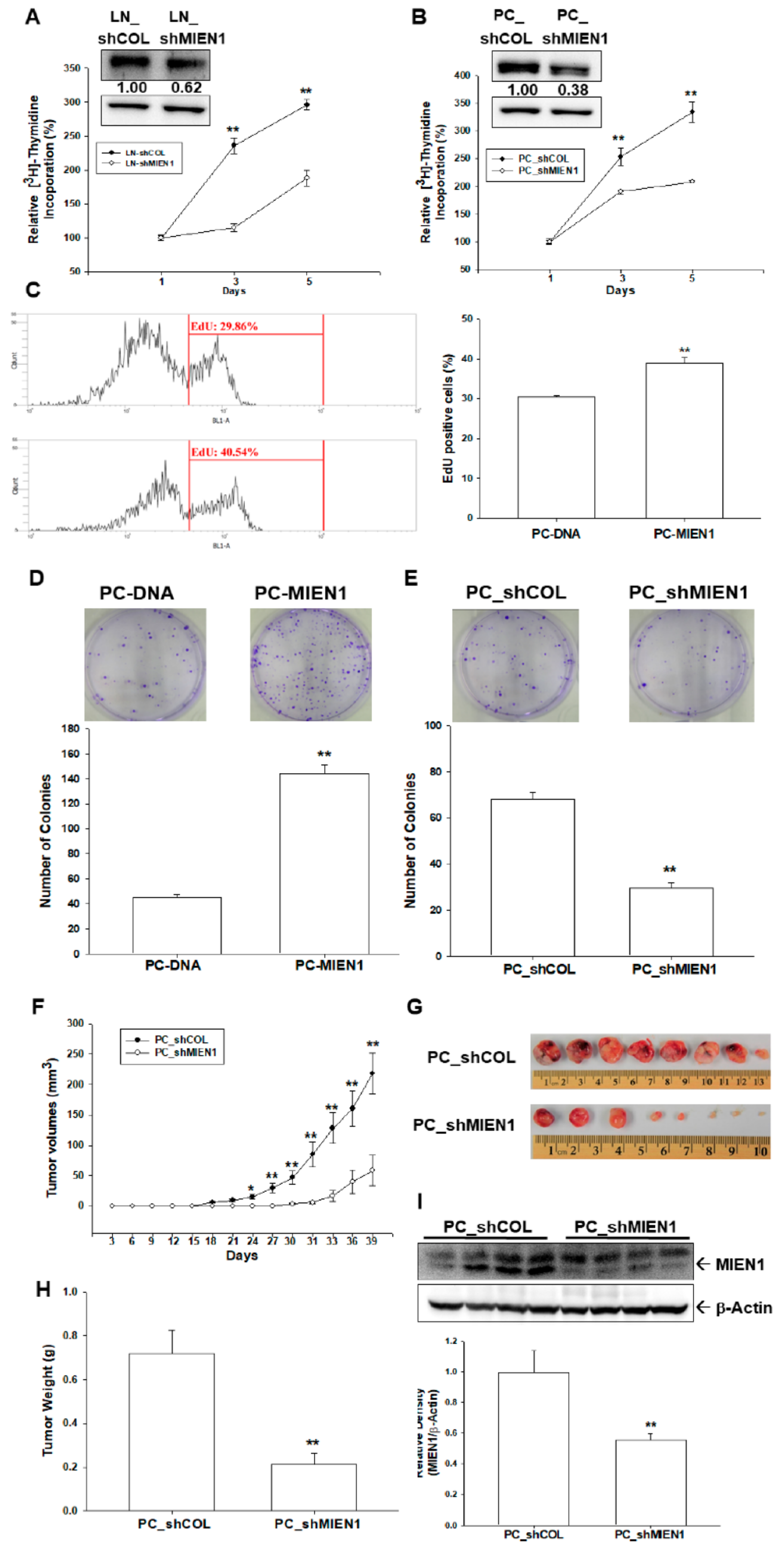
2.3. MIEN1 Enhances Cell Growth of Prostate Carcinoma Cells In Vitro and In Vivo
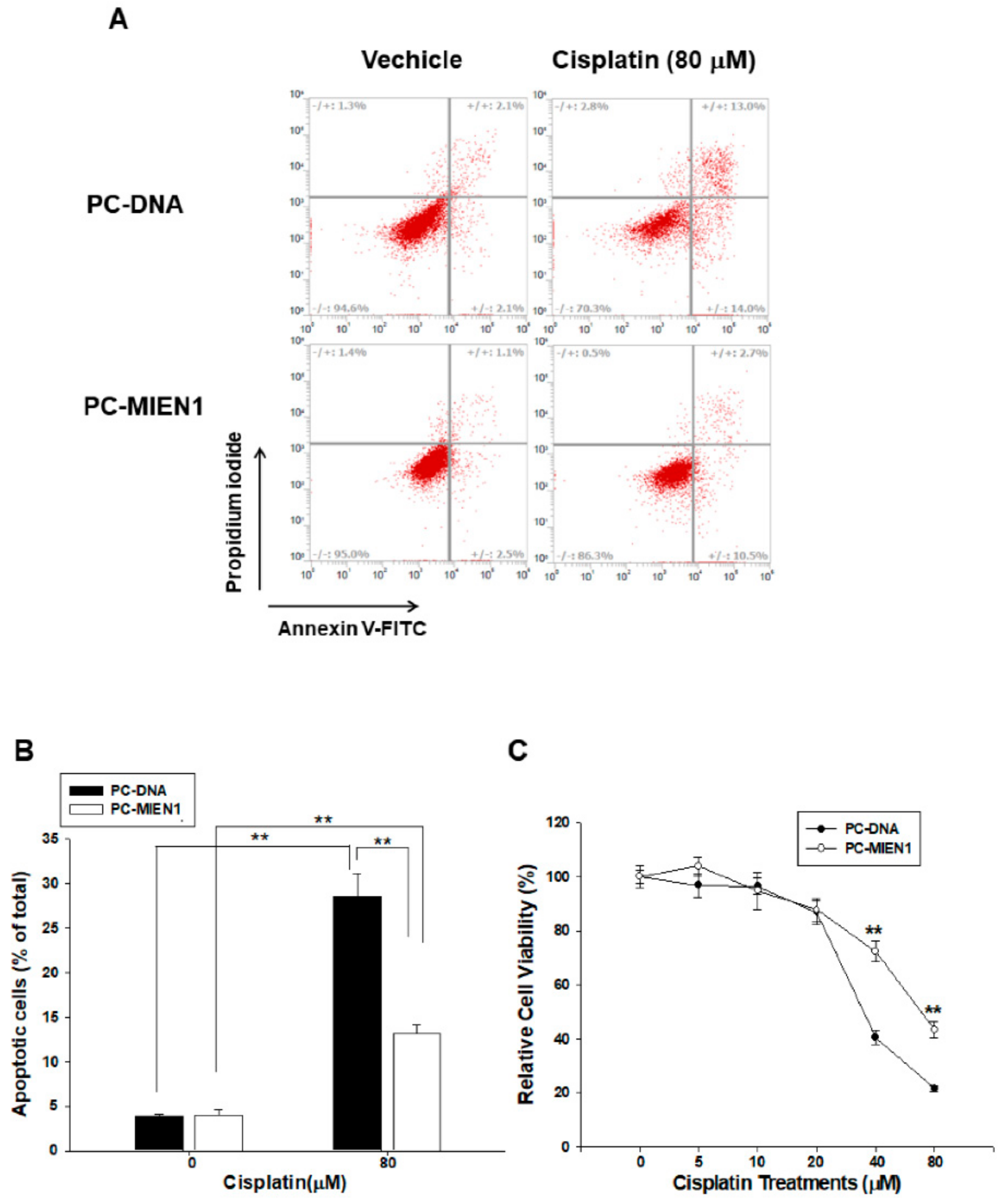
2.4. Ectopic Overexpression of MIEN1 Attenuates Cisplatin-Induced Cell Apoptosis in PC-3 Cells
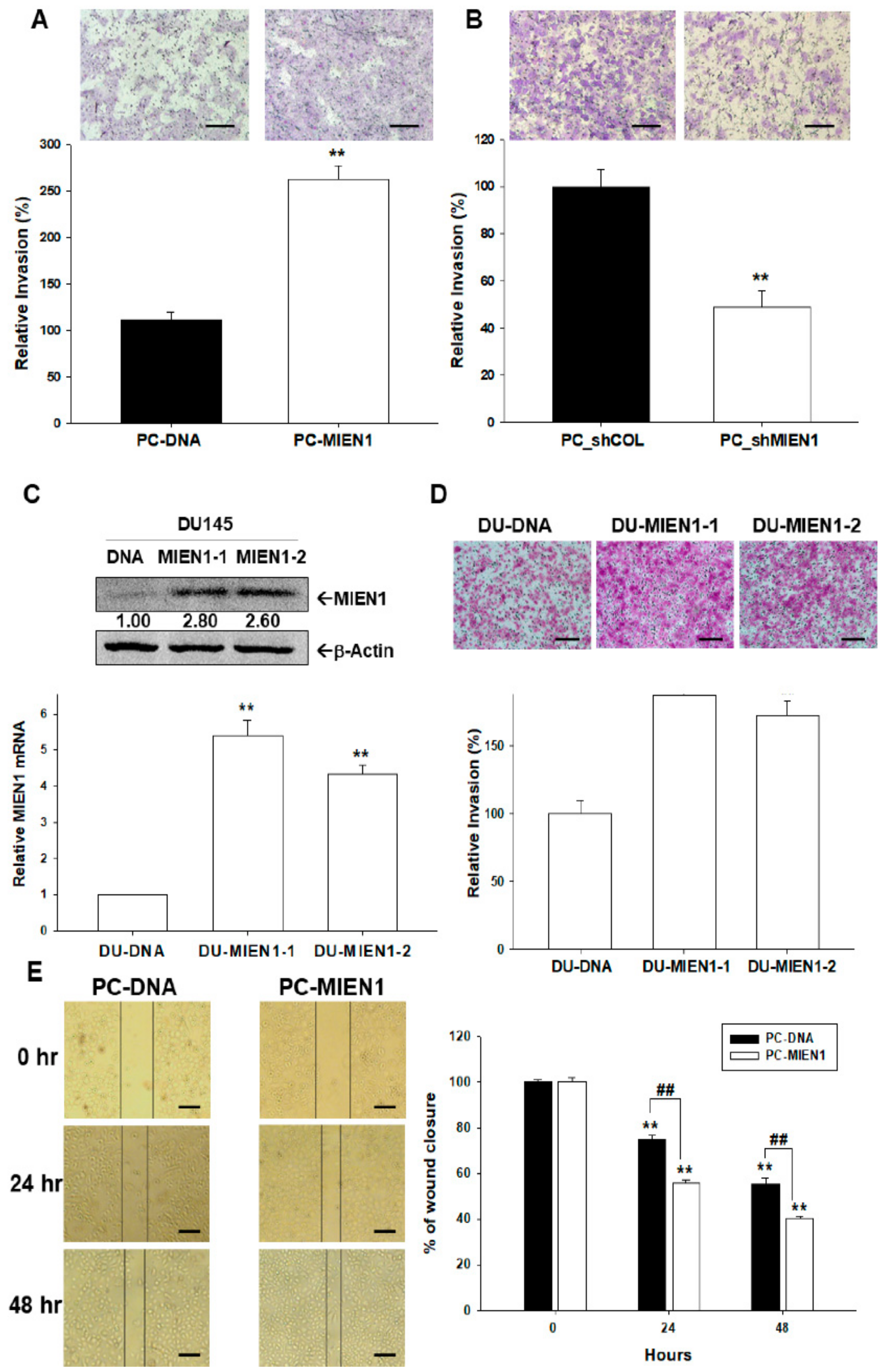
2.5. Effects of MIEN1 on Cell Invasion and Migration in Prostate Carcinoma Cells
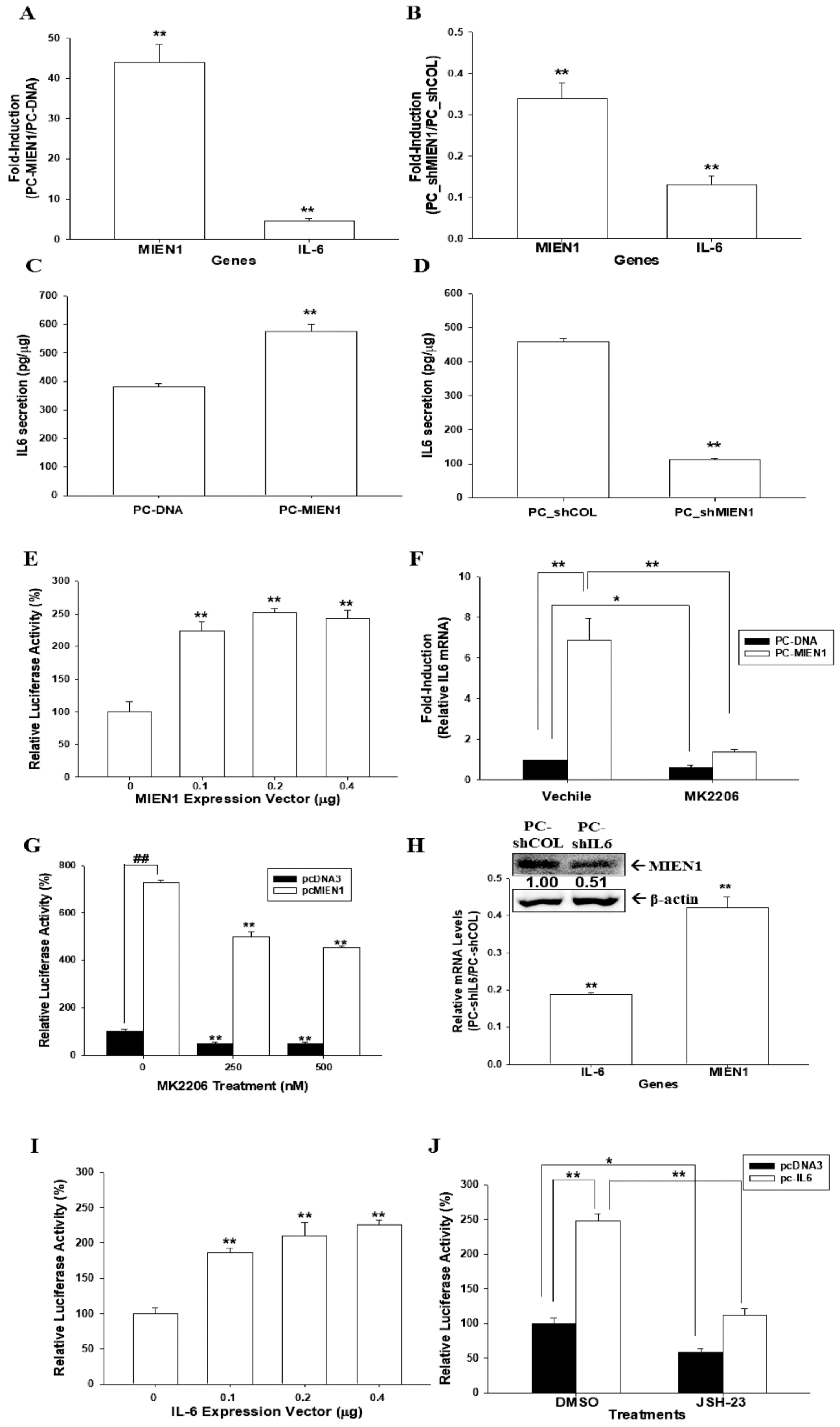
2.6. Regulation between MIEN1 and IL-6 in Prostate Carcinoma PC-3 Cells
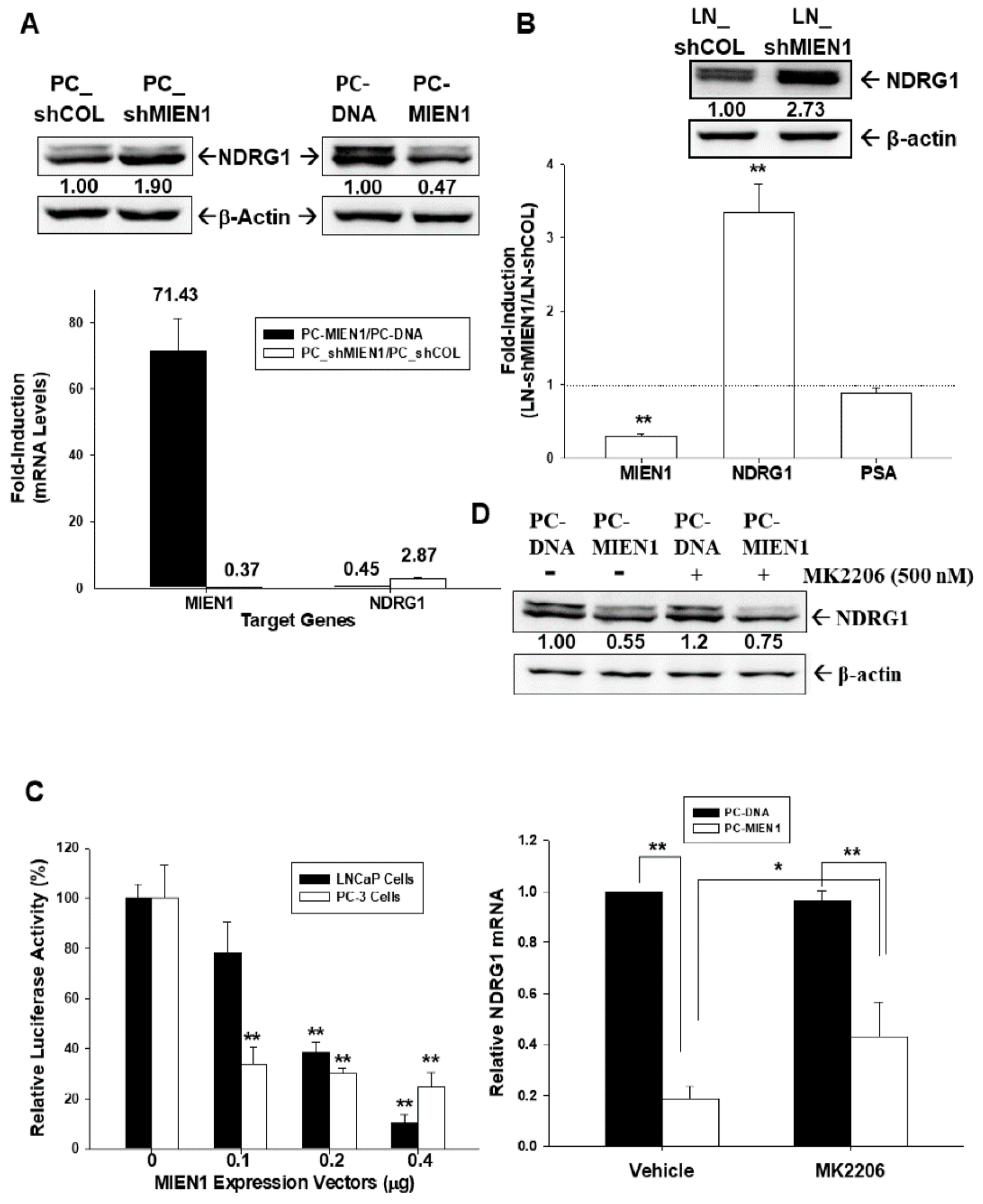
2.7. MIEN1 Downregulates NDRG1 Gene Expression in Prostate Carcinoma Cells
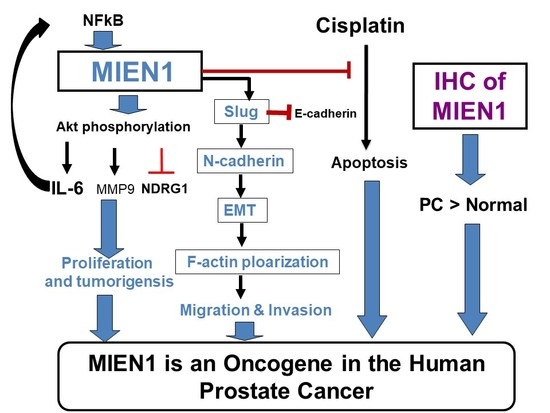
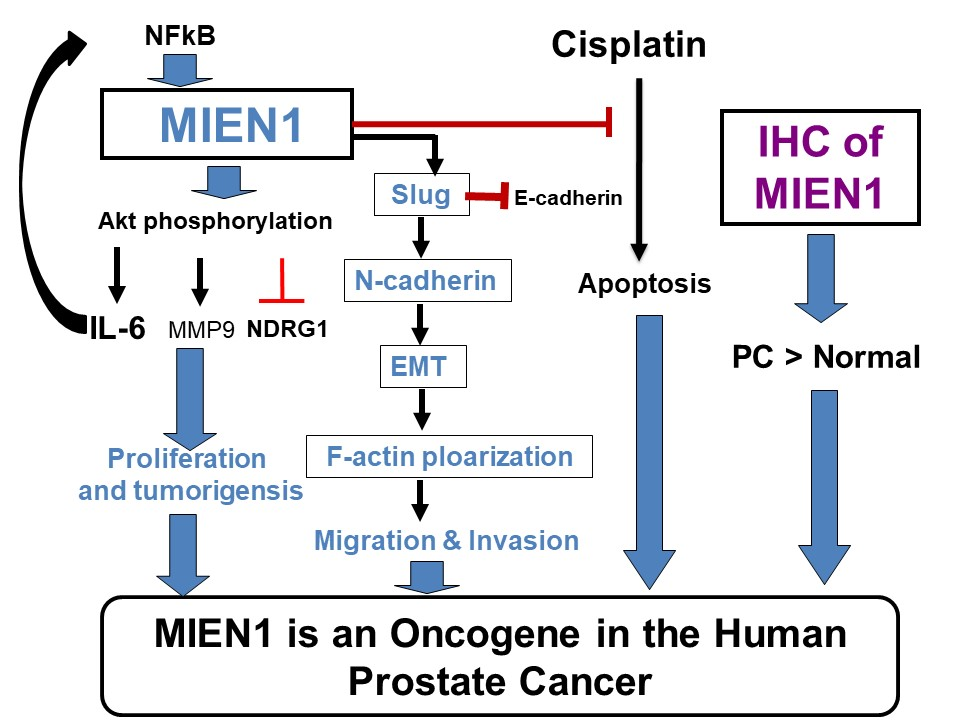
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Chemicals
4.2. Immunohistochemical Assays
4.3. Expression Vector Constructs and Stable Transfection
4.4. Knockdown of MIEN1
4.5. Immunoblot Assays
4.6. Real-Time Reverse Transcriptase Polymerase Chain Reaction (RT-qPCR)
4.7. Thymidine Incorporation Assays
4.8. EdU Staining Proliferation Assay
4.9. MTS Assay
4.10. Colony Formation Assay
4.11. Annexin V-FITC Apoptosis Detection
4.12. Matrigel Invasion Assay
4.13. Cell Migration Assay
4.14. Xenograft Animal Model
4.15. F-Actin Staining
4.16. IL-6 ELISA
4.17. Reporter Vector and Reporter Assays
4.18. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Availability of Data and Materials
References
