
The Mechanism of Diarrhetic Shellfish Poisoning Toxin Production in Prorocentrum spp.: Physiological and Molecular Perspectives


Abstract
:1. Introduction
2. Nutritional Factors
2.1. Nitrogen
2.2. Phosphorus
2.3. Chelating Reagents
3. Physical Factors
3.1. Temperature
3.2. Salinity
3.3. Light
4. Bacteria Interaction
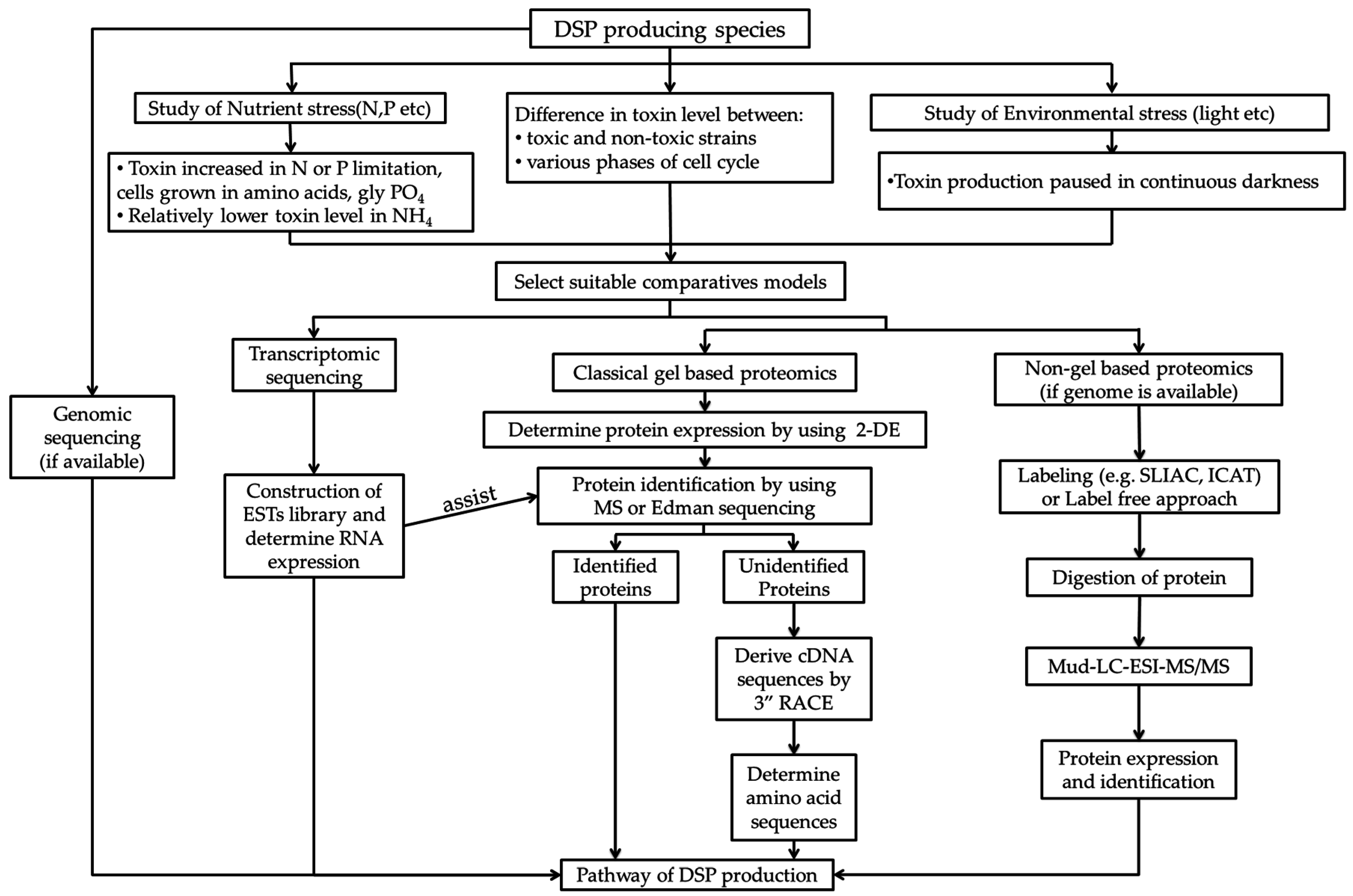
5. Application of Molecular Tools for Studying the Mechanism of Algal Toxin Production
5.1. Genomics and Transcriptomics Studies
5.2. Proteomics
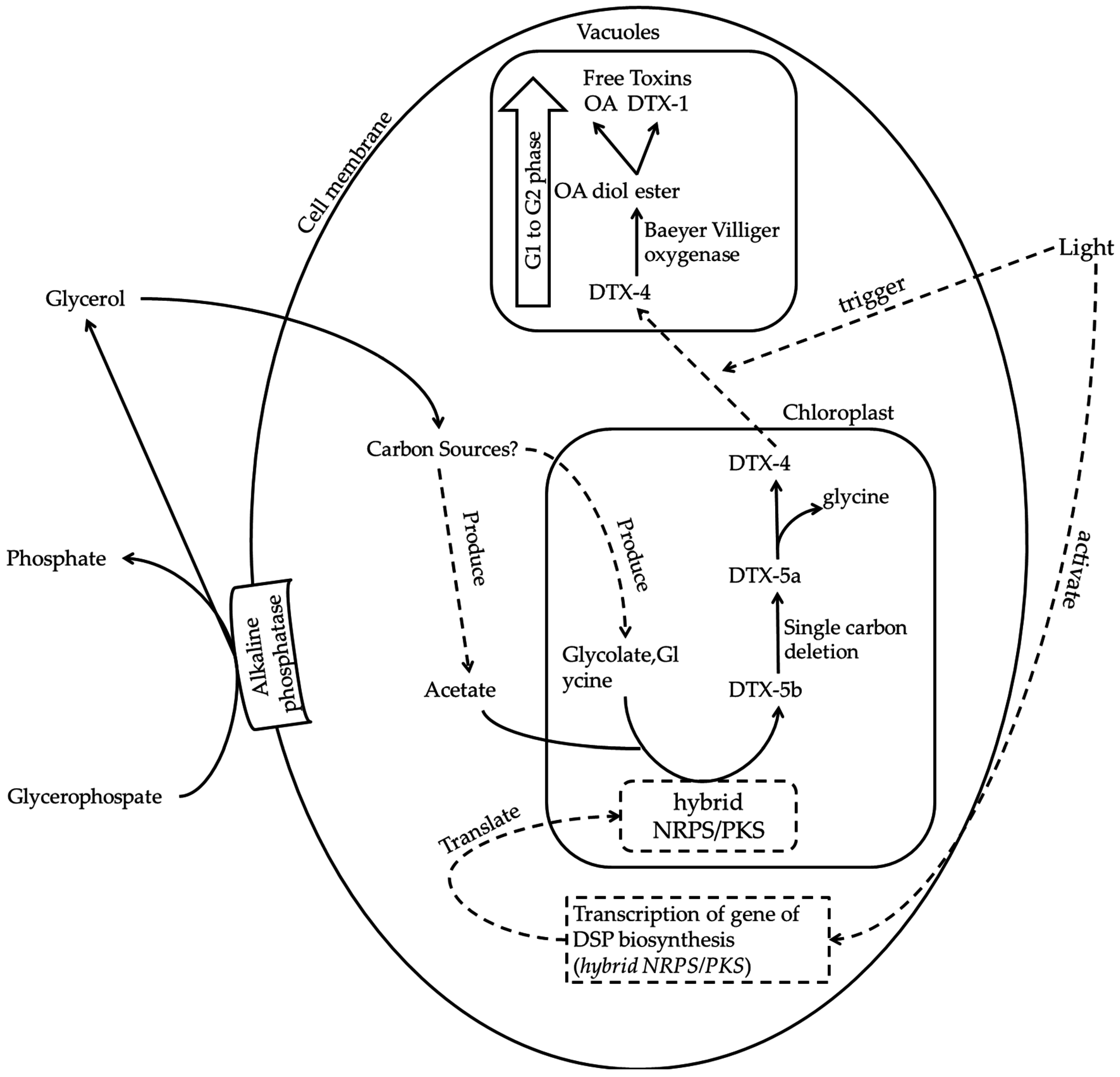
6. Putative Mechanism of DSP Toxin Production
7. Conclusions and Perspectives
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kat, M. Diarrhetic mussel poisoning in the Netherlands related to the dinoflagellate Dinophysis acuminata. Antonie van Leeuwenhoek 1983, 49, 417–427. [Google Scholar] [PubMed]
- Aune, T.; Yndestad, M. Diarrhetic shellfish poisoning. In Algal Toxins in Seafood and Drinking Water; Ian, R.F., Ed.; Academic Press: San Diego, CA, USA, 1993; pp. 87–104. [Google Scholar]
- Hallegraeff, G.M. A review of harmful algal blooms and their apparent global increase. Phycologia 1993, 32, 79–99. [Google Scholar] [CrossRef]
- Pitcher, G.C.; Krock, B.; Cembella, A.D. Accumulation of diarrhetic shellfish poisoning toxins in the oyster Crassostrea gigas and the mussel Choromytilus meridionalis in the Southern Benguela ecosystem. Afr. J. Mar. Sci. 2011, 33, 273–281. [Google Scholar] [CrossRef]
- Li, A.; Ma, J.; Cao, J.; McCarron, P. Toxins in mussels (Mytilus galloprovincialis) associated with diarrhetic shellfish poisoning episodes in China. Toxicon 2012, 60, 420–425. [Google Scholar] [CrossRef] [PubMed]
- Madigan, T.L.; Lee, K.G.; Padula, D.J.; McNabb, P.; Pointon, A.M. Diarrhetic shellfish poisoning (DSP) toxins in South Australian shellfish. Harmful Algae 2006, 5, 119–123. [Google Scholar] [CrossRef]
- Sim, J.; Wilson, N. Surveillance of marine biotoxins, 1993–96. N. Z. Public Health Rep. 1997, 4, 9–16. [Google Scholar]
- Fernández, J.J.; Suárez-Gómez, B.; Souto, M.L.; Norte, M. Identification of new okadaic acid derivatives from laboratory cultures of Prorocentrum lima. J. Nat. Prod. 2003, 66, 1294–1296. [Google Scholar] [CrossRef] [PubMed]
- Yasumoto, T.; Oshima, Y.; Sugawara, W.; Fukuyo, Y.; Oguri, H.; Igarashi, T.; Fujita, N. Identification of Dinophysis fortii as the causative organism of diarrhetic shellfish poisoning. Nippon Suisan Gakk 1980, 46, 1405–1411. [Google Scholar] [CrossRef]
- Zhou, J.; Fritz, L. Okadaic acid antibody localizes to chloroplasts in the DSP-toxin-producing dinoflagellates Prorocentrum lima and Prorocentrum maculosum. Phycologia 1994, 33, 455–461. [Google Scholar] [CrossRef]
- Granéli, E.; Flynn, K. Chemical and physical factors influencing toxin content. In Ecology of Harmful Algae; Granéli, E., Turner, J.T., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; Volume 189, pp. 229–241. [Google Scholar]
- Li, L.X.; Tang, J.Q.; Yang, W.D.; Liu, J.S.; Zhang, J.J.; Li, H.Y. Studies on toxin production of Prorocentrum lima. Asian J. Ecotoxicol. 2009, 4, 745–750. [Google Scholar]
- Yang, W.d.; Zhong, N.; Liu, J.S.; Zhang, J.L.; Yang, H. Effects of different phosphorus sources on the growth and toxin production of Prorocentrum lima. Environ. Sci. 2008, 29, 2760–2765. [Google Scholar]
- Zhong, N.; Yang, W.; Liu, J.; Zhang, J.; He, Y. Effects of different nitrogen sources on the growth and toxin production of Prorocentrum lima. Acta Sci. Circumst. 2008, 28, 1186–1191. [Google Scholar]
- Vanucci, S.; Guerrini, F.; Milandri, A.; Pistocchi, R. Effects of different levels of N- and P-deficiency on cell yield, okadaic acid, DTX-1, protein and carbohydrate dynamics in the benthic dinoflagellate Prorocentrum lima. Harmful Algae 2010, 9, 590–599. [Google Scholar] [CrossRef]
- Varkitzi, I.; Pagou, K.; Granéli, E.; Hatzianestis, I.; Pyrgaki, C.; Pavlidou, A.; Montesanto, B.; Economou-Amilli, A. Unbalanced N:P ratios and nutrient stress controlling growth and toxin production of the harmful dinoflagellate Prorocentrum lima (Ehrenberg) dodge. Harmful Algae 2010, 9, 304–311. [Google Scholar] [CrossRef]
- McLachlan, J.L.; Marr, J.C.; Conlon-Keily, A.; Adamson, A. Effects of nitrogen concentration and cold temperature on DSP-toxin concentrations in the dinoflagellate Prorocentrum lima (Prorocentrales, Dinophyceae). Nat. Toxins 1994, 2, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Carmelo, R.T.; Daniel, G.B. The influence of phosphorus sources on the growth and cellular toxin content of the benthic dinoflagellate Prorocentrum lima. In Toxic Phytoplankton Blooms in the Sea; Smayda, T.J., Shimizu, Y., Eds.; Elsevier Science Publishers: Amsterdam, The Netherlands, 1993; pp. 565–570. [Google Scholar]
- Sohet, K.; Pereira, A.; Braekman, J.C.; Houvenaghel, G. Growth and toxicity of Prorocentrum lima (Ehrenberg) dodge in different culture media: Effect of humic acids and organic phosphorus. In Harmful Marine Algal Blooms: Proceedings of the Sixth International Conference on Toxic Marine Phytoplankton; Lassus, P., Arzul, G., Erard, E., Gentlen, P., Marcailiou, C., Eds.; Nantes: Pays de la Loire, France, 1995; pp. 669–674. [Google Scholar]
- Jackson, A.E.; Marr, J.C.; McLachlan, J.L. The production of diarrhetic shellfish toxins by an isolate of Prorocentrum lima from Nova Soctia, Canada. In Toxic Phytoplankton Blooms in the Sea: Proceedings of the Fifth International Conference on Toxic Marine Phytoplankton; Smayda, T.J., Shimizu, Y., Eds.; Elsevier: Newport, RI, USA, 1993; pp. 513–518. [Google Scholar]
