Tracing Monotreme Venom Evolution in the Genomics Era

Abstract
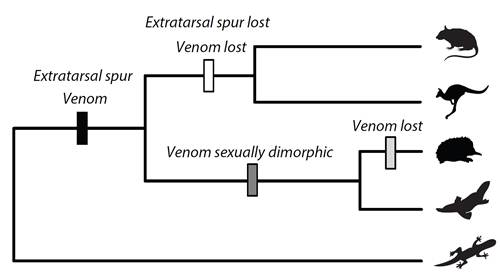
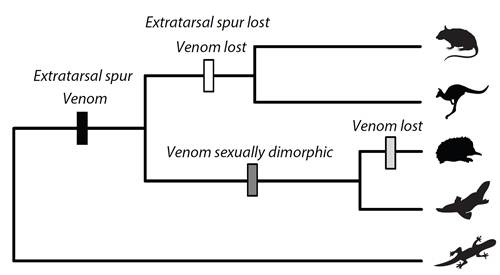
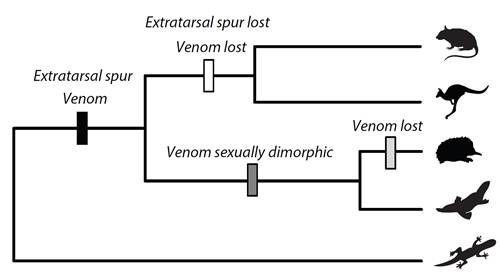
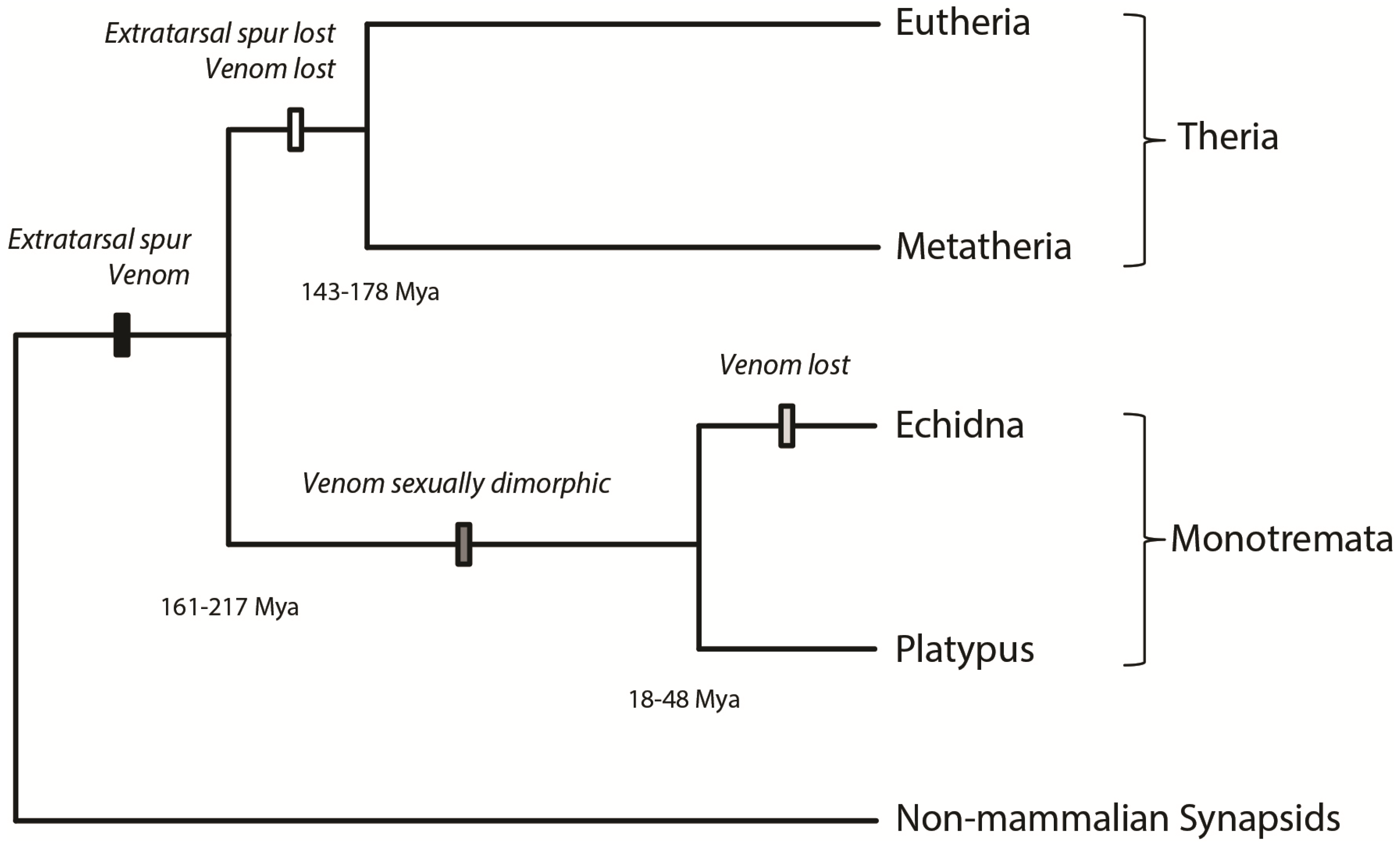
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction: Mammalian Venom
2. Venom in Ancient Monotremes

3. Derived Venom System: The Platypus
3.1. The Crural System

3.2. Venom Function
3.3. Platypus Venom Composition: Pharmacological and Proteomic Characterization
3.4. Platypus Venom Composition: Genomic Insights
3.5. Platypus Venom Gene Evolution
4. Secondary Losses of the Venom System: The Echidnas
5. Looking Forward: Genomics and Mammalian Venom
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Fry, B.G.; Roelants, K.; Champagne, D.E.; Scheib, H.; Tyndall, J.D.; King, G.F.; Nevalainen, T.J.; Norman, J.A.; Lewis, R.J.; Norton, R.S.; et al. The toxicogenomic multiverse: Convergent recruitment of proteins into animal venoms. Annu. Rev. Genomics Hum. Genet. 2009, 10, 483–511. [Google Scholar]
- Dufton, M.J. Venomous mammals. Pharmacol. Ther. 1992, 53, 199–215. [Google Scholar] [CrossRef]
- Tellgren-Roth, A.; Dittmar, K.; Massey, S.E.; Kemi, C.; Tellgren-Roth, C.; Savolainen, P.; Lyons, L.A.; Liberles, D.A. Keeping the blood flowing-plasminogen activator genes and feeding behavior in vampire bats. Naturwissenschaften 2009, 96, 39–47. [Google Scholar]
- Nekaris, K.A.-I.; Moore, R.; Rode, E.; Fry, B. Mad, bad and dangerous to know: The biochemistry, ecology and evolution of slow loris venom. J. Venom. Anim. Toxins Incl. Trop. Dis. 2013, 19, 21. [Google Scholar] [CrossRef]
- Whittington, C.M.; Koh, J.M.S.; Warren, W.C.; Papenfuss, A.T.; Torres, A.M.; Kuchel, P.W.; Belov, K. Understanding and utilising mammalian venom via a platypus venom transcriptome. J. Proteomics 2009, 72, 155–164. [Google Scholar] [CrossRef]
- Flannery, T.F.; Groves, C.P. A revision of the genus Zaglossus (Monotremata, Tachyglossidae), with description of new species and subspecies. Mammalia 1998, 62, 367–396. [Google Scholar]
- Warren, W.C.; Hillier, L.W.; Graves, J.A.M.; Birney, E.; Ponting, C.P.; Grutzner, F.; Belov, K.; Miller, W.; Clarke, L.; Chinwalla, A.T.; et al. Genome analysis of the platypus reveals unique signatures of evolution. Nature 2008, 453, 175–183. [Google Scholar] [CrossRef]
- Musser, A.M. Review of the monotreme fossil record and comparison of palaeontological and molecular data. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2003, 136, 927–942. [Google Scholar] [CrossRef]
- Rowe, T.; Rich, T.H.; Vickers-Rich, P.; Springer, M.; Woodburne, M.O. The oldest platypus and its bearing on divergence timing of the platypus and echidna clades. Proc. Natl. Acad. Sci. USA 2008, 105, 1238–1242. [Google Scholar]