- Evans, E.E.; Henn, A.D.; Jonason, A.; Paris, M.J.; Schiffhauer, L.M.; Borrello, M.A.; Smith, E.S.; Sahasrabudhe, D.M.; Zauderer, M. C35 (C17orf37) is a novel tumor biomarker abundantly expressed in breast cancer. Mol. Cancer Ther. 2006, 5, 2919–2930. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Katz, E.; Dubois-Marshall, S.; Sims, A.H.; Faratian, D.; Li, J.; Smith, E.S.; Quinn, J.A.; Edward, M.; Meehan, R.R.; Evans, E.E.; et al. A gene on the HER2 amplicon, C35, is an oncogene in breast cancer whose actions are prevented by inhibition of Syk. Br. J. Cancer 2010, 103, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.B.; Zhang, X.F.; Wang, H.B.; Zhang, M.Z. Migration and invasion enhancer 1 (MIEN1) is overexpressed in breast cancer and is a potential new therapeutic molecular target. Genet. Mol. Res. 2017, 16, 6019380. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.; Qi, Y.; Yin, X.; Gao, J. MiR-136 targets MIEN1 and involves the metastasis of colon cancer by suppressing epithelial-to-mesenchymal transition. OncoTargets Ther. 2017, 11, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Rajendiran, S.; Kpetemey, M.; Maji, S.; Gibbs, L.D.; Dasgupta, S.; Mantsch, R.; Hare, R.J.; Vishwanatha, J.K. MIEN1 promotes oral cancer progression and implicates poor overall survival. Cancer Biol. Ther. 2015, 16, 876–885. [Google Scholar] [CrossRef] [PubMed]
- Rajendiran, S.; Gibbs, L.D.; Van Treuren, T.; Klinkebiel, D.L.; Vishwanatha, J.K. MIEN1 is tightly regulated by SINE Alu methylation in its promoter. Oncotarget 2016, 7, 65307–65319. [Google Scholar] [CrossRef]
- Li, D.; Wei, Y.; Wang, D.; Gao, H.; Liu, K. MicroRNA-26b suppresses the metastasis of non-small cell lung cancer by targeting MIEN1 via NF-κB/MMP-9/VEGF pathways. Biochem. Biophys. Res. Commun. 2016, 472, 465–470. [Google Scholar] [CrossRef]
