
Isolation and Preliminary Characterization of Proteinaceous Toxins with Insecticidal and Antibacterial Activities from Black Widow Spider (L. tredecimguttatus) Eggs

Abstract
:1. Introduction
2. Results
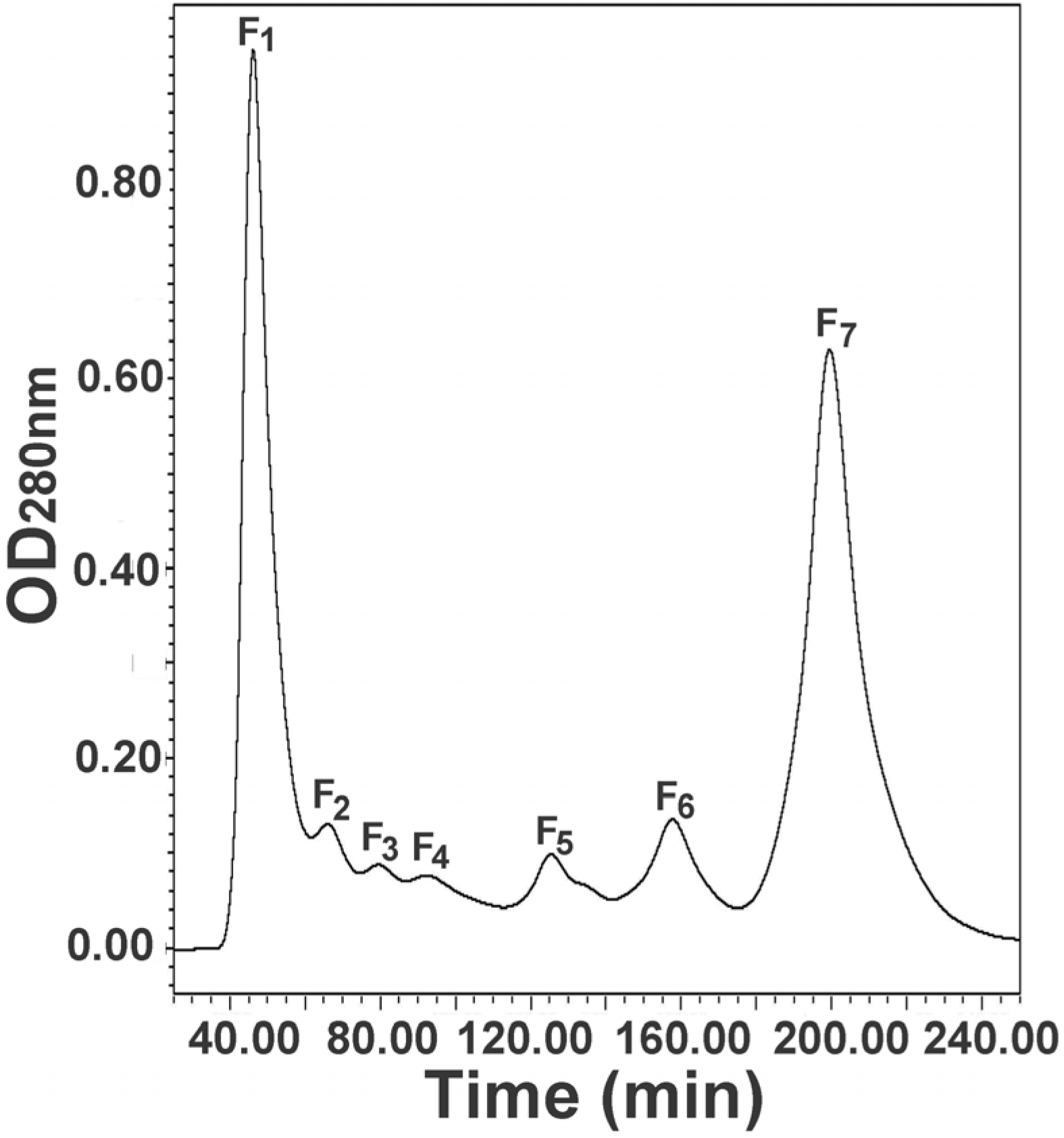
2.1. Molecular Sieve Chromatography of Egg Extract
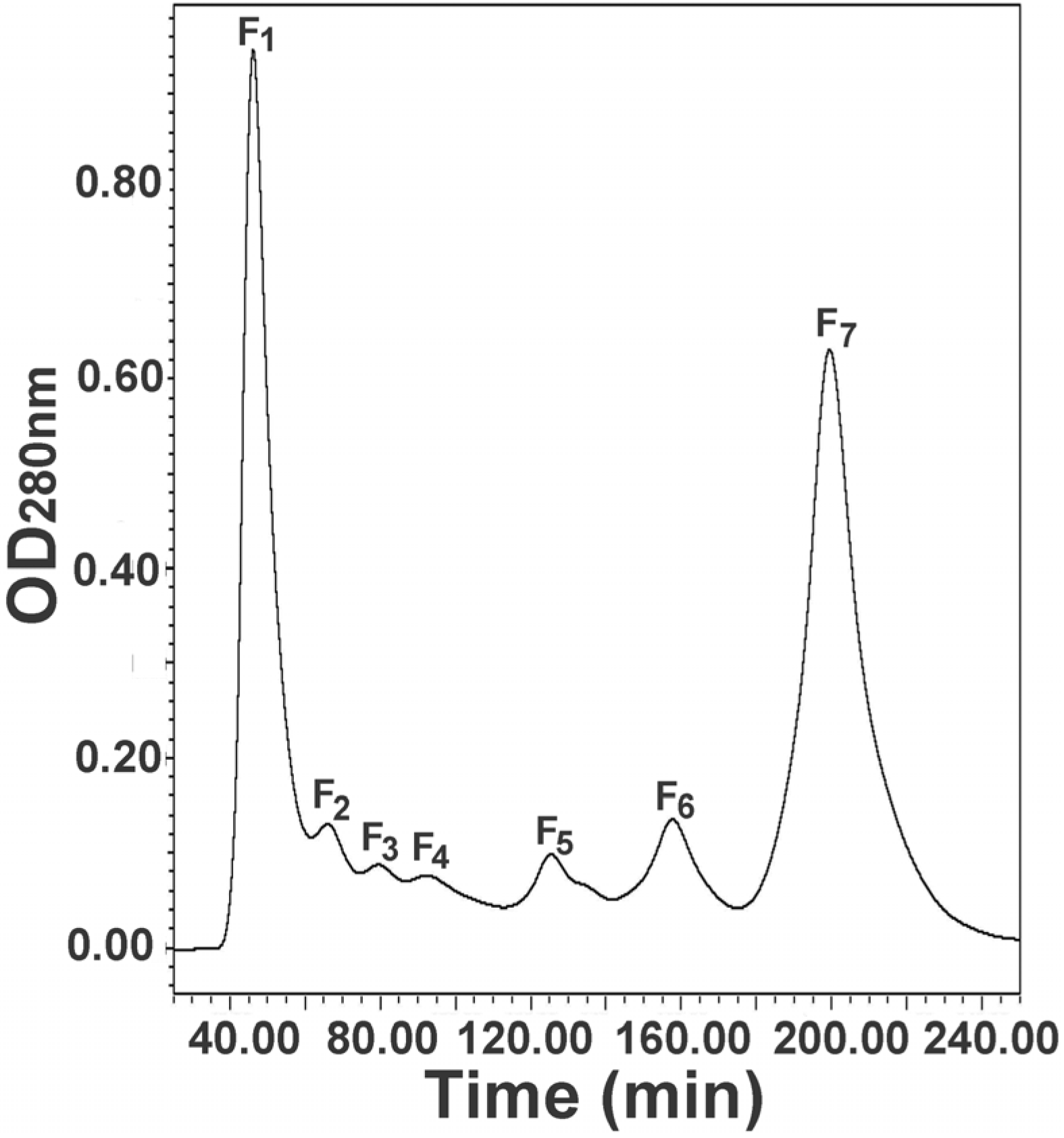
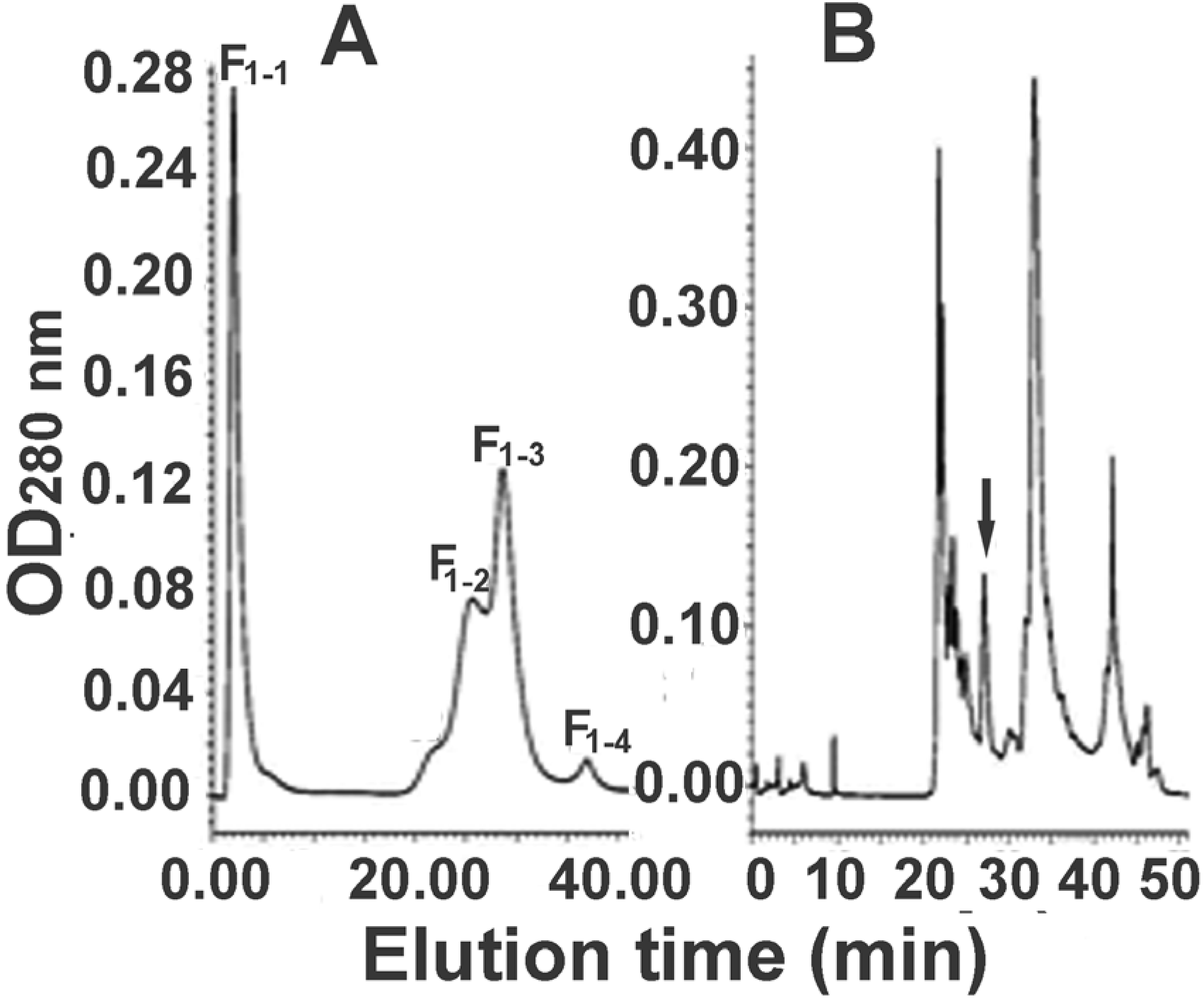
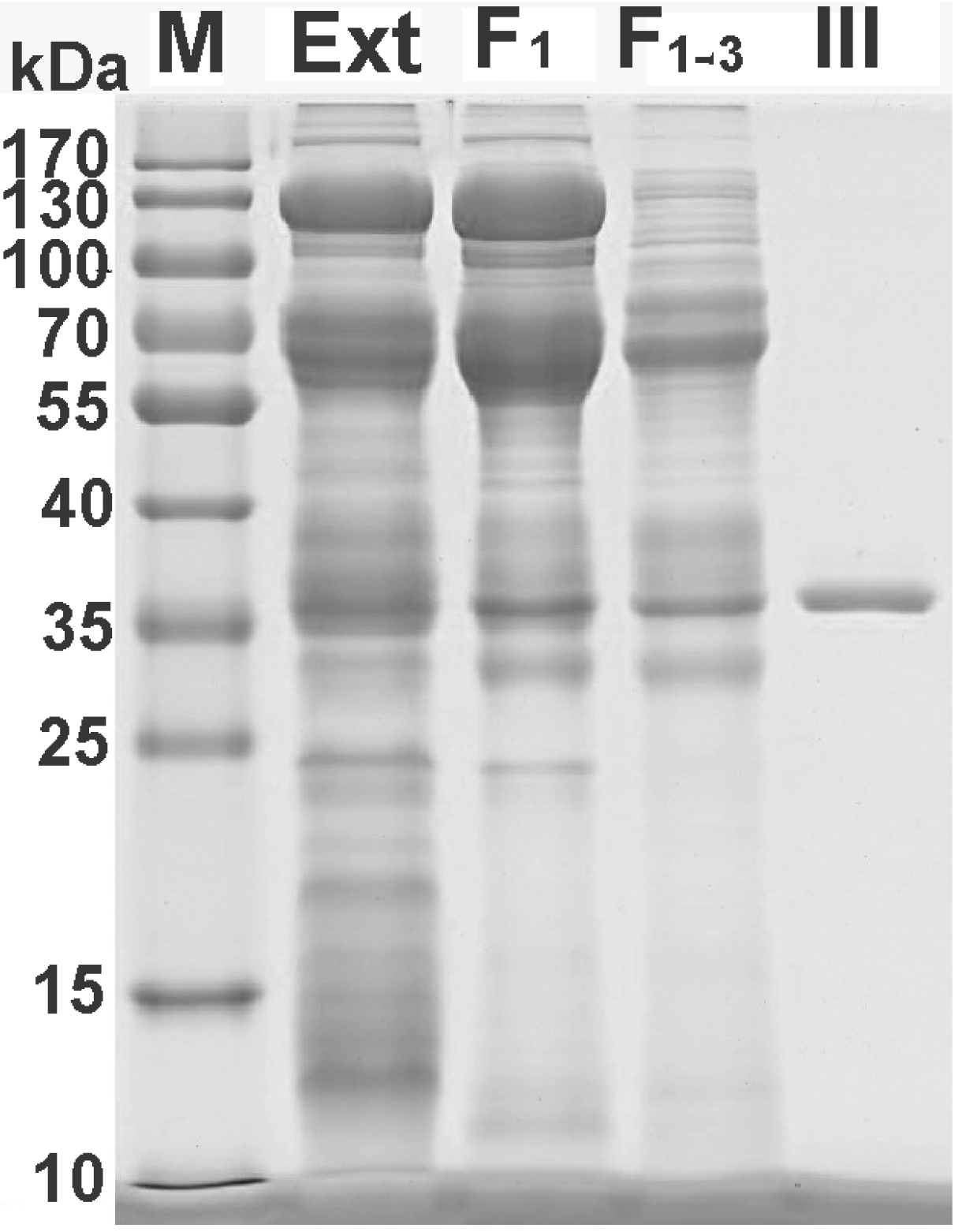
2.2. Further Separation of Protein Fraction F1
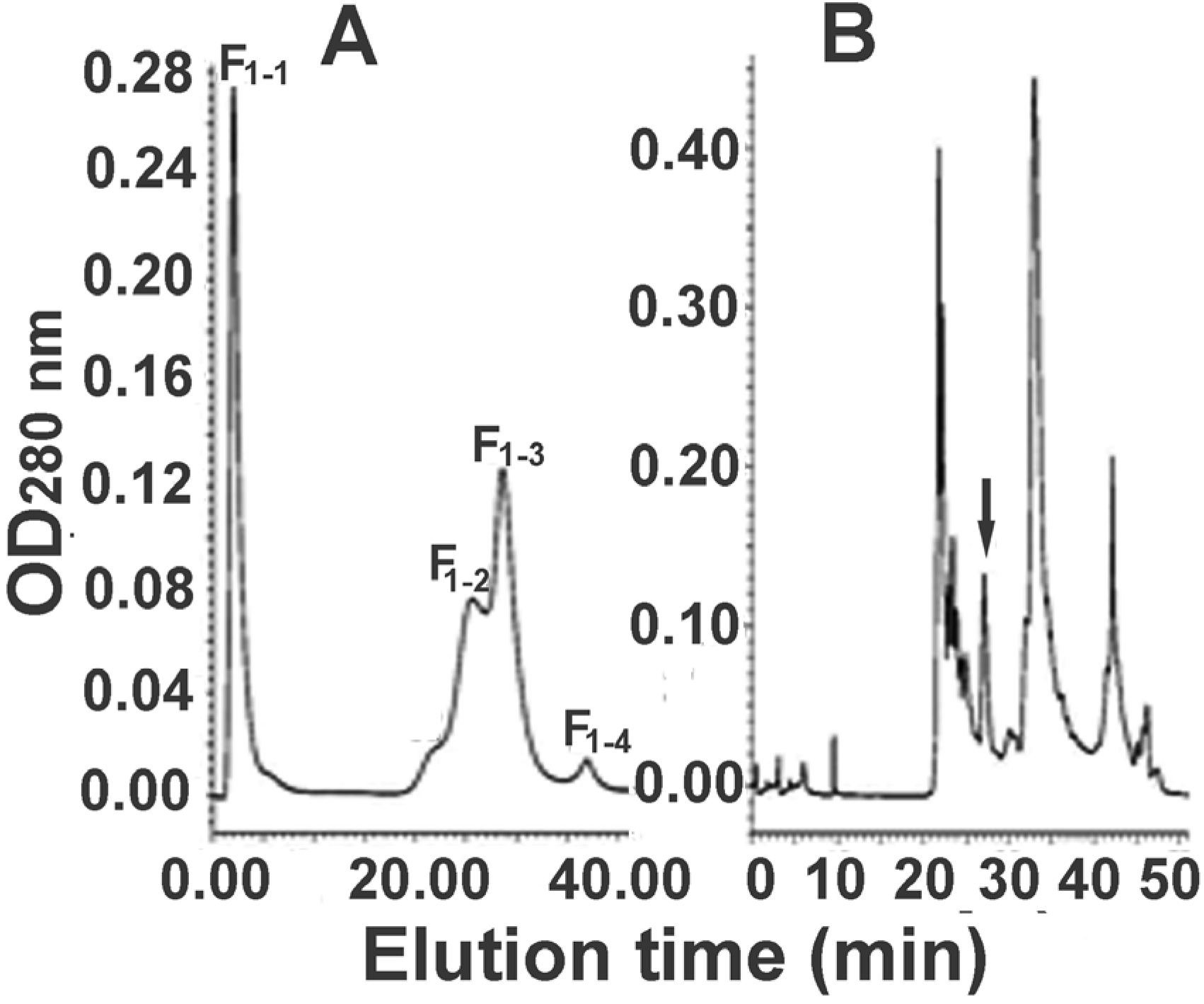

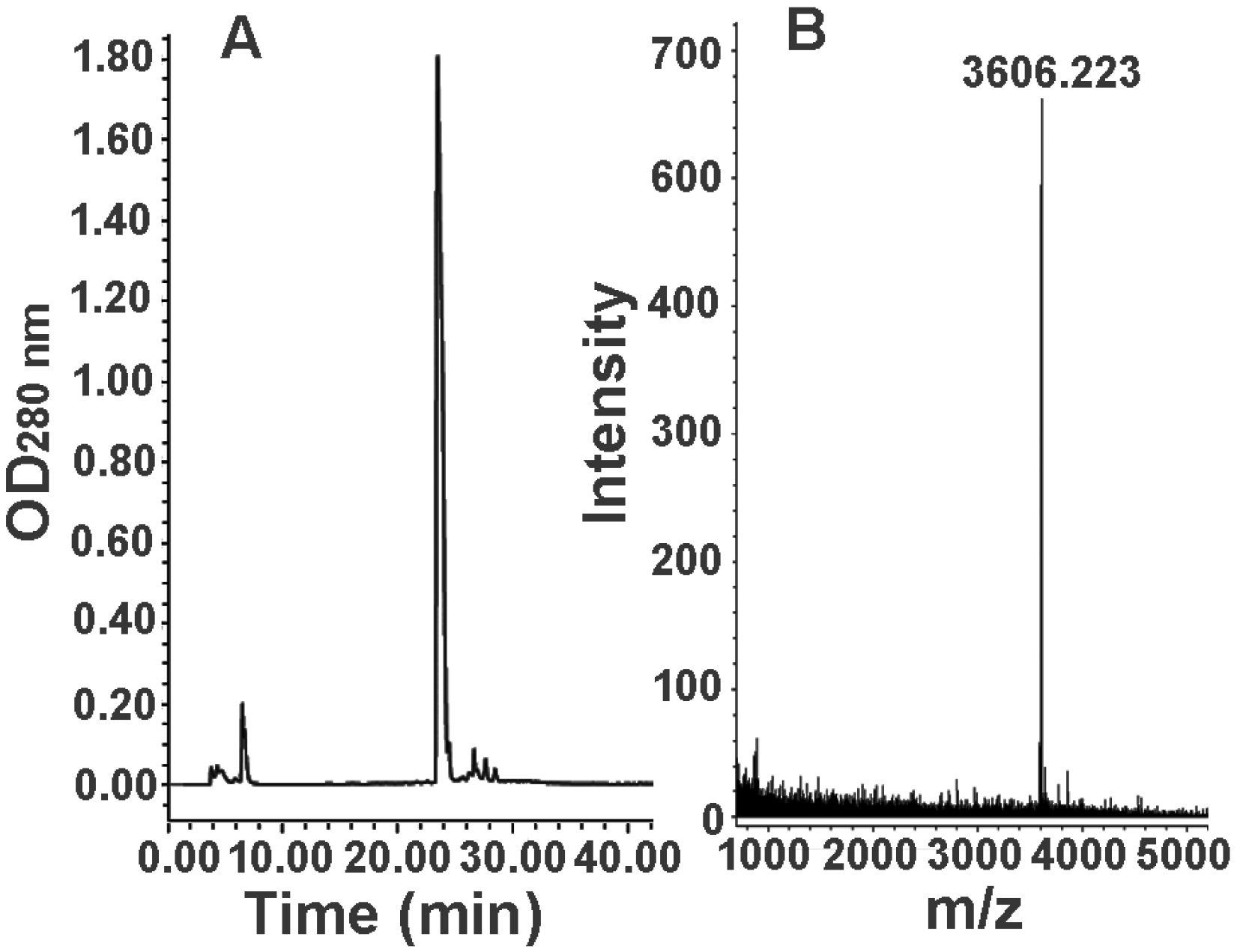
2.3. Further Separation of Peptide Fraction F7
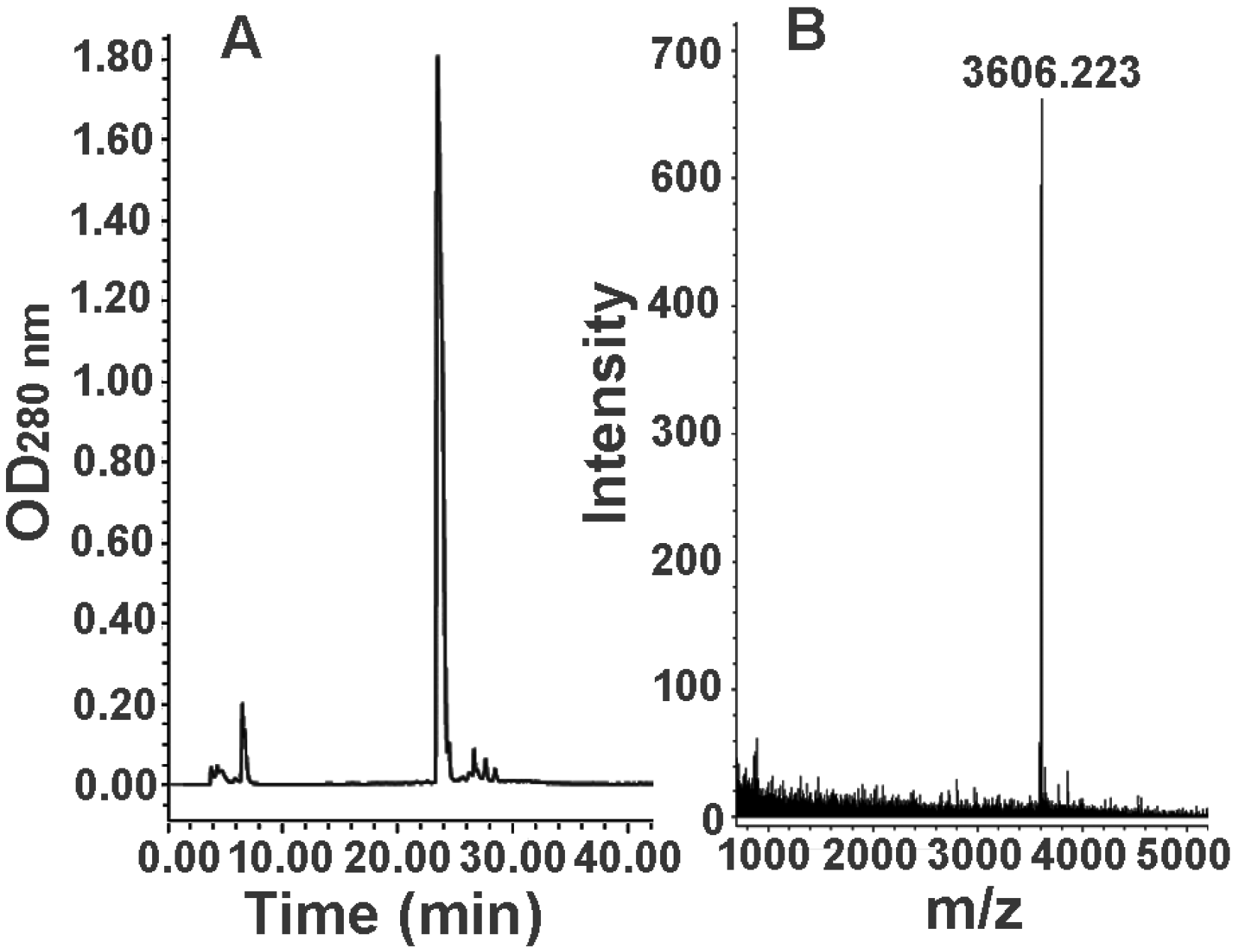
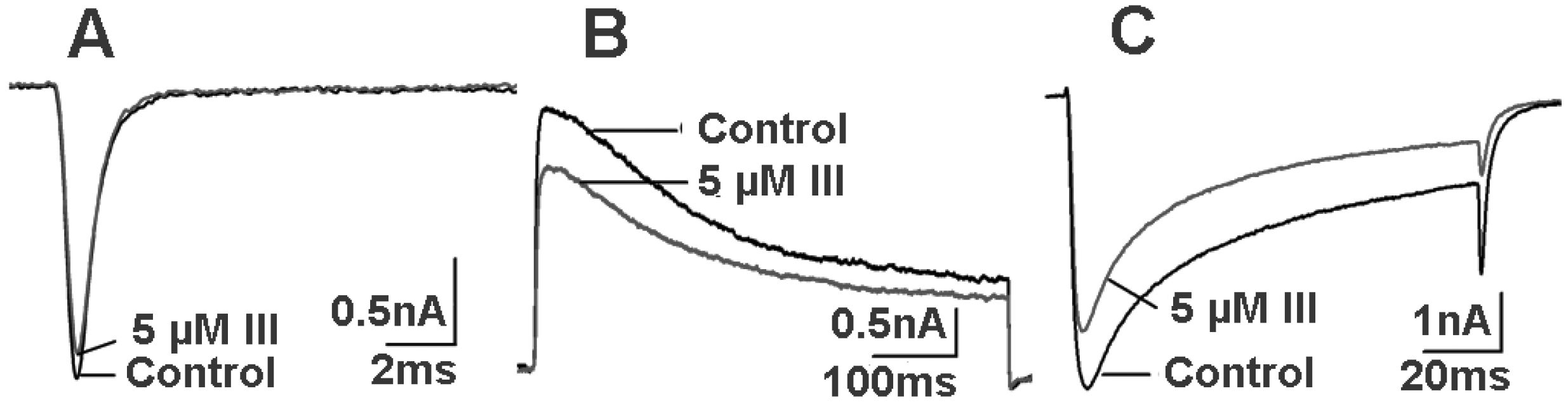
2.4. Bioactivity and N-terminal Sequence of Latroeggtoxin-III
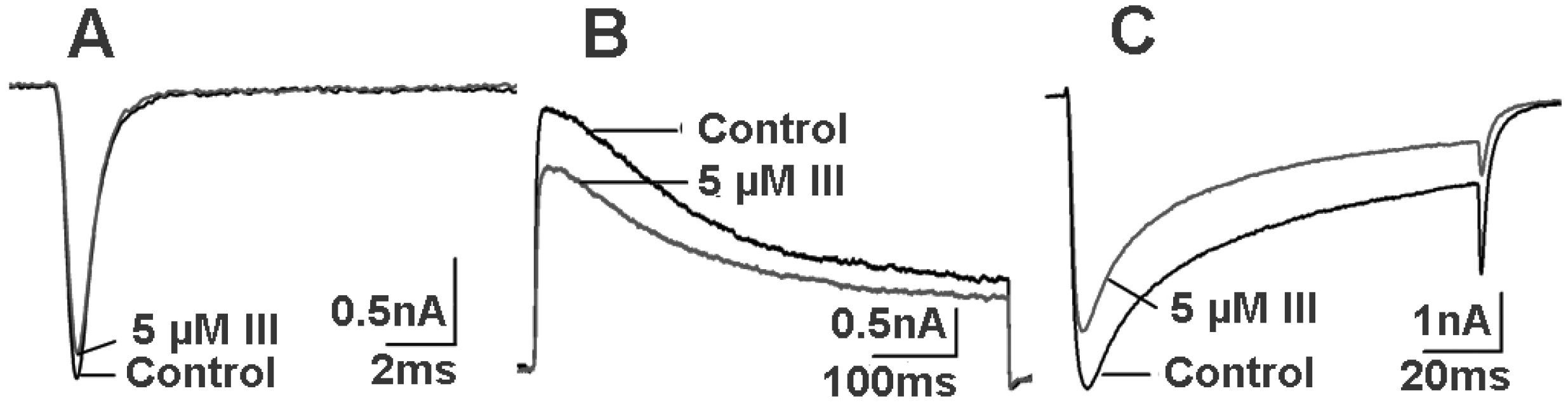
2.5. Bioactivity of Latroeggtoxin-IV

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | IZD b | IZD c |
|---|---|---|
| S. aureus | 14.73 ± 0.96 | 11.17 ± 0.41 |
| S. typhimurium | 11.53 ± 0.16 | 12.58 ± 0.41 |
| B. subtilis | 11.40 ± 0.89 | 13.28 ± 0.44 |
| E. coli | 10.82 ± 2.00 | 13.49 ± 1.57 |
| P. aeruginosa | 9.46 ± 0.19 | 12.67 ± 0.33 |
3. Discussion
4. Experimental Section
4.1. Materials
4.2. Preparation of Egg Extract
4.3. Fractionation of Egg Extract
