Flow-Based Three-Dimensional Co-Culture Model for Long-Term Hepatotoxicity Prediction


Abstract
1. Introduction
2. Materials and Methods
2.1. Fabrication of Concave Chip with Flow
2.2. Isolation and Culture of Primary Hepatocyte and HSCs
2.3. Formation of Hepatospheres and Heterospheres in the Concave Chamber
2.4. SEM
2.5. Immunofluorescence Staining
2.6. Cell Viability
2.7. Functional Assessment
2.8. Reverse Transcriptase Polymerase Chain Reaction (RT-PCR)
2.9. Preparation and Treatment of Drugs, Cytotoxicity Test
2.10. Statistical Analysis
3. Results
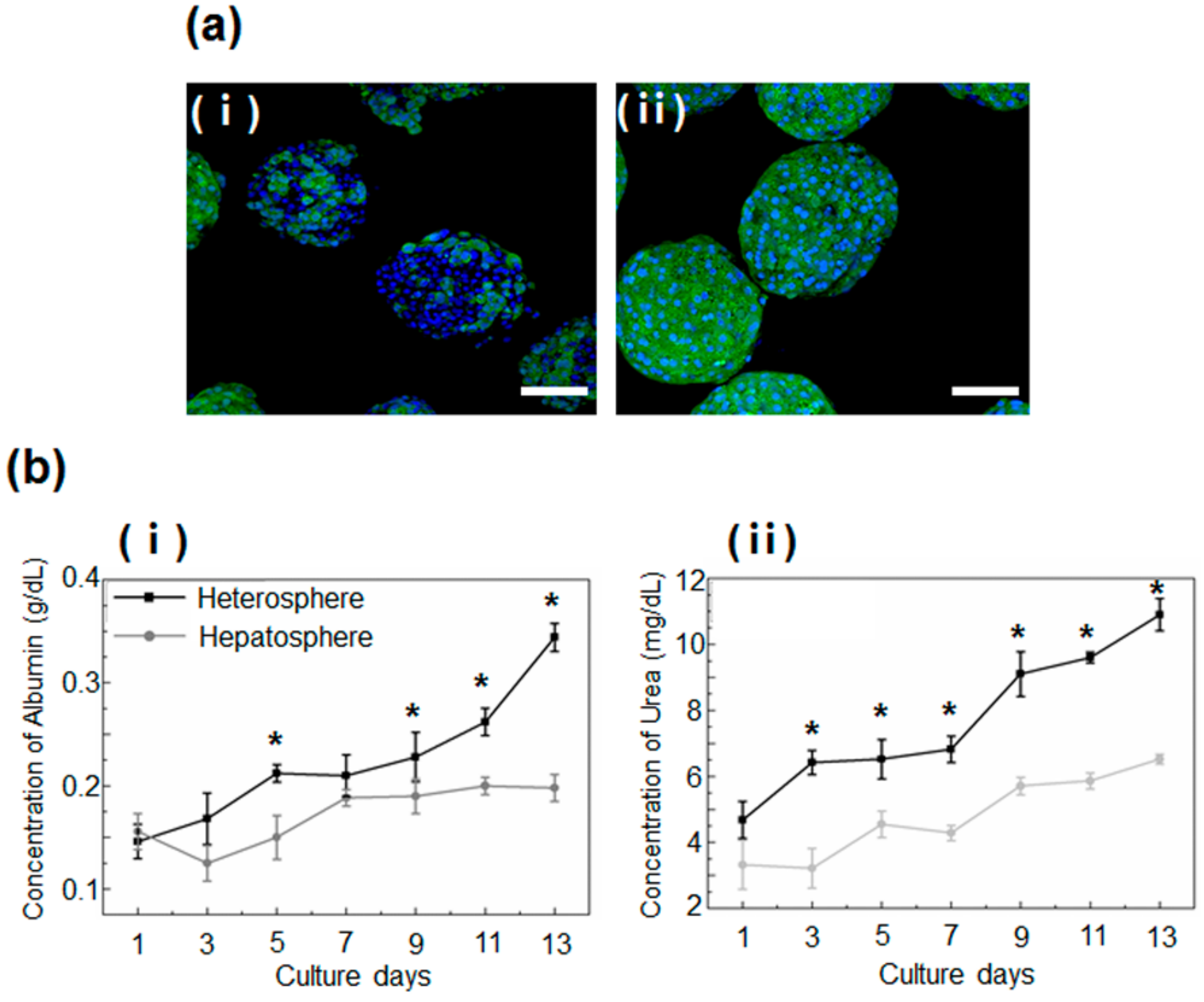
3.1. Morphological Observation and Viability of Spheroids Cultured in Concave Flow Chips
3.2. Functional Evaluation of Spheroids Culture in Concave Flow Chips
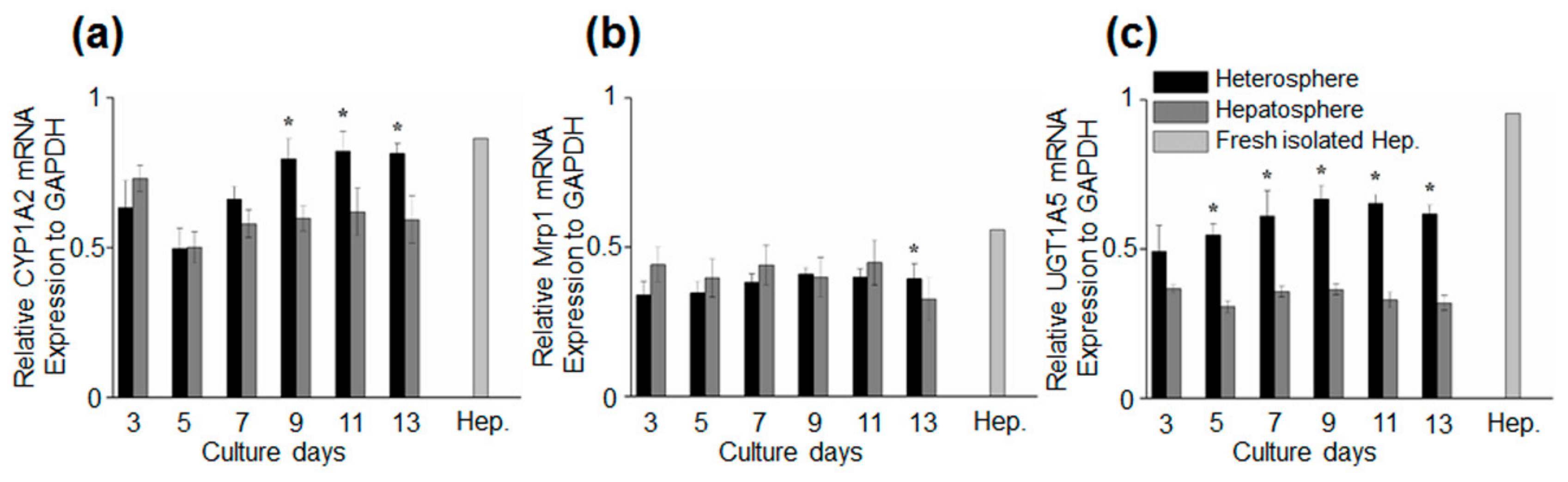
3.3. Hepatocyte-Specific Gene Expression of Spheroids Culture in Concave Flow Chips
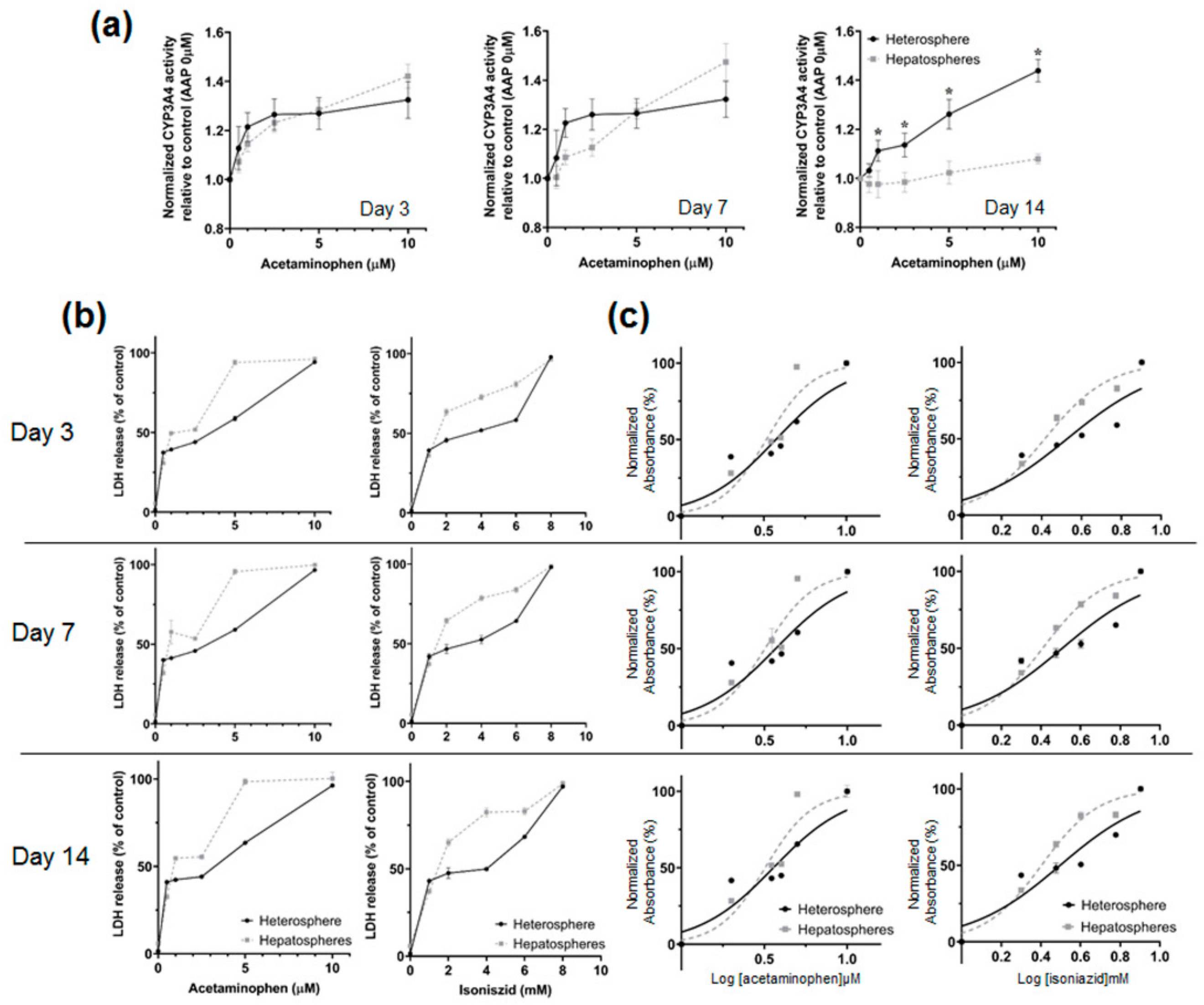
3.4. Drug Induced Hepatotoxicity and Metabolic Function Evaluation of Spheroids Culture in Concave Flow Chips
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Gao, B.; Jeong, W.I.; Tian, Z. Liver: An organ with predominant innate immunity. Hepatology 2008, 47, 729–736. [Google Scholar] [CrossRef] [PubMed]
- Friedman, S.L. Molecular regulation of hepatic fibrosis, an integrated cellular response to tissue injury. J. Biol. Chem. 2000, 275, 2247–2250. [Google Scholar] [CrossRef] [PubMed]
- Hewitt, N.J.; Lechón, M.J.; Houston, J.B.; Hallifax, D.; Brown, H.S.; Maurel, P.; Kenna, J.G.; Gustavsson, L.; Lohmann, C.; Skonberg, C.; et al. Primary hepatocytes: Current understanding of the regulation of metabolic enzymes and transporter proteins, and pharmaceutical practice for the use of hepatocytes in metabolism, enzyme induction, transporter, clearance, and hepatotoxicity studies. Drug Metab. Rev. 2007, 39, 159–234. [Google Scholar] [CrossRef] [PubMed]