- Leung, T.H.; Wong, S.C.; Chan, K.K.; Chan, D.W.; Cheung, A.N.; Ngan, H.Y. The interaction between C35 and ΔNp73 promotes chemo-resistance in ovarian cancer cells. Br. J. Cancer 2013, 109, 965–975. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Zhang, Z.Y.; Du, W.D.; Ye, L.; Xu, S.; Zuo, X.B.; Zhou, F.S.; Chen, G.; Ma, X.L.; Schneider, M.E.; et al. Association analysis of ERBB2 amplicon genetic polymorphisms and STARD3 expression with risk of gastric cancer in the Chinese population. Gene 2014, 535, 225–232. [Google Scholar] [CrossRef]
- Yang, C.J.; Chang, C.F.; Huang, L.Y.; Chang, Y.Y.; Shen, T.L.; Hsu, C.H. Resonance assignments of human C35 (C17orf37) protein, a novel tumor biomarker. Biomol. NMR Assign. 2010, 4, 191–193. [Google Scholar] [CrossRef]
- Van Treuren, T.; Vishwanatha, J.K. CRISPR deletion of MIEN1 in breast cancer cells. PLoS ONE 2018, 13, e0204976. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, S.; Wasson, L.M.; Rauniyar, N.; Prokai, L.; Borejdo, J.; Vishwanatha, J.K. Novel gene C17orf37 in 17q12 amplicon promotes migration and invasion of prostate cancer cells. Oncogene 2009, 28, 2860–2872. [Google Scholar] [CrossRef] [PubMed]
- Rajendiran, S.; Parwani, A.V.; Hare, R.J.; Dasgupta, S.; Roby, R.K.; Vishwanatha, J.K. MicroRNA-940 suppresses prostate cancer migration and invasion by regulating MIEN1. Mol. Cancer 2014, 13, 250. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hsu, C.H.; Shen, T.L.; Chang, C.F.; Chang, Y.Y.; Huang, L.Y. Solution structure of the oncogenic MIEN1 protein reveals a thioredoxin-like fold with a redox-active motif. PLoS ONE 2012, 7, e52292. [Google Scholar] [CrossRef] [PubMed]