4.4. Detection of Molecular Weight Distribution of the Fractions
4.5. Further Isolation of Protein Fraction F1 and Peptide Fraction F7
4.6. Animal Toxicity Detection
4.7. Antibacterial Activity Detection
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Escoubas, P.; Diochot, S.; Corzo, G. Structure and pharmacology of spider venom neurotoxins. Biochimie 2000, 82, 893–907. [Google Scholar] [CrossRef] [PubMed]
- Cestèle, S.; Catterall, W.A. Molecular mechanisms of neurotoxin action on voltage-gated sodium channels. Biochimie 2000, 82, 883–892. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.Y.; Yu, Q. Isolation and characterization of Hainantoxin-II, a new neurotoxic peptide from the Chinese bird spider (Haplopelma hainanum). Zool. Res. 2010, 31, 570–574. [Google Scholar] [PubMed]
- Akhunov, A.; Golubenko, Z.; Abdurashidova, N.A.; Mustakimova, E.Ch.; Ibragimov, F.A.; Mackessy, S. Comparative biochemistry of the physiologically active components of venom, hemolymphy, and eggs of the karakurt spider (Latrodectus tredecimguttatus). Chem. Nat. Compd. 2001, 37, 562–565. [Google Scholar] [CrossRef]
- Buffkin, D.C.; Russell, F.E. A poison from the eggs and spiderlings of the black widow spider. Proc. West. Pharmac. Soc. 1971, 14, 166. [Google Scholar]
