
Vitamin D3 and Monomethyl Fumarate Enhance Natural Killer Cell Lysis of Dendritic Cells and Ameliorate the Clinical Score in Mice Suffering from Experimental Autoimmune Encephalomyelitis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
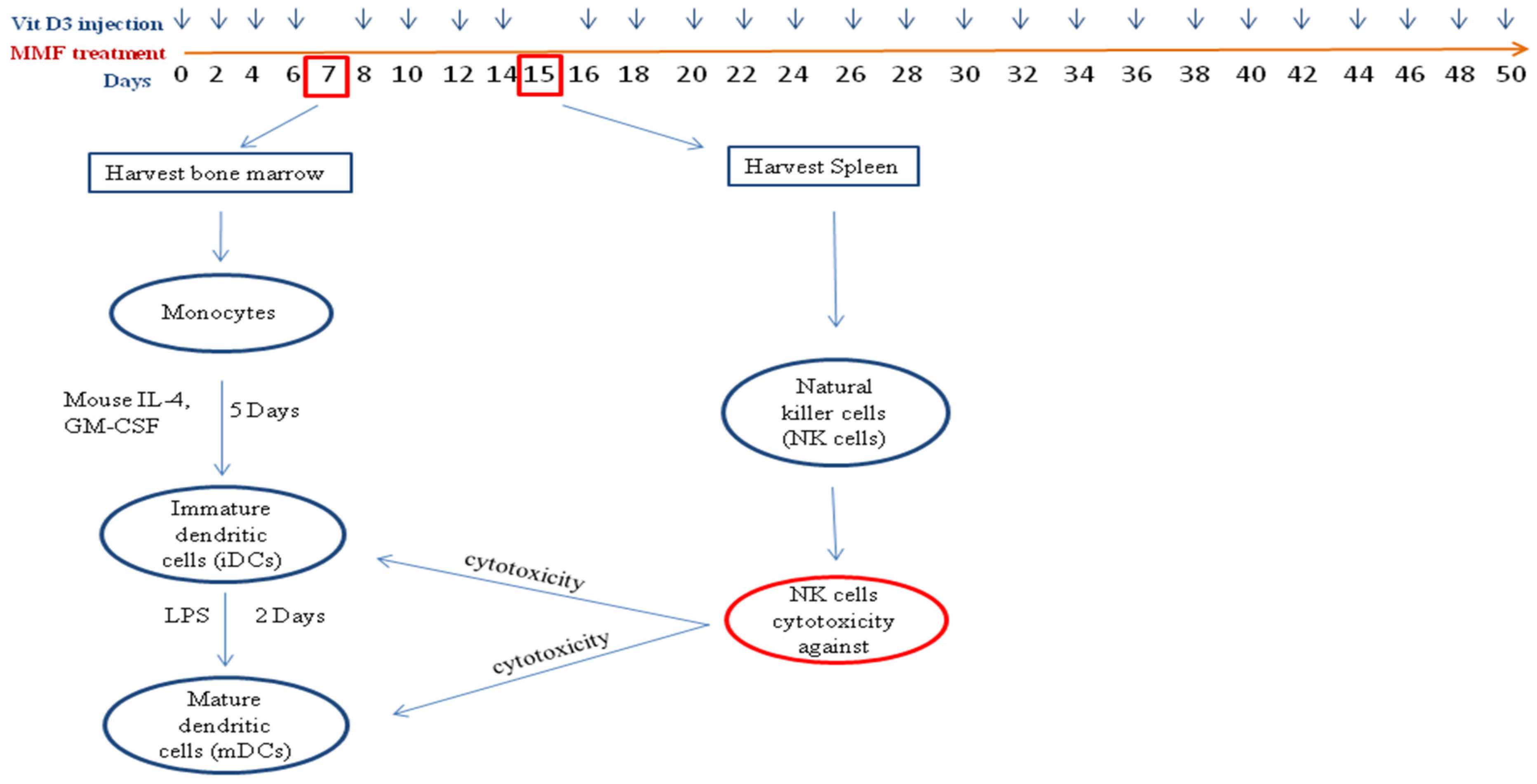
2. Results
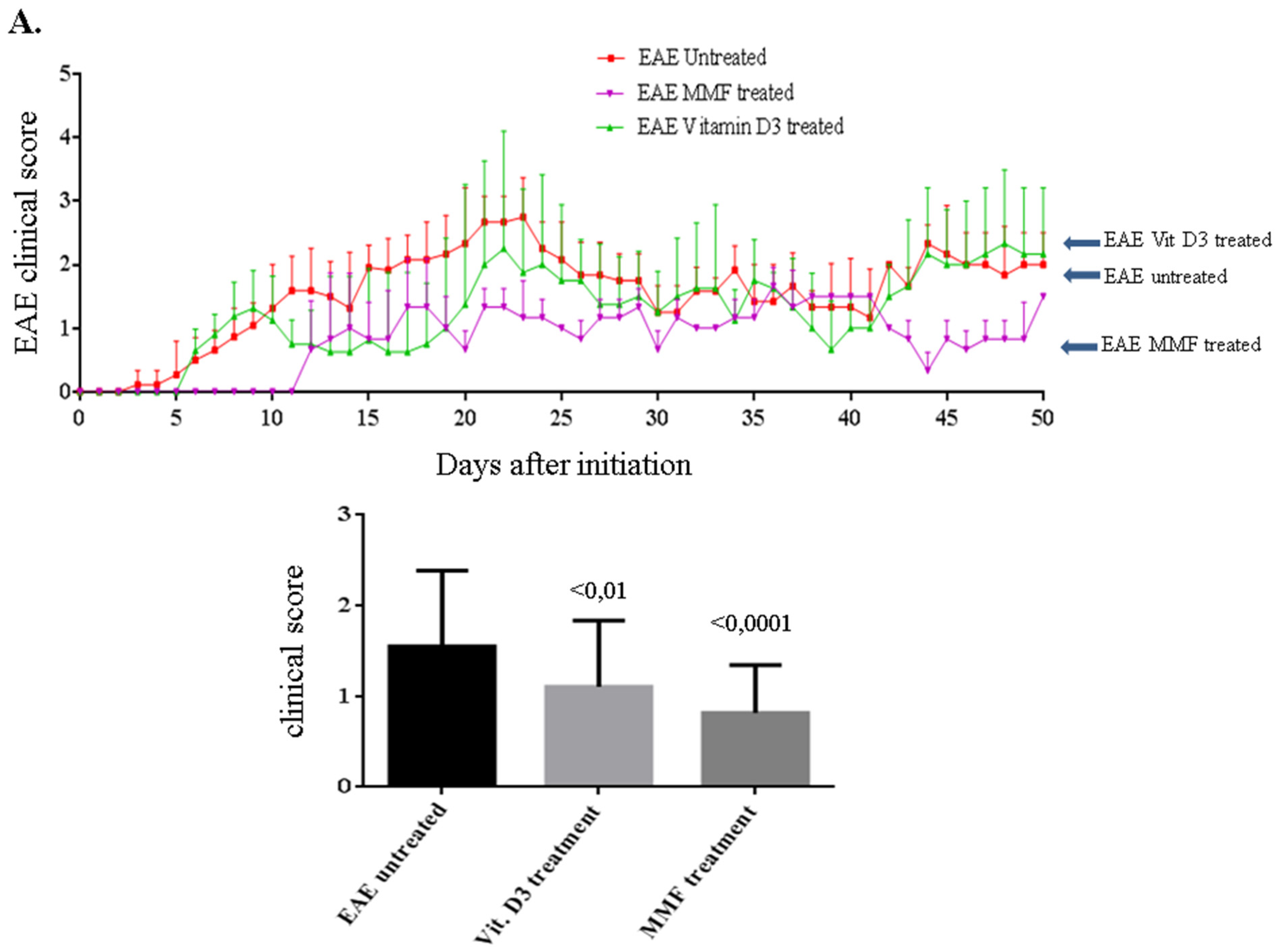
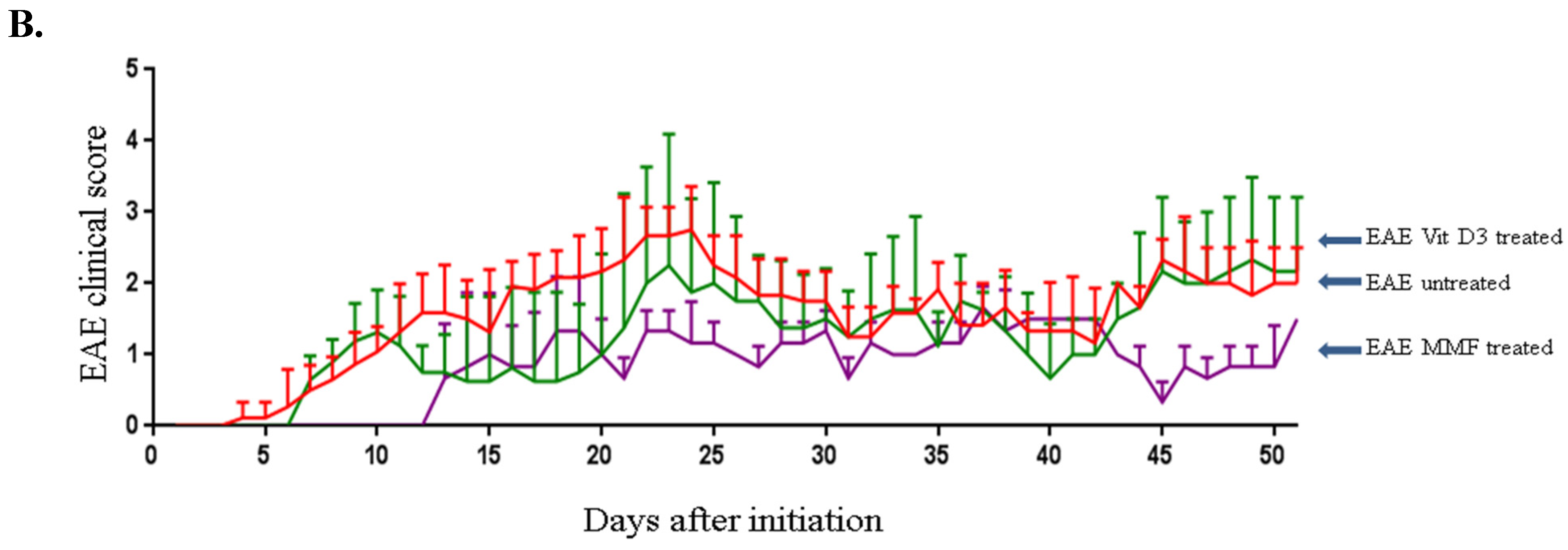
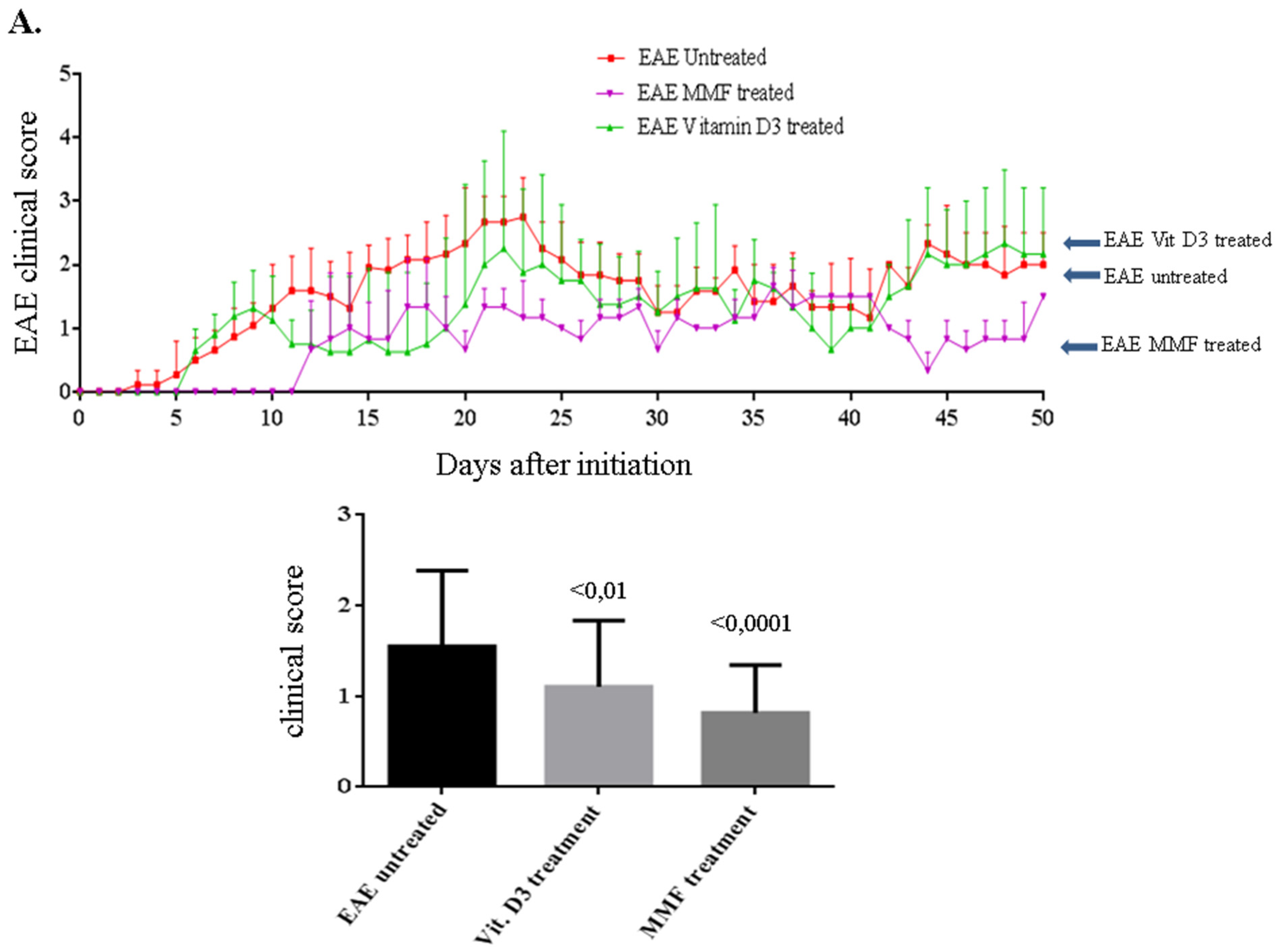
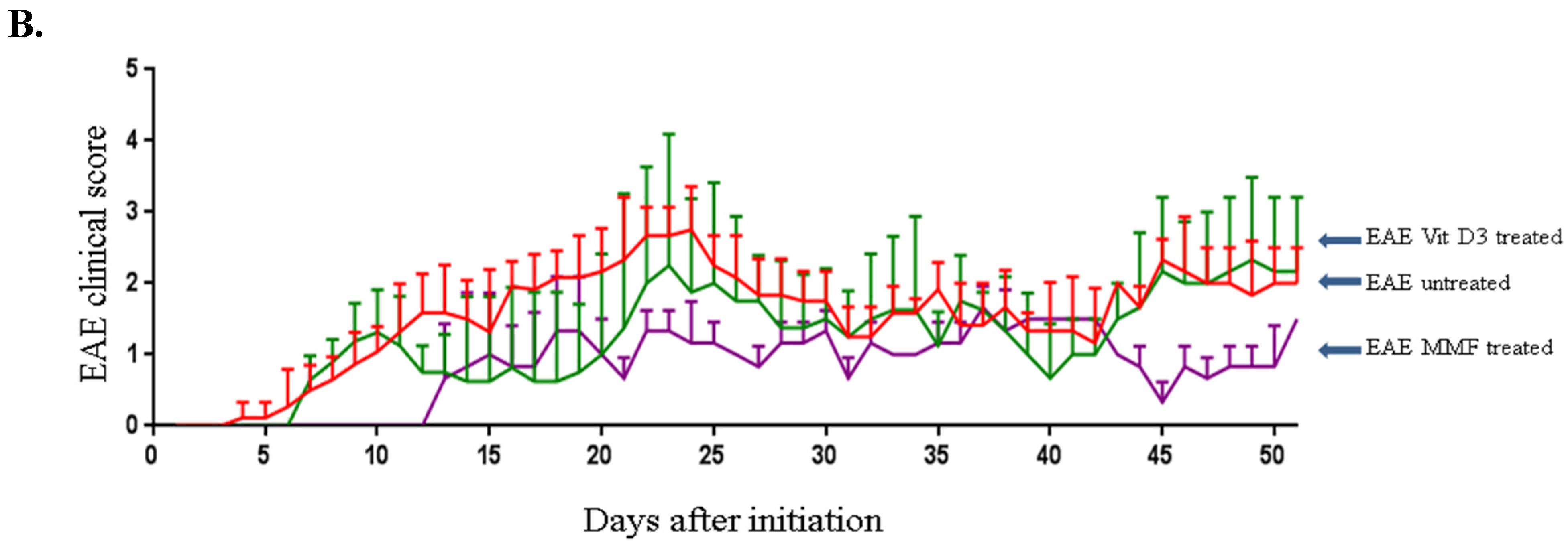
2.1. Vitamin D3 or MMF Reduces the EAE Clinical Score
2.2. Vitamin D3 Induces NK Cell Lysis of Dendritic Cells

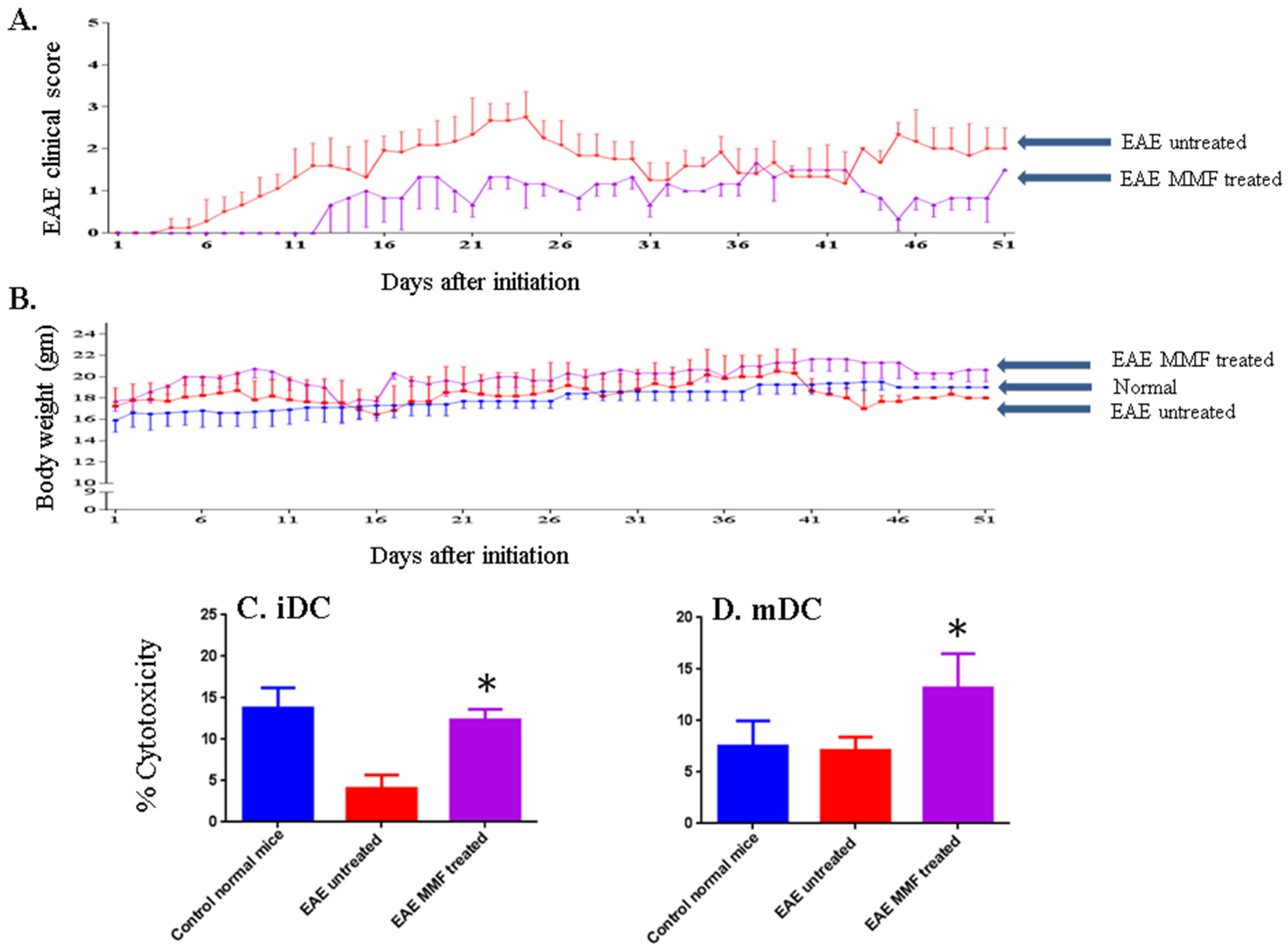
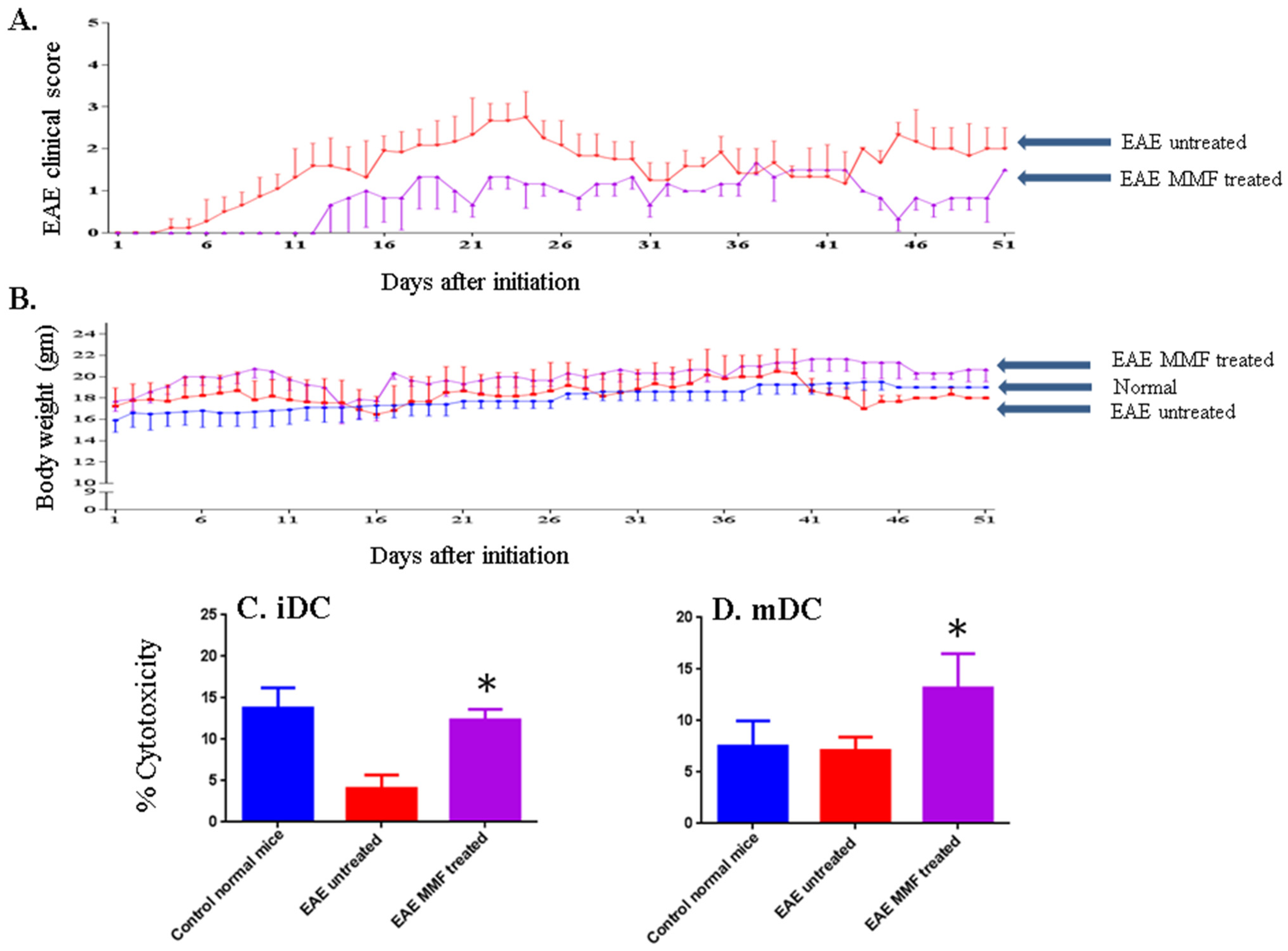
2.3. MMF Reduces the EAE Clinical Score Corroborated with Enhancing NK Cell Activity
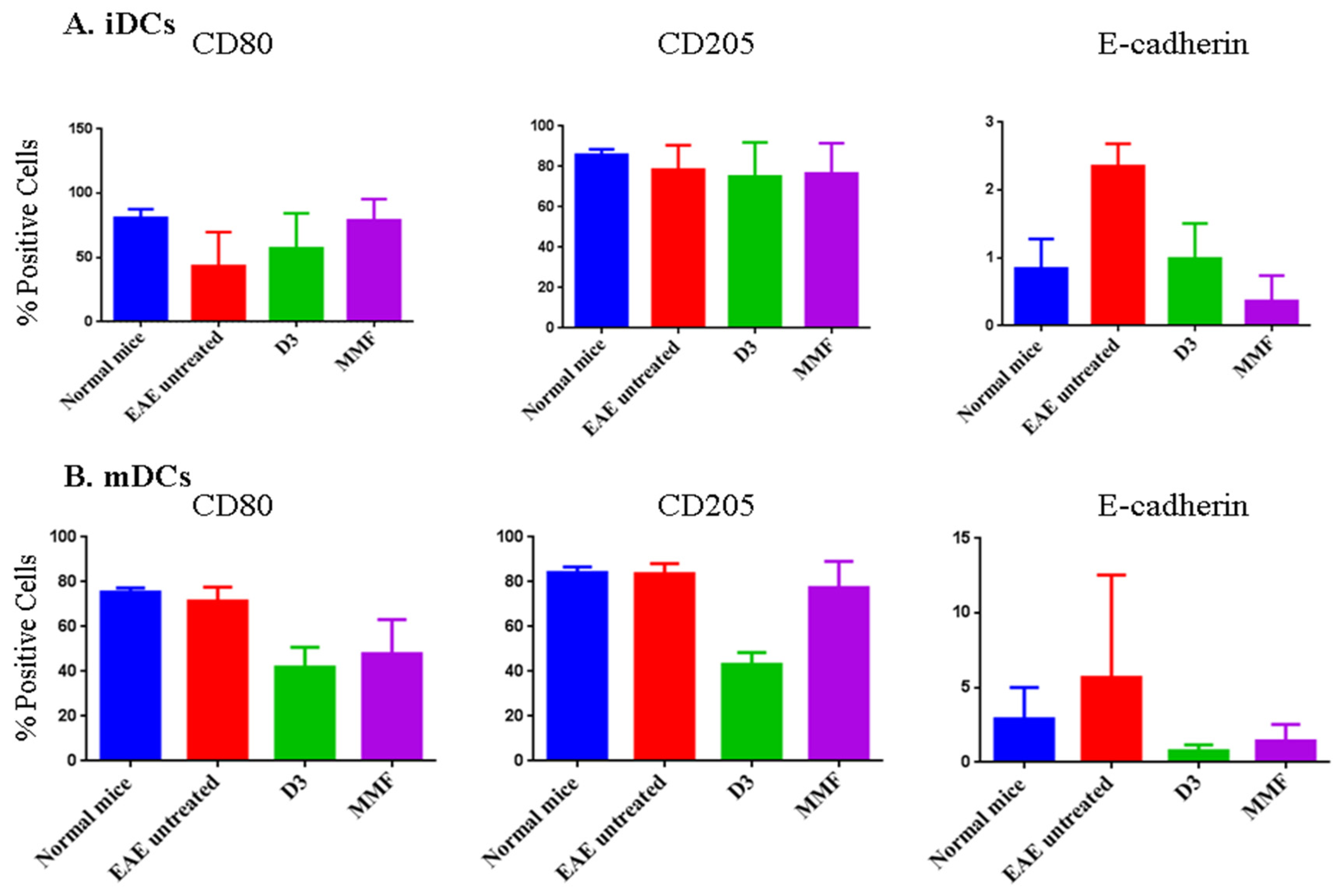
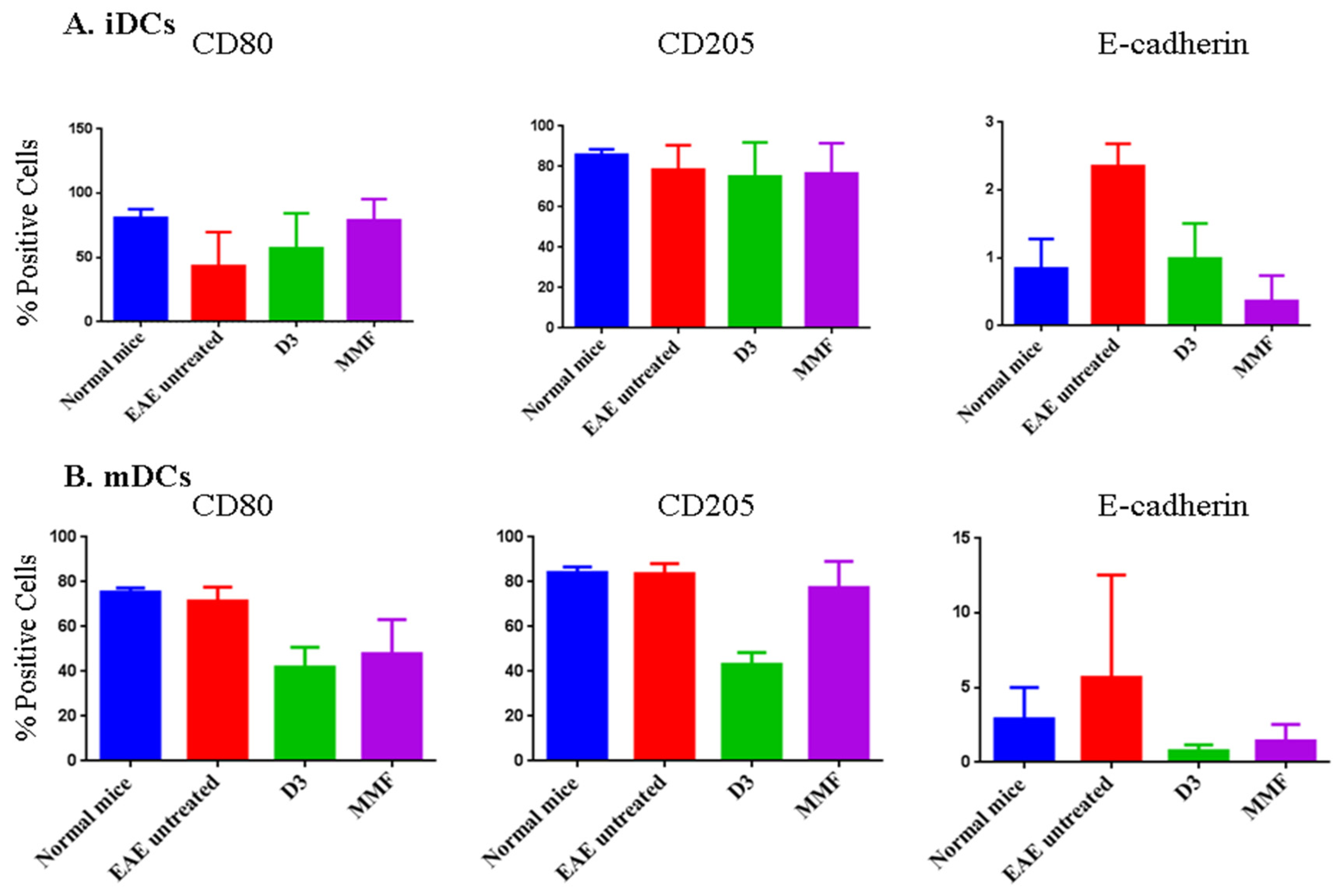
2.4. Effects of Vitamin D3 and MMF on Dendritic Cell Phenotypic Expression
3. Discussion
4. Experimental Sections
4.1. Ethical Statement
4.2. Induction of EAE in SJL Mice
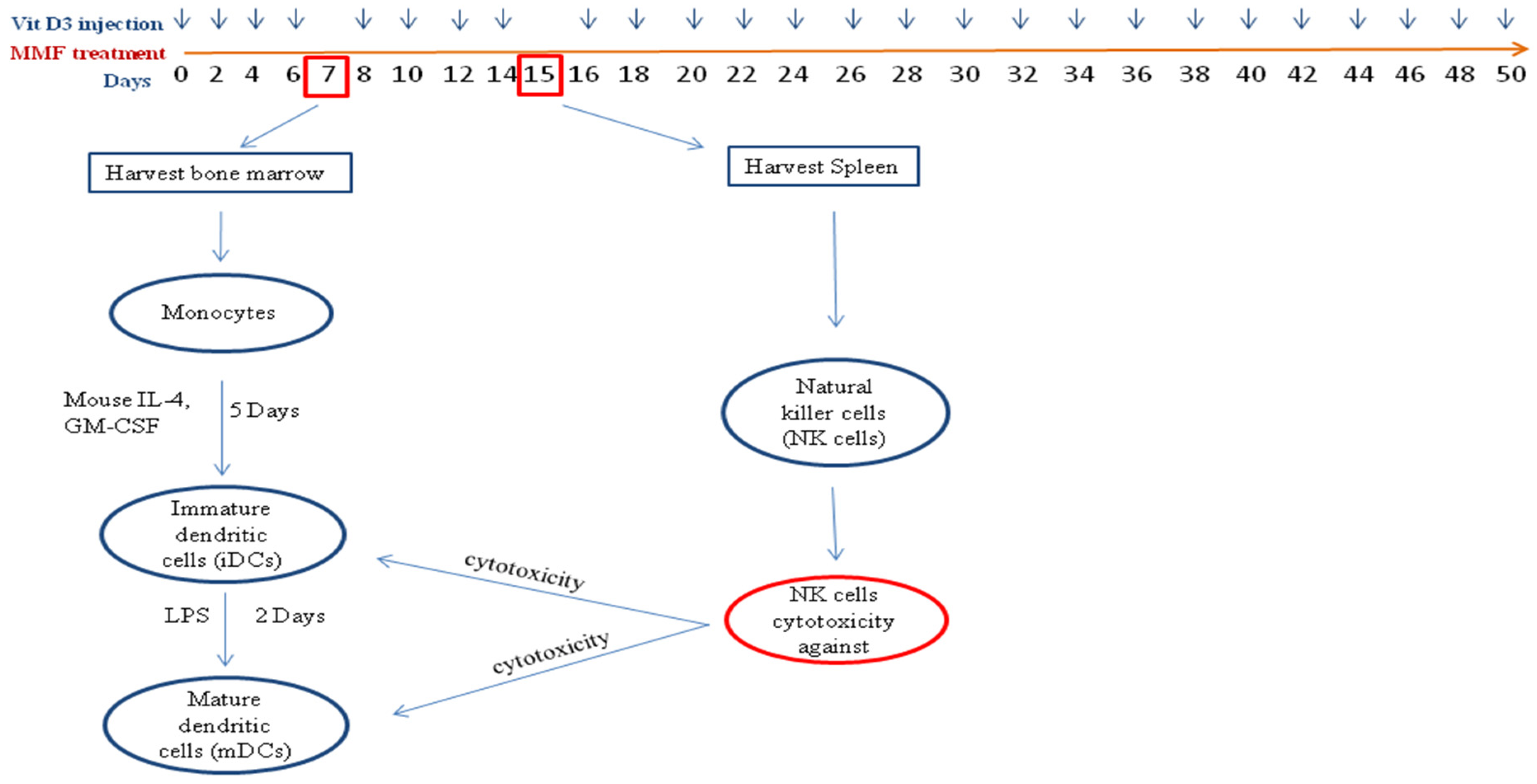
4.3. Treatment of Mice and Isolation of NK Cells and DCs
4.4. NK Cells Cytotoxicity Assay
4.5. Flow Cytometry Analysis (FACS)
4.6. Statistical Analysis
5. Conclusions
Supplementary Files
Supplementary File 1Author Contributions
Conflicts of Interest
References
- Cooper, M.A.; Fehniger, T.A.; Caligiuri, M.A. The biology of human natural killer-cell subsets. Trends Immunol. 2001, 22, 633–640. [Google Scholar] [CrossRef]
- Moretta, A. Natural killer cells and dendritic cells: Rendezvous in abused tissues. Nat. Rev. Immunol. 2002, 2, 957–964. [Google Scholar] [CrossRef] [PubMed]
- Biron, C.A.; Nguyen, K.B.; Pien, G.C.; Cousens, L.P.; Salazar-Mather, T.P. Natural killer cells in antiviral defense: Function and regulation by innate cytokines. Annu. Rev. Immunol. 1999, 17, 189–220. [Google Scholar] [CrossRef] [PubMed]