- Culig, Z.; Puhr, M. Interleukin-6 and prostate cancer: Current developments and unsolved questions. Mol. Cell. Endocrinol. 2018, 462, 25–30. [Google Scholar] [CrossRef]
- George, D.J.; Halabi, S.; Shepard, T.F.; Sanford, B.; Vogelzang, N.J.; Small, E.J.; Kantoff, P.W. The prognostic significance of plasma interleukin-6 levels in patients with metastatic hormone-refractory prostate cancer: Results from cancer and leukemia group B 9480. Clin. Cancer Res. 2005, 11, 1815–1820. [Google Scholar] [CrossRef]
- Fang, B.A.; Kovačević, Ž.; Park, K.C.; Kalinowski, D.S.; Jansson, P.J.; Lane, D.J.; Sahni, S.; Richardson, D.R. Molecular functions of the iron-regulated metastasis suppressor, NDRG1, and its potential as a molecular target for cancer therapy. Biochim. Biophys. Acta 2014, 1845, 1–19. [Google Scholar] [CrossRef]
- Li, Y.; Pan, P.; Qiao, P.; Liu, R. Downregulation of N-myc downstream regulated gene 1 caused by the methylation of CpG islands of NDRG1 promoter promotes proliferation and invasion of prostate cancer cells. Int. J. Oncol. 2015, 47, 1001–1008. [Google Scholar] [CrossRef]
- Sharma, A.; Mendonca, J.; Ying, J.; Kim, H.; Verdone, J.E.; Zarif, J.C.; Carducci, M.; Hammers, H.; Pienta, K.J.; Kachhap, S. The prostate metastasis suppressor gene NDRG1 differentially regulates cell motility and invasion. Mol. Oncol. 2017, 11, 655–669. [Google Scholar] [CrossRef]