- Morton, S.L.; Bomber, J.W.; Tindall, P.M. Environmental effects on the production of okadaic acid from Prorocentrum hoffmannianum faust I. Temperature, light, and salinity. J. Exp. Mar. Biol. Ecol. 1994, 178, 67–77. [Google Scholar] [CrossRef]
- Wang, S.; Chen, J.; Li, Z.; Wang, Y.; Fu, B.; Han, X.; Zheng, L. Cultivation of the benthic microalga Prorocentrum lima for the production of diarrhetic shellfish poisoning toxins in a vertical flat photobioreactor. Bioresour. Technol. 2015, 179, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Hou, D.Y.; Liang, J.J.; Zou, C.; Li, H.Y.; Liu, J.S.; Yang, W.D. MRP functional activity and character in the dinoflagellate Prorocentrum lima. J. Appl. Phycol. 2016, 28, 1667–1676. [Google Scholar] [CrossRef]
- López-Rosales, L.; Gallardo-Rodríguez, J.; Sánchez-Mirón, A.; Cerón-García, M.; Belarbi, E.; García-Camacho, F.; Molina-Grima, E. Simultaneous effect of temperature and irradiance on growth and okadaic acid production from the marine dinoflagellate Prorocentrum belizeanum. Toxins 2014, 6, 229. [Google Scholar] [CrossRef] [PubMed]
- Blanco, J.; Moroño, Á.; Fernández, M.L. Toxic episodes in shellfish, produced by lipophilic phycotoxins: An overview. Revista Galega de Recursos Mariños (Monog.) 2005, 1, 1–70. [Google Scholar]
- Cohen, P.; Holmes, C.F.B.; Tsukitani, Y. Okadaic acid: A new probe for the study of cellular regulation. Trends Biochem. Sci. 1990, 15, 98–102. [Google Scholar] [CrossRef]
- EFSA Panel on Contaminants in the Food Chain (CONTAM). Opinion of the scientific panel on contaminants in the food chain on a request from the european commission on marine biotoxins in shellfish – okadaic acid and analogues. EFSA J. 2008, 589, 1–62. [Google Scholar]
- Tachibana, K.; Scheuer, P.J.; Tsukitani, Y.; Kikuchi, H.; Van Engen, D.; Clardy, J.; Gopichand, Y.; Schmitz, F.J. Okadaic acid, a cytotoxic polyether from two marine sponges of the genus Halichondria. J. Am. Chem. Soc. 1981, 103, 2469–2471. [Google Scholar] [CrossRef]
- Schmitz, F.J.; Prasad, R.S.; Gopichand, Y.; Hossain, M.B.; Van der Helm, D.; Schmidt, P. Acanthifolicin, a new episulfide-containing polyether carboxylic acid from extracts of the marine sponge Pandaros acanthifolium. J. Am. Chem. Soc. 1981, 103, 2467–2469. [Google Scholar] [CrossRef]
- Dominguez, H.J.; Paz, B.; Daranas, A.H.; Norte, M.; Franco, J.M.; Fernández, J.J. Dinoflagellate polyether within the yessotoxin, pectenotoxin and okadaic acid toxin groups: Characterization, analysis and human health implications. Toxicon 2010, 56, 191–217. [Google Scholar] [CrossRef] [PubMed]
- Yasumoto, T.; Murata, M.; Oshima, Y.; Sano, M.; Matsumoto, G.K.; Clardy, J. Diarrhetic shellfish toxins. Tetrahedron 1985, 41, 1019–1025. [Google Scholar] [CrossRef]
- García, C.; Truan, D.; Lagos, M.; Santelices, J.P.; Dêaz, J.C.; Lagos, N. Metabolic transformation of dinophysistoxin-3 into dinophysistoxin-1 causes human intoxication by consumption of O-acyl-derivatives dinophysistoxins contaminated shellfish. J. Toxicol. Sci. 2005, 30, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Kamiyama, T.; Okumura, Y.; Ishihara, K.; Matsushima, R.; Kaneniwa, M. Liquid-chromatographic hybrid triple–quadrupole linear-ion-trap ms/ms analysis of fatty-acid esters of dinophysistoxin-1 in bivalves and toxic dinoflagellates in japan. Fisheries Sci. 2009, 75, 1039–1048. [Google Scholar] [CrossRef]
- Vanden Heuvel, J.P. Xenobiotic receptor systems: Introduction and overview. In Comprehensive Toxicology: Cellular and Molecular Toxicology; Elsevier Science Limited: Amsterdam, The Netherlands, 2002. [Google Scholar]
- Suganuma, M.; Fujiki, H.; Suguri, H.; Yoshizawa, S.; Hirota, M.; Nakayasu, M.; Ojika, M.; Wakamatsu, K.; Yamada, K.; Sugimura, T. Okadaic acid: An additional non-phorbol-12-tetradecanoate-13-acetate-type tumor promoter. Proc. Natl. Acad. Sci. USA 1988, 85, 1768–1771. [Google Scholar] [CrossRef] [PubMed]
- Fujiki, H.; Suganuma, M.; Suguri, H.; Yoshizawa, S.; Takagi, K.; Uda, N.; Wakamatsu, K.; Yamada, K.; Murata, M.; Yasumoto, T.; et al. Diarrhetic shellfish toxin, dinophysistoxin-1, is a potent tumor promoter on mouse skin. Cancer Sci. 1988, 79, 1089–1093. [Google Scholar] [CrossRef]
- Fujiki, H.; Suganuma, M. Tumor promotion by inhibitors of protein phosphatases 1 and 2a: The okadaic acid class of compounds. Adv. Cancer Res. 1993, 61, 143–194. [Google Scholar] [PubMed]
- Cordier, S.; Monfort, C.; Miossec, L.; Richardson, S.; Belin, C. Ecological analysis of digestive cancer mortality related to contamination by diarrhetic shellfish poisoning toxins along the coasts of france. Environ. Res. 2000, 84, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Bøe, R.; Gjertsen, B.T.; Vintermyr, O.K.; Houge, G.; Lanotte, M.; Døskeland, S.O. The protein phosphatase inhibitor okadaic acid induces morphological changes typical of apoptosis in mammalian cells. Exp. Cell Res. 1991, 195, 237–246. [Google Scholar] [CrossRef]
- Lerga, A.; Richard, C.; Delgado, M.D.; Cañelles, M.; Frade, P.; Cuadrado, M.A.; León, J. Apoptosis and mitotic arrest are two independent effects of the protein phosphatases inhibitor okadaic acid in K562 leukemia cells. Biochem. Bioph. Res. Commun. 1999, 260, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Jayaraj, R.; Gupta, N.; Rao, P.V.L. Multiple signal transduction pathways in okadaic acid induced apoptosis in hela cells. Toxicology 2009, 256, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Traoré, A.; Baudrimont, I.; Ambaliou, S.; Dano, S.D.; Creppy, E.E. DNA breaks and cell cycle arrest induced by okadaic acid in Caco-2 cells, a human colonic epithelial cell line. Arch. Toxicol. 2001, 75, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Chen, L. Okadaic acid induces apoptosis through the PKR, NF-κB and caspase pathway in human osteoblastic osteosarcoma MG63 cells. Toxicol. in Vitro 2011, 25, 1796–1802. [Google Scholar] [CrossRef] [PubMed]