- Kobert, R. Ueber die giftigen Spinnen Russlads. Sber. Naturf. Ges. Dorpot. 1889, 8, 340–362. [Google Scholar]
- Peng, X.; Zhang, Y.; Liu, J.; Yu, H.; Chen, J.; Lei, Q.; Wang, X.; Liang, S. Physiological and biochemical analysis to reveal the molecular basis for black widow spiderling toxicity. J. Biochem. Mol. Toxicol. 2014, 28, 198–205. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.Z.; Li, J.J.; Zhang, Y.Y.; Peng, X.Z.; Guo, T.Y.; Wang, J.R.; Hu, W.J.; Duan, Z.G.; Wang, X.C. Physiological and biochemical characterization of egg extract of black widow spiders to uncover molecular basis of egg toxicity. Biol. Res. 2014, 47, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liu, H.; Duan, Z.; Cao, R.; Wang, X.; Liang, S. Protein compositional analysis of the eggs of black widow spider (L. tredecimguttatus): Implications for the understanding of egg toxicity. J. Biochem. Mol. Toxicol. 2012, 26, 510–515. [Google Scholar] [CrossRef] [PubMed]
- Li, J.J.; Yan, Y.Z.; Wang, J.J.; Guo, T.Y.; Hu, W.J.; Duan, Z.G.; Wang, X.C.; Liang, S.P. Purification and partial characterization of a novel neurotoxic Protein from eggs of black widow spiders (Latrodectus tredecimguttatus). J. Biochem. Mol. Toxicol. 