Quo Vadis Venomics? A Roadmap to Neglected Venomous Invertebrates


Abstract
:1. Introduction
2. Arthropoda
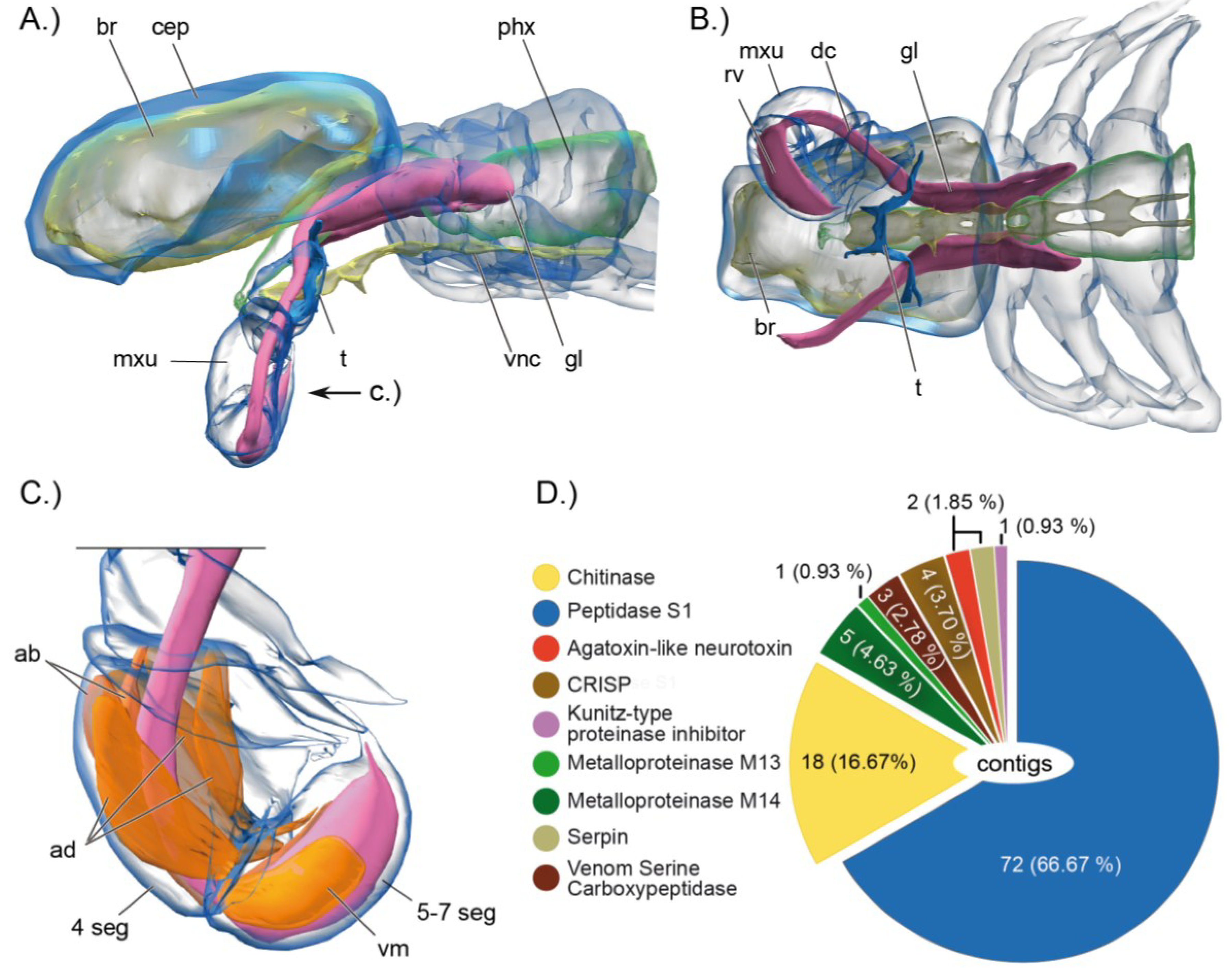
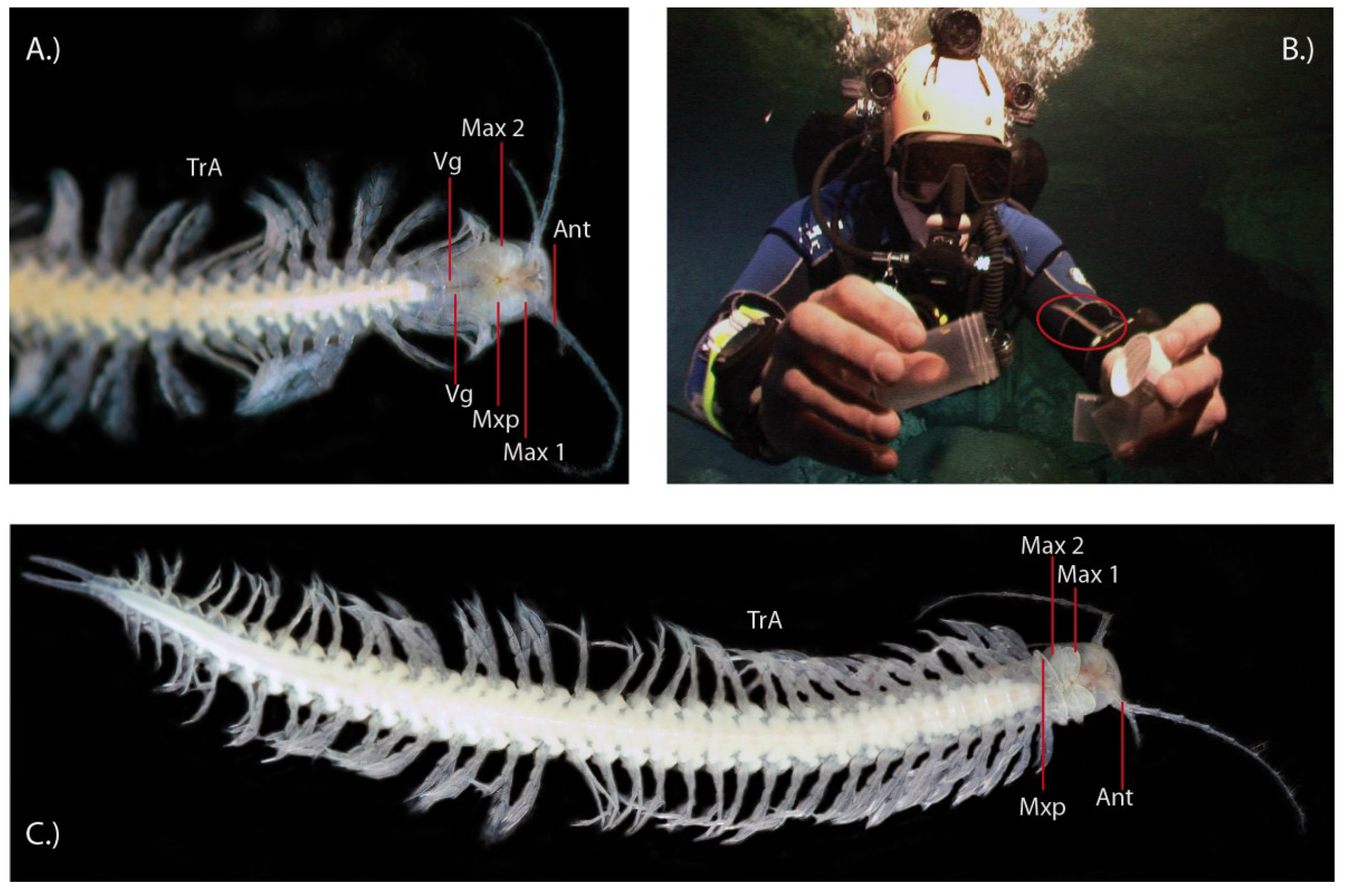
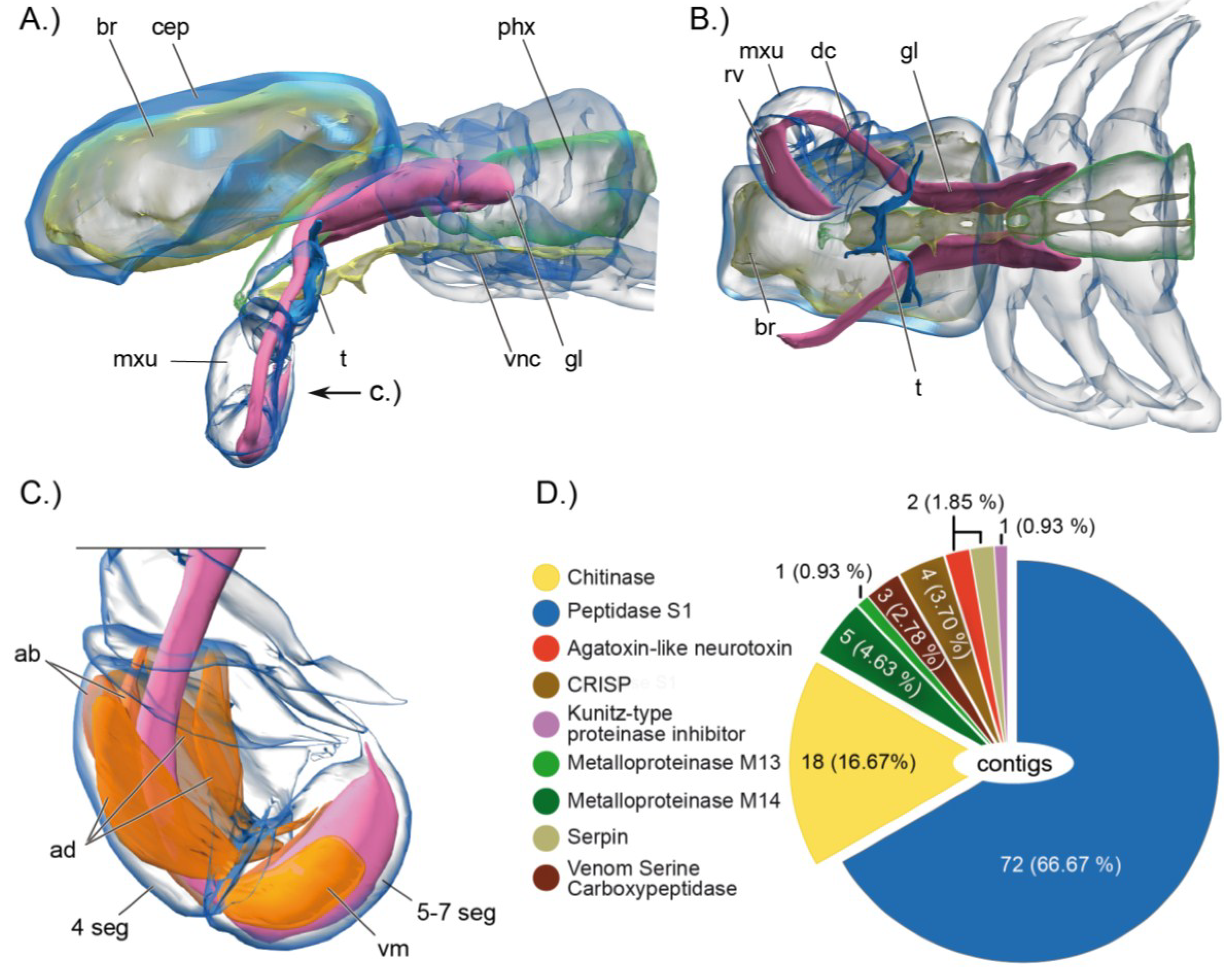
2.1. Remipedes, the First Venomous Crustaceans
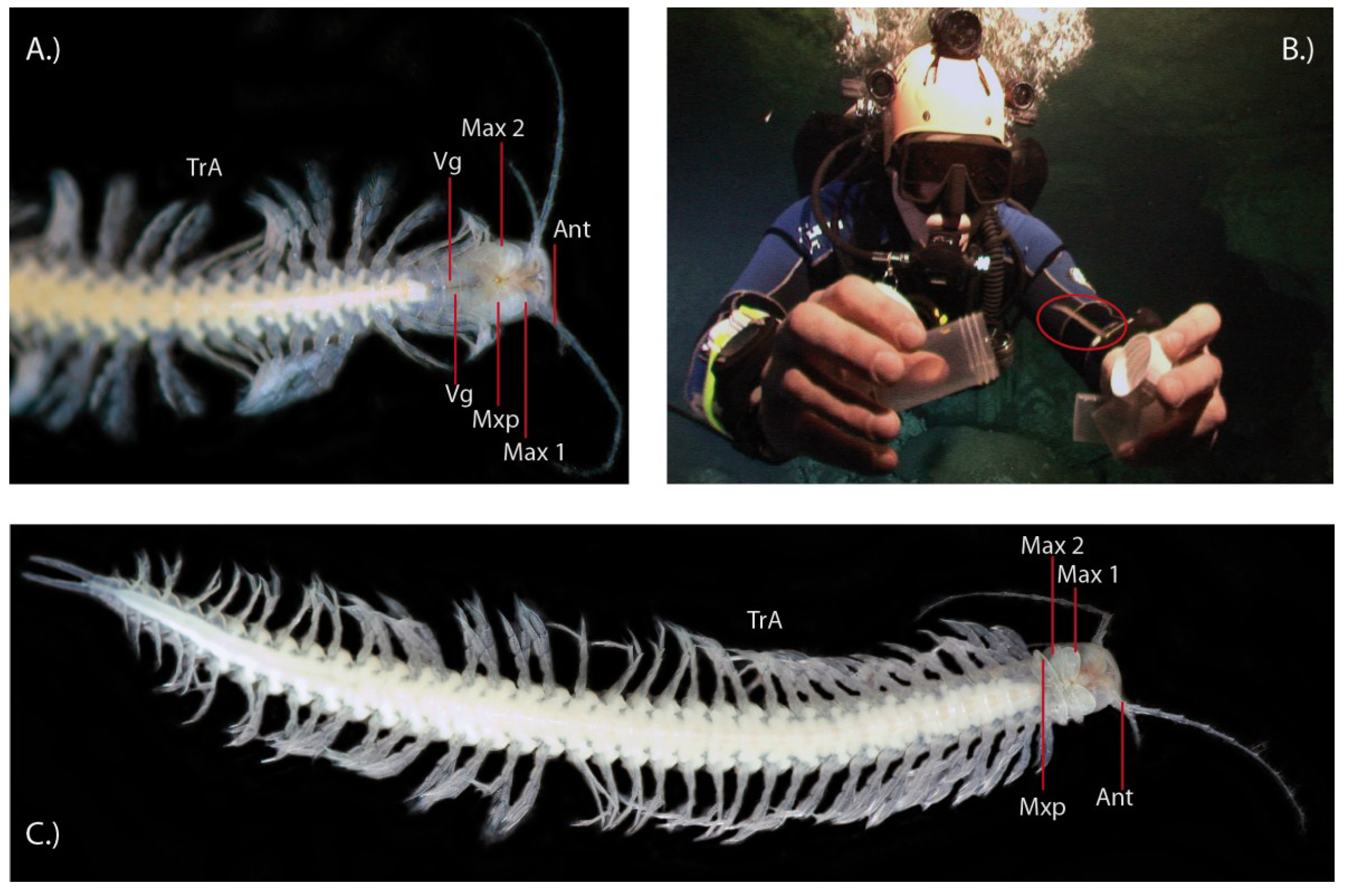
2.2. Other Neglected Putatively Venomous Crustaceans
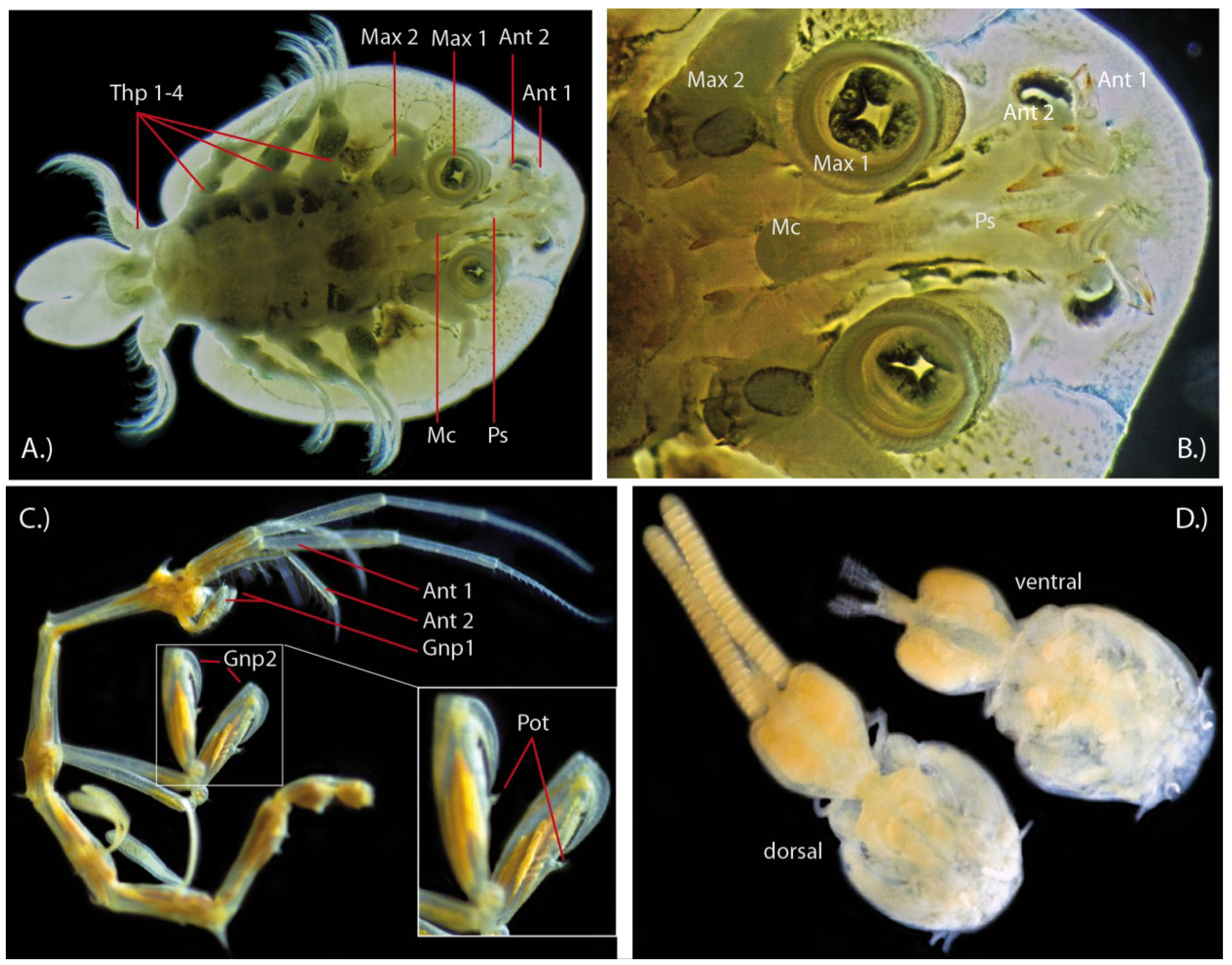
2.2.1. Fish Lice (Branchiura)

2.2.2. Skeleton Shrimp (Caprellidae)
2.2.3. Copepoda
2.2.4. Gnathiid Isopods
2.3. Neglected Venomous Insects
2.3.1. Robber Flies (Asilidae)

2.3.2. Horse Flies (Tabanidae)
2.3.3. Marsh Flies (Sciomyzidae)
2.3.4. Further Neglected Fly Groups
2.4. Neglected Centipedes

2.5. Arachnida
2.5.1. Pseudoscorpions


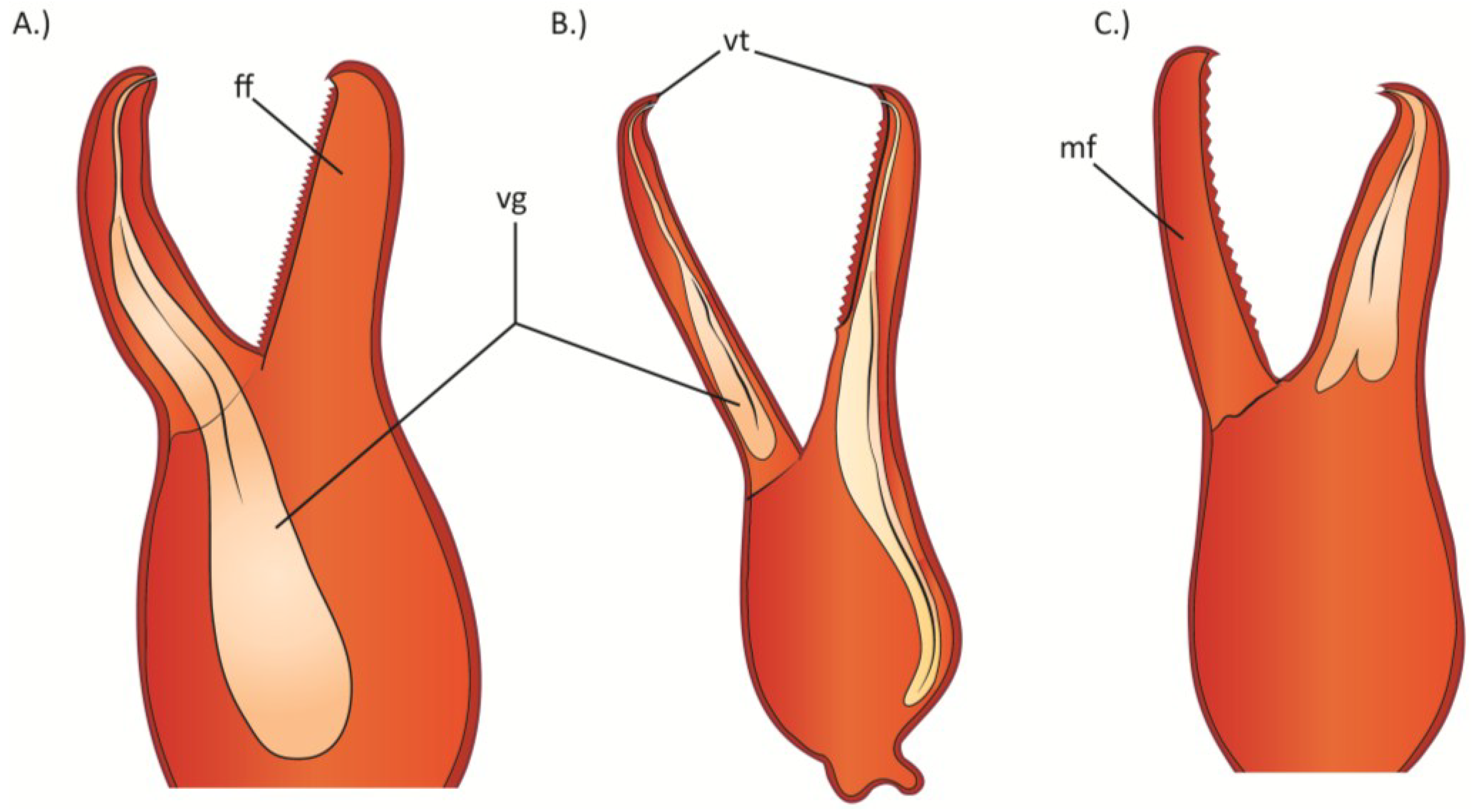
2.5.2. Camel Spiders or Wind Scorpions (Solifugae)
3. Lophotrochozoa (=Spiralia)
3.1. Annelida
3.1.1. Leeches (Hirudinea, Clitellata)
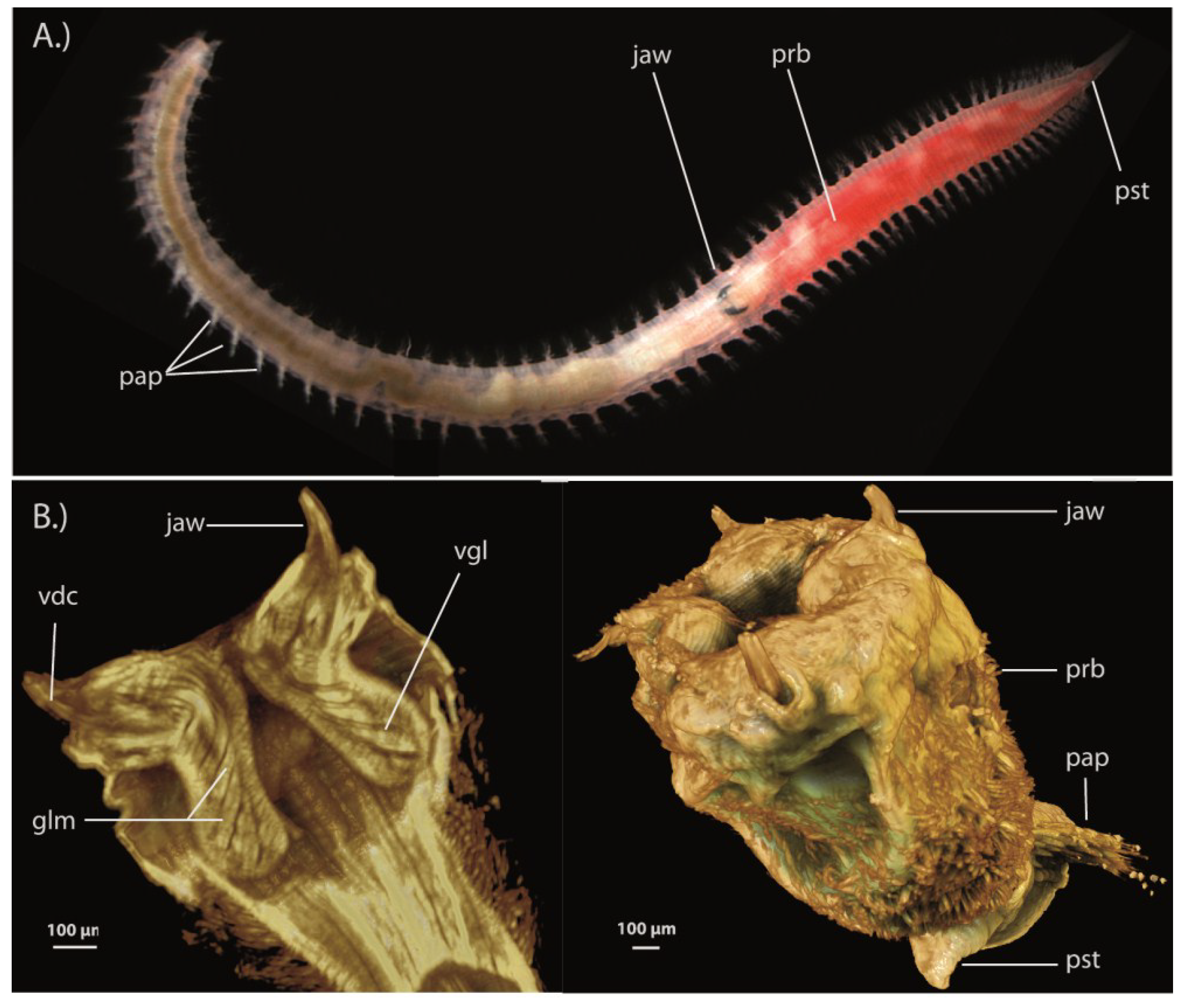
3.1.2. Bristleworms (Amphinomida)
3.1.3. Earthworms (Eisenia)
3.1.4. Bloodworms (Glyceridae)
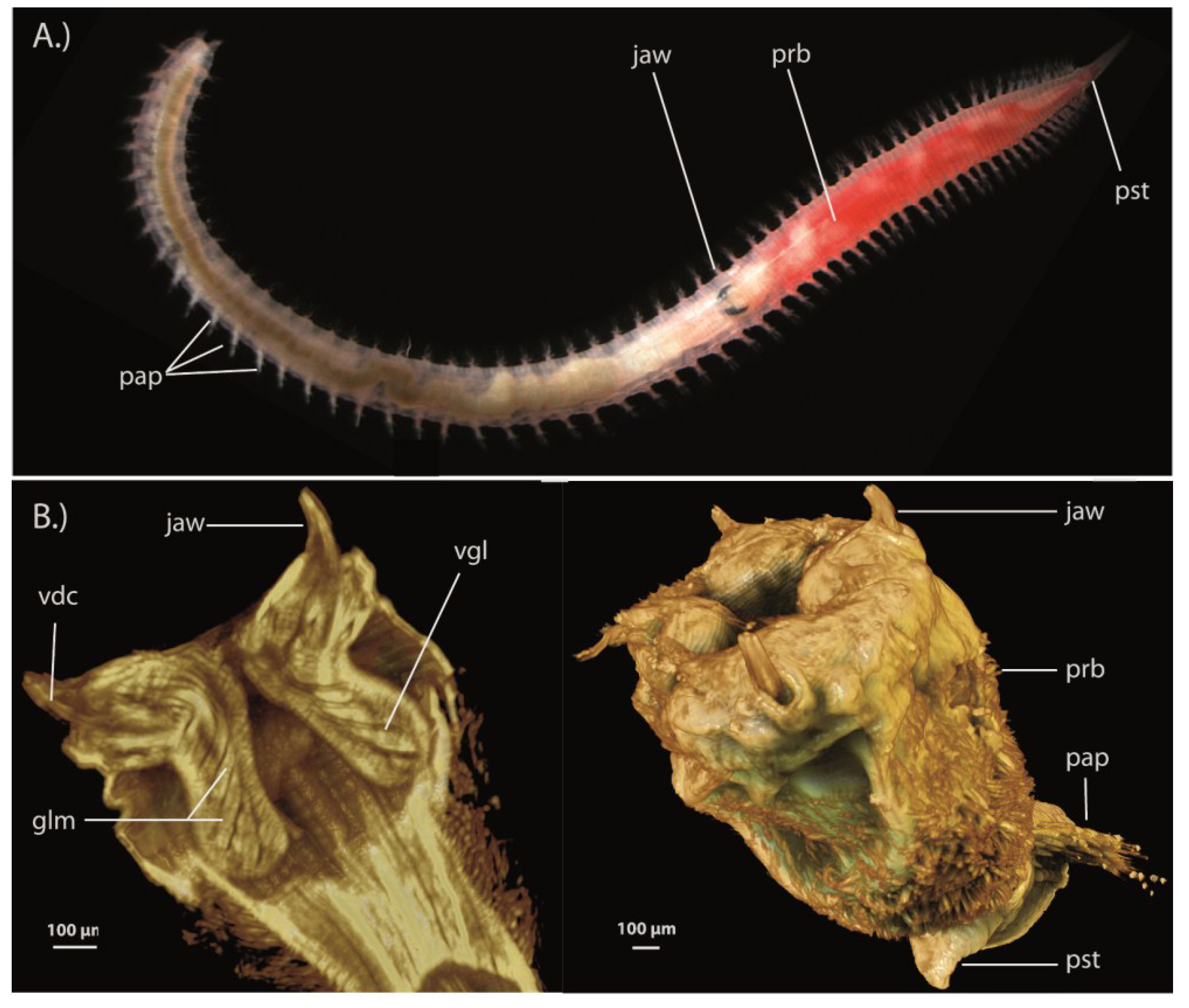
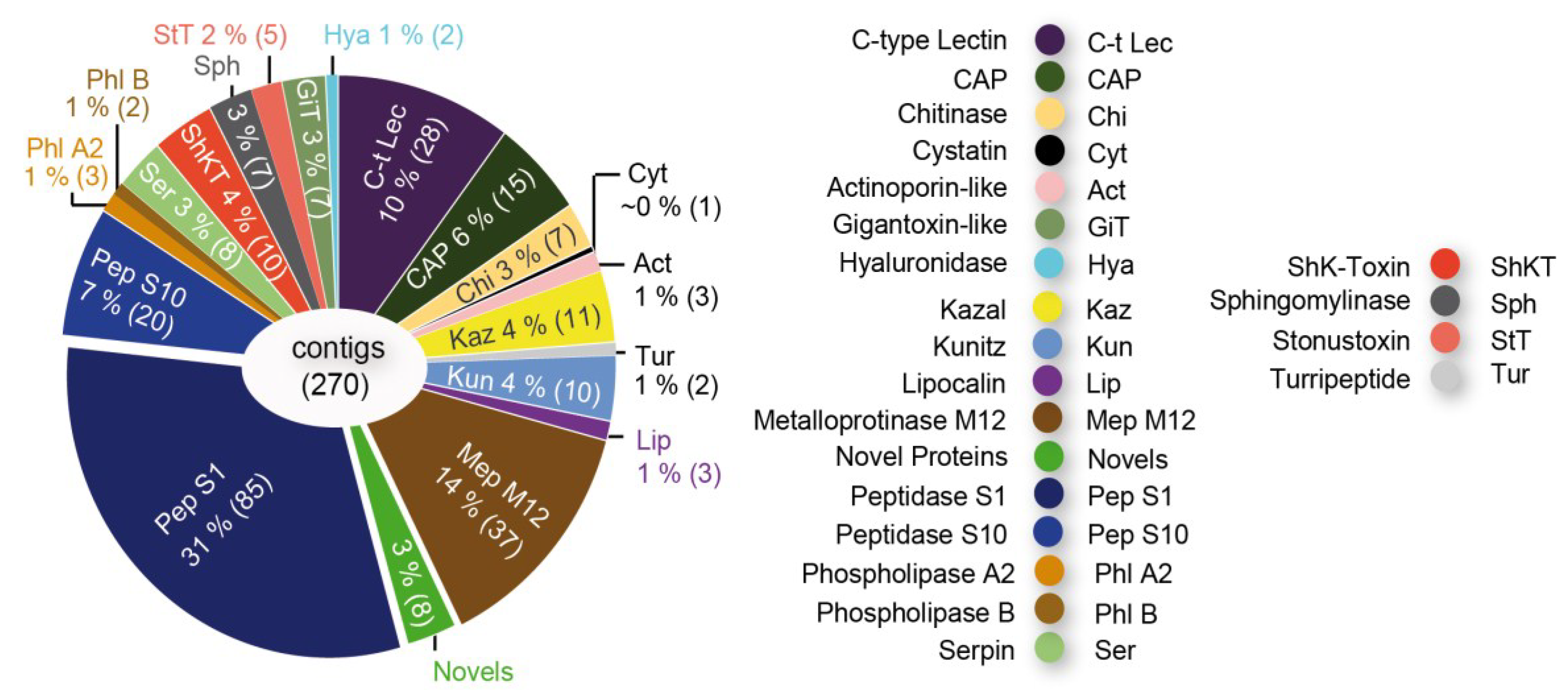
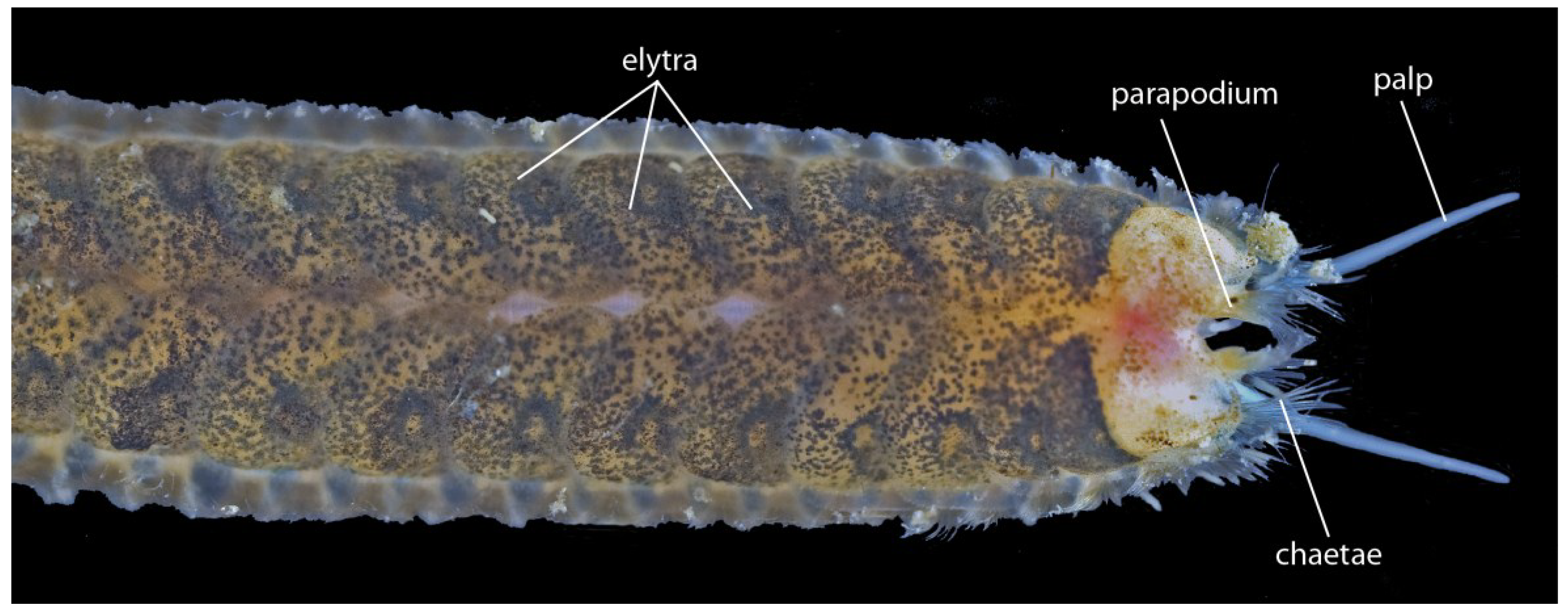
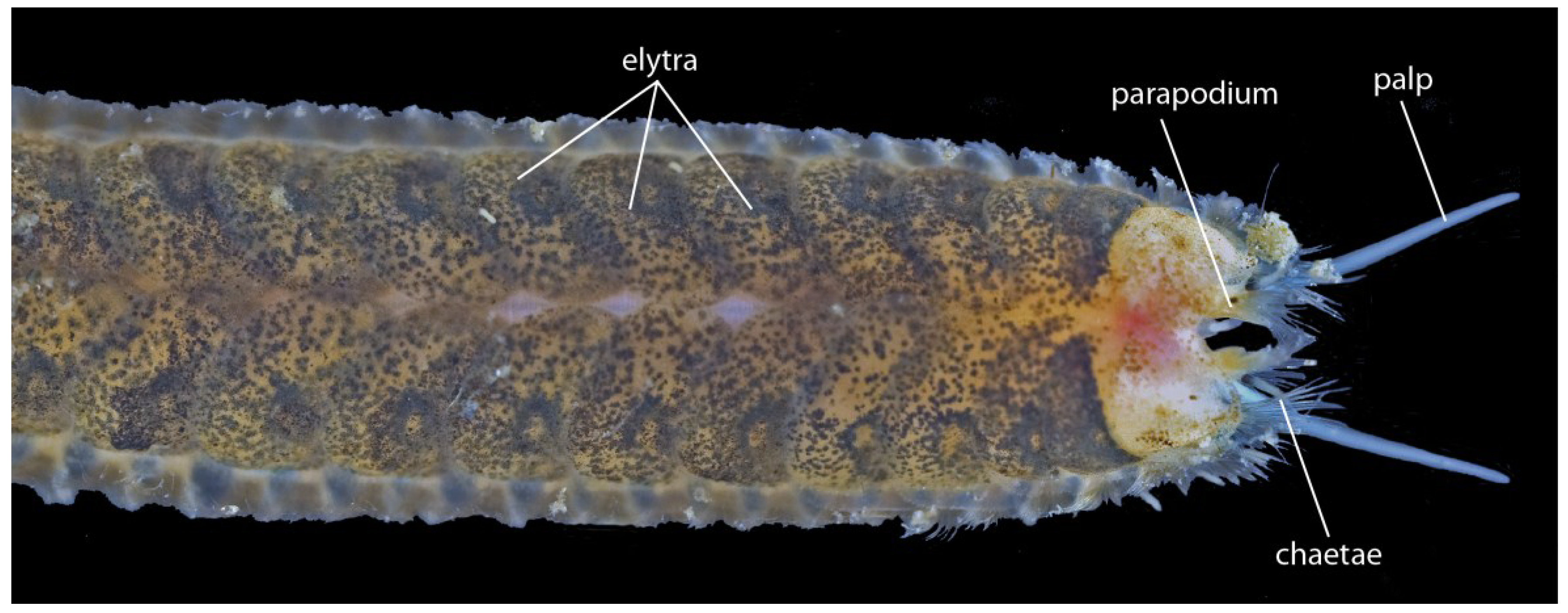
3.1.5. Scale-Worms (Aphroditifomia)
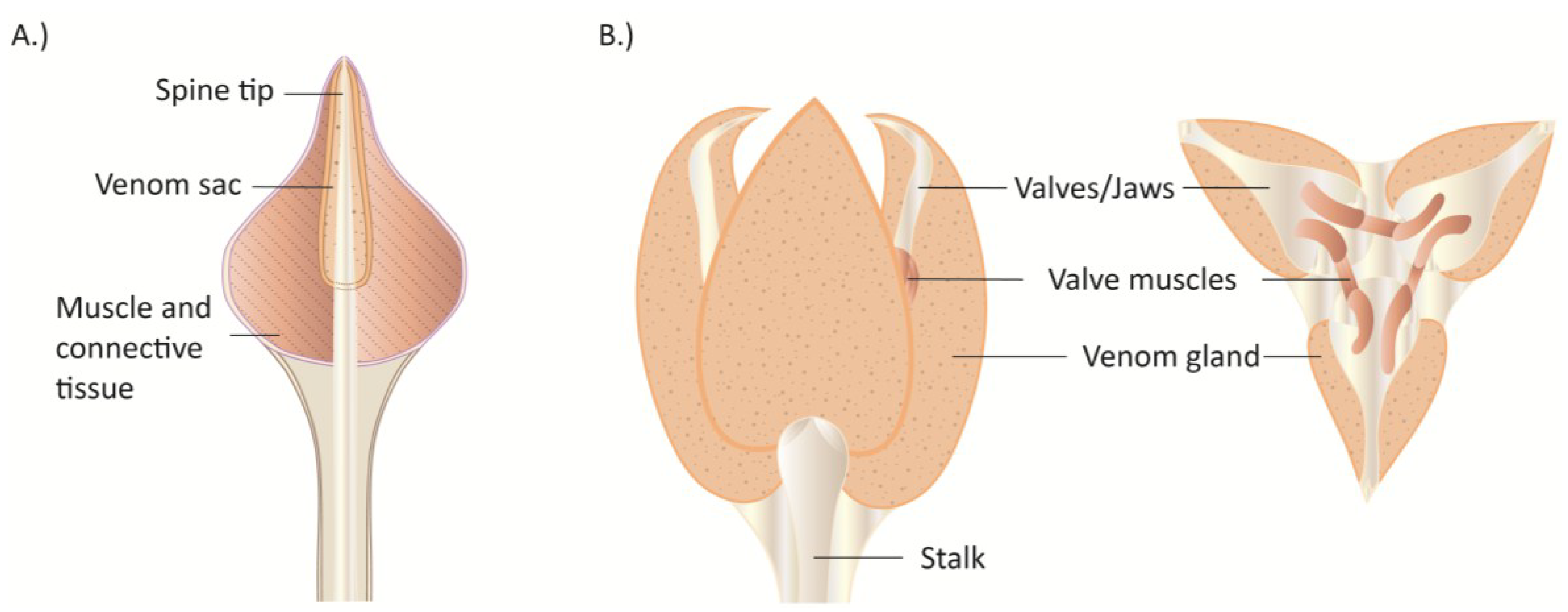
3.2. Flatworms (Platyhelminthes)
3.3. Ribbon Worms (Nemertea)

3.4. Mollusca
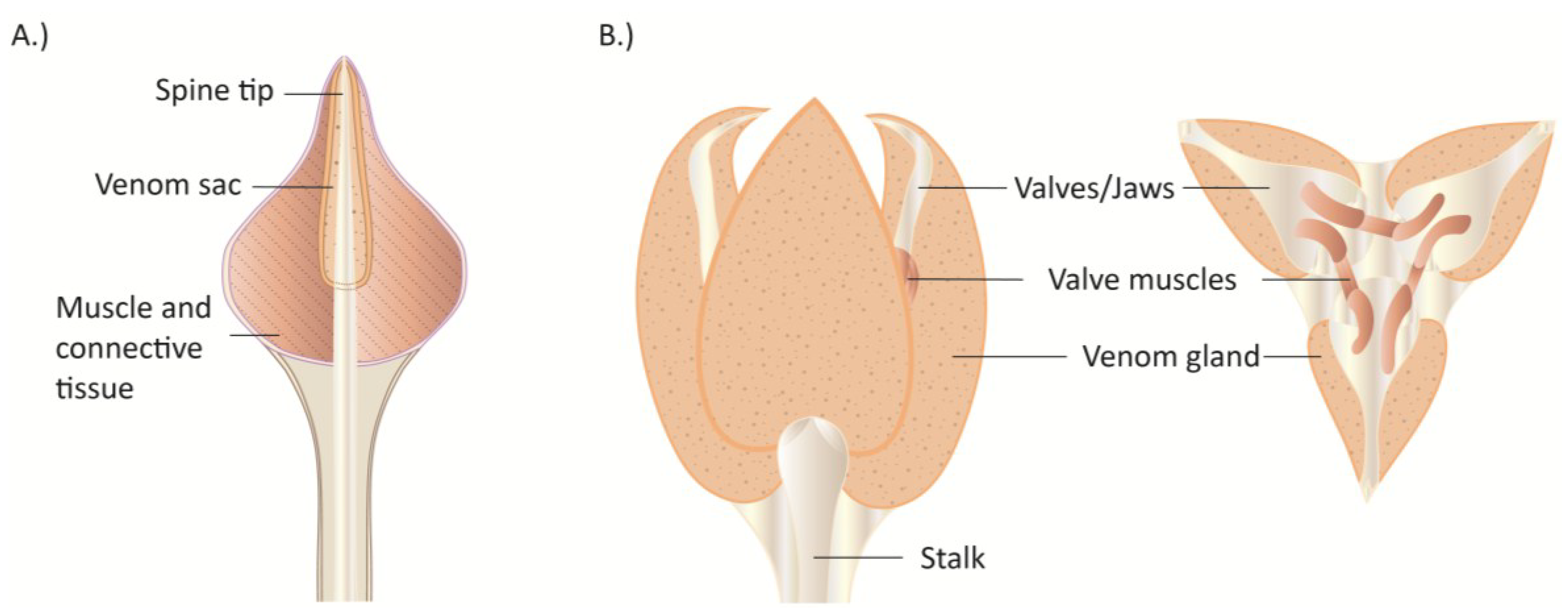
4. Echinoderms (Sea Stars, Sea Urchins)
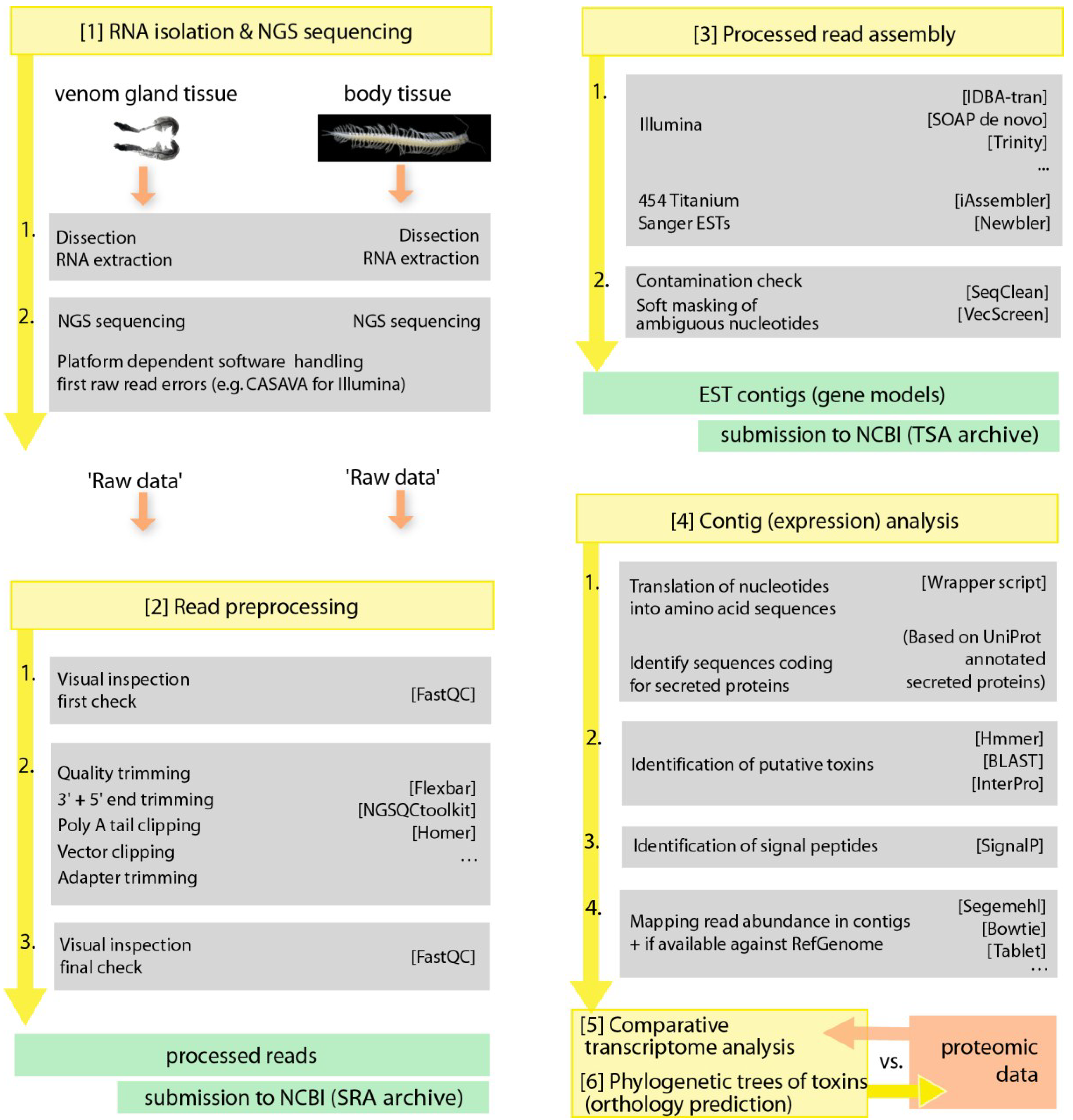
5. Methodological and Future Prospects
5.1. Separating Fact from Fiction
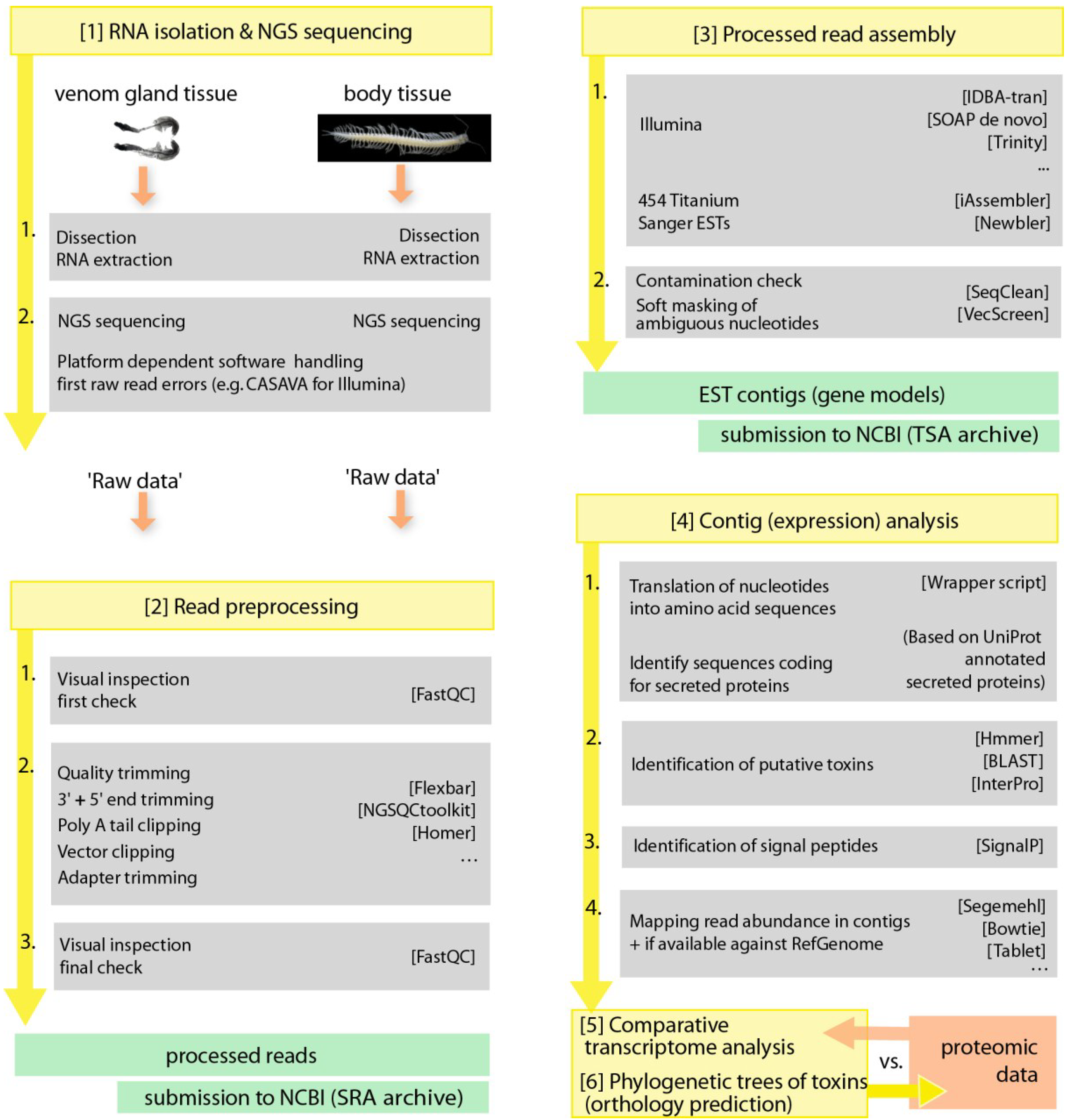
5.2. Book of Venom Revelation—-omics Technology as a Game Changer

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SL_0243 | Protein sequences | Match “toxins” | Match “venom” | Match conotoxin | Match snake venom |
|---|---|---|---|---|---|
| 01.2013 | 53,796 | 3,638 | 332 | 960 | 130 |
| 07.2014 | 85,146 | 4,524 | 528 | 1,515 | 225 |
5.3. The Unique Value of Incorporating Neglected Taxa into Venomics
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Brodie, E.D. Toxins and venoms. Curr. Biol. 2009, 19, R931–R935. [Google Scholar] [CrossRef] [PubMed]
- Calvete, J.J.; Sanz, L.; Angulo, Y.; Lomonte, B.; Gutiérrez, J.M. Venoms, venomics, antivenomics. FEBS Lett. 2009, 583, 1736–1743. [Google Scholar] [CrossRef] [PubMed]
- Casewell, N.R.; Wüster, W.; Vonk, F.J.; Harrison, R.A.; Fry, B.G. Complex cocktails: The evolutionary novelty of venoms. Trends Ecol. Evol. 2012, 28, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Fry, B.G.; Roelants, K.; Champagne, D.E.; Scheib, H.; Tyndall, J.D.A.; King, G.F.; Nevalainen, T.J.; Norman, J.A.; Lewis, R.J.; Norton, R.S.; et al. The toxicogenomic multiverse: Convergent recruitment of proteins into animal venoms. Annu. Rev. Genom. Hum. Genet. 2009, 10, 483–511. [Google Scholar] [CrossRef]
- Dutertre, S.; Jin, A.-H.; Vetter, I.; Hamilton, B.; Sunagar, K.; Lavergne, V.; Dutertre, V.; Fry, B.G.; Antunes, A.; Venter, D.J.; et al. Evolution of separate predation- and defence-evoked venoms in carnivorous cone snails. Nat. Commun. 2014, 5, 1–9. [Google Scholar]
- Vonk, F.J.; Casewell, N.R.; Henkel, C.V.; Heimberg, A.M.; Jansen, H.J.; McCleary, R.J.R.; Kerkkamp, H.M.E.; Vos, R.A.; Guerreiro, I.; Calvete, J.J.; et al. The king cobra genome reveals dynamic gene evolution and adaptation in the snake venom system. Proc. Natl. Acad. Sci. USA 2013, 110, 20651–20656. [Google Scholar] [CrossRef] [PubMed]
- Von Reumont, B.M.; Blanke, A.; Richter, S.; Alvarez, F.; Bleidorn, C.; Jenner, R.A. The first venomous crustacean revealed by transcriptomics and functional morphology: Remipede venom glands express a unique toxin cocktail dominated by enzymes and a neurotoxin. Mol. Biol. Evol. 2014, 31, 48–58. [Google Scholar]
- Undheim, E.A.B.; Jones, A.; Clauser, K.R.; Holland, J.W.; Pineda, S.S.; King, G.F.; Fry, B.G. Clawing through evolution: Toxin diversification and convergence in the ancient lineage Chilopoda (centipedes). Mol. Biol. Evol. 2014, 31, 2124–2148. [Google Scholar] [CrossRef] [PubMed]
- Wong, E.S.W.; Morgenstern, D.; Mofiz, E.; Gombert, S.; Morris, K.M.; Temple-Smith, P.; Renfree, M.B.; Whittington, C.M.; King, G.F.; Warren, W.C.; et al. Proteomics and deep sequencing comparison of seasonally active venom glands in the platypus reveals novel venom peptides and distinct expression profiles. Mol. Cell Proteomics 2012, 11, 1354–1364. [Google Scholar] [CrossRef] [PubMed]