- Jones, C.N.; Tuleuova, N.; Lee, J.Y.; Ramanculov, E.; Reddi, A.H.; Zern, M.A.; Revzin, A. Cultivating hepatocytes on printed arrays of HGF and BMP7 to characterize protective effects of these growth factors during in vitro alcohol injury. Biomaterials 2010, 31, 5936–5944. [Google Scholar] [CrossRef]
- Sivaraman, A.; Leach, J.K.; Townsend, S.; Iida, T.; Hogan, B.J.; Stolz, D.B.; Fry, R.; Samson, L.D.; Tannenbaum, S.R.; Griffith, L.G. A microscale in vitro physiological model of the liver: Predictive screens for drug metabolism and enzyme induction. Curr. Drug. Metab. 2005, 6, 569–591. [Google Scholar] [CrossRef]
- Blouin, A.; Bolender, R.P.; Weibel, E.R. Distribution of organelles and membranes between hepatocytes and nonhepatocytes in the rat liver parenchyma. A stereological study. J. Cell Biol. 1977, 72, 441–455. [Google Scholar] [CrossRef]
- Gressner, A.M.; Krull, N.; Bachem, M.G. Regulation of proteoglycan expression in fibrotic liver and cultured fat-storing cells. Pathol. Res. Pract. 1994, 190, 864–882. [Google Scholar] [CrossRef]
- De Minicis, S.; Seki, E.; Uchinami, H.; Kluwe, J.; Zhang, Y.; Brenner, D.A.; Schwabe, R.F. Gene expression profiles during hepatic stellate cell activation in culture and in vivo. Gastroenterology 2007, 132, 1937–1946. [Google Scholar] [CrossRef]
- Gressner, A.M.; Weiskirchen, R. Modern pathogenetic concepts of liver fibrosis suggest stellate cells and TGF-beta as major players and therapeutic targets. J. Cell Mol. Med. 2006, 10, 76–99. [Google Scholar] [CrossRef]