2013, 27, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Yan, Y.; Yu, H.; Peng, X.; Zhang, Y.; Hu, W.; Duan, Z.; Wang, X.; Liang, S. Isolation and identification of a sodium channel-inhibiting protein from eggs of black widow spiders. Int. J. Biol. Macromol. 2014, 65, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Seehuus, S.C.; Norberg, K.; Gimsa, U.; Krekling, T.; Amdam, G.V. Reproductive protein protects functionally sterile honey bee workers from oxidative stress. Proc. Natl. Acad. Sci. USA 2006, 103, 962–967. [Google Scholar] [CrossRef] [PubMed]
- Sattar Khan, M.A.; Nakamura, S.; Ogawa, M.; Akita, E.; Azakami, H.; Kato, A. Bactericidal action of egg yolk phosvitin against Escherichia coli under thermal stress. J. Agric. Food. Chem. 2000, 48, 1503–1506. [Google Scholar] [CrossRef] [PubMed]
- Dreon, M.S.; Frassa, M.V.; Ceolín, M.; Ituarte, S.; Qiu, J.W.; Sun, J.; Fernández, P.E.; Heras, H. Novel animal defenses against predation: A snail egg neurotoxin combining lectin and pore-forming chains that resembles plant defense and bacteria attack toxins. PLoS One 2013, 8, e63782. [Google Scholar] [CrossRef] [PubMed]
- Corzo, G.; Escoubas, P. Pharmacologically active spider peptide toxins. Cell. Mol. Life Sci. 2003, 60, 2409–2426. [Google Scholar] [CrossRef] [PubMed]
- Tedford, H.W.; Sollod, B.L.; Maggio, F.; King, G.F. Australian funnel-web spiders: Master insecticide chemists. Toxicon 2004, 43, 601–618. [Google Scholar] [CrossRef] [PubMed]
- Ushkaryov, Y.A.; Volynski, K.E.; Ashton, A.C. The multiple actions of black widow spider toxins and their selective use in neurosecretion studies. Toxicon 2004, 43, 527–542. [Google Scholar] [CrossRef] [PubMed]
- Rohou, A.; Nield, J.; Ushkaryov, Y.A. Insecticidal toxins from black widow spider venom. Toxicon 2007, 49, 531–549. [Google Scholar] [CrossRef] [PubMed]
- Kiyatkin, N.I.; Kulikovskaya, I.M.; Grishin, E.V.; Beadle, D.J.; King, L.A. Functional characterization of black widow spider neurotoxins synthesized in insect cells. Eur. J. Biochem. 1995, 230, 854–859. [Google Scholar] [CrossRef] [PubMed]
- Russell, F.E.; Maretć, Z. Effects of Latrodectus egg poison on web building. Toxicon 1979, 17, 649–650. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Adams, M.E. Lycotoxins, antimicrobial peptides from venom of the wolf spider Lycosa carolinensis. J. Biol. Chem. 1998, 273, 2059–2066. [Google Scholar] [CrossRef] [PubMed]