- Undheim, E.A.B.; Sunagar, K.; Hamilton, B.R.; Jones, A.; Venter, D.J.; Fry, B.G.; King, G.F. Multifunctional warheads: Diversification of the toxin arsenal of centipedes via novel multidomain transcripts. J. Proteomics 2014, 102, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Giribet, G.; Edgecombe, G.D. Reevaluating the arthropod tree of life. Annu. Rev. Entomol. 2012, 57, 167–186. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, E.F.; Mourão, C.B.; Moreira, K.G.; Camargos, T.S.; Mortari, M.R. Arthropod venoms: A vast arsenal of insecticidal neuropeptides. Biopolymers 2012, 98, 385–405. [Google Scholar] [CrossRef] [PubMed]
- Edgecombe, G.D. Palaeontological and molecular evidence linking arthropods, onychophorans, and other ecdysozoa. Evol. Educ. Outreach 2009, 2, 178–190. [Google Scholar] [CrossRef]
- Von Reumont, B.M.; Jenner, R.A.; Wills, M.A.; Dell’Ampio, E.; Pass, G.; Ebersberger, I.; Koenemann, S.; Iliffe, T.M.; Stamatakis, A.; Niehuis, O.; et al. Pancrustacean phylogeny in the light of new phylogenomic data: Support for remipedia as the possible sister group of Hexapoda. Mol. Biol. Evol. 2012, 29, 1031–1045. [Google Scholar] [CrossRef] [PubMed]
- King, G.F.; Hardy, M.C. Spider-venom peptides: Structure, pharmacology, and potential for control of insect Pests. Annu. Rev. Entomol. 2013, 58, 475–496. [Google Scholar] [CrossRef] [PubMed]
- Cherniack, E.P.; Cherniack, E.P. Bugs as drugs, part two: Worms, leeches, scorpions, snails, ticks, centipedes, and spiders. Altern. Med. Rev. 2011, 16, 50–58. [Google Scholar] [PubMed]
- Pimenta, A.M.C.; de Lima, M.E. Small peptides, big world: Biotechnological potential in neglected bioactive peptides from arthropod venoms. J. Peptide Sci. 2005, 11, 670–676. [Google Scholar] [CrossRef]
- Moreau, S.J. “‘It stings a bit but it cleans well’”: Venoms of Hymenoptera and their antimicrobial potential. J. Insect. Physiol. 2013, 59, 186–204. [Google Scholar] [CrossRef] [PubMed]
- Anderson, N.L.; Anderson, N.G. Proteome and proteomics: New technologies, new concepts, and new words. Electrophoresis 1998, 19, 1853–1861. [Google Scholar] [CrossRef] [PubMed]
- Blackstock, W.P.; Weir, M.P. Proteomics: Quantitative and physical mapping of cellular proteins. Trends Biotechnol. 1999, 17, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Burki, F.; Burki, F.; Shalchian-Tabrizi, K.; Shalchian-Tabrizi, K.; Pawlowski, J.; Pawlowski, J. Phylogenomics reveals a new “megagroup” including most photosynthetic eukaryotes. Biol. Lett. 2008, 4, 366–369. [Google Scholar] [CrossRef] [PubMed]
- Dunn, C.W.; Hejnol, A.; Matus, D.Q.; Pang, K.; Browne, W.E.; Smith, S.A.; Seaver, E.; Rouse, G.W.; Obst, M.; Edgecombe, G.D.; et al. Broad phylogenomic sampling improves resolution of the animal tree of life. Nature 2008, 452, 745–749. [Google Scholar] [CrossRef] [PubMed]
- James, P. Protein identification in the post-genome era: the rapid rise of proteomics. Q. Rev. Biophys. 1997, 30, 279–331. [Google Scholar] [CrossRef] [PubMed]
- Danneels, E.L.; Gerlo, S.; Heyninck, K.; van Craenenbroeck, K.; de Bosscher, K.; Haegeman, G.; de Graaf, D.C. How the venom from the ectoparasitoid Wasp Nasonia vitripennis exhibits anti-inflammatory properties on mammalian cell lines. PLoS One 2014, 9, e96825. [Google Scholar] [CrossRef] [PubMed]
- de Graaf, D.C.; Aerts, M.; Danneels, E.; Devreese, B. Bee, wasp and ant venomics pave the way for a component-resolved diagnosis of sting allergy. J. Proteomics 2009, 72, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Luengo, O.; Cardona, V. Component resolved diagnosis: When should it be used? Clin. Transl. Allergy 2014, 4, 28. [Google Scholar] [CrossRef] [PubMed]
- Antolín-Amérigo, D.; Moreno Aguilar, C.; Vega, A.; Alvarez-Mon, M. Venom immunotherapy: An updated review. Curr. Allergy Asthma Rep. 2014, 14, 449. [Google Scholar] [CrossRef] [PubMed]
- Etheridge, S.M. Paralytic shellfish poisoning: seafood safety and human health perspectives. Toxicon 2010, 56, 108–122. [Google Scholar] [CrossRef] [PubMed]
- Maillaud, C.; Barguil, Y.; Mikulski, M.; Cheze, M.; Pivert, C.; Deveaux, M.; Lapostolle, F. First successful curative use of digoxin-specific Fab antibody fragments in a life-threatening coconut crab (Birgus latro L.) poisoning. Toxicon 2012, 60, 1013–1017. [Google Scholar] [CrossRef] [PubMed]
- Hoenemann, M.; Neiber, M.T.; Humphreys, W.F.; Iliffe, T.M.; Li, D.; Schram, F.R.; Koenemann, S. Phylogenetic analysis and systematic revision of Remipedia (Nectiopoda) from bayesian analysis of molecular data. J. Crustac. Biol. 2013, 33, 603–619. [Google Scholar] [CrossRef]
- Yager, J. Remipedia, a new class of Crustacea from a marine cave in the Bahamas. J. Crustac. Biol. 1981, 1, 328–333. [Google Scholar] [CrossRef]
- Schram, F.R. Nectiopoda and Enantiopoda. In Crustacea; Oxford University Press: New York, NY, USA, 1986; p. 620. [Google Scholar]