- Nuydens, R.; De Jong, M.; Van Den Kieboom, G.; Heers, C.; Dispersyn, G.; Cornelissen, F.; Nuyens, R.; Borgers, M.; Geerts, H. Okadaic acid-induced apoptosis in neuronal cells: Evidence for an abortive mitotic attempt. J. Neurochem. 1998, 70, 1124–1133. [Google Scholar] [CrossRef] [PubMed]
- Souid-Mensi, G.; Moukha, S.; Mobio, T.A.; Maaroufi, K.; Creppy, E.E. The cytotoxicity and genotoxicity of okadaic acid are cell-line dependent. Toxicon 2008, 51, 1338–1344. [Google Scholar] [CrossRef] [PubMed]
- Berven, G.; Sætre, F.; Halvorsen, K.; Seglen, P.O. Effects of the diarrhetic shellfish toxin, okadaic acid, on cytoskeletal elements, viability and functionality of rat liver and intestinal cells. Toxicon 2001, 39, 349–362. [Google Scholar] [CrossRef]
- Fessard, V.; Grosse, Y.; Pfohl-Leszkowicz, A.; Puiseux-Dao, S. Okadaic acid treatment induces DNA adduct formation in BHK21 C13 fibroblasts and HESV keratinocytes. Mutat. Res. 1996, 361, 133–141. [Google Scholar] [CrossRef]
- Hégarat, L.L.; Orsière, T.; Botta, A.; Fessard, V. Okadaic acid: Chromosomal non-disjunction analysis in human lymphocytes and study of aneugenic pathway in CHO-K1 cells. Mutat. Res. 2005, 578, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Louzao, M.C.; Fernández, D.A.; Abal, P.; Fraga, M.; Vilariño, N.; Vieytes, M.R.; Botana, L.M. Diarrhetic effect of okadaic acid could be related with its neuronal action: Changes in neuropeptide y. Toxicol. Lett. 2015, 237, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Bechemin, C.; Grzebyk, D.; Hachame, F.; Hummert, C.; Maestrini, S.Y. Effect of different nitrogen/phosphorus nutrient ratios on the toxin content in Alexandrium minutum. Aquat. Microb. Ecol. 1999, 20, 157–165. [Google Scholar] [CrossRef]
- Flynn, K.; Franco, J.M.; Fernandez, P.; Reguera, B.; Zapata, M.; Wood, G.; Flynn, K.J. Changes in toxin content, biomass and pigments of the dinoflagellate Alexandrium minutum during nitrogen refeeding and growth into nitrogen or phosphorus stress. Mar. Ecol. Prog. Ser. 1994, 111, 99–109. [Google Scholar] [CrossRef]
- Hwang, D.F.; Lu, Y.H. Influence of environmental and nutritional factors on growth, toxicity, and toxin profile of dinoflagellate Alexandrium minutum. Toxicon 2000, 38, 1491–1503. [Google Scholar] [CrossRef]
- John, E.H.; Flynn, K.J. Growth dynamics and toxicity of Alexandrium fundyense (Dinophyceae): The effect of changing n:P supply ratios on internal toxin and nutrient levels. Eur. J. Phycol. 2000, 35, 11–23. [Google Scholar] [CrossRef]
- Oh, S.J.; Yamamoto, T.; Kataoka, Y.; Matsuda, O.; Matsuyama, Y.; Kotani, Y. Utilization of dissolved organic phosphorus by the two toxic dinoflagellates, Alexandrium tamarense and Gymnodinium catenatum (dinophyceae). Fish. Sci. 2002, 68, 416–424. [Google Scholar] [CrossRef]
- Wang, D.Z.; Hsieh, D.P.H. Effects of nitrate and phosphate on growth and C2 toxin productivity of Alexandrium tamarense CI01 in culture. Mar. Pollut. Bull. 2002, 45, 286–289. [Google Scholar] [CrossRef]
- Murata, A.; Leong, S.C.Y.; Nagashima, Y.; Taguchi, S. Nitrogen:Phosphorus supply ratio may control the protein and total toxin of dinoflagellate Alexandrium tamarense. Toxicon 2006, 48, 683–689. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.C.H.; Kwok, O.T.; Ho, K.C.; Lee, F.W.F. Effects of different nitrate and phosphate concentrations on the growth and toxin production of an Alexandrium tamarense strain collected from Drake passage. Mar. Environ. Res. 2012, 81, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Ho, A.Y.T.; He, L.; Yin, K.; Hung, C.; Choi, N.; Lam, P.K.S.; Wu, R.S.S.; Anderson, D.M.; Harrison, P.J. Effects of inorganic and organic nitrogen and phosphorus on the growth and toxicity of two Alexandrium species from hong kong. Harmful Algae 2012, 16, 89–97. [Google Scholar] [CrossRef]
- Morton, S.L.; Norris, D.R. Role of temperature, salinity, and light on the seasonality of Prorocentrum lima (ehrenberg) dodge. In Toxic Marine Phytoplankton: Proceedings of the Fourth International Conference on Toxic Marine Phytoplankton; Granéli, E., Sundstorm, B., Edler, L., Anderson, D.M., Eds.; Elsevier: Lund, Sweden, 1990. [Google Scholar]
- Morton, S.L.; Norris, D.R.; Bomber, J.W. Effect of temperature, salinity and light intensity on the growth and seasonality of toxic dinoflagellates associated with ciguatera. J. Exp. Mar. Biol. Ecol. 1992, 157, 79–90. [Google Scholar] [CrossRef]
- Pan, Y.; Cembella, A.D.; Quilliam, M.A. Cell cycle and toxin production in the benthic dinoflagellate Prorocentrum lima. Mar. Biol. 1999, 134, 541–549. [Google Scholar] [CrossRef]
- Park, M.G.; Kim, S.; Kim, H.S.; Myung, G.; Kang, Y.G.; Yih, W. First successful culture of the marine dinoflagellate Dinophysis acuminata. Aquat. Microb. Ecol. 2006, 45, 101–106. [Google Scholar] [CrossRef]
- Nielsen, L.T.; Krock, B.; Hansen, P.J. Production and excretion of okadaic acid, pectenotoxin-2 and a novel dinophysistoxin from the DSP-causing marine dinoflagellate Dinophysis acuta—Effects of light, food availability and growth phase. Harmful Algae 2013, 23, 34–45. [Google Scholar] [CrossRef]
- Tong, M.; Kulis, D.M.; Fux, E.; Smith, J.L.; Hess, P.; Zhou, Q.; Anderson, D.M. The effects of growth phase and light intensity on toxin production by Dinophysis acuminata from the Northeastern United States. Harmful Algae 2011, 10, 254–264. [Google Scholar] [CrossRef]