- Tan, H.; Ding, X.; Meng, S.; Liu, C.; Wang, H.; Xia, L.; Liu, Z.; Liang, S. Antimicrobial potential of lycosin-I, a cationic and amphiphilic peptide from the venom of the spider Lycosa singorensis. Curr. Mol. Med. 2013, 13, 900–910. [Google Scholar] [CrossRef] [PubMed]
- Kuhn-Nentwig, L.; Muller, J.; Schaller, J.; Walz, A.; Dathe, M.; Nentwig, W. Cupiennin 1, a new family of highly basic antimicrobial peptides in the venom of the spider Cupiennius salei (Ctenidae). J. Biol. Chem. 2002, 277, 11208–11216. [Google Scholar] [CrossRef] [PubMed]
- Shulov, A.; Weissman, A. Notes on the life habits and potency of the venom of the three Latrodectus spider species of Israël. Ecology 1959, 40, 515–518. [Google Scholar] [CrossRef]
- LaemmLi, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.A.; Xie, X.J.; Li, M.; Yang, D.M.; Xie, Z.P.; Zong, X.; Liang, S.P. Blockade of neuromuscular transmission by huwentoxin-I, purified from the venom of the Chinese bird spider Selenocosmia huwena. Toxicon 1997, 35, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.Z.; Li, Z.W. Modulation by adenosine of GABA-activated current in rat dorsal root ganglion neurons. J. Physiol. 1997, 501, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Deng, M.C.; Kuang, F.; Sun, Z.H.; Tao, H.; Cai, T.F.; Zhong, L.L.; Chen, Z.R.; Xiao, Y.C.; Liang, S.P. Jingzhaotoxin-IX, a novel gating modifier of both sodium and potassium channels from Chinese tarantula Chilobrachys jingzhao. Neuropharmacology 2009, 57, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Gunning, S.J.; Maggio, F.; Windley, M.J.; Valenzuela, S.M.; King, G.F.; Nicholson, G.M. The Janus-faced atracotoxins are specific blockers of invertebrate KCa channels. FEBS J. 2008, 275, 4045–4059. [Google Scholar] [CrossRef] [PubMed]
- Windley, M.J.; Escoubas, P.; Valenzuela, S.M.; Nicholson, G.M. A novel family of insect-selective peptide neurotoxins targeting insect large-conductance calcium-activated K+ channels isolated from the venom of the theraphosid spider Eucratoscelus constrictus. Mol. Pharmacol. 2011, 80, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Rong, M.; Xiao, Y.; Liang, S. The effects of huwentoxin-I on the voltage-gated sodium channels of rat hippocampal and cockroach dorsal unpaired median neurons. Peptides 2012, 34, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Zarai, Z.; Kadri, A.; Ben Chobba, I.; Ben Mansour, R.; Bekir, A.; Mejdoub, H.; Gharsallah, N. The in-vitro evaluation of antibacterial, antifungal and cytotoxic properties of Marrubium vulgare L. essential oil grown in Tunisia. Lipids Health Dis. 2011, 10, 161–168. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lei, Q.; Yu, H.; Peng, X.; Yan, S.; Wang, J.; Yan, Y.; Wang, X. Isolation and Preliminary Characterization of Proteinaceous Toxins with Insecticidal and Antibacterial Activities from Black Widow Spider (L. tredecimguttatus) Eggs. Toxins 2015, 7, 886-899. https://doi.org/10.3390/toxins7030886
Lei Q, Yu H, Peng X, Yan S, Wang J, Yan Y, Wang X. Isolation and Preliminary Characterization of Proteinaceous Toxins with Insecticidal and Antibacterial Activities from Black Widow Spider (L. tredecimguttatus) Eggs. Toxins. 2015; 7(3):886-899. https://doi.org/10.3390/toxins7030886
Chicago/Turabian StyleLei, Qian, Hai Yu, Xiaozhen Peng, Shuai Yan, Jirong Wang, Yizhong Yan, and Xianchun Wang. 2015. "Isolation and Preliminary Characterization of Proteinaceous Toxins with Insecticidal and Antibacterial Activities from Black Widow Spider (L. tredecimguttatus) Eggs" Toxins 7, no. 3: 886-899. https://doi.org/10.3390/toxins7030886
APA StyleLei, Q., Yu, H., Peng, X., Yan, S., Wang, J., Yan, Y., & Wang, X. (2015). Isolation and Preliminary Characterization of Proteinaceous Toxins with Insecticidal and Antibacterial Activities from Black Widow Spider (L. tredecimguttatus) Eggs. Toxins, 7(3), 886-899. https://doi.org/10.3390/toxins7030886