- Ertas, B.; von Reumont, B.M.; Wagele, J.W.; Misof, B.; Burmester, T. Hemocyanin suggests a close relationship of Remipedia and Hexapoda. Mol. Biol. Evol. 2009, 26, 2711–2718. [Google Scholar] [CrossRef] [PubMed]
- Fanenbruck, M.; Harzsch, S.; Wägele, J.-W. The brain of the Remipedia (Crustacea) and an alternative hypothesis on their phylogenetic relationships. Proc. Natl. Acad. Sci. USA 2004, 101, 3868–3873. [Google Scholar] [CrossRef] [PubMed]
- Stemme, T.; Iliffe, T.M.; Bicker, G.; Harzsch, S.; Koenemann, S. Serotonin immunoreactive interneurons in the brain of the Remipedia: New insights into the phylogenetic affinities of an enigmatic crustacean taxon. BMC Evol. Biol. 2012, 12, 168. [Google Scholar] [CrossRef] [PubMed]
- Van der Ham, J.L.; Felgenhauer, B.E. On possible venomous effects of Speleonectes sp (Remipedia). Crustaceana 2007, 80, 755–765. [Google Scholar] [CrossRef]
- Van der Ham, J.L.; Felgenhauer, B.E. The functional morphology of the putative injecting apparatus of Speleonectes Tanumekes (Remipedia). J. Crustac. Biol. 2007, 27, 1–9. [Google Scholar] [CrossRef]
- Billen, B.; Vassilevski, A.; Nikolsky, A.; Debaveye, S.; Tytgat, J.; Grishin, E. Unique bell-shaped voltage-dependent modulation of Na+ channel gating by novel insect-selective toxins from the spider Agelena orientalis. J. Biol. Chem. 2010, 285, 18545–18554. [Google Scholar] [CrossRef] [PubMed]
- Schram, F.R.; Lewis, C.A. Functional morphology of feeding in the Nectiopoda. In Functional morphology of feeding and grooming in Crustacea; Crustacean Issues: Rotterdam, The Netherlands, 1989; Volume 6, pp. 115–122. [Google Scholar]
- Felgenhauer, B.E.; Abele, L.G.; Felder, D.L. Remipedia. In Microscopic anatomy of invertebrates; Crustacea: Wiley-Liss Presss: New York, NY, USA, 1992; Volume 9, pp. 225–247. [Google Scholar]
- Carpenter, J.H. Behavior and ecology of Speleonectes epilimnius (Remipedia, Speleonectidae) from surface water of an anchialine cave on San Salvador Island, Bahamas. Crustaceana 1999, 72, 979–991. [Google Scholar] [CrossRef]
- Koenemann, S.; Schram, F.R.; Iliffe, T.M.; Hinderstein, L.M.; Bloechl, A. Behavior of Remipedia in the laboratory, with supporting field observations. J. Crustac. Biol. 2007, 27, 534–542. [Google Scholar] [CrossRef]
- Møller, O.S. Branchiura (Crustacea)-survey of historical literature and taxonomy. Arthropod. Syst. Phylogeny 2009, 67, 41–55. [Google Scholar]
- Cabiddu, S.; Atzori, G.; Gallo, M.; Cau, A.; Gravina, M.F. Occurence of the alien species Caprella scaura (Amphipoda: Caprellidae) in three Tyrrhenian lagoons. Transit. Water Bull. 2013, 7, 62–71. [Google Scholar]
- Martínez, J.; Adarraga, I. First record of invasive caprellid Caprella scaura Templeton, 1836 sensu lato (Crustacea: Amphipoda: Caprellidae) from the Iberian Peninsula. Aquat. Invasions 2008, 3, 165–171. [Google Scholar] [CrossRef]
- Willis, K.J.; Cook, E.J.; Lozano-Fernandez, M.; Takeuchi, I. First record of the alien caprellid amphipod, Caprella mutica, for the UK. J. Mar. Biol. Assoc. 2004, 84, 1027–1028. [Google Scholar] [CrossRef]
- Møller, O.S.; Olesen, J. The little-known Dipteropeltis hirundo Calman, 1912 (Crustacea, Branchiura): SEM investigations of paratype material in light of recent phylogenetic analyses. Exp. Parasitol. 2010, 125, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Møller, O.S.; Olesen, J.; Avenant-Oldewage, A.; Thomsen, P.F.; Glenner, H.; Glenner, H. First maxillae suction discs in Branchiura (Crustacea): Development and evolution in light of the first molecular phylogeny of Branchiura, Pentastomida, and other “Maxillopoda”. Arthropod Struct. Dev. 2008, 37, 333–346. [Google Scholar] [CrossRef] [PubMed]
- Ider, D.; Ramdane, Z.; Courcot, L.; Amara, R.; Trilles, J.-P. A scanning electron microscopy study of Argulus vittatus (Rafinesque-Schmaltz, 1814) (Crustacea: Branchiura) from Algerian coast. Parasitol. Res. 2014, 113, 2265–2276. [Google Scholar] [CrossRef] [PubMed]
- Claus, C. Über die Entwicklung, Organisation und systematische Stellung der Arguliden. Zeitschrift für Wissenschaftliche Zoologie 1875, 15, 1–68. [Google Scholar]
- Saha, S.K.; Guha, A.; Banerjee, A. Feeding apparatus and associated glands in the freshwater fish ectoparasite Argulus Siamensis Wilson, 1926 (Branchiura). Crustaceana 2011, 84, 1153–1168. [Google Scholar] [CrossRef]
- Swanepoel, J.H.; Avenant-Oldewage, A. Comments on the morphology of the pre-oral spine in Argulus (Crustacea: Branchiura). J. Morphol. 1992, 212, 155–162. [Google Scholar] [CrossRef]
- Gresty, K.A.; Boxshall, G.A.; Nagasawa, K. The fine structure and function of the cephalic appendages of the branchiuran parasite, Argulus japonicus Thiele. Philos. Trans. R. Soc. Lond., B, Biol. Sci. 1993, 339, 119–135. [Google Scholar] [CrossRef]