- Smith, J.L.; Tong, M.; Fux, E.; Anderson, D.M. Toxin production, retention, and extracellular release by Dinophysis acuminata during extended stationary phase and culture decline. Harmful Algae 2012, 19, 125–132. [Google Scholar] [CrossRef]
- Lourenço, S.O.; Barbarino, E.; Lavín, P.L.; Lanfer Marquez, U.M.; Aidar, E. Distribution of intracellular nitrogen in marine microalgae: Calculation of new nitrogen-to-protein conversion factors. Eur. J. Phycol. 2004, 39, 17–32. [Google Scholar] [CrossRef]
- Lai, J.; Yu, Z.; Song, X.; Cao, X.; Han, X. Responses of the growth and biochemical composition of Prorocentrum donghaiense to different nitrogen and phosphorus concentrations. J. Exp. Mar. Biol. Ecol. 2011, 405, 6–17. [Google Scholar] [CrossRef]
- Cary, L.B.; Jonathan, R.P. The effect of nitrogen source on the growth and toxicity of species of the genus Prorocentrum. In Symposium on Harmful Marine Algae in the U.S.; Marine Biological Laboratory: Woods Hole, MA, USA, 2000; p. 25. [Google Scholar]
- Souto, M.L.; Fernández, J.J.; Norte, M.; Fernández, M.L.; Martı́nez, A. Influence of amino acids on okadaic acid production. Toxicon 2001, 39, 659–664. [Google Scholar] [CrossRef]
- Aikman, K.E.; Tindall, D.R.; Morton, S.L. Physiology, potency of the dinoflagellate Prorocentrum hoffmannianum (faust) during one complete growth cycle. In Toxic Phytoplankton Blooms in the Sea; Smayda, T.J., Shimizu, Y., Eds.; Elsevier: Amsterdam, The Netherlands, 1993; pp. 463–468. [Google Scholar]
- Morel, F.M.M.; Milligan, A.J.; Saito, M.A. Marine bioinorganic chemistry: The role of trace metals in the oceanic cycles of major nutrients. In The Oceans and Marine Geochemistry; Elderfield, H., Holland, H.D., Turekian, K.K., Eds.; Elsevier Science Limited: Kidlington, Oxford, UK, 2006; pp. 113–140. [Google Scholar]
- Azad, H.S.; Borchardt, J.A. Variations in phosphorus uptake by algae. Environ. Sci. Technol. 1970, 4, 737–743. [Google Scholar] [CrossRef]
- Nascimento, S.M.; Purdie, D.A.; Morris, S. Morphology, toxin composition and pigment content of Prorocentrum lima strains isolated from a coastal lagoon in Southern UK. Toxicon 2005, 45, 633–649. [Google Scholar] [CrossRef] [PubMed]
- Glibert, P.M.; Legrand, C. The diverse nutrient strategies of harmful algae: Focus on osmotrophy. In Ecology of Harmful Algae; Granéli, E., Turner, J.T., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; Volume 189, pp. 163–175. [Google Scholar]
- Andersen, R.A. Algal Culturing Techniques, 1st ed.; Elsevier: San Diego, CA, USA, 2005; p. 596. [Google Scholar]
- Tindall, D.R.; Morton, S.L. Community dynamics and physiology of epiphytic/benthic dinoflagellates associated with ciguatera. In Physiological Ecology of Harmful Algal Blooms; Anderson, D.M., Cembella, A.D., Hallegraeff, G.M., Eds.; Springer: Berlin, Germany, 1998. [Google Scholar]
- Faust, M.A. Three new benthic species of Prorocentrum (dinophyceae) from Twin Cays, Belize: P. maculosum sp. nov., P. foraminosum sp. nov. and P. formosum sp. nov. Phycologia 1993, 32, 410–418. [Google Scholar] [CrossRef]
- Aligizaki, K.; Nikolaidis, G.; Katikou, P.; Baxevanis, A.D.; Abatzopoulos, T.J. Potentially toxic epiphytic Prorocentrum (Dinophyceae) species in greek coastal waters. Harmful Algae 2009, 8, 299–311. [Google Scholar] [CrossRef]
- Glibert, P.M.; Burkholder, J.M.; Kana, T.M. Recent insights about relationships between nutrient availability, forms, and stoichiometry, and the distribution, ecophysiology, and food web effects of pelagic and benthic Prorocentrum species. Harmful Algae 2012, 14, 231–259. [Google Scholar] [CrossRef]
- Koike, K.; Sato, S.; Yamaji, M.; Nagahama, Y.; Kotaki, Y.; Ogata, T.; Kodama, M. Occurrence of okadaic acid-producing Prorocentrum lima on the sanriku coast, northern japan. Toxicon 1998, 36, 2039–2042. [Google Scholar] [CrossRef]
- Samuelsson, G.; Richardson, K. Photoinhibition at low quantum flux densities in a marine dinoflagellate (Amphidinium carterae). Mar. Biol. 1982, 70, 21–26. [Google Scholar] [CrossRef]
- Woodward, F.I.; Sheehy, J.E. Radiation. In Principles and Measurements in Environmental Biology; Woodward, F.I., Sheehy, J.E., Eds.; Elsevier Science: Gateshead, UK, 1983; p. 35. [Google Scholar]
- Kiang, N.Y.; Siefert, J.; Blankenship, R.E. Spectral signatures of photosynthesis. I. Review of earth organisms. Astrobiology 2007, 7, 222–251. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Iida, S.; Takaichi, S.; Mimuro, M.; Murakami, A.; Akimoto, S. Excitation relaxation dynamics and energy transfer in pigment–protein complexes of a dinoflagellate, revealed by ultrafast fluorescence spectroscopy. Photosynth. Res. 2016, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Rausch de Traubenberg, C.; Géraud, M.L.; Soyer-Gobillard, M.O.; Emdadi, D. The toxic dinoflagellate Prorocentrum lima and its associated bacteria: I. An ultrastructural study. Eur. J. Protistol. 1995, 31, 318–326. [Google Scholar] [CrossRef]
- Prokic, I.; Brümmer, F.; Brigge, T.; Görtz, H.D.; Gerdts, G.; Schütt, C.; Elbrächter, M.; Müller, W.E.G. Bacteria of the genus Roseobacter associated with the toxic dinoflagellate Prorocentrum lima. Protist 1998, 149, 347–357. [Google Scholar] [CrossRef]
- Lafay, B.; Ruimy, R.; Rausch De Traubenber, C.; Breittmayer, V.; Gauthier, M.J.; Christen, R. Roseobacter algicola sp. Nov., a new marine bacterium isolated from the phycosphere of the toxin-producing dinoflagellate Prorocentrum lima. Int. J. Syst. Bacteriol. 1995, 45, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Rausch de Traubenberg, C. Interactions between a Dinoflagellate and It Associated Bacterial Microflora: Role of Bacteria in the Toxicity of Prorocentrun lima Ehrenberg (Dodge). Ph.D. Thesis, Université de Nantes, Lombardie, Eyjeaux, France, December 1993. [Google Scholar]