- Cahill, C.M.; Rogers, J.T. Interleukin (IL) 1beta induction of IL-6 is mediated by a novel phosphatidylinositol 3-kinase-dependent AKT/IkappaB kinase alpha pathway targeting activator protein-1. J. Biol. Chem. 2008, 283, 25900–25912. [Google Scholar] [CrossRef]
- Sun, J.; Zhang, D.; Bae, D.H.; Sahni, S.; Jansson, P.; Zhang, Y.; Zhao, Q.; Yue, F.; Zheng, M.; Kovacevic, A.; et al. Metastasis suppressor, NDRG1, mediates its activity through signaling pathways and molecular motors. Carcinogenesis 2013, 34, 1943–1954. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.Q.; Yin, K.; Zhu, S.; Zhang, L.; Wen, P.E.; Li, C.L.; Zhang, D.B.; Liu, M.; Yan, G. Inhibition of C35 gene expression by small interfering RNA induces apoptosis of breast cancer cells. Biosci. Trends 2010, 4, 254–259. [Google Scholar] [PubMed]
- Kpetemey, M.; Dasgupta, S.; Rajendiran, S.; Das, S.; Gibbs, L.D.; Shetty, P.; Gryczynski, Z.; Vishwanatha, J.K. MIEN1, a novel interactor of Annexin A2, promotes tumor cell migration by enhancing AnxA2 cell surface expression. Mol. Cancer 2015, 14, 156. [Google Scholar] [CrossRef] [PubMed]
- Kpetemey, M.; Chaudhary, P.; Van Treuren, T.; Vishwanatha, J.K. MIEN1 drives breast tumor cell migration by regulating cytoskeletal-focal adhesion dynamics. Oncotarget 2016, 7, 54913–54924. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Tsui, K.H.; Feng, T.H.; Hsieh, W.C.; Chang, P.L.; Juang, H.H. Expression of interleukin-6 is downregulated by 17-(allylamino)-17-demethoxygeldanamycin in human prostatic carcinoma cells. Acta Pharmacol. Sin. 2008, 29, 1334–1341. [Google Scholar] [CrossRef][Green Version]