- Uyama, N.; Shimahara, Y.; Kawada, N.; Seki, S.; Okuyama, H.; Iimuro, Y.; Yamaoka, Y. Regulation of cultured rat hepatocyte proliferation by stellate cells. J. Hepatol. 2002, 36, 590–599. [Google Scholar] [CrossRef]
- Ostrovidov, S.; Jiang, J.; Sakai, Y.; Fujii, T. Membrane-based PDMS microbioreactor for perfused 3D primary rat hepatocyte cultures. Biomed. Microdevices 2004, 6, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Miyazawa, M.; Torii, T.; Toshimitsu, Y.; Koyama, I. Effect of mechanical stress imposition on co-culture of hepatic parenchymal and nonparenchymal cells: Possibility of stimulating production of regenerating factor. Transplant. Proc. 2005, 37, 2398–2401. [Google Scholar] [CrossRef] [PubMed]
- Sudo, R.; Mitaka, T.; Ikeda, M.; Tanishita, K. Reconstruction of 3D stacked-up structures by rat small hepatocytes on microporous membranes. FASEB J. 2005, 19, 1695–1697. [Google Scholar] [CrossRef] [PubMed]
- Glicklis, R.; Merchuk, J.C.; Cohen, S. Modeling mass transfer in hepatocyte spheroids via cell viability, spheroid size, and hepatocellular functions. Biotechnol. Bioeng. 2004, 86, 672–680. [Google Scholar] [CrossRef] [PubMed]
- Sakai, Y.; Yamagami, S.; Nakazawa, K. Comparative analysis of gene expression in rat liver tissue and monolayer- and spheroid-cultured hepatocytes. Cells Tissues Organs 2010, 191, 281–288. [Google Scholar] [CrossRef]
- Thomas, R.J.; Bhandari, R.; Barrett, D.A.; Bennett, A.J.; Fry, J.R.; Powe, D.; Thomson, B.J.; Shakesheff, K.M. The effect of three-dimensional co-culture of hepatocytes and hepatic stellate cells on key hepatocyte functions in vitro. Cells Tissues Organs 2005, 181, 67–79. [Google Scholar] [CrossRef]
- Choi, Y.Y.; Kim, J.; Lee, S.H.; Kim, D.S. Lab on a chip-based hepatic sinusoidal system simulator for optimal primary hepatocyte culture. Biomed. Microdevices 2016, 18, 58. [Google Scholar] [CrossRef]