- Barbier, M.; Amzil, Z.; Mondeguer, F.; Bhaud, Y.; Soyer-Gobillard, M.-O.; Lassus, P. Okadaic acid and PP2A cellular immunolocalization in Prorocentrum lima (dinophyceae). Phycologia 1999, 38, 41–46. [Google Scholar] [CrossRef]
- Spector, D.L. Dinoflagellate nuclei. In Dinoflagellates; Spector, D.L., Ed.; Academic: Orlando, FL, USA, 1984; pp. 107–147. [Google Scholar]
- Rizzo, P.J. Biochemistry of the dinoflagellate nucleus. In The Biology of Dinoflagellates; Taylor, F.J.R., Ed.; Blackwell Scientific Publications: Palo Alto, CA, USA, 1987; pp. 143–173. [Google Scholar]
- Hackett, J.D.; Anderson, D.M.; Erdner, D.L.; Bhattacharya, D. Dinoflagellates: A remarkable evolutionary experiment. Am. J. Bot. 2004, 91, 1523–1534. [Google Scholar] [CrossRef] [PubMed]
- McLean, T.I. “Eco-omics”: A review of the application of genomics, transcriptomics, and proteomics for the study of the ecology of harmful algae. Microb. Ecol. 2013, 65, 901–915. [Google Scholar] [CrossRef] [PubMed]
- Yang, I.; John, U.; Beszteri, S.; Glockner, G.; Krock, B.; Goesmann, A.; Cembella, A. Comparative gene expression in toxic versus non-toxic strains of the marine dinoflagellate Alexandrium minutum. BMC Genom. 2010, 11, 248. [Google Scholar] [CrossRef] [PubMed]
- Yang, I.; Beszteri, S.; Tillmann, U.; Cembella, A.; John, U. Growth- and nutrient-dependent gene expression in the toxigenic marine dinoflagellate Alexandrium minutum. Harmful Algae 2011, 12, 55–69. [Google Scholar] [CrossRef]
- Yang, I.; Beszteri, S.; Tillmann, U.; Cembella, A.; John, U. Physiollogical and gene expression response to salinity stress in Alexandrium minutum. In Proceedings of the 13th International Conference on Harmful Algae; Ho, K.C., Zhou, M.J., Qi, Y.Z., Eds.; Environmental Publication House Hong Kong: Hong Kong, 2008; pp. 180–183. [Google Scholar]
- Morey, J.; Monroe, E.; Kinney, A.; Beal, M.; Johnson, J.; Hitchcock, G.; Van Dolah, F. Transcriptomic response of the red tide dinoflagellate, Karenia brevis, to nitrogen and phosphorus depletion and addition. BMC Genom. 2011, 12, 346. [Google Scholar] [CrossRef] [PubMed]
- Bayer, T.; Aranda, M.; Sunagawa, S.; Yum, L.K.; DeSalvo, M.K.; Lindquist, E.; Coffroth, M.A.; Voolstra, C.R.; Medina, M. Symbiodinium transcriptomes: Genome insights into the dinoflagellate symbionts of reef-building corals. PLoS ONE 2012, 7, e35269. [Google Scholar] [CrossRef] [PubMed]
- Lowe, C.; Mello, L.; Samatar, N.; Martin, L.; Montagnes, D.; Watts, P. The transcriptome of the novel dinoflagellate Oxyrrhis marina (Alveolata: Dinophyceae): Response to salinity examined by 454 sequencing. BMC Genom. 2011, 12, 519. [Google Scholar] [CrossRef] [PubMed]
- Jaeckisch, N.; Yang, I.; Wohlrab, S.; Glöckner, G.; Kroymann, J.; Vogel, H.; Cembella, A.; John, U. Comparative genomic and transcriptomic characterization of the toxigenic marine dinoflagellate Alexandrium ostenfeldii. PLoS ONE 2011, 6, e28012. [Google Scholar] [CrossRef] [PubMed]
- Toulza, E.; Shin, M.S.; Blanc, G.; Audic, S.; Laabir, M.; Collos, Y.; Claverie, J.-M.; Grzebyk, D. Gene expression in proliferating cells of the dinoflagellate Alexandrium catenella (dinophyceae). Appl. Environ. Microb. 2010, 76, 4521–4529. [Google Scholar] [CrossRef] [PubMed]
- Wisecaver, J.; Hackett, J. Transcriptome analysis reveals nuclear-encoded proteins for the maintenance of temporary plastids in the dinoflagellate Dinophysis acuminata. BMC Genom. 2010, 11, 366. [Google Scholar] [CrossRef] [PubMed]
- Hackett, J.; Scheetz, T.; Yoon, H.; Soares, M.; Bonaldo, M.; Casavant, T.; Bhattacharya, D. Insights into a dinoflagellate genome through expressed sequence tag analysis. BMC Genom. 2005, 6, 80. [Google Scholar] [CrossRef] [PubMed]
- Tanikawa, N.; Akimoto, H.; Ogoh, K.; Chun, W.; Ohmiya, Y. Expressed sequence tag analysis of the dinoflagellate Lingulodinium polyedrum during dark phase. Photochem. Photobiol. 2004, 80, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Lidie, K.B.; Ryan, J.C.; Barbier, M.; Dolah, F.M. Gene expression in florida red tide dinoflagellate Karenia brevis: Analysis of an expressed sequence tag library and development of DNA microarray. Mar. Biotechnol. 2005, 7, 481–493. [Google Scholar] [CrossRef] [PubMed]
- Velculescu, V.E.; Zhang, L.; Vogelstein, B.; Kinzler, K.W. Serial analysis of gene expression. Science 1995, 270, 484–487. [Google Scholar] [CrossRef] [PubMed]
- Brenner, S.; Johnson, M.; Bridgham, J.; Golda, G.; Lloyd, D.H.; Johnson, D.; Luo, S.; McCurdy, S.; Foy, M.; Ewan, M.; et al. Gene expression analysis by massively parallel signature sequencing (MPSS) on microbead arrays. Nat. Biotechnol. 2000, 18, 630–634. [Google Scholar] [CrossRef] [PubMed]
- Dyhrman, S.T. Molecular approaches to diagnosing nutritional physiology in harmful algae: Implications for studying the effects of eutrophication. Harmful Algae 2008, 8, 167–174. [Google Scholar] [CrossRef]
- Coyne, K.J.; Burkholder, J.M.; Feldman, R.A.; Hutchins, D.A.; Cary, S.C. Modified serial analysis of gene expression method for construction of gene expression profiles of microbial eukaryotic species. Appl. Environ. Microb. 2004, 70, 5298–5304. [Google Scholar] [CrossRef] [PubMed]