- Maghazachi, A.A. Compartmentalization of human natural killer cells. Mol. Immunol. 2005, 42, 523–529. [Google Scholar] [CrossRef] [PubMed]
- Maghazachi, A.A. Insights into seven and single transmembrane-spanning domain receptors and their signaling pathways in human natural killer cells. Pharmacol. Rev. 2005, 57, 339–357. [Google Scholar] [CrossRef] [PubMed]
- Lanier, L.L.; Testi, R.; Bindl, J.; Phillips, J.H. Identity of Leu-19 (CD56) leukocyte differentiation antigen and neural cell adhesion molecule. J. Exp. Med. 1989, 169, 2233–2238. [Google Scholar] [CrossRef] [PubMed]
- Walzer, T.; Jaeger, S.; Chaix, J.; Vivier, E. Natural killer cells: From CD3(-)NKp46(+) to post-genomics meta-analyses. Curr. Opin. Immunol. 2007, 19, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Glimcher, L.; Shen, F.W.; Cantor, H. Identification of a cell-surface antigen selectively expressed on the natural killer cell. J. Exp. Med. 1977, 145, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Barnett, M.H.; Sutton, I. The pathology of multiple sclerosis: A paradigm shift. Curr. Opin. Neurol. 2006, 19, 242–247. [Google Scholar] [CrossRef] [PubMed]
- Maghazachi, A.A. On the role of natural killer cells in neurodegenerative diseases. Toxins 2013, 5, 363–375. [Google Scholar] [CrossRef] [PubMed]
- Sand, K.L.; Knudsen, E.; Rolin, J.; Al-Falahi, Y.; Maghazachi, A.A. Modulation of natural killer cell cytotoxicity and cytokine release by the drug glatiramer acetate. Cell. Mol. Life Sci. 2009, 66, 1446–1456. [Google Scholar] [CrossRef] [PubMed]
- Al-Falahi, Y.; Sand, K.L.; Knudsen, E.; Damaj, B.B.; Rolin, J.; Maghazachi, A.A. Splenic natural killer cell activity in two models of experimental neurodegenerative diseases. J. Cell. Mol. Med. 2009, 13, 2693–2703. [Google Scholar] [CrossRef] [PubMed]
- Hoglund, R.A.; Holmoy, T.; Harbo, H.F.; Maghazachi, A.A. A one year follow-up study of natural killer and dendritic cells activities in multiple sclerosis patients receiving glatiramer acetate (GA). PLoS ONE 2013, 8, e62237. [Google Scholar] [CrossRef] [PubMed]
- Ascherio, A.; Munger, K.L. Environmental risk factors for multiple sclerosis. Part II: Noninfectious factors. Ann. Neurol. 2007, 61, 504–513. [Google Scholar] [CrossRef] [PubMed]
- Ascherio, A.; Munger, K.L.; Simon, K.C. Vitamin D and multiple sclerosis. Lancet Neurol. 2010, 9, 599–612. [Google Scholar] [CrossRef]
- Brustad, M.; Sandanger, T.; Aksnes, L.; Lund, E. Vitamin D status in a rural population of northern Norway with high fish liver consumption. Public Health Nutr. 2004, 7, 783–789. [Google Scholar] [CrossRef] [PubMed]
- Lemire, J.M.; Archer, D.C. 1,25-Dihydroxyvitamin D3 prevents the in vivo induction of murine experimental autoimmune encephalomyelitis. J. Clin. Investig. 1991, 87, 1103–1107. [Google Scholar] [CrossRef] [PubMed]
- Cantorna, M.T.; Hayes, C.E.; Deluca, H.F. 1,25-Dihydroxyvitamin D3 reversibly blocks the progression of relapsing encephalomyelitis, a model of multiple sclerosis. Proc. Natl. Acad. Sci. USA 1996, 93, 7861–7864. [Google Scholar] [CrossRef] [PubMed]
- Burton, J.M.; Kimball, S.; Vieth, R.; Bar-Or, A.; Dosch, H.M.; Cheung, R.; Gagne, D.; D’Souza, C.; Ursell, M.; O’Connor, P. A phase I/II dose-escalation trial of vitamin D3 and calcium in multiple sclerosis. Neurology 2010, 74, 1852–1859. [Google Scholar] [CrossRef] [PubMed]
- Smolders, J.; Thewissen, M.; Peelen, E.; Menheere, P.; Tervaert, J.W.; Damoiseaux, J.; Hupperts, R. Vitamin D status is positively correlated with regulatory T cell function in patients with multiple sclerosis. PLoS ONE 2009, 4, e6635. [Google Scholar] [CrossRef] [PubMed]
- Smolders, J.; Peelen, E.; Thewissen, M.; Cohen Tervaert, J.W.; Menheere, P.; Hupperts, R.; Damoiseaux, J. Safety and T cell modulating effects of high dose vitamin D3 supplementation in multiple sclerosis. PLoS ONE 2010, 5, e15235. [Google Scholar] [CrossRef] [PubMed]
- Wergeland, S.; Torkildsen, O.; Myhr, K.M.; Aksnes, L.; Mork, S.J.; Bo, L. Dietary vitamin D3 supplements reduce demyelination in the cuprizone model. PLoS ONE 2011, 6, e26262. [Google Scholar] [CrossRef] [PubMed]
- Al-Jaderi, Z.; Maghazachi, A.A. Effects of vitamin D3, calcipotriol and FTY720 on the expression of surface molecules and cytolytic activities of human natural killer cells and dendritic cells. Toxins 2013, 5, 1932–1947. [Google Scholar] [CrossRef] [PubMed]
- Linker, R.A.; Lee, D.H.; Ryan, S.; van Dam, A.M.; Conrad, R.; Bista, P.; Zeng, W.; Hronowsky, X.; Buko, A.; Chollate, S.; et al. Fumaric acid esters exert neuroprotective effects in neuroinflammation via activation of the Nrf2 antioxidant pathway. Brain 2011, 134 (Pt 3), 678–692. [Google Scholar] [CrossRef] [PubMed]
- Schimrigk, S.; Brune, N.; Hellwig, K.; Lukas, C.; Bellenberg, B.; Rieks, M.; Hoffmann, V.; Pohlau, D.; Przuntek, H. Oral fumaric acid esters for the treatment of active multiple sclerosis: An open-label, baseline-controlled pilot study. Eur. J. Neurol. 2006, 13, 604–610. [Google Scholar] [CrossRef] [PubMed]
- Ghoreschi, K.; Bruck, J.; Kellerer, C.; Deng, C.; Peng, H.; Rothfuss, O.; Hussain, R.Z.; Gocke, A.R.; Respa, A.; Glocova, I.; et al. Fumarates improve psoriasis and multiple sclerosis by inducing type II dendritic cells. J. Exp. Med. 2011, 208, 2291–2303. [Google Scholar] [CrossRef] [PubMed]
- De Jong, R.; Bezemer, A.C.; Zomerdijk, T.P.; Pouw-Kraan, T.; Ottenhoff, T.H.; Nibbering, P.H. Selective stimulation of T helper 2 cytokine responses by the anti-psoriasis agent monomethylfumarate. Eur. J. Immunol. 1996, 26, 2067–2074. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Guerau-de-Arellano, M.; Mehta, V.B.; Yang, Y.; Huss, D.J.; Papenfuss, T.L.; Lovett-Racke, A.E.; Racke, M.K. Dimethyl fumarate inhibits dendritic cell maturation via nuclear factor kappaB (NF-kappaB) and extracellular signal-regulated kinase 1 and 2 (ERK1/2) and mitogen stress-activated kinase 1 (MSK1) signaling. J. Biol. Chem. 2012, 287, 28017–28026. [Google Scholar] [CrossRef] [PubMed]
- Vego, H.; Sand, K.L.; Hoglund, R.A.; Fallang, L.E.; Gundersen, G.; Holmoy, T.; Maghazachi, A.A. Monomethyl fumarate augments NK cell lysis of tumor cells through degranulation and the upregulation of NKp46 and CD107a. Cell. Mol. Immunol. 2014. [Google Scholar] [CrossRef] [PubMed]
- Chiuso-Minicucci, F.; Ishikawa, L.L.; Mimura, L.A.; Fraga-Silva, T.F.; Franca, T.G.; Zorzella-Pezavento, S.F.; Marques, C.; Ikoma, M.R.; Sartori, A. Treatment with Vitamin D/MOG Association Suppresses Experimental Autoimmune Encephalomyelitis. PLoS ONE 2015, 10, e0125836. [Google Scholar] [CrossRef] [PubMed]
- Litjens, N.H.; Rademaker, M.; Ravensbergen, B.; Thio, H.B.; van Dissel, J.T.; Nibbering, P.H. Effects of monomethylfumarate on dendritic cell differentiation. Br. J. Dermatol. 2006, 154, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Litjens, N.H.; Rademaker, M.; Ravensbergen, B.; Rea, D.; van der Plas, M.J.; Thio, B.; Walding, A.; van Dissel, J.T.; Nibbering, P.H. Monomethylfumarate affects polarization of monocyte-derived dendritic cells resulting in down-regulated Th1 lymphocyte responses. Eur. J. Immunol. 2004, 34, 565–575. [Google Scholar] [CrossRef] [PubMed]
- Steinman, R.M. Decisions about dendritic cells; past, present, and future. Annu. Rev. Immunol. 2012, 30, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Becklund, B.R.; Hansen, D.W., Jr.; DeLuca, H.F. Enhancement of 1,25-dihydroxyvitamin D3-mediated suppression of experimental autoimmune encephalomyelitis by calcitonin. Proc. Natl. Acad. Sci. USA 2009, 106, 5276–5281. [Google Scholar] [CrossRef] [PubMed]
- Cantorna, M.T.; Woodward, W.D.; Hayes, C.E.; Deluca, H.F. 1,25-Dihydroxyvitamin D3 is a positive regulator for the two anti-encephalitogenic cytokines TGF-beta 1 and IL-4. J. Immunol. 1998, 160, 5314–5319. [Google Scholar] [PubMed]
- Lemire, J.M.; Archer, D.C.; Beck, L.; Spiegelberg, H.L. Immunosuppressive actions of 1,25-dihydroxyvitamin D3: Preferential inhibition of Th1 functions. J. Nutr. 1995, 125 (Suppl. 6), 1704S–1708S. [Google Scholar] [PubMed]
- Joshi, S.; Pantalena, L.C.; Liu, X.K.; Gaffen, S.L.; Liu, H.; Rohowsky-Kochan, C.; Ichiyama, K.; Yoshimura, A.; Steinman, L.; Christakos, S.; et al. 1,25-Dihydroxyvitamin D(3) ameliorates Th17 autoimmunity via transcriptional modulation of interleukin-17A. Mol. Cell. Biol. 2011, 31, 3653–3669. [Google Scholar] [CrossRef] [PubMed]
- Mayne, C.G.; Spanier, J.A.; Relland, L.M.; Williams, C.B.; Hayes, C.E. 1,25-Dihydroxyvitamin D3 acts directly on the T lymphocyte vitamin D receptor to inhibit experimental autoimmune encephalomyelitis. Eur. J. Immunol. 2011, 41, 822–832. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.H.; Cha, H.R.; Lee, D.S.; Seo, K.Y.; Kweon, M.N. 1,25-Dihydroxyvitamin D3 inhibits the differentiation and migration of T(H)17 cells to protect against experimental autoimmune encephalomyelitis. PLoS ONE 2010, 5, e12925. [Google Scholar] [CrossRef] [PubMed]
- Boonstra, A.; Barrat, F.J.; Crain, C.; Heath, V.L.; Savelkoul, H.F.; O’Garra, A. 1alpha,25-Dihydroxyvitamin D3 has a direct effect on naive CD4(+) T cells to enhance the development of Th2 cells. J. Immunol. 2001, 167, 4974–4980. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Park, S.; Cheon, S.; Lee, J.H.; Kim, S.; Hur, D.Y.; Kim, T.S.; Yoon, S.R.; Yang, Y.; Bang, S.I.; et al. 1,25-Dihydroxyvitamin D(3) enhances NK susceptibility of human melanoma cells via Hsp60-mediated FAS expression. Eur. J. Immunol. 2011, 41, 2937–2946. [Google Scholar] [CrossRef] [PubMed]
- Rinaldi, A.O.; Sanseverino, I.; Purificato, C.; Cortese, A.; Mechelli, R.; Francisci, S.; Salvetti, M.; Millefiorini, E.; Gessani, S.; Gauzzi, M.C. Increased circulating levels of vitamin D binding protein in MS patients. Toxins 2015, 7, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Dibbert, S.; Clement, B.; Skak-Nielsen, T.; Mrowietz, U.; Rostami-Yazdi, M. Detection of fumarate-glutathione adducts in the portal vein blood of rats: Evidence for rapid dimethylfumarate metabolism. Arch. Dermatol. Res. 2013, 305, 447–451. [Google Scholar] [CrossRef] [PubMed]
- Scannevin, R.H.; Chollate, S.; Jung, M.Y.; Shackett, M.; Patel, H.; Bista, P.; Zeng, W.; Ryan, S.; Yamamoto, M.; Lukashev, M.; et al. Fumarates promote cytoprotection of central nervous system cells against oxidative stress via the nuclear factor (erythroid-derived 2)-like 2 pathway. J. Pharmacol. Exp. Ther. 2012, 341, 274–284. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Lu, J.Y.; Zheng, X.; Yang, Y.; Reagan, J.D. The psoriasis drug monomethylfumarate is a potent nicotinic acid receptor agonist. Biochem. Biophys. Res. Commun. 2008, 375, 562–556. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Assmann, J.C.; Krenz, A.; Rahman, M.; Grimm, M.; Karsten, C.M.; Köhl, J.; Offermanns, S.; Wettschureck, N.; Schwaninger, M. Hydroxycarboxylic acid receptor 2 mediates dimethyl fumarate’s protective effect in EAE. J. Clin. Investig. 2014, 124, 2188–2192. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Jaderi, Z.; Maghazachi, A.A. Vitamin D3 and Monomethyl Fumarate Enhance Natural Killer Cell Lysis of Dendritic Cells and Ameliorate the Clinical Score in Mice Suffering from Experimental Autoimmune Encephalomyelitis. Toxins 2015, 7, 4730-4744. https://doi.org/10.3390/toxins7114730
Al-Jaderi Z, Maghazachi AA. Vitamin D3 and Monomethyl Fumarate Enhance Natural Killer Cell Lysis of Dendritic Cells and Ameliorate the Clinical Score in Mice Suffering from Experimental Autoimmune Encephalomyelitis. Toxins. 2015; 7(11):4730-4744. https://doi.org/10.3390/toxins7114730
Chicago/Turabian StyleAl-Jaderi, Zaidoon, and Azzam A. Maghazachi. 2015. "Vitamin D3 and Monomethyl Fumarate Enhance Natural Killer Cell Lysis of Dendritic Cells and Ameliorate the Clinical Score in Mice Suffering from Experimental Autoimmune Encephalomyelitis" Toxins 7, no. 11: 4730-4744. https://doi.org/10.3390/toxins7114730
APA StyleAl-Jaderi, Z., & Maghazachi, A. A. (2015). Vitamin D3 and Monomethyl Fumarate Enhance Natural Killer Cell Lysis of Dendritic Cells and Ameliorate the Clinical Score in Mice Suffering from Experimental Autoimmune Encephalomyelitis. Toxins, 7(11), 4730-4744. https://doi.org/10.3390/toxins7114730