- Phillips, M.J.; Bennett, T.H.; Lee, M.S.Y. Molecules, morphology, and ecology indicate a recent, amphibious ancestry for echidnas. Proc. Natl. Acad. Sci. USA 2009, 106, 17089–17094. [Google Scholar] [CrossRef]
- Whittington, C.M.; Papenfuss, A.T.; Bansal, P.; Torres, A.M.; Wong, E.S.W.; Deakin, J.E.; Graves, T.; Alsop, A.; Schatzkamer, K.; Kremitzki, C.; et al. Defensins and the convergent evolution of platypus and reptile venom genes. Genome Res. 2008, 18, 986–994. [Google Scholar] [CrossRef]
- Whittington, C.M.; Belov, K. Platypus venom: A review. Aust. Mammal. 2007, 29, 57–62. [Google Scholar] [CrossRef]
- Schondube, J.E.; Herrera-M, L.G.; Martínez del Rio, C. Diet and the evolution of digestion and renal function in phyllostomid bats. Zoology 2001, 104, 59–73. [Google Scholar] [CrossRef]
- Hurum, J.H.; Luo, Z.X.; Kielan-Jaworowska, Z. Were mammals originally venomous? Acta Palaeontol. Pol. 2006, 51, 1–11. [Google Scholar]
- Luo, Z.X.; Kielan-Jaworowska, Z.; Cifelli, R.L. In quest for a phylogeny of Mesozoic mammals. Acta Palaeontol. Pol. 2002, 47, 1–78. [Google Scholar]
- Grant, T. Platypus, 4th ed.; CSIRO Publishing: Collingwood, Australia, 2007. [Google Scholar]
- Temple-Smith, P.D. Seasonal Breeding Biology of the Platypus, Ornithorhynchus Anatinus (Shaw, 1799), with Special Reference to the Male; Australian National University: Canberra, Australia, 1973. [Google Scholar]
- Krause, W.J. Morphological and histochemical observations on the crural gland-spur apparatus of the echidna (Tachyglossus aculeatus) together with comparative observations on the femoral gland-spur apparatus of the duckbilled platypus (Ornithorhyncus anatinus). Cells Tissues Organs 2010, 191, 336–354. [Google Scholar] [CrossRef]
- Fenner, P.J.; Williamson, J.A.; Myers, D. Platypus envenomation—A painful learning experience. Med. J. Aust. 1992, 157, 829–832. [Google Scholar]
- Burrell, H. The Platypus; Angus & Robinson Ltd: Sydney, Australia, 1927. [Google Scholar]
- Munday, B.L.; Whittington, R.J.; Stewart, N.J. Disease conditions and subclinical infections of the platypus (Ornithorhynchus anatinus). Philos. Trans. R. Soc. Lond. B Biol. Sci. 1998, 353, 1093–1099. [Google Scholar] [CrossRef]
- Morgenstern, D.; King, G.F. The venom optimization hypothesis revisited. Toxicon 2013, 63, 120–128. [Google Scholar] [CrossRef]
- King, G.F. Venoms as a platform for human drugs: Translating toxins into therapeutics. Expert Opin. Biol. Ther. 2011, 11, 1469–1484. [Google Scholar] [CrossRef]
- Martin, C.J.; Tidswell, F. Observations on the femoral gland of Ornithorhynchus and its secretion; together with an experimental enquiry concerning its supposed toxic action. Proc. Linn. Soc. N. S. W. 1895, 9, 471–500. [Google Scholar]