- Moustafa, A.; Evans, A.N.; Kulis, D.M.; Hackett, J.D.; Erdner, D.L.; Anderson, D.M.; Bhattacharya, D. Transcriptome profiling of a toxic dinoflagellate reveals a gene-rich protist and a potential impact on gene expression due to bacterial presence. PLoS ONE 2010, 5, e9688. [Google Scholar] [CrossRef] [PubMed]
- Erdner, D.; Anderson, D. Global transcriptional profiling of the toxic dinoflagellate Alexandrium fundyense using massively parallel signature sequencing. BMC Genom. 2006, 7, 88. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Sui, Z.; Chang, L.; Kang, K.; Ma, J.; Kong, F.; Zhou, W.; Wang, J.; Guo, L.; Geng, H.; et al. Transcriptome de novo assembly sequencing and analysis of the toxic dinoflagellate Alexandrium catenella using the illumina platform. Gene 2014, 537, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, S.F.; Lin, L.; Wang, D.Z. Comparative transcriptome analysis of a toxin-producing dinoflagellate Alexandrium catenella and its non-toxic mutant. Mar. Drugs 2014, 12, 5698–5718. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, O.K.; Robertson, D.L.; Fagan, T.F.; Hastings, J.W.; Colepicolo, P. Different regulatory mechanisms modulate the expression of a dinoflagellate iron-superoxide dismutase. J. Biol. Chem. 2001, 276, 19989–19993. [Google Scholar] [CrossRef] [PubMed]
- Fagan, T.; Morse, D.; Hastings, J.W. Circadian synthesis of a nuclear-encoded chloroplast glyceraldehyde-3-phosphate dehydrogenase in the dinoflagellate Gonyaulax polyedra is translationally controlled. Biochemistry 1999, 38, 7689–7695. [Google Scholar] [CrossRef] [PubMed]
- Van Dolah, F.M.; Leighfield, T.A.; Sandel, H.D.; Hsu, C.K. Cell division in the dinoflagellate Gambierdiscus toxicus is phased to the diurnal cycle and accompanied by activation of the cell cycle regulatory protein, cdc2 kinase1. J. Phycol. 1995, 31, 395–400. [Google Scholar] [CrossRef]
- Lin, S.; Zhang, H.; Gray, M.W. RNA editing in dinoflagellates and its implications for the evolutionary history of the editing machinery. In RNA and DNA Editing: Molecular Mechanisms and Their Integration into Biological Systems; Harold, C.S., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2008; pp. 280–309. [Google Scholar]
- Lee, F.W.F.; Morse, D.; Lo, S.C.L. Identification of two plastid proteins in the dinoflagellate Alexandrium affine that are substantially down-regulated by nitrogen-depletion. J. Proteome Res. 2009, 8, 5080–5092. [Google Scholar] [CrossRef] [PubMed]
- Lee, F.W.F.; Lo, S.C.L. The use of trizol reagent (phenol/guanidine isothiocyanate) for producing high quality two-dimensional gel electrophoretograms (2-DE) of dinoflagellates. J. Microbiol. Methods 2008, 73, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.-Z.; Zhang, Y.-J.; Zhang, S.-F.; Lin, L.; Hong, H.-S. Quantitative proteomic analysis of cell cycle of the dinoflagellate Prorocentrum donghaiense (dinophyceae). PLoS ONE 2013, 8, e63659. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.Z.; Li, C.; Xie, Z.X.; Dong, H.P.; Lin, L.; Hong, H.S. Homology-Driven proteomics of dinoflagellates with unsequenced genomes using MALDI-TOF/TOF and automated de novo sequencing. Evid. Based Complement. Altern. Med. 2011, 2011, 16. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Wang, D.Z.; Dong, H.P.; Xie, Z.X.; Hong, H.S. Proteomics of a toxic dinoflagellate Alexandrium catenella DH01: Detection and identification of cell surface proteins using fluorescent labeling. Chin. Sci. Bull. 2012, 57, 3320–3327. [Google Scholar] [CrossRef]
- Wang, D.Z.; Li, C.; Zhang, Y.; Wang, Y.Y.; He, Z.P.; Lin, L.; Hong, H.S. Quantitative proteomic analysis of differentially expressed proteins in the toxicity-lost mutant of Alexandrium catenella (Dinophyceae) in the exponential phase. J. Proteom. 2012, 75, 5564–5577. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.Z.; Gao, Y.; Lin, L.; Hong, H.S. Comparative proteomic analysis reveals proteins putatively involved in toxin biosynthesis in the marine dinoflagellate Alexandrium catenella. Mar. Drugs 2013, 11, 213–232. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Lo, S.C.-L.; Matton, D.P.; Lang, B.F.; Morse, D. Daily changes in the phosphoproteome of the dinoflagellate Lingulodinium. Protist 2012, 163, 746–754. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.Z.; Lin, L.; Chan, L.L.; Hong, H.S. Comparative studies of four protein preparation methods for proteomic study of the dinoflagellate Alexandrium sp. Using two-dimensional electrophoresis. Harmful Algae 2009, 8, 685–691. [Google Scholar] [CrossRef]
- Lee, T.C.H.; Ho, K.C.; Xu, S.J.l.; Lee, F.W.F. Applications and challenges of proteomic technology in the study of harmful algal blooms (habs). In International Conference on Biodiversity and Conservation of Wetland in South China—From Preservation to Green Development; Environmental Publication House Hong Kong: Hong Kong, China, 2014; pp. 53–64. [Google Scholar]
- Chan, L.L.; Sit, W.H.; Lam, P.K.S.; Hsieh, D.P.H.; Hodgkiss, I.J.; Wan, J.M.F.; Ho, A.Y.T.; Choi, N.M.C.; Wang, D.Z.; Dudgeon, D. Identification and characterization of a “biomarker of toxicity” from the proteome of the paralytic shellfish toxin-producing dinoflagellate Alexandrium tamarense (Dinophyceae). Proteomics 2006, 6, 654–666. [Google Scholar] [CrossRef] [PubMed]
- Chan, L.L.; Hodgkiss, I.J.; Lam, P.K.S.; Wan, J.M.F.; Chou, H.N.; Lum, J.H.K.; Lo, M.G.Y.; Mak, A.S.C.; Sit, W.H.; Lo, S.C.L. Use of two-dimensional gel electrophoresis to differentiate morphospecies of Alexandrium minutum, a paralytic shellfish poisoning toxin-producing dinoflagellate of harmful algal blooms. Proteomics 2005, 5, 1580–1593. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.Z.; Lin, L.; Wang, M.H.; Li, C.; Hong, H.S. Proteomic analysis of a toxic dinoflagellate Alexandrium catenella under different growth phases and conditions. Chin. Sci. Bull. 2012, 57, 3328–3341. [Google Scholar] [CrossRef]