- Lee, S.A.; No da, Y.; Kang, E.; Ju, J.; Kim, D.S.; Lee, S.H. Spheroid-based three-dimensional liver-on-a-chip to investigate hepatocyte-hepatic stellate cell interactions and flow effects. Lab. chip 2013, 18, 3529–3537. [Google Scholar] [CrossRef]
- Park, J.Y.; Hwang, C.M.; Lee, S.H. Gradient generation by an osmotic pump and the behavior of human mesenchymal stem cells under the fetal bovine serum concentration gradient. Lab. Chip 2007, 7, 1673–1680. [Google Scholar] [CrossRef]
- Riccalton-Banks, L.; Bhandari, R.; Fry, J.; Shakesheff, K.M. A simple method for the simultaneous isolation of stellate cells and hepatocytes from rat liver tissue. Mol. Cell Biochem. 2003, 248, 97–102. [Google Scholar] [CrossRef]
- Ahmed, H.M.M.; Salerno, S.; Morelli, S.; Giorno, L.; De Bartolo, L. 3D liver membrane system by co-culturing human hepatocytes, sinusoidal endothelial and stellate cells. Biofabrication 2017, 9, 025022. [Google Scholar] [CrossRef] [PubMed]
- Matis, G.; Kulcsár, A.; Petrilla, J.; Talapka, P.; Neogrády, Z. Porcine hepatocyte-Kupffer cell co-culture as an in vitro model for testing the efficacy of anti-inflammatory substances. J. Anim. Physiol. Anim. Nutr. 2017, 101, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Xiao, W.; Perry, G.; Komori, K.; Sakai, Y. New physiologically-relevant liver tissue model based on hierarchically cocultured primary rat hepatocytes with liver endothelial cells. Integr. Biol. 2015, 7, 1412–1422. [Google Scholar] [CrossRef] [PubMed]
- Zinchenko, Y.S.; Culberson, C.R.; Coger, R.N. Contribution of non-parenchymal cells to the performance of micropatterned hepatocytes. Tissue Eng. 2006, 12, 2241–2251. [Google Scholar] [CrossRef] [PubMed]