- Kellaway, C.H.; Le Messurier, D.H. The venom of the platypus (Ornithorhynchus anatinus). Aust. J. Exp. Biol. Med. Sci. 1935, 13, 205–221. [Google Scholar] [CrossRef]
- De Plater, G.; Martin, R.L.; Milburn, P.J. A pharmacological and biochemical investigation of the venom from the platypus (Ornithorhynchus anatinus). Toxicon 1995, 33, 157–169. [Google Scholar] [CrossRef]
- De Plater, G.M.; Milburn, P.J.; Martin, R.L. Venom from the platypus, Ornithorhynchus anatinus, induces a calcium-dependent current in cultured dorsal root ganglion cells. J. Neurophysiol. 2001, 85, 1340–1345. [Google Scholar]
- Kourie, J.I. A component of platypus (Ornithorhynchus anatinus) venom forms slow-kinetic cation channels. J. Membr. Biol. 1999, 172, 37–45. [Google Scholar] [CrossRef]
- Torres, A.M.; de Plater, G.M.; Doverskog, M.; Birinyi-Strachan, L.C.; Nicholson, G.M.; Gallagher, C.H.; Kuchel, P.W. Defensin-like peptide-2 from platypus venom: member of a class of peptides with a distinct structural fold. Biochem. J. 2000, 348, 649–656. [Google Scholar] [CrossRef]
- Torres, A.M.; Wang, X.H.; Fletcher, J.I.; Alewood, D.; Alewood, P.F.; Smith, R.; Simpson, R.J.; Nicholson, G.M.; Sutherland, S.K.; Gallagher, C.H.; et al. Solution structure of a defensin-like peptide from platypus venom. Biochem. J. 1999, 341, 785–794. [Google Scholar] [CrossRef]
- De Plater, G.M.; Martin, R.L.; Milburn, P.J. The natriuretic peptide (OvCNP-39) from platypus (Ornithorhynchus anatinus) venom relaxes the isolated rat uterus and promotes oedema and mast cell histamine release. Toxicon 1998, 36, 847–857. [Google Scholar] [CrossRef]
- De Plater, G. Fractionation, Primary Structural Characterisation and Biological Activities of Polypeptides from the Venom of the Platypus (Ornithorhynchus anatinus); Australian National University: Canberra, Australia, 1998. [Google Scholar]
- Torres, A.M.; Tsampazi, M.; Tsampazi, C.; Kennett, E.C.; Belov, K.; Geraghty, D.P.; Bansal, P.S.; Alewood, P.F.; Kuchel, P.W. Mammalian l-to-d-amino-acid-residue isomerase from platypus venom. FEBS Lett. 2006, 580, 1587–1591. [Google Scholar] [CrossRef]
- Bansal, P.S.; Torres, A.M.; Crossett, B.; Wong, K.K.Y.; Koh, J.M.S.; Geraghty, D.P.; Vandenberg, J.I.; Kuchel, P.W. Substrate specificity of platypus venom l-to-d-peptide isomerase. J. Biol. Chem. 2008, 283, 8969–8975. [Google Scholar] [CrossRef]
- Whittington, C.M.; Belov, K. Platypus venom genes expressed in non-venom tissues. Aust. J. Zool. 2009, 57, 199–202. [Google Scholar] [CrossRef]
- Whittington, C.M.; Papenfuss, A.T.; Kuchel, P.W.; Belov, K. Expression patterns of platypus defensin and related venom genes across a range of tissue types reveal the possibility of broader functions for OvDLPs than previously suspected. Toxicon 2008, 52, 559–565. [Google Scholar] [CrossRef]
- Whittington, C.M.; Papenfuss, A.T.; Locke, D.P.; Mardis, E.R.; Wilson, R.K.; Abubucker, S.; Mitreva, M.; Wong, E.S.W.; Hsu, A.L.; Kuchel, P.W.; et al. Novel venom gene discovery in the platypus. Genome Biol. 2010, 11. [Google Scholar] [CrossRef]
- Wong, E.S.W.; Morgenstern, D.; Mofiz, E.; Gombert, S.; Morris, K.M.; Temple-Smith, P.; Renfree, M.B.; Whittington, C.M.; King, G.F.; Warren, W.C.; et al. Proteomics and deep sequencing comparison of seasonally active venom glands in the platypus reveals novel venom peptides and distinct expression profiles. Mol. Cell. Proteomics 2012, 11, 1354–1364. [Google Scholar] [CrossRef]
- Koh, J.M.S.; Bansal, P.S.; Torres, A.M.; Kuchel, P.W. Platypus venom: Source of novel compounds. Aust. J. Zool. 2009, 57, 203–210. [Google Scholar] [CrossRef]
- Kochva, E. The origin of snakes and evolution of the venom apparatus. Toxicon 1987, 25, 65–106. [Google Scholar] [CrossRef]
- Fry, B.G. From genome to “venome”: Molecular origin and evolution of the snake venom proteome inferred from phylogenetic analysis of toxin sequences and related body proteins. Genome Res. 2005, 15, 403–420. [Google Scholar] [CrossRef]
- Nei, M.; Rooney, A.P. Concerted birth-and-death evolution of multigene families. Annu. Rev. Genet. 2005, 39, 121–152. [Google Scholar] [CrossRef]
- Whittington, C.M.; Sharp, J.A.; Papenfuss, A.; Belov, K. No evidence of expression of two classes of natural antibiotics (cathelicidins and defensins) in a sample of platypus milk. Aust. J. Zool. 2009, 57, 211–217. [Google Scholar] [CrossRef]
- Casewell, N.R.; Wuster, W.; Vonk, F.J.; Harrison, R.A.; Fry, B.G. Complex cocktails: The evolutionary novelty of venoms. Trends Ecol. Evol. 2013, 28, 219–229. [Google Scholar]
- Duda, T.F.; Palumbi, S.R. Molecular genetics of ecological diversification: Duplication and rapid evolution of toxin genes of the venomous gastropod Conus. Proc. Natl. Acad. Sci. USA 1999, 96, 6820–6823. [Google Scholar] [CrossRef]
- Escoubas, P. Molecular diversification in spider venoms: A web of combinatorial peptide libraries. Mol. Divers. 2006, 10, 545–554. [Google Scholar]
- Vonk, F.J.; Casewell, N.R.; Henkel, C.V.; Heimberg, A.M.; Jansen, H.J.; McCleary, R.J.R.; Kerkkamp, H.M.E.; Vos, R.A.; Guerreiro, I.; Calvete, J.J.; et al. The king cobra genome reveals dynamic gene evolution and adaptation in the snake venom system. Proc. Natl. Acad. Sci. USA 2013, 110, 20651–20656. [Google Scholar] [CrossRef]