- Li, C.; Zhang, Y.; Xie, Z.X.; He, Z.P.; Lin, L.; Wang, D.Z. Quantitative proteomic analysis reveals evolutionary divergence and species-specific peptides in the Alexandrium tamarense complex (dinophyceae). J. Proteom. 2013, 86, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Snyder, R.V.; Gibbs, P.D.L.; Palacios, A.; Abiy, L.; Dickey, R.; Lopez, J.V.; Rein, K.S. Polyketide synthase genes from marine dinoflagellates. Mar. Biotechnol. 2003, 5, 1–12. [Google Scholar] [PubMed]
- Tang, J.Q.; Li, T.; Yang, W.D.; Liu, J.S.; Li, H.Y. Cloning and analysis of PKS gene from Prorocentrum lima. Acta Ecol. Sin. 2009, 29, 2383–2390. [Google Scholar]
- Ivanka, T.; Plamen, S.; Detelina, B.; Ivanka, D.D.; Rumen, M.; Balik, D. Production of cyanobacterial toxins from two Nostoc species (Nostocales) and evaluation of their cytotoxicity in vitro. J. BioSci. Biotechnol. 2012, 1, 33–43. [Google Scholar]
- Christiansen, G.; Fastner, J.; Erhard, M.; Börner, T.; Dittmann, E. Microcystin biosynthesis in planktothrix: Genes, evolution, and manipulation. J. Bacteriol. 2003, 185, 564–572. [Google Scholar] [CrossRef] [PubMed]
- Tillett, D.; Dittmann, E.; Erhard, M.; von Döhren, H.; Börner, T.; Neilan, B.A. Structural organization of microcystin biosynthesis in Microcystis aeruginosa PCC7806: An integrated peptide–polyketide synthetase system. Chem. Biol. 2000, 7, 753–764. [Google Scholar] [CrossRef]
- Norte, M.; Padilla, A.; Fernández, J.J. Studies on the biosynthesis of the polyether marine toxin dinophysistoxin-1 (DTX-1). Tetrahedron Lett. 1994, 35, 1441–1444. [Google Scholar] [CrossRef]
- Macpherson, G.R.; Burton, I.W.; LeBlanc, P.; Walter, J.A.; Wright, J.L.C. Studies of the biosynthesis of DTX-5a and DTX-5b by the dinoflagellate Prorocentrum maculosum: Regiospecificity of the putative Baeyer−villigerase and insertion of a single amino acid in a polyketide chain. J. Org. Chem. 2002, 68, 1659–1664. [Google Scholar] [CrossRef] [PubMed]
- Needham, J.; Hu, T.; McLachlan, J.L.; Walter, J.A.; Wright, J.L.C. Biosynthetic studies of the DSP toxin DTX-4 and an okadaic acid diol ester. J. Chem. Soc. Chem. Commun. 1995, 1623–1624. [Google Scholar] [CrossRef]
- Wright, J.L.C.; Hu, T.; McLachlan, J.L.; Needham, J.; Walter, J.A. Biosynthesis of DTX-4: Confirmation of a polyketide pathway, proof of a Baeyer–villiger oxidation step, and evidence for an unusual carbon deletion process. J. Am. Chem. Soc. 1996, 118, 8757–8758. [Google Scholar] [CrossRef]
- Daranas, A.H.; Fernández, J.J.; Norte, M.; Gavín, J.A.; Suárez-Gómez, B.; Souto, M.L. Biosynthetic studies of the DSP toxin skeleton. Chem. Rec. 2004, 4, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Nishizawa, T.; Ueda, A.; Asayama, M.; Fujii, K.; Harada, K.-I.; Ochi, K.; Shirai, M. Polyketide synthase gene coupled to the peptide synthetase module involved in the biosynthesis of the cyclic heptapeptide microcystin. J. Biochem. 2000, 127, 779–789. [Google Scholar] [CrossRef] [PubMed]
- Kaebernick, M.; Neilan, B.A.; Börner, T.; Dittmann, E. Light and the transcriptional response of the microcystin biosynthesis gene cluster. Appl. Environ. Microb. 2000, 66, 3387–3392. [Google Scholar] [CrossRef]
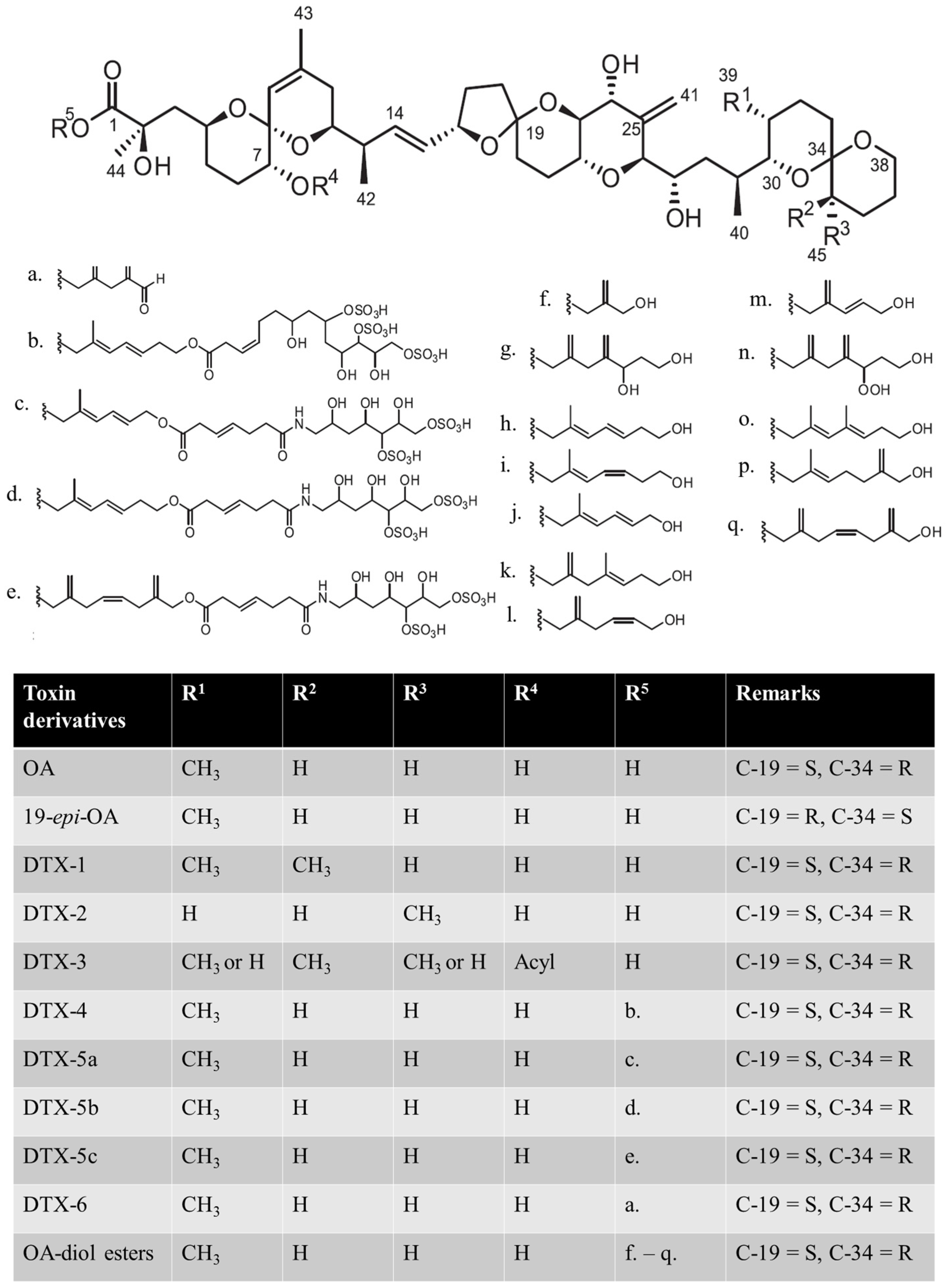
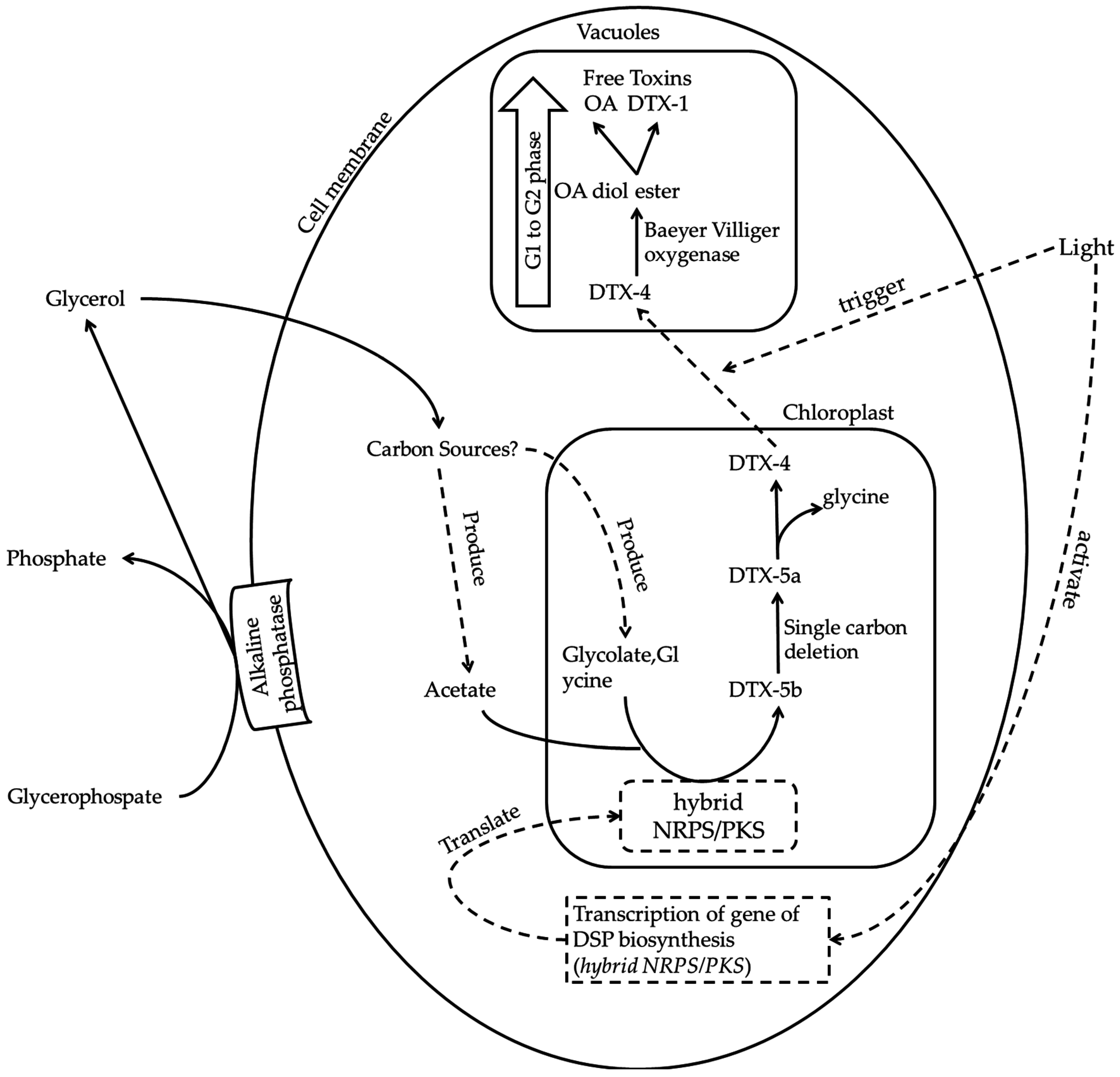
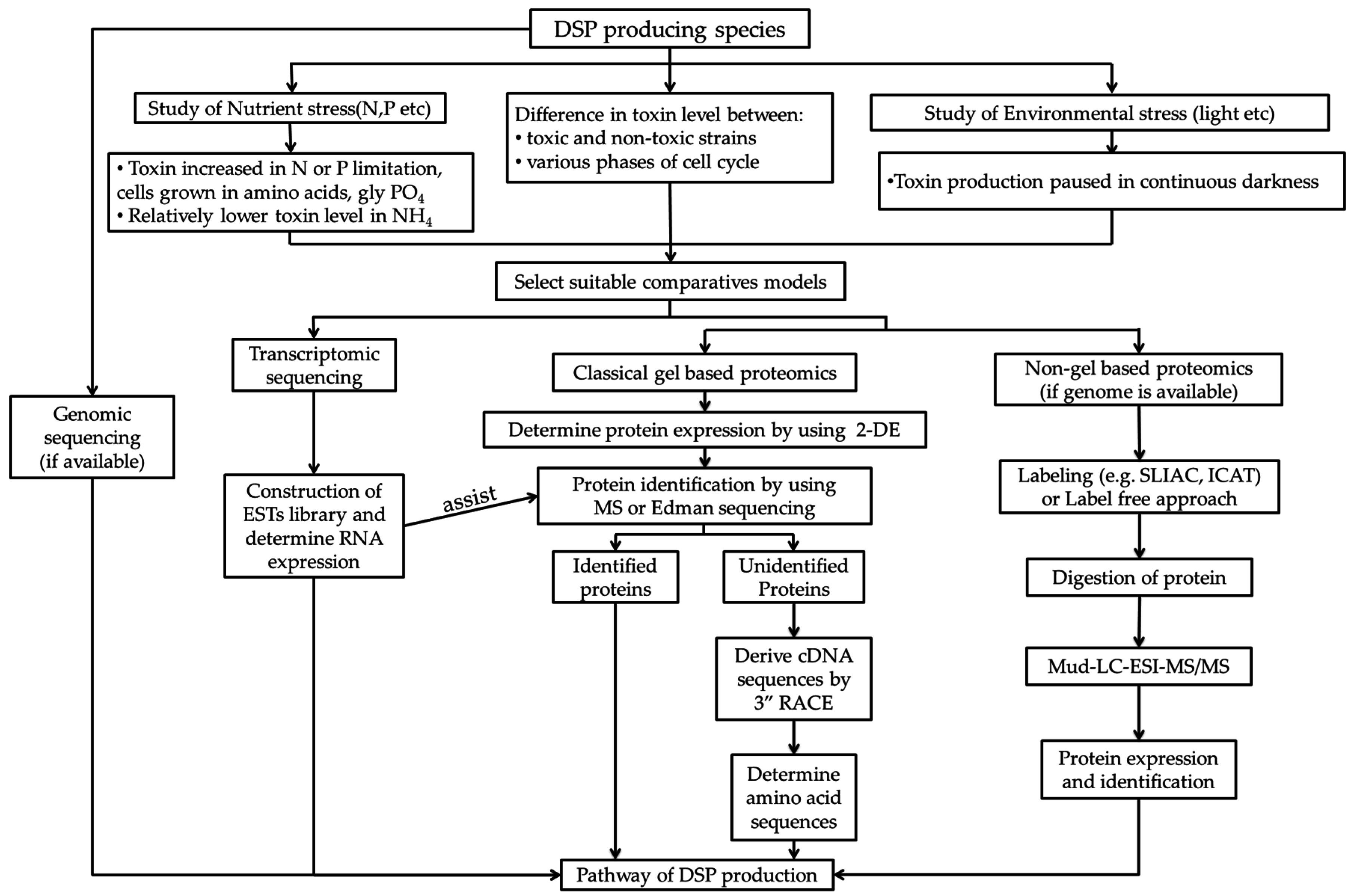

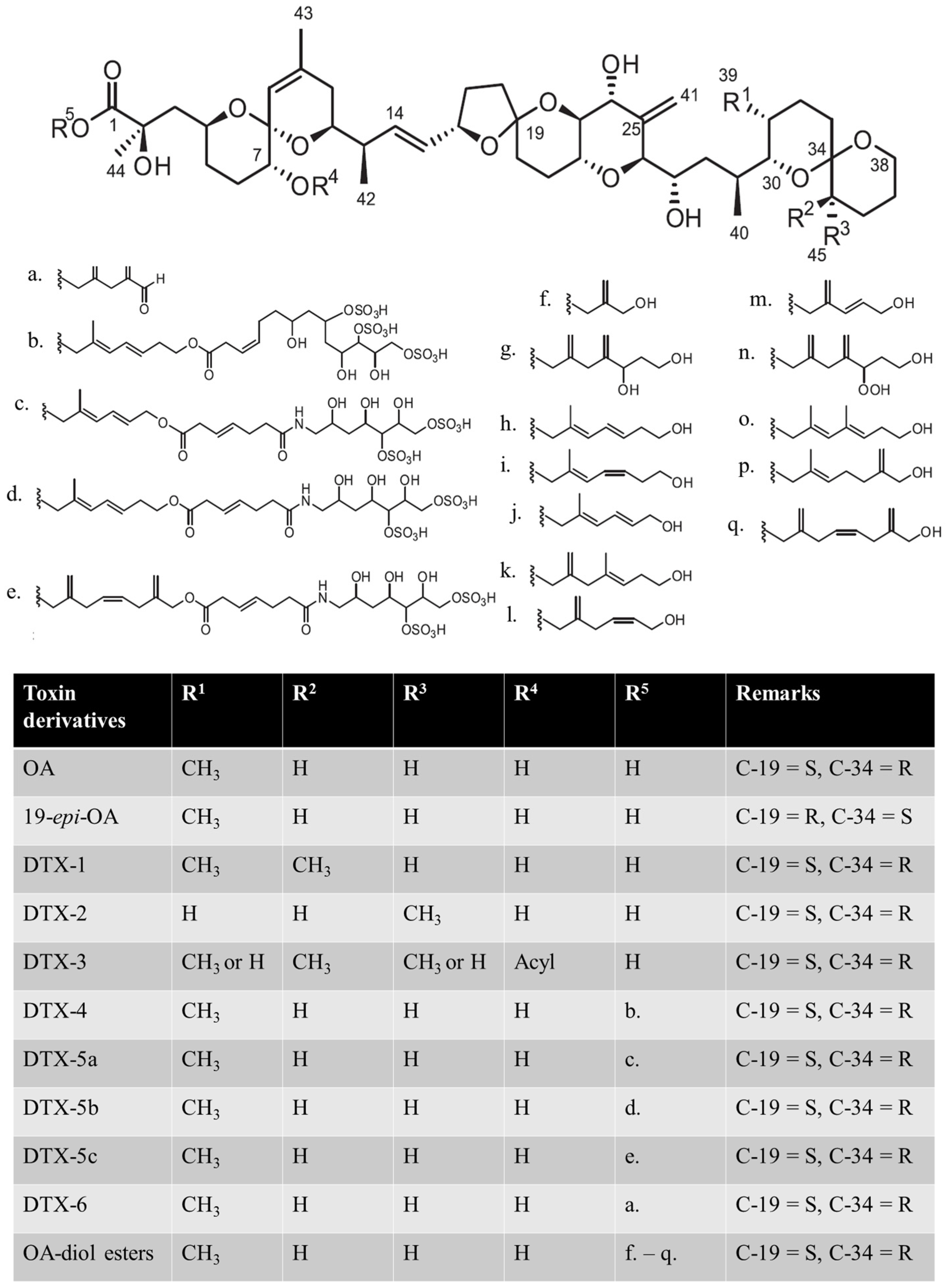{kind=link}
{kind=link}
{kind=link}
| Strains | N Source: P Source | Initial N (μM):P (μM) | N/P | Growth Rate (μ, Day−1) | Max. Cell Density (Cells/mL) | Cellular Toxin Level (pg/Cell) | Reference | |
|---|---|---|---|---|---|---|---|---|
| P. lima (Isolates from coastal lagoon of Goro) | NO3:PO4 | 17.7:36.3 | 0.49 |  | 0.22 | 7040 | OA: 12.5 b, DTX-1:0.39 b | [15] |
| 44.2:36.3 | 1.22 | ~10000 c | OA: ~11 b,c, DTX-1: ~0.25 b,c | |||||
| 88.3:36.3 | 2.43 | ~14000 c | OA: ~11.74 b,c, DTX-1: ~0.18 b,c | |||||
| 294:36.3 | 8.10 | ~24000 c | OA: ~10.34 b,c, DTX-1: ~ 0.15 b,c | |||||
| 882:36.3 | 24.30 | 0.23 | 33100–35000 | OA: 6.69–6.87 b, DTX-1: 0.12 b | ||||
| P. lima (CCAP1136/11) | NO3:PO4 | 882:36.3 | 24.30 | 0.22 | 20950 | OA: ~6 b,c | [16] | |
| NH4:PO4 | 882:36.3 | 24.30 | 0.21 | 10790 | OA:~4.7 b,c | |||
| P. lima (CCMP2579) | NO3:PO4 | 88.2:36.3 | 2.43 | ~0.029 a | ~5220 c | OA: 192.69 b | [12] | |
| 882:36.3 | 24.30 | ~0.039 a | ~10000 c | OA: 100.66 b | ||||
| P. lima (CCMP2579) | NO3:PO4 | 12:36.3 | 0.33 | | 0.058 | ~13180 c | OA: ~210 b,c | [14] |
| 25:36.3 | 0.69 | ~13180 c | OA: ~275 b,c | |||||
| 50:36.3 | 1.38 | ~14540 c | OA: ~240 b,c | |||||
| 100:36.3 | 2.75 | ~15000 c | OA: ~338 b,c | |||||
| NH4:PO4 | 12:36.3 | 0.33 | | 0.059 | ~13210 c | OA: ~85 b,c | ||
| 25:36.3 | 0.69 | ~11740 c | OA: ~68 b,c | |||||
| 50:36.3 | 1.38 | ~12500 c | OA: ~85 b,c | |||||
| 100:36.3 | 2.75 | ~13620 c | OA: ~100 b,c | |||||
| Urea:PO4 | 12:36.3 | 0.33 | | 0.060 | ~13880 c | OA: ~100 b,c | ||
| 25:36.3 | 0.69 | ~13330 c | OA: ~58 b,c | |||||
| 50:36.3 | 1.38 | ~15550 c | OA: ~55 b,c | |||||
| 100:36.3 | 2.75 | 18330 | OA: ~113 b,c | |||||
| P. lima (Clone from Mahone Bay, Nova Scotia) | NO3:PO4 | 0:36.3 | 0.00 | ~0.013 a | ~8077 c | OA+DTX-1 at Day 20: 34 b | [17] | |
| 300:36.3 | 8.26 | ~0.032 a | ~20000 c | OA+DTX-1 at Day 30: 25 b | ||||
| 1000:36.3 | 27.55 | ~0.033 a | ~20833 c | OA+DTX-1 at Day 30: 7 b | ||||
| P. lima (CCMP2579) | NO3:PO4 | 17.7:36.3 | 0.49 | ~0.103 a | ~35454 c | OA: ~30.1 b,c | [23] | |
| 882:36.3 | 24.30 | ~0.097 a | ~40000 c | OA: ~17.5 b,c | ||||
| Strains | N Source:P Source | Initial N(μM):P(μM) | N/P | Growth Rate (μ, day−1) | Max. Cell Density (cells/mL) | Cellular Toxin Level (pg/cell) | Reference | |
|---|---|---|---|---|---|---|---|---|
| P. lima (Isolates from coastal lagoon of Goro) | NO3:PO4 | 882:0.73 | 1208.22 | | 0.22 | 6250 | OA: 10.75 b, DTX-1: ~ 0.26 b,c | [15] |
| 882:1.81 | 487.29 | ~11,000 c | OA: 15.80 b, DTX-1: 0.32 b | |||||
| 882:3.63 | 242.98 | ~17,000 c | OA: ~11.27 b,c, DTX-1: ~ 0.26 b,c | |||||
| 882:12.1 | 72.89 | ~31,000 c | OA: ~9.7 b,c, DTX-1: ~ 0.16 b,c | |||||
| 882:36.3 | 24.30 | 0.23 | 33,100–35,000 | OA: 6.69–6.87 b, DTX-1: 0.13 b | ||||
| P. lima (CCMP2579) | NO3:PO4 | 882:3.63 | 242.98 | ~0.029 a | ~3333 c | OA: 268.68 b | [12] | |
| 882:36.3 | 24.30 | ~0.039 a | ~10,000 c | OA: 100.66 b | ||||
| P. lima (CCMP2579) | NO3:PO4 | 882:0.5 | 1764.00 | | 0.059 | ~12,230 c | OA: ~88 b c | [13] |
| 882:1 | 882.00 | ~13,290 c | OA: ~76 b,c | |||||
| 882:2 | 441.00 | ~14,200 c | OA: ~86 b,c | |||||
| 882:5 | 176.40 | ~15,000 c | OA: ~60 b,c | |||||
| 882:10 | 88.20 | ~16,000 c | OA: ~68 b,c | |||||
| NO3:Gly PO4 | 882: 0.5 | 1764.00 | | 0.048 | ~11,840 c | OA: ~116 b,c | ||
| 882:1 | 882.00 | ~11,840 c | OA: ~97 b,c | |||||
| 882:2 | 441.00 | ~11,440 c | OA: ~100 b,c | |||||
| 882:5 | 176.40 | ~13,600 c | OA: ~94 b,c | |||||
| 882:10 | 88.20 | ~16,310 c | OA: ~88 b,c | |||||
| NO3:ATP | 882:0.5 | 1764.00 | | 0.053 | ~13,160 c | OA:~51 b,c | ||
| 882:1 | 882.00 | ~13,830 c | OA: ~57 b,c | |||||
| 882:2 | 441.00 | ~17,000c | OA: ~57 b,c | |||||
| 882:5 | 176.40 | ~18,160 c | OA: ~60 b,c | |||||
| 882:10 | 88.20 | ~19,160 c | OA: ~57 b,c | |||||
| P. lima (Isolates from dry Tortugas, Florida) | NO3 + NH4:PO4 | 932:10 | 93.20 | ~0.5 a | 135,185 | OA at Day30: 8.9 b | [18] | |
| NO3 + NH4:gly PO4 | 932:10 | 93.20 | ~0.144 a | 221,445 | OA at Day30: 14.2 b | |||
| P. lima (CCMP685) | NO3:Gly PO4 | 882:10 | 88.20 | ~0.082 a | 26,637 at Day 40 | OA at Day47: 9.96 | [19] | |
| 882:20 | 44.10 | ~0.094 a | 43,128 at Day 40 | N/A | ||||
| 882: 30 | 29.40 | ~0.101 a | 55,903 at Day 40 | OA at Day47: ~4.4 c | ||||
| NO3:PO4 | 882:10 | 88.20 | ~0.08 a | 25,014 at Day 40 | OA at Day47: ~7 c | |||
| 882:20 | 44.10 | ~0.092 a | 39,997 at Day 40 | N/A | ||||
| 882:30 | 29.40 | ~0.098 a | 50,016 at Day 40 | OA at Day47: ~4 c | ||||
| P. lima (CCMP2579) | NO3:PO4 | 882:1.81 | 487.29 | ~0.021 a | ~25,909 c | OA: ~35.2 b,c | [23] | |
| 882:36.3 | 24.30 | ~0.097 a | ~40,000 c | OA: ~17.5 b,c | ||||
| Strains | Temperature (°C) | Growth Rate (μ, day−1) | Max. Cell Density (Cells/mL) | Cellular Toxin Level (pg/cell) | Reference |
|---|---|---|---|---|---|
| P. lima (Isolates from Nova Scotia, Canada) | 5 °C | ~0.000 a | ~1000 at Day 28 | OA at Day 28:8 | [20] |
| 10 °C | ~0.073 a | ~7800 at Day 28 | OA at Day 28:2.5 | ||
| 15 °C | ~0.150 a | ~11,000 at Day 16 | OA at Day 16:4.4 | ||
| 20 °C | ~0.198 a | ~23,800 at Day 16 | OA at Day 16:2.5 | ||
| 25 °C | ~0.229 a | ~39,000 at Day 16 | OA at Day 16:1.4 | ||
| P. lima (PL100A, Isolates from Knight Key, Florida) | 19–33 °C | ~0.05 c–0.3 (max. at 26 °C) | N/A | N/A | [59] |
| P. lima (PL100A) | 21–33 °C | ~0.066–~0.3 a (max. at 27 °C) | N/A | N/A | [60] |
| P. concavum (PC100A, Isolates from Knight Key, Florida) | 21–31 °C | ~0.05–~0.28 a (max. at 26 °C) | N/A | N/A | |
| P. hoffmannianum (882a, Isolates from Little Lameshur Bay, St. John, US Virgin Island) | 23–31 °C | - 2000 lux: ~0.06–~0.2 c (max. at 27 °C) - 3000 lux: ~0.1–~0.425 c (max. at 29 °C) - 4000 lux: ~0.07–~0.425 c (max. at 27 °C) - 5000 lux: ~0.08–~0.53 c (max. at 27 °C) | N/A | - OA at 2000 lux:~10–~53.75 c (max. at 23 °C) - OA at 3000 lux:~11.25–~38 c (max. at 29 °C) - OA at 4000 lux:~5–45 c (max. at 29 °C) - OA at 5000 lux: ~2.5 ~ 17.5 c (max. at 25 and 31 °C) | [21] |
| P. belizeanum (VGO1029, Isolates from La Puntilla, Las Palmas de Gran Canaria) | 18 °C | ~0.08–0.125 a (max. at 40 µE m−2 s−1) | ~135,000 c at 40 µE m−2 s−1 | OA: ~4.75 b,c | [24] |
| 25 °C | ~0.06–0.205 a (max. at 40 µE m−2 s−1) | ~120,000 c at 40 µE m−2 s−1 | OA: ~1.1 b,c | ||
| 28 °C | −0.075–0.125 a (max. at 40 µE m−2 s−1) | ~40,000 c at 40 µE m−2 s−1 | OA: ~2.625 b,c | ||
| P. lima (CCMP2579) | 15 °C | ~0.048 a | ~16,700 c | OA:~5.5 b,c, DTX-1: ~14.1 b,c | [22] |
| 20 °C | ~0.050 a | ~25,700 c | OA:~1.5 b,c, DTX-1: ~11 b,c | ||
| 25 °C | ~0.056 a | ~25,100 c | OA:~1 b,c, DTX-1: ~4.7 b,c | ||
| 30 °C | ~0.036 a | ~8200 c | OA:~12.731 b, DTX-1: ~16.587 b |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, T.C.-H.; Fong, F.L.-Y.; Ho, K.-C.; Lee, F.W.-F. The Mechanism of Diarrhetic Shellfish Poisoning Toxin Production in Prorocentrum spp.: Physiological and Molecular Perspectives. Toxins 2016, 8, 272. https://doi.org/10.3390/toxins8100272
Lee TC-H, Fong FL-Y, Ho K-C, Lee FW-F. The Mechanism of Diarrhetic Shellfish Poisoning Toxin Production in Prorocentrum spp.: Physiological and Molecular Perspectives. Toxins. 2016; 8(10):272. https://doi.org/10.3390/toxins8100272
Chicago/Turabian StyleLee, Thomas Chun-Hung, Fiona Long-Yan Fong, Kin-Chung Ho, and Fred Wang-Fat Lee. 2016. "The Mechanism of Diarrhetic Shellfish Poisoning Toxin Production in Prorocentrum spp.: Physiological and Molecular Perspectives" Toxins 8, no. 10: 272. https://doi.org/10.3390/toxins8100272
APA StyleLee, T. C.-H., Fong, F. L.-Y., Ho, K.-C., & Lee, F. W.-F. (2016). The Mechanism of Diarrhetic Shellfish Poisoning Toxin Production in Prorocentrum spp.: Physiological and Molecular Perspectives. Toxins, 8(10), 272. https://doi.org/10.3390/toxins8100272