- Chiang, K.C.; Tsui, K.H.; Chung, L.C.; Yeh, C.N.; Chen, W.T.; Chang, P.L.; Juang, H.H. Celastrol blocks interleukin-6 gene expression via downregulation of NF-κB in prostate carcinoma cells. PLoS ONE 2014, 9, e93151. [Google Scholar] [CrossRef]
- Zegeye, M.M.; Lindkvist, M.; Fälker, K.; Kumawat, A.K.; Paramel, G.; Grenegård, M.; Sirsjö, A.; Ljungberg, L.U. Activation of the JAK/STAT3 and PI3K/AKT pathways are crucial for IL-6 trans-signaling-mediated pro-inflammatory response in human vascular endothelial cells. Cell Commun. Signal. 2018, 16, 55. [Google Scholar] [CrossRef]
- Culig, Z. Proinflammatory cytokine interleukin-6 in prostate carcinogenesis. Am. J. Clin. Exp. Urol. 2014, 2, 231–238. [Google Scholar]
- Wang, L.; Walia, B.; Evans, J.; Gewirtz, A.T.; Merlin, D.; Sitaraman, S.V. IL-6 induces NF-kappa B activation in the intestinal epithelia. J. Immunol. 2003, 171, 3194–3201. [Google Scholar] [CrossRef]
- Park, K.C.; Paluncic, J.; Kovacevic, Z.; Richardson, D.R. Pharmacological targeting and the diverse functions of the metastasis suppressor, NDRG1, in cancer. Free Radic. Biol Med. 2019, S0891–5849(19)30256–30264. [Google Scholar] [CrossRef]
- Chung, L.C.; Chiang, K.C.; Feng, T.H.; Chang, K.S.; Chuang, S.T.; Chen, Y.J.; Tsui, K.H.; Lee, J.C. Caffeic acid phenethyl ester upregulates N-myc downstream regulated gene 1 via ERK pathway to inhibit human oral cancer cell growth in vitro and in vivo. Mol. Nutr. Food Res. 2017, 61, 1600842. [Google Scholar] [CrossRef] [PubMed]
- Dixon, K.M.; Lui, G.Y.; Kovačević, Ž.; Zhang, D.; Yao, M.; Chen, Z.; Dong, Q.; Assinder, S.J.; Richardson, D.R. Dp44mT targets the AKT, TGF-b and ERK pathways via the metastasis suppressor NDRG1 in normal prostate epithelial cells and prostate cancer cells. Br. J. Cancer 2013, 108, 409–419. [Google Scholar] [CrossRef]
- Tsui, K.H.; Chung, L.C.; Feng, T.H.; Lee, T.Y.; Chang, P.L.; Chen, W.T.; Juang, H.H. Divergent effect of liver X receptor agonists on prostate-specific antigen expression is dependent on androgen receptor in prostate carcinoma cells. Prostate 2015, 75, 603–615. [Google Scholar] [CrossRef]
- Tsui, K.H.; Lin, Y.H.; Chung, L.C.; Chuang, S.T.; Feng, T.H.; Chiang, K.C.; Chang, P.L.; Yen, C.L.; Juang, H.H. Prostate-derived ets factor represses tumorigenesis and modulates epithelial-to-mesenchymal transition in bladder carcinoma cells. Cancer Lett. 2016, 375, 142–151. [Google Scholar] [CrossRef]
- Tsui, K.H.; Chiang, K.C.; Lin, Y.H.; Chang, K.S.; Feng, T.H.; Juang, H.H. BTG2 is a tumor suppressor gene upregulated by p53 and PTEN in human bladder carcinoma cells. Cancer Med. 2018, 7, 184–195. [Google Scholar] [CrossRef]
- Tsui, K.H.; Hou, C.P.; Chang, K.S.; Lin, Y.H.; Feng, T.H.; Chen, C.C.; Shin, Y.S.; Juang, H.H. Metallothionein 3 is a hypoxia-upregulated oncogene enhancing cell invasion and tumorigenesis in human bladder carcinoma cells. Int. J. Mol. Sci. 2019, 20, 980. [Google Scholar] [CrossRef]
- Lee, J.C.; Chiang, K.C.; Feng, T.H.; Chen, Y.J.; Chuang, S.T.; Tsui, K.H.; Chung, L.C.; Juang, H.H. The iron chelator, Dp44mT, effectively inhibit human oral squamous cell carcinoma cell growth in vitro and in vivo. Int. J. Mol. Sci. 2016, 17, 1435. [Google Scholar] [CrossRef] [PubMed]
- Tsui, K.H.; Chang, Y.L.; Yang, P.S.; Hou, C.P.; Lin, Y.H.; Lin, B.W.; Feng, T.H.; Juang, H.H. The inhibitory effects of capillarisin on cell proliferation and invasion of prostate carcinoma cells. Cell Prolif. 2018, 51, e12429. [Google Scholar] [CrossRef] [PubMed]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, K.-S.; Tsui, K.-H.; Lin, Y.-H.; Hou, C.-P.; Feng, T.-H.; Juang, H.-H. Migration and Invasion Enhancer 1 Is an NF-ĸB-Inducing Gene Enhancing the Cell Proliferation and Invasion Ability of Human Prostate Carcinoma Cells In Vitro and In Vivo. Cancers 2019, 11, 1486. https://doi.org/10.3390/cancers11101486
Chang K-S, Tsui K-H, Lin Y-H, Hou C-P, Feng T-H, Juang H-H. Migration and Invasion Enhancer 1 Is an NF-ĸB-Inducing Gene Enhancing the Cell Proliferation and Invasion Ability of Human Prostate Carcinoma Cells In Vitro and In Vivo. Cancers. 2019; 11(10):1486. https://doi.org/10.3390/cancers11101486
Chicago/Turabian StyleChang, Kang-Shuo, Ke-Hung Tsui, Yu-Hsiang Lin, Chen-Pang Hou, Tsui-Hsia Feng, and Horng-Heng Juang. 2019. "Migration and Invasion Enhancer 1 Is an NF-ĸB-Inducing Gene Enhancing the Cell Proliferation and Invasion Ability of Human Prostate Carcinoma Cells In Vitro and In Vivo" Cancers 11, no. 10: 1486. https://doi.org/10.3390/cancers11101486
APA StyleChang, K.-S., Tsui, K.-H., Lin, Y.-H., Hou, C.-P., Feng, T.-H., & Juang, H.-H. (2019). Migration and Invasion Enhancer 1 Is an NF-ĸB-Inducing Gene Enhancing the Cell Proliferation and Invasion Ability of Human Prostate Carcinoma Cells In Vitro and In Vivo. Cancers, 11(10), 1486. https://doi.org/10.3390/cancers11101486