- Wilson, B.C. North American parasitic copepods of the family Argulidae: With a bibliography of the group and a systematic review of all known species. Proc. U. S. Natl. Mus. 1902, 25, 635–742. [Google Scholar] [CrossRef]
- Madsen, N. The anatomy of Argulus foliaceus Linnè: With notes on Argulus coregoni Thorell and Argulus africanus Thiele. Part I. Integument, central nervous system, sense organs, pre-oral spine and digestive organs. In Kungliga Fysiografiska Sälkabets Handligar N.F: Lunds universitets arsskrift; University of Lund: Lund, Sweden, 1964; pp. 1–32. [Google Scholar]
- Martin, J.W.; Davis, G.E. An updated classification of the recent Crustacea; Science Series Natural History Museum of Los Angeles County; Natural History Museum of Los Angeles County: Los Angeles, CA, USA, 2001; Volume 39. [Google Scholar]
- Caine, E.A. Reproductive behavior and sexual dimorphism of a caprellid amphipod. J. Crust. Biol. 1991, 11, 56. [Google Scholar] [CrossRef]
- Boos, K.; Ashton, G.V.; Cook, E.J. The Japanese skeleton shrimp Caprella mutica (Crustacea, Amphipoda): A global invader of coastal waters. the Wrong Place-Alien Marine Crustaceans: Distribution, Biology and Impacts 2011, 6, 129–156. [Google Scholar]
- Schulz, M.K.; Alexander, C.G. Aggressive behaviour of Caprella scaura typicamayer, 1890. (Crustacea: Amphipoda). Mar. Fresh. Behav. Physiol. 2001, 34, 181–187. [Google Scholar] [CrossRef]
- Takeshita, F.; Wada, S. Morphological comparison of the second gnathopod in males of four caprellid species (Amphipoda: Caprellidae). J. Crust. Biol. 2012, 32, 673–676. [Google Scholar] [CrossRef]
- Lewbel, G.S. Sexual dimorphism and intraspecific aggression, and their relationship to sex ratios in Caprella gorgonia Laubitz & Lewbel (Crustacea: Amphipoda: Caprellidae). J. Exp. Mar. Biol. Ecol. 1978, 33, 133–151. [Google Scholar] [CrossRef]
- Nekaris, K.; Moore, R.; Rode, E.; Fry, B. Mad, bad and dangerous to know: The biochemistry, ecology and evolution of slow loris venom. J. Venom Anim. Toxins Incl. Trop. Dis. 2013, 19, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Nishida, S.; Ohtsuka, S. Specialized feeding mechanism in the pelagic copepod genus Heterorhabdus (Calanoida: Heterorhabdidae), with special reference to the mandibular tooth and labral glands. Mar. Biol. 1996, 126, 619–632. [Google Scholar] [CrossRef]
- Ohtsuka, S.; Soh, H.Y.; Nishida, S. Evolutionary switching from suspension feeding to carnivory in the calanoid family Heterorhabdidae (Copepoda). J. Crust. Biol. 1997, 17, 577–595. [Google Scholar] [CrossRef]
- Johnson, S.C.; Albright, L.J. Comparative susceptibility and histopathology of the response of naive Atlantic, chinook and coho salmon to experimental infection with Lepeophtheirus salmonis (Copepoda: Caligidae). Dis. Aquat. Org. 1992, 14, 179–193. [Google Scholar] [CrossRef]
- Fast, M.D.; Johnson, S.C.; Eddy, T.D.; Pinto, D.; Ross, N.W. Lepeophtheirus salmonis secretory/excretory products and their effects on Atlantic salmon immune gene regulation. Parasite Immunol. 2007, 29, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Nelsen, D.R.; Nisani, Z.; Cooper, A.M.; Fox, G.A.; Gren, E.C.K.; Corbit, A.G.; Hayes, W.K. Poisons, toxungens, and venoms: Redefining and classifying toxic biological secretions and the organisms that employ them. Biol. Rev. 2014, 89, 450–465. [Google Scholar] [CrossRef] [PubMed]
- Manship, B.M.; Walker, A.J.; Jones, L.A.; Davies, A.J. Blood feeding in juvenile Paragnathia formica (Isopoda: Gnathiidae): Biochemical characterization of trypsin inhibitors, detection of anticoagulants, and molecular identification of fish hosts. Parasitology 2012, 139, 744–754. [Google Scholar] [CrossRef] [PubMed]
- Lambkin, C.; Sinclair, B.J.; Pape, T.; Courtney, G.W.; Skevington, J.H.; Meier, R.; Yeates, D.K.; Blagoderov, V.; Wiegmann, B.M. The phylogenetic relationships among infraorders and superfamilies of Diptera based on morphological evidence. Syst. Entomol. 2012, 38, 164–179. [Google Scholar] [CrossRef]
- Yeates, D.K.; Wiegmann, B.M.; Courtney, G.W.; Meier, R.; Lambkin, C.; Pape, T. Phylogeny and systematics of Diptera: Two decades of progress and prospects. Zootaxa 2007, 1668, 565–590. [Google Scholar]
- Ribeiro, J.; Mans, B.J.; Arcà, B. An insight into the sialome of blood-feeding Nematocera. Insect Biochem. Mol. Biol. 2010, 40, 767–784. [Google Scholar] [CrossRef] [PubMed]
- Dennis, D.S.; Barnes, J.K.; Knutson, L. Review and analysis of information on the biology and morphology of immature stages of robber flies (Diptera: Asilidae). Zootaxa 2013, 3673, 1–64. [Google Scholar] [CrossRef]
- Dikow, T.; Grimaldi, D.A. Robber flies in Cretaceous ambers (Insecta, Diptera, Asilidae). Am. Mus. Novit. 2014, 3799, 1–19. [Google Scholar] [CrossRef]