- Ikeda, N.; Chijiwa, T.; Matsubara, K.; Oda-Ueda, N.; Hattori, S.; Matsuda, Y.; Ohno, M. Unique structural characteristics and evolution of a cluster of venom phospholipase A2 isozyme genes of Protobothrops flavoviridis snake. Gene 2010, 461, 15–25. [Google Scholar] [CrossRef]
- Wong, E.S.W.; Papenfuss, A.T.; Whittington, C.M.; Warren, W.C.; Belov, K. A Limited role for gene duplications in the evolution of platypus venom. Mol. Biol. Evol. 2012, 29, 167–177. [Google Scholar] [CrossRef]
- Cousin, X.; Bon, S.; Massoulié, J.; Bon, C. Identification of a novel type of alternatively spliced exon from the acetylcholinesterase gene of Bungarus fasciatus: Molecular forms of acetylcholinesterase in the snake liver and muscle. J. Biol. Chem. 1998, 273, 9812–9820. [Google Scholar] [CrossRef]
- Camens, A.B. Were early Tertiary monotremes really all aquatic? Inferring paleobiology and phylogeny from a depauperate fossil record. Proc. Natl. Acad. Sci. USA 2010, 107. [Google Scholar] [CrossRef]
- Griffiths, M. The Biology of the Monotremes; Academic Press, Inc.: New York, NY, USA, 1978. [Google Scholar]
- Augee, M.L.; Gooden, B.; Musser, A. Echidna: Extraordinary Egg-Laying Mammal; CSIRO Publishing: Collingwood, Australia, 2006. [Google Scholar]
- Harris, R.L.; Davies, N.W.; Nicol, S.C. Chemical composition of odorous secretions in the Tasmanian short-beaked echidna (Tachyglossus aculeatus setosus). Chem. Senses 2012, 37, 819–836. [Google Scholar] [CrossRef]
- Wong, E.S.W.; Nicol, S.; Warren, W.C.; Belov, K. Echidna venom gland transcriptome provides insights into the evolution of monotreme venom. PLoS ONE 2013, 8, e79092. [Google Scholar]
- Koh, J.M.S.; Haynes, L.; Belov, K.; Kuchel, P.W. l-to-d-peptide isomerase in male echidna venom. Aust. J. Zool. 2010, 58, 284–288. [Google Scholar] [CrossRef]
- Pupko, T.; Pe, I.; Shamir, R.; Graur, D. A fast algorithm for joint reconstruction of ancestral amino acid sequences. Mol. Biol. Evol. 2000, 17, 890–896. [Google Scholar] [CrossRef]
- Casewell, N.R. On the ancestral recruitment of metalloproteinases into the venom of snakes. Toxicon 2012, 60, 449–454. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Whittington, C.M.; Belov, K. Tracing Monotreme Venom Evolution in the Genomics Era. Toxins 2014, 6, 1260-1273. https://doi.org/10.3390/toxins6041260
Whittington CM, Belov K. Tracing Monotreme Venom Evolution in the Genomics Era. Toxins. 2014; 6(4):1260-1273. https://doi.org/10.3390/toxins6041260
Chicago/Turabian StyleWhittington, Camilla M., and Katherine Belov. 2014. "Tracing Monotreme Venom Evolution in the Genomics Era" Toxins 6, no. 4: 1260-1273. https://doi.org/10.3390/toxins6041260
APA StyleWhittington, C. M., & Belov, K. (2014). Tracing Monotreme Venom Evolution in the Genomics Era. Toxins, 6(4), 1260-1273. https://doi.org/10.3390/toxins6041260