- Zinchenko, Y.S.; Schrum, L.W.; Clemens, M.; Coger, R.N. Hepatocyte and kupffer cells co-cultured on micropatterned surfaces to optimize hepatocyte function. Tissue Eng. 2006, 12, 751–761. [Google Scholar] [CrossRef]
- Lu, H.F.; Chua, K.N.; Zhang, P.C.; Lim, W.S.; Ramakrishna, S.; Leong, K.W.; Mao, H.Q. Three-dimensional co-culture of rat hepatocyte spheroids and NIH/3T3 fibroblasts enhances hepatocyte functional maintenance. Acta Biomater. 2005, 1, 399–410. [Google Scholar] [CrossRef]
- Racanelli, V.; Rehermann, B. The liver as an immunological organ. Hepatology 2006, 43, S54–S62. [Google Scholar] [CrossRef]
- Gandhi, C.R. Oxidative Stress and Hepatic stellate cell: A PATADOXICAL RELATIONSHIP. Trends Cell Mol. Biol. 2012, 7, 1–10. [Google Scholar]
- Wong, S.F.; No, D.Y.; Choi, Y.Y.; Kim, D.S.; Chung, B.G.; Lee, S.H. Concave microwell based size-controllable hepatosphere as a three-dimensional liver tissue model. Biomaterials 2011, 32, 8087–8096. [Google Scholar] [CrossRef]
- Raucy, J.L.; Lasker, J.M.; Lieber, C.S.; Black, M. Acetaminophen activation by human liver cytochromes P450IIE1 and P450IA2. Arch. Biochem. Biophys. 1989, 271, 270–283. [Google Scholar] [CrossRef]
- Schyschka, L.; Sánchez, J.J.; Wang, Z.; Burkhardt, B.; Müller-Vieira, U.; Zeilinger, K.; Bachmann, A.; Nadalin, S.; Damm, G.; Nussler, A.K. Hepatic 3D cultures but not 2D cultures preserve specific transporter activity for acetaminophen-induced hepatotoxicity. Arch. Toxicol. 2013, 87, 1581–1593. [Google Scholar] [CrossRef] [PubMed]
- DelRaso, N.J. In vitro methodologies for enhanced toxicity testing. Toxicol. Lett. 1993, 68, 91–99. [Google Scholar] [CrossRef]
- Soldatow, V.Y.; LeCluyse, L.; Griffith, L.G.; Rusyn, I. In vitro models for liver toxicity testing. Toxicol. Res. 2013, 2, 23–39. [Google Scholar] [CrossRef] [PubMed]