- Bybee, S.M.; Taylor, S.D.; Riley Nelson, C.; Whiting, M.F. A phylogeny of robber flies (Diptera: Asilidae) at the subfamilial level: Molecular evidence. Mol. Phylogenet. Evol. 2004, 30, 789–797. [Google Scholar] [CrossRef] [PubMed]
- Melin, D.E. Contributions to the Knowledge of the Biology, Metamorphosis and Distribution of the Swedish Asilids in Relation to the Whole famIly of Asilids; Almqvist & Wiksells: Stockholm, Sweden, 1923. [Google Scholar]
- Dennis, D.S. Ethology of Proctacanthus. brevipennis (Wiedemann, 1828) (Diptera: Asilidae) in Northeastern Florida, USA. J. Entomol. Res. Soc. 2012, 14, 91–109. [Google Scholar]
- Whitfield, F.G. The Relation between the Feeding-habits and the Structure of the Month-parts in the Asilidæ (Diptera). Proc. Zool. Soc. Lond. 1925, 95, 599–638. [Google Scholar] [CrossRef]
- Bromley, S.W. Ohio robber flies IV (Diptera: Asilidae). Ohio. J. Sci. 1947, 47, 67–68. [Google Scholar]
- Morse, R.A.; Flottum, K. Honey Bee Pests, Predators, and Diseases; Wicwas Press: Kalamazoo, MI, USA, 1997. [Google Scholar]
- Owsley, W.B. The comparative morphology of internal structures of the Asilidae (Diptera). Ann. Entomol. Soc. Am. 1946, 39, 33–68. [Google Scholar]
- Kahan, D. The toxic effect of the bite and the proteolytic activity of the saliva and stomach contents of the robber flies (Diptera Asilidae). Isr. J. Zool. 1964, 13, 47–57. [Google Scholar]
- Musso, J.J. Observations sur le comortement alimentaires l’anatomie e l’histologie des glandes salivaires de deux asilides mediterraneens. Ann. Soc. Entomol. Fr. 1968, 4, 245–255. [Google Scholar]
- Musso, J.J.; Garnier, R.; Legier, F. Comparison of toxicity of venom of some asilids (Diptera Brachycera) on locusts. Ann. Soc. Entomol. Fr. 1978, 14, 177–184. [Google Scholar]
- Mackessy, S.P. Morphology and ultrastructure of the venom glands of the northern pacific rattlesnake Crotalus viridis oreganus. J. Morphol. 1991, 208, 109–128. [Google Scholar] [CrossRef]
- Mackessy, S.P.; Baxter, L.M. Bioweapons synthesis and storage: The venom gland of front-fanged snakes. Zool. Anz. 2006, 245, 147–159. [Google Scholar] [CrossRef]
- Schmidt, J.O. Biochemistry of insect venoms. Annu. Rev. Entomol. 1982, 27, 339–368. [Google Scholar] [CrossRef] [PubMed]
- Baldacchino, F.; Desquesnes, M.; Mihok, S.; Foil, L.D.; Duvallet, G.; Jittapalapong, S. Tabanids: Neglected subjects of research, but important vectors of disease agents! Infect. Genet. Evol. 2014. [Google Scholar] [CrossRef]
- Lessard, B.D.; Cameron, S.L.; Bayless, K.M.; Wiegmann, B.M.; Yeates, D.K. The evolution and biogeography of the austral horse fly tribe Scionini (Diptera: Tabanidae: Pangoniinae) inferred from multiple mitochondrial and nuclear genes. Mol. Phylogenet. Evol. 2013, 68, 516–540. [Google Scholar] [CrossRef] [PubMed]
- Wiegmann, B.M.; Trautwein, M.D.; Winkler, I.S.; Barr, N.B.; Kim, J.-W.; Lambkin, C.; Bertone, M.A.; Cassel, B.K.; Bayless, K.M.; Heimberg, A.M.; et al. Episodic radiations in the fly tree of life. Proc. Natl. Acad. Sci. U.S.A. 2011, 108, 5690–5695. [Google Scholar] [CrossRef] [PubMed]
- Beard, R.L. Insect toxins and venoms. Annu. Rev. Entomol. 1963, 8, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Philip, C.B. The Tabanidae (horseflies) of Minnesota; University Farm: Cambridge, UK, 1931. [Google Scholar]
- Jackman, R.; Nowicki, S.; Aneshansley, D.J.; Eisner, T. Predatory capture of toads by fly larvae. Science 1983, 222, 515–516. [Google Scholar] [CrossRef] [PubMed]
- Nowicki, S.; Eisner, T. Predatory Capture of Bombardier Beetles by a Tabanid Fly Larva. Psyche J. Entom. 1983, 90, 119–122. [Google Scholar] [CrossRef]
- Otsuru, M.; Ogawa, S. Observations on the bite of horsefly larvae in the paddy-fields. Eisei Dobutsu 1959, 10, 27–34. [Google Scholar]
- BugGuide. Available online: http://www.bugguide.net (accessed on 10 December 2014).
- Trelka, D.G.; Berg, C.O. Behavioral studies of the slug-killing larvae of two species of Tetanocera (Diptera: Sciomyzidae). Proc. Entomol. Soc. Wash. 1977, 79, 475–486. [Google Scholar]
- Berg, C.O.; Knutson, L. Biology and systematics of the Sciomyzidae. Annu. Rev. Entomol. 1978, 23, 239–258. [Google Scholar] [CrossRef]
- Chapman, E.G.; Przhiboro, A.A.; Harwood, J.D.; Foote, B.A.; Hoeh, W.R. Widespread and persistent invasions of terrestrial habitats coincident with larval feeding behavior transitions during snail-killing fly evolution (Diptera: Sciomyzidae). BMC. Evol. Biol. 2012, 12, 175. [Google Scholar] [CrossRef] [PubMed]
- Foote, B.A. Biology and immature stages of snail-killing flies belonging to the genus Tetanocera (Diptera: Sciomyzidae). IV. Life histories of predators of land snails and slugs. Ann. Carnegie Mus. 2008, 77, 301–312. [Google Scholar] [CrossRef]
- Murphy, W.L.; Knutson, L.V.; Chapman, E.G.; Donnell, R.J.M.; Williams, C.D.; Foote, B.A.; Vala, J.-C. Key aspects of the biology of snail-killing Sciomyzidae flies. Annu. Rev. Entomol. 2012, 57, 425–447. [Google Scholar] [CrossRef] [PubMed]
- Vala, J.C.; Murphy, W.L.; Knutson, L. A cornucopia for Sciomyzidae (Diptera). Stud. Dipterol. 2012, 19, 67–137. [Google Scholar]
- Berg, C.O. Sciomyzid larvae (Diptera) that feed on snails. J. Parasitol. 1953, 39, 630–636. [Google Scholar] [CrossRef] [PubMed]
- Catterall, W.A. Neurotoxins that act on voltage-sensitive sodium channels in excitable membranes. Annu. Rev. Pharmacol. Toxicol. 1980, 20, 15–43. [Google Scholar] [CrossRef] [PubMed]
- Howard, B.D.; Gundersen, C.B. Effects and mechanisms of polypeptide neurotoxins that act presynaptically. Annu. Rev. Pharmacol. Toxicol. 1980, 20, 307–336. [Google Scholar] [CrossRef] [PubMed]
- Giordani, I.; Hynes, T.; Reich, I.; Donnell, R.M. Tetanocera elata (Diptera: Sciomyzidae) larvae feed on protected slug species Geomalacus maculosus (Gastropoda: Arionidae): First record of predation. J. Insect. Behav. 2014, 27, 652–656. [Google Scholar] [CrossRef]
- Donnell, R.J.M.; Paine, T.D.; Mulkeen, C.J.; Gormally, M.J. Effects of temperature and prey availability on the malacophagous larval stage of Sepedon spinipes (Scopoli) (Diptera: Sciomyzidae): Potential biocontrol for gastropod vectors of parasitic diseases. Bio. Control. 2014, 70, 42–47. [Google Scholar] [CrossRef]
- Hynes, T.; Giordani, I.; Larkin, M.; McDonnell, R.J.; Gormally, M. Larval feeding behaviour of Tetanocera elata (Diptera: Sciomyzidae): Potential biocontrol agent of pestiferous slugs. Biocontrol. Sci. Techn. 2014, 24, 1–11. [Google Scholar] [CrossRef]
- Mayr, L. Untersuchungen zur Funktion der Speicheldrüsen räuberischer Gallmückenlarven (Aphidoletes aphidimyza Rond.). Z. Ang. Ent. 1975, 77, 270–273. [Google Scholar] [CrossRef]
- Dugon, M.M.; Arthur, W. Comparative studies on the structure and development of the venom-delivery system of centipedes, and a hypothesis on the origin of this evolutionary novelty. Evol. Dev. 2012, 14, 128–137. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Xiao, Y.; Kang, D.; Liu, J.; Li, Y.; Undheim, E.A.B.; Klint, J.K.; Rong, M.; Lai, R.; King, G.F. Discovery of a selective NaV1.7 inhibitor from centipede venom with analgesic efficacy exceeding morphine in rodent pain models. Proc. Natl. Acad. Sci. USA 2013, 110, 17534–17539. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.-Q. Animal Biodiversity: An Outline of Higher-Level Classification and Survey of Taxonomic Richness; Magnolia press: Auckland, New Zealand, 2011. [Google Scholar]
- Undheim, E.A.B.; King, G.F. On the venom system of centipedes (Chilopoda), a neglected group of venomous animals. Toxicon 2011, 57, 512–524. [Google Scholar] [CrossRef] [PubMed]
- Chippaux, J.-P.; Goyffon, M. Epidemiology of scorpionism: A global appraisal. Acta Trop. 2008, 107, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Antoniazzi, M.M.; Pedroso, C.M.; Knysak, I.; Martins, R.; Guizze, S.P.; Jared, C.; Barbaro, K.C. Comparative morphological study of the venom glands of the centipede Cryptops iheringi, Otostigmus pradoiand Scolopendra viridicornis. Toxicon 2009, 53, 367–374. [Google Scholar] [CrossRef] [PubMed]
- Haug, J.T.; Haug, C.; Schweigert, G.; Sombke, A. The evolution of centipede venom claws—Open questions and possible answers. Arthropod Struct. Dev. 2014, 43, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Shear, W.A.; Edgecombe, G.D. The geological record and phylogeny of the Myriapoda. Arthropod Struct. Dev. 2010, 39, 174–190. [Google Scholar] [CrossRef] [PubMed]
- Dugon, M.M.; Hayden, L.; Black, A.; Arthur, W. Development of the venom ducts in the centipede Scolopendra: An example of recapitulation. Evol. Dev. 2012, 14, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Dugon, M.M.; Black, A.; Arthur, W. Variation and specialisation of the forcipular apparatus of centipedes (Arthropoda: Chilopoda): A comparative morphometric and microscopic investigation of an evolutionary novelty. Arthropod Struct. Dev. 2012, 41, 231–243. [Google Scholar] [CrossRef] [PubMed]
- Dugon, M.M.; Arthur, W. Prey orientation and the role of venom availability in the predatory behaviour of the centipede Scolopendra subspinipes mutilans (Arthropoda: Chilopoda). J. Insect. Physiol. 2012, 58, 874–880. [Google Scholar] [CrossRef] [PubMed]
- Rates, B.; Bemquerer, M.P.; Richardson, M.; Borges, M.H.; Morales, R.A.; de Lima, M.E.; Pimenta, A.M. Venomic analyses of Scolopendra viridicornis nigraand Scolopendra angulata (Centipede, Scolopendromorpha): Shedding light on venoms from a neglected group. Toxicon 2007, 49, 810–826. [Google Scholar] [CrossRef] [PubMed]
- Malta, M.B.; Lira, M.S.; Soares, S.L.; Rocha, G.C.; Knysak, I.; Martins, R.; Guizze, S.P.G.; Santoro, M.L.; Barbaro, K.C. Toxic activities of Brazilian centipede venoms. Toxicon 2008, 52, 255–263. [Google Scholar] [CrossRef] [PubMed]
- GonzAlez-Morales, L.; Diego-García, E.; Segovia, L.; del Carmen GutiErrez, M.; Possani, L.D. Venom from the centipede Scolopendra viridis Say: Purification, gene cloning and phylogenetic analysis of a phospholipase A2. Toxicon 2009, 54, 8–15. [Google Scholar] [CrossRef] [PubMed]
- González-Moralesa, L.; Pedraza-Escalonaa, M.; Diego-Garciaa, E.; Restano-Cassulinia, R.; Batistaa, C.V.F.; del Carmen Gutiérrezb, M.; Possania, L.D. Proteomic characterization of the venom and transcriptomic analysis of the venomous gland from the Mexican centipede Scolopendra viridis. J. Proteomics 2014, 111, 224–237. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Liu, Z.; Xiao, Y.; Li, Y.; Rong, M.; Liang, S.; Zhang, Z.; Yu, H.; King, G.F.; Lai, R. Chemical punch packed in venoms makes centipedes excellent predators. Mol. Cell Proteomics 2012, 11, 640–650. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.-C.; Zhang, R.; Zhao, F.; Chen, Z.-M.; Liu, H.-W.; Wang, Y.-J.; Jiang, P.; Zhang, Y.; Wu, Y.; Ding, J.-P.; Lee, W.-H.; Zhang, Y. Venomic and transcriptomic analysis of centipede Scolopendra subspinipes dehaani. J. Proteome Res. 2012, 11, 6197–6212. [Google Scholar] [PubMed]
- Cabezas-Cruz, A.; Valdés, J.J. Are ticks venomous animals? Front. Zool. 2014, 11, 47. [Google Scholar] [CrossRef] [PubMed]
- Kuhn-Nentwig, L.; Stöcklin, R.; Nentwig, W. Venom composition and strategies in spiders. Adv.In Insect Phys. 2011, 40, 1–86. [Google Scholar]
- Goyffon, M.; Tournier, J.-N. Scorpions: A presentation. Toxins 2014, 6, 2137–2148. [Google Scholar] [CrossRef] [PubMed]
- Harvey, M. Order Pseudoscorpiones. Zootaxa 2013, 3703, 34–35. [Google Scholar] [CrossRef]
- Weygoldt, P. The Biology of Pseudoscorpions; Harvard University Press: Cambridge, MA, USA, 1969. [Google Scholar]
- Murienne, J.; Harvey, M.S.; Giribet, G. First molecular phylogeny of the major clades of Pseudoscorpiones (Arthropoda: Chelicerata). Mol. Phylogenet. Evol. 2008, 49, 170–184. [Google Scholar] [CrossRef] [PubMed]
- Fagan, L.L.; Nelson, W.R.; Meenken, E.D.; Howlett, B.G.; Walker, M.K.; Donovan, B.J. Varroa management in small bites. J. Appl. Entomol. 2012, 136, 473–475. [Google Scholar] [CrossRef]
- Harvey, M.S. A review and redescription of the cosmopolitan pseudoscorpion Chelifer cancroides (Pseudoscorpiones: Cheliferidae). J. Arachnol. 2014, 42, 86–104. [Google Scholar] [CrossRef]
- Harvey, M.S. The phylogeny and classification of the Pseudoscorpionida (Chelicerata: Arachnida). Invertebr. Syst. 1992, 6, 1373–1435. [Google Scholar] [CrossRef]
- Turk, F.A. A new genus and species of pseudoscorpion with some notes on its biology. Proc. Zool. Soc. Lond. 1952, 122, 951–954. [Google Scholar] [CrossRef]
- Read, S.; Howlett, B.G.; Donovan, B.J.; Nelson, W.R.; van Toor, R.F. Culturing chelifers (Pseudoscorpions) that consume Varroa mites. J. Appl. Entomol. 2014, 138, 260–266. [Google Scholar] [CrossRef]
- Brach, V. Social behavior in the pseudoscorpion Paratemnus elongatus (Banks) (Pseudoscorpionida: Atemnidae). Insectes Soc. 1978, 25, 3–11. [Google Scholar] [CrossRef]
- Zeh, J.; Zeh, D. Cooperative foraging for large prey by Paratemnus elongatus (Pseudoscorpionida, Atemnidae). J. Arachnol. 1990, 18, 307–311. [Google Scholar]
- Dos Santos, W.F.; Coutinho-Netto, J. Effects of the Paratemnus elongatus pseudoscorpion venom in the uptake and binding of the l-glutamate and GABA from rat cerebral cortex. J. Biochem. Mol. Toxicol. 2006, 20, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Aruchami, M.; Sundara Rajulu, G. An investigation on the poison glands and the nature of the venom of Rhagodes nicrocinctus (Solifugae: Arachnida). Natl. Acad. Sci. Lett. 1978, 1, 191–192. [Google Scholar]
- Van der Ham, J.L.; Felgenhauer, B.E. Ultrastructure and functional morphology of glandular setae and distal claws of cephalic appendages of Speleonectes tanumekes (Crustacea: Remipedia). Arthropod Struct. Dev. 2008, 37, 235–247. [Google Scholar] [CrossRef] [PubMed]
- Van der Meijden, A.; Langer, F.; Boistel, R.; Vagovic, P.; Heethoff, M. Functional morphology and bite performance of raptorial chelicerae of camel spiders (Solifugae). J. Exp. Biol. 2012, 215, 3411–3418. [Google Scholar] [CrossRef] [PubMed]
- Philippe, H. Multigene analyses of bilaterian animals corroborate the monophyly of ecdysozoa, lophotrochozoa, and protostomia. Mol. Biol. Evol. 2005, 22, 1246–1253. [Google Scholar] [CrossRef] [PubMed]
- Erwin, D.H.; Laflamme, M.; Tweedt, S.M.; Sperling, E.A.; Pisani, D.; Peterson, K.J. The cambrian conundrum: Early divergence and later ecological success in the early history of animals. Science 2011, 334, 1091–1097. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, C. Animal Evolution: Interrelationships of the Living Phyla; Oxford University Press: Oxford, UK, 2012. [Google Scholar]
- Weigert, A.; Helm, C.; Meyer, M.; Nickel, B.; Arendt, D.; Hausdorf, B.; Santos, S.R.; Halanych, K.M.; Purschke, G.; Bleidorn, C.; et al. Illuminating the base of the annelid tree using transcriptomics. Mol. Biol. Evol. 2014, 31, 1391–1401. [Google Scholar] [CrossRef] [PubMed]
- Fauchald, K.; Jumars, P.A. The diet of worms: a study of polychaete feeding guilds. Oceanogr. Mari. Biol. Ann. Rev. 1979, 17, 193–284. [Google Scholar]
- Darwin, C. The Formation of Vegetable Mould, Through the Action of Worms, with Observations on Their Habits; J. Murray: Stevens Point, WI, USA, 1892. [Google Scholar]
- Elliott, J.M.; Kutschera, U. Medicinal leeches: Historical use, ecology, genetics and conservation. Freshw. Rev. 2011, 4, 21–41. [Google Scholar] [CrossRef]
- Kvist, S.; Min, G.-S.; Siddall, M.E. Diversity and selective pressures of anticoagulants in three medicinal leeches (Hirudinida: Hirudinidae, Macrobdellidae). Ecol. Evol. 2013, 3, 918–933. [Google Scholar] [CrossRef] [PubMed]
- Klawe, W.L.; Dickie, L.M. Biology of the Bloodworm, Glycera dibranchiata Ehlers, and Its Relation to the Bloodworm Fishery of the Maritime Provinces; Fisheries Research Board of Canada: Toronto, ON, Canada, 1957. [Google Scholar]
- Basanova, A.V.; Baskova, I.P.; Zavalova, L.L. Vascular-platelet and plasma hemostasis regulators from bloodsucking animals. Biochemistry Mosc. 2002, 67, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Freiberg, M.; Walls, J. The World of Venomous Animals; T.F.H. Publications Inc.: Neptune city, NJ, USA, 1984. [Google Scholar]
- Wolf, P.S. A New genus and species of interstitial sigalionidae and a report on the presence of venom glands in some scale worm families (Annelida, Polychaeta). Proc. Biol. Soc. Wash. 1986, 99, 79–83. [Google Scholar]
- Von Reumont, B.M.; Campbell, L.I.; Richter, S.; Hering, L.; Sykes, D.; Hetmank, J.; Jenner, R.A.; Bleidorn, C. A polychaete’s powerful punch: Venom gland transcriptomics of Glycera reveals a complex cocktail of toxin homologs. Genome Biol. Evol. 2014, 6, 2406–2423. [Google Scholar] [CrossRef] [PubMed]
- Borda, E.; Oceguera-Figueroa, A.; Siddall, M.E. On the classification, evolution and biogeography of terrestrial haemadipsoid leeches (Hirudinida: Arhynchobdellida: Hirudiniformes). Mol. Phylogenet. Evol. 2008, 46, 142–154. [Google Scholar] [CrossRef] [PubMed]
- Struck, T.H.; Schult, N.; Kusen, T.; Hickman, E.; Bleidorn, C.; McHugh, D.; Halanych, K.M. Annelid phylogeny and the status of Sipuncula and Echiura. BMC. Evol. Biol. 2007, 7, 57. [Google Scholar] [CrossRef] [PubMed]
- Sawyer, R.T. Leech Biology and Behaviour. Vol. I Anatomy, Physiology, and Behaviour; Clarendon Press: Oxford, UK, 1986. [Google Scholar]
- Wilkin, P.J.; Scofield, A.M. Growth of the medicinal leech, Himudo medicinalis, under natural and laboratory conditions. Freshw. Biol. 1991, 25, 547–553. [Google Scholar] [CrossRef]
- Lemke, S.; Müller, C.; Lipke, E.; Uhl, G.; Hildebrandt, J.-P. May salivary gland secretory proteins from hematophagous leeches (Hirudo verbana) reach pharmacologically relevant concentrations in the vertebrate host? PLoS One 2013, 8, e73809. [Google Scholar] [CrossRef] [PubMed]
- Sawyer, R.T. Leech Biology and Behaviour. Vol. II Feeding Biology, Ecology, and Systematics; Clarendon Press: Oxford, UK, 1986. [Google Scholar]
- Borda, E.; Siddall, M.E. Arhynchobdellida (Annelida: Oligochaeta: Hirudinida): Phylogenetic relationships and evolution. Mol. Phylogenet. Evol. 2004, 30, 213–225. [Google Scholar] [CrossRef] [PubMed]
- Phillips, A.J.; Siddall, M.E. Poly-paraphyly of Hirudinidae: Many lineages of medicinal leeches. BMC. Evol. Biol. 2009, 9, 246. [Google Scholar] [CrossRef] [PubMed]
- Jansson, I.-M.; McLoughlin, S.; Vajda, V. Early Jurassic annelid cocoons from eastern Australia. Alcheringa 2008, 32, 285–296. [Google Scholar] [CrossRef]
- Min, G.-S.; Sarkar, I.N.; Siddall, M.E. Salivary transcriptome of the North American medicinal leech, Macrobdella decora. J. Parasitol. 2010, 96, 1211–1221. [Google Scholar] [CrossRef] [PubMed]
- Kvist, S.; Brugler, M.R.; Goh, T.G.; Giribet, G.; Siddall, M.E. Pyrosequencing the salivary transcriptome of Haemadipsa interrupta (Annelida: Clitellata: Haemadipsidae): Anticoagulant diversity and insight into the evolution of anticoagulation capabilities in leeches. