Development of Microfluidic Stretch System for Studying Recovery of Damaged Skeletal Muscle Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. PDMS Device Design and Fabrication
2.2. Cell Culture
2.3. Stretcher Design
2.4. Creating Damage Model of Muscles Cells
2.5. Recovery of Damaged Muscle Cells
2.6. Staining and Image Processing
3. Results
3.1. Confirmation of Myotube Formation in a Microfluidic Channel
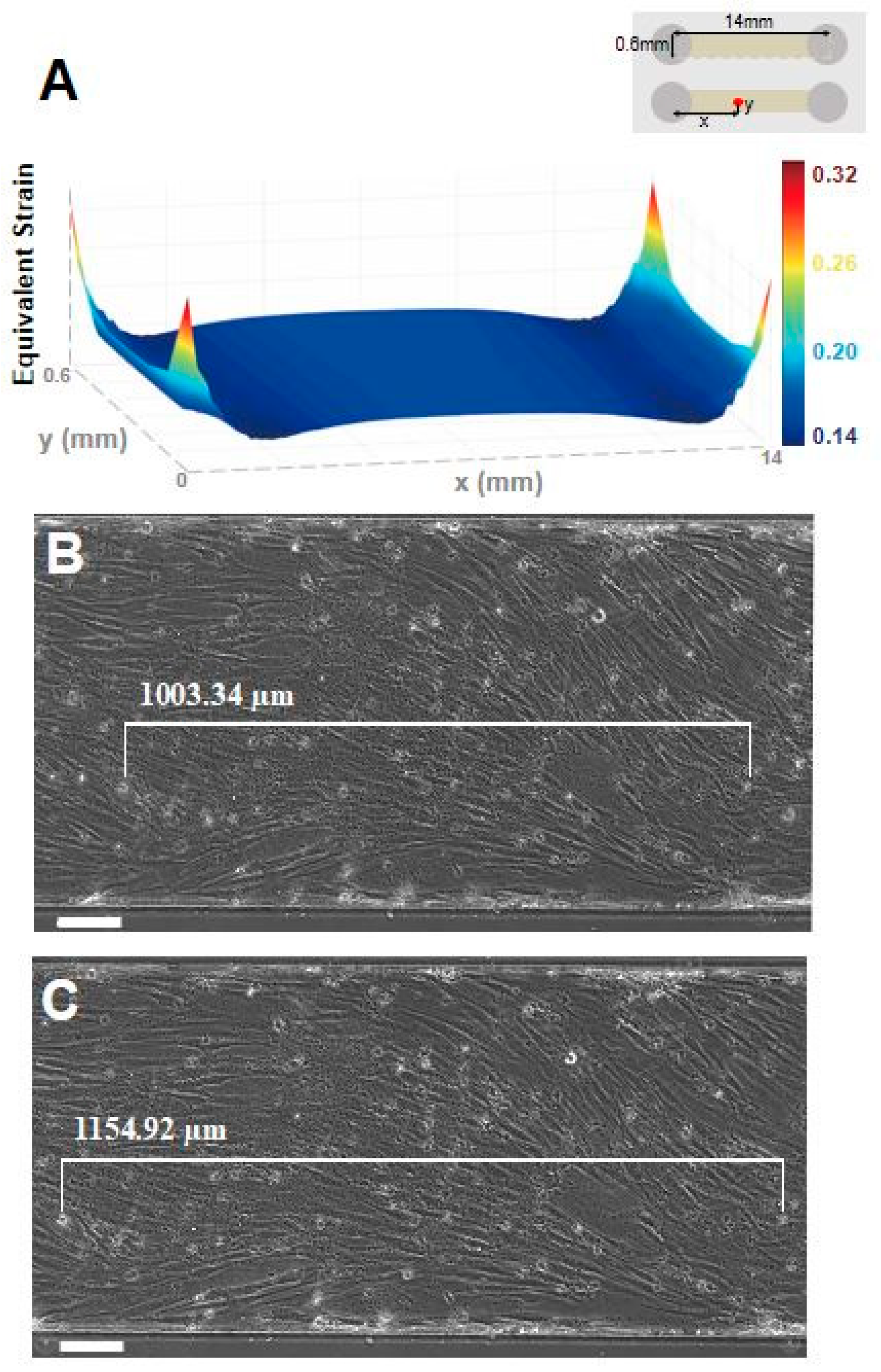
3.2. Stretcher Simulation and Validation
3.3. Damage Confirmation
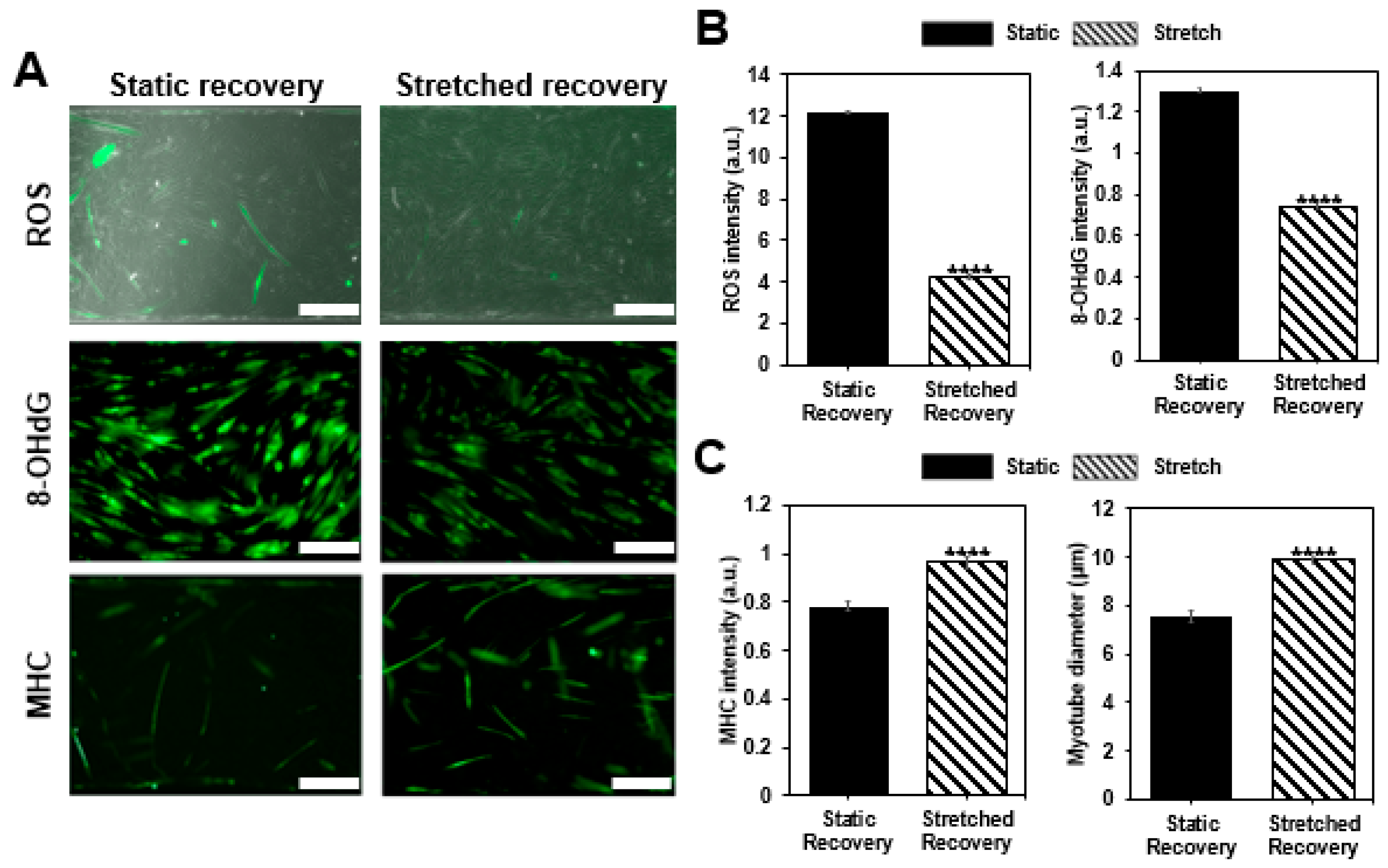
3.4. Recovery Confirmation
4. Discussions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Tero, A.H.; Järvinen, M.J.; Kalimo, H. Regeneration of injured skeletal muscle after the injury. Muscles Ligaments Tendons J. 2013, 3, 337. [Google Scholar]
- Ekstrand, J.; Hagglund, M.; Walden, M. Epidemiology of muscle injuries in professional football (soccer). Am. J. Sports Med. 2011, 39, 1226–1232. [Google Scholar] [CrossRef] [PubMed]
- Huard, J.; Li, Y.; Fu, F.H. Muscle injuries and repair: Current trends in research. J. Bone Joint Surg. Am. Vol. 2002, 84, 822–832. [Google Scholar] [CrossRef]
- Grounds, M.D.; Yablonka-Reuveni, Z. Molecular and cell biology of skeletal muscle regeneration. In Molecular and Cell Biology of Muscular Dystrophy; Partridge, T., Ed.; Springer: Dordrecht, The Netherlands, 1993. [Google Scholar]
- Aarimaa, V.; Rantanen, J.; Best, T.; Schultz, E.; Corr, D.; Kalimo, H. Mild eccentric stretch injury in skeletal muscle causes transient effects on tensile load and cell proliferation. Scand. J. Med. Sci Sports 2004, 14, 367–372. [Google Scholar] [CrossRef] [PubMed]
- Jarvinen, T.A.; Jarvinen, T.L.; Kaariainen, M.; Kalimo, H.; Jarvinen, M. Muscle injuries: Biology and treatment. Am. J. Sports Med. 2005, 33, 745–764. [Google Scholar] [CrossRef] [PubMed]
- Jankowski, R.J.; Deasy, B.M.; Cao, B.; Gates, C.; Huard, J. The role of CD34 expression and cellular fusion in the regeneration capacity of myogenic progenitor cells. J. Cell Sci. 2002, 115, 4361–4374. [Google Scholar] [CrossRef] [PubMed]
- Ekstrand, J.; Askling, C.; Magnusson, H.; Mithoefer, K. Return to play after thigh muscle injury in elite football players: Implementation and validation of the Munich muscle injury classification. Br. J. Sports Med. 2013, 47, 769–774. [Google Scholar] [CrossRef] [PubMed]
- Järvinen, M. Healing of a crush injury in rat striated muscle: 2. A histological study of the effect of early mobilization and immobilization on the repair processes. Acta Pathol. Microbiol. Scand. Sect. A Pathol. 1975, 83, 269–282. [Google Scholar] [CrossRef]
- Valkering, K.P.; Aufwerber, S.; Ranuccio, F.; Lunini, E.; Edman, G.; Ackermann, P.W. Functional weight-bearing mobilization after Achilles tendon rupture enhances early healing response: A single-blinded randomized controlled trial. Sports Traumatol. 2017, 25, 1807–1816. [Google Scholar] [CrossRef] [PubMed]
- McCormack, R.; Bovard, J. Early functional rehabilitation or cast immobilisation for the postoperative management of acute Achilles tendon rupture? A systematic review and meta-analysis of randomised controlled trials. Br. J. Sports Med. 2015, 49, 1329–1335. [Google Scholar] [CrossRef] [PubMed]
- Witard, O.; Wardle, S.; Macnaughton, L.; Hodgson, A.; Tipton, K. Protein Considerations for Optimising Skeletal Muscle Mass in Healthy Young and Older Adults. Nutrients 2016, 8, 181. [Google Scholar] [CrossRef] [PubMed]
- McGlory, C.; Devries, M.; Phillips, S. Skeletal muscle and resistance exercise training; the role of protein synthesis in recovery and remodeling. J. Appl. Physiol. 2016, 122, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Morelli, K.M.; Brown, L.B.; Warren, G.L. Effect of NSAIDs on Recovery from Acute Skeletal Muscle Injury: A Systematic Review and Meta-analysis. Am. J. Sports Med. 2018, 46, 224–233. [Google Scholar] [CrossRef] [PubMed]
- Järvinen, M.; Lehto, M.; Sorvari, T.; Mikola, A. Effect of some anti-inflammatory agents on the healing of ruptured muscle: An experimental study in rats. J. Sport Traumatol. Relat. Res. 1992, 14, 19–28. [Google Scholar]
- Rahusen, F.T.; Weinhold, P.S.; Almekinders, L.C. Nonsteroidal anti-inflammatory drugs and acetaminophen in the treatment of an acute muscle injury. Am. J. Sports Med. 2004, 32, 1856–1859. [Google Scholar] [CrossRef] [PubMed]
- Beiner, J.M.; Jokl, P.; Cholewicki, J.; Panjabi, M.M. The effect of anabolic steroids and corticosteroids on healing of muscle contusion injury. Am. J. Sports Med. 1999, 27, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Carson, J.A.; Manolagas, S.C. Effects of sex steroids on bones and muscles: Similarities, parallels, and putative interactions in health and disease. Bone 2015, 80, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Levine, W.N.; Bergfeld, J.A.; Tessendorf, W.; Moorman, C.T., 3rd. Intramuscular corticosteroid injection for hamstring injuries. A 13-year experience in the National Football League. Am. J. Sports Med. 2000, 28, 297–300. [Google Scholar] [CrossRef] [PubMed]
- Quattrocelli, M.; Barefield, D.Y.; Warner, J.L.; Vo, A.H.; Hadhazy, M.; Earley, J.U.; Demonbreun, A.R.; McNally, E.M. Intermittent glucocorticoid steroid dosing enhances muscle repair without eliciting muscle atrophy. J. Clin. Investig. 2017, 127, 2418–2432. [Google Scholar] [CrossRef] [PubMed]
- Menetrey, J.; Kasemkijwattana, C.; Day, C.S.; Bosch, P.; Vogt, M.; Fu, F.H.; Moreland, M.S.; Huard, J. Growth factors improve muscle healing in vivo. J. Bone Joint Surg. Br. Vol. 2000, 82, 131–137. [Google Scholar] [CrossRef]
- Kasemkijwattana, C.; Menetrey, J.; Bosch, P.; Somogyi, G.; Moreland, M.S.; Fu, F.H.; Buranapanitkit, B.; Watkins, S.S.; Huard, J. Use of growth factors to improve muscle healing after strain injury. Clin. Orthop. Relat. Res. 2000, 370, 272–285. [Google Scholar] [CrossRef]
- Diao, Y.-P.; Cui, F.-K.; Yan, S.; Chen, Z.-G.; Lian, L.-S.; Guo, L.-L.; Li, Y.-J. Nerve Growth Factor Promotes Angiogenesis and Skeletal Muscle Fiber Remodeling in a Murine Model of Hindlimb Ischemia. Chin. Med. J. 2016, 129, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.J.; Thaloor, D.; Matteson, S.; Pavlath, G.K. Hepatocyte growth factor affects satellite cell activation and differentiation in regenerating skeletal muscle. Am. J. Physiol. Cell. Physiol. 2000, 278, 174–181. [Google Scholar] [CrossRef] [PubMed]
- White, J.; Smythe, G. Growth Factors and Cytokines in Skeletal Muscle Development, Growth, Regeneration and Disease; Springer: Berlin, Germany, 2016; Volume 900. [Google Scholar]
- Rantanen, J.; Thorsson, O.; Wollmer, P.; Hurme, T.; Kalimo, H. Effects of therapeutic ultrasound on the regeneration of skeletal myofibers after experimental muscle injury. Am. J. Sports Med. 1999, 27, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Markert, C.D.; Merrick, M.A.; Kirby, T.E.; Devor, S.T. Nonthermal ultrasound and exercise in skeletal muscle regeneration. Arch. Phys. Med. Rehabil. 2005, 86, 1304–1310. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Diaz, D.C.; Gordon, B.S.; Dompier, T.; Burgess, S.; Dumke, C.; Mazoué, C.; Caldwell, T.; Kostek, M.C. Therapeutic Ultrasound Affects IGF-1 Splice Variant Expression in Human Skeletal Muscle. Am. J. Sports Med. 2011, 39, 2233–2241. [Google Scholar] [CrossRef] [PubMed]
- Chongsatientam, A.; Yimlamai, T. Therapeutic Pulsed Ultrasound Promotes Revascularization and Functional Recovery of Rat Skeletal Muscle after Contusion Injury. Ultrasound Med. Biol. 2016, 42, 2938–2949. [Google Scholar] [CrossRef] [PubMed]
- Koike, T.; Camargo, R.C.; Ozaki, G.; Castoldi, R.; Seraphim, P.; Oikawa, S.; Camargo Filho, J.C. Morphometric and fractal analysis of injured skeletal muscle tissue subjected to a combination of treatments; cryotherapy and therapeutic ultrasound. Int. J. Morphol. 2016, 34, 1076–1082. [Google Scholar] [CrossRef]
- Best, T.M.; Loitz-Ramage, B.; Corr, D.T.; Vanderby, R. Hyperbaric oxygen in the treatment of acute muscle stretch injuries. Results in an animal model. Am. J. Sports Med. 1998, 26, 367–372. [Google Scholar] [CrossRef] [PubMed]
- Bennett, M.; Best, T.M.; Babul, S.; Taunton, J.; Lepawsky, M. Hyperbaric oxygen therapy for delayed onset muscle soreness and closed soft tissue injury. Cochrane Database Syst Rev. 2005, 4, CD004713. [Google Scholar] [CrossRef] [PubMed]
- Hyngstrom, A.S.; Murphy, S.A.; Nguyen, J.; Schmit, B.D.; Negro, F.; Gutterman, D.D.; Durand, M.J. Ischemic conditioning increases strength and volitional activation of paretic muscle in chronic stroke: A pilot study. J. Appl. Physiol. 2018, 124, 1140–1147. [Google Scholar] [CrossRef] [PubMed]
- Taylor, N.A.; Wilkinson, J.G. Exercise-induced skeletal muscle growth. Hypertrophy or hyperplasia? Sports Med. 1986, 3, 190–200. [Google Scholar] [CrossRef] [PubMed]
- Bian, W.; Bursac, N. Tissue engineering of functional skeletal muscle: Challenges and recent advances. IEEE Eng. Med. Biol. Mag. 2008, 27, 109–113. [Google Scholar] [CrossRef] [PubMed]
- Otis, J.S.; Burkholder, T.J.; Pavlath, G.K. Stretch-induced myoblast proliferation is dependent on the COX2 pathway. Exp. Cell Res. 2005, 310, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Andersen, J.I.; Juhl, M.; Nielsen, T.; Emmersen, J.; Fink, T.; Zachar, V.; Pennisi, C.P. Uniaxial cyclic strain enhances adipose-derived stem cell fusion with skeletal myocytes. Biochem. Biophys. Res. Commun. 2014, 450, 1083–1088. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.J.; Chen, Y.J.; Huang, C.W.; Fan, S.C.; Huang, B.M.; Chang, W.T.; Tsai, Y.S.; Su, F.C.; Wu, C.C. Cyclic Stretch Facilitates Myogenesis in C2C12 Myoblasts and Rescues Thiazolidinedione-Inhibited Myotube Formation. Front. Bioeng. Biotechnol. 2016, 4, 27. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Kuang, W.; Jin, Z.; Jin, F.; Xu, L.; Yu, Q.; Kong, L.; Zeng, G.; Yuan, X.; Duan, Y. Inhibition of NFkappaB by activated c-Jun NH2 terminal kinase 1 acts as a switch for C2C12 cell death under excessive stretch. Apoptosis 2009, 14, 764–770. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Wu, X.; Young, A.T.; Haynes, C.L. Microfluidics-based in vivo mimetic systems for the study of cellular biology. Acc. Chem. Res. 2014, 47, 1165–1173. [Google Scholar] [CrossRef] [PubMed]
- Shin, Y.; Han, S.; Jeon, J.S.; Yamamoto, K.; Zervantonakis, I.K.; Sudo, R.; Kamm, R.D.; Chung, S. Microfluidic assay for simultaneous culture of multiple cell types on surfaces or within hydrogels. Nat. Protoc. 2012, 7, 1247. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kim, W.; Lim, S.; Jeon, J. Vasculature-On-A-Chip for In Vitro Disease Models. Bioengineering 2017, 4, 8. [Google Scholar] [CrossRef] [PubMed]
- Bersini, S.; Jeon, J.S.; Moretti, M.; Kamm, R.D. In vitro models of the metastatic cascade: From local invasion to extravasation. Drug Discov. Today 2014, 19, 735–742. [Google Scholar] [CrossRef] [PubMed]
- Madden, L.; Juhas, M.; Kraus, W.E.; Truskey, G.A.; Bursac, N. Bioengineered human myobundles mimic clinical responses of skeletal muscle to drugs. Elife 2015, 4, e04885. [Google Scholar] [CrossRef] [PubMed]
- Uzel, S.G.M.; Platt, R.J.; Subramanian, V.; Pearl, T.M.; Rowlands, C.J.; Chan, V.; Boyer, L.A.; So, P.T.C.; Kamm, R.D. Microfluidic device for the formation of optically excitable, three-dimensional, compartmentalized motor units. Sci. Adv. 2016, 2. [Google Scholar] [CrossRef] [PubMed]
- Osaki, T.; Uzel, S.G.M.; Kamm, R.D. Microphysiological 3D model of amyotrophic lateral sclerosis (ALS) from human iPS-derived muscle cells and optogenetic motor neurons. Sci. Adv. 2018, 4. [Google Scholar] [CrossRef] [PubMed]
- Hyun, K.U.; Sukhee, P.; Hyunseung, B.; Mina, K.; Hyunjun, S.; Shin, J.H. Promotion of Myogenic Maturation by Timely Application of Electric Field Along the Topographical Alignment. Tissue Eng. Part A 2018, 24, 752–760. [Google Scholar] [CrossRef]
- Agrawal, G.; Aung, A.; Varghese, S. Skeletal muscle-on-a-chip: An in vitro model to evaluate tissue formation and injury. Lab Chip 2017, 17, 3447–3461. [Google Scholar] [CrossRef] [PubMed]
- Huh, D.; Matthews, B.D.; Mammoto, A.; Montoya-Zavala, M.; Hsin, H.Y.; Ingber, D.E. Reconstituting Organ-Level Lung Functions on a Chip. Science 2010, 328, 1662–1668. [Google Scholar] [CrossRef] [PubMed]
- Bächtold, P.R. Stretchable, High–Throughput, Continuously Perfused Cell Culture System; Swiss Federal Institute of Technology Zurich: Zürich, Switzerland, 2009. [Google Scholar]
- Lee, J.N.; Jiang, X.; Ryan, D.; Whitesides, G.M. Compatibility of Mammalian Cells on Surfaces of Poly(dimethylsiloxane). Langmuir 2004, 20, 11684–11691. [Google Scholar] [CrossRef] [PubMed]
- Wan, C.R.; Chung, S.; Kamm, R.D. Differentiation of embryonic stem cells into cardiomyocytes in a compliant microfluidic system. Ann. Biomed. Eng. 2011, 39, 1840–1847. [Google Scholar] [CrossRef] [PubMed]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [PubMed]
- Wells, L.; Edwards, K.A.; Bernstein, S.I. Myosin heavy chain isoforms regulate muscle function but not myofibril assembly. The EMBO journal 1996, 15, 4454–4459. [Google Scholar] [CrossRef] [PubMed]
- van der Vliet, A.; Janssen-Heininger, Y.M. Hydrogen peroxide as a damage signal in tissue injury and inflammation: Murderer, mediator, or messenger? J. Cell. Biochem. 2014, 115, 427–435. [Google Scholar] [CrossRef] [PubMed]
- Kozakowska, M.; Pietraszek-Gremplewicz, K.; Jozkowicz, A.; Dulak, J. The role of oxidative stress in skeletal muscle injury and regeneration: Focus on antioxidant enzymes. J. Muscle Res. Cell Motil. 2015, 36, 377–393. [Google Scholar] [CrossRef] [PubMed]
- Brancaccio, P.; Lippi, G.; Maffulli, N. Biochemical markers of muscular damage. Clin. Chem. Lab. Med. 2010, 48, 757–767. [Google Scholar] [CrossRef] [PubMed]
- Beiner, J.; Jokl, P. Muscle contusion injuries: Current treatment options. J. Am. Acad. Orthop. Surg. 2001, 9, 227–237. [Google Scholar] [CrossRef] [PubMed]




© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, W.; Kim, J.; Park, H.-S.; Jeon, J.S. Development of Microfluidic Stretch System for Studying Recovery of Damaged Skeletal Muscle Cells. Micromachines 2018, 9, 671. https://doi.org/10.3390/mi9120671
Kim W, Kim J, Park H-S, Jeon JS. Development of Microfluidic Stretch System for Studying Recovery of Damaged Skeletal Muscle Cells. Micromachines. 2018; 9(12):671. https://doi.org/10.3390/mi9120671
Chicago/Turabian StyleKim, Wanho, Jaesang Kim, Hyung-Soon Park, and Jessie S. Jeon. 2018. "Development of Microfluidic Stretch System for Studying Recovery of Damaged Skeletal Muscle Cells" Micromachines 9, no. 12: 671. https://doi.org/10.3390/mi9120671
APA StyleKim, W., Kim, J., Park, H.-S., & Jeon, J. S. (2018). Development of Microfluidic Stretch System for Studying Recovery of Damaged Skeletal Muscle Cells. Micromachines, 9(12), 671. https://doi.org/10.3390/mi9120671