- Guguen-Guillouzo, C.; Corlu, A.; Guillouzo, A. Stem cell-derived hepatocytes and their use in toxicology. Toxicology 2010, 270, 3–9. [Google Scholar]
- LeCluyse, E.L. Human hepatocyte culture systems for the in vitro evaluation of cytochrome P450 expression and regulation. Eur. J. Pharm. Sci. 2001, 13, 343–368. [Google Scholar] [CrossRef]
- O’Brien, P.J.; Siraki, A.G. Accelerated cytotoxicity mechanism screening using drug metabolising enzyme modulators. Curr. Drug. Metab. 2005, 6, 101–109. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IC50 Values | Day 3 | Day 7 | Day 14 | |||
|---|---|---|---|---|---|---|
| Heterosphere | Hepatosphere | Heterosphere | Hepatosphere | Heterosphere | Hepatosphere | |
| IC50 of acetaminophen (µM) | 2.23 | 2.13 | 2.11 | 1.57 | 2.15 | 1.02 |
| IC50 of isoniazid (mM) | 2.96 | 2.35 | 2.81 | 1.80 | 2.66 | 1.25 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, Y.Y.; Seok, J.-I.; Kim, D.-S. Flow-Based Three-Dimensional Co-Culture Model for Long-Term Hepatotoxicity Prediction. Micromachines 2020, 11, 36. https://doi.org/10.3390/mi11010036
Choi YY, Seok J-I, Kim D-S. Flow-Based Three-Dimensional Co-Culture Model for Long-Term Hepatotoxicity Prediction. Micromachines. 2020; 11(1):36. https://doi.org/10.3390/mi11010036
Chicago/Turabian StyleChoi, Yoon Young, Jin-I Seok, and Dong-Sik Kim. 2020. "Flow-Based Three-Dimensional Co-Culture Model for Long-Term Hepatotoxicity Prediction" Micromachines 11, no. 1: 36. https://doi.org/10.3390/mi11010036
APA StyleChoi, Y. Y., Seok, J.-I., & Kim, D.-S. (2020). Flow-Based Three-Dimensional Co-Culture Model for Long-Term Hepatotoxicity Prediction. Micromachines, 11(1), 36. https://doi.org/10.3390/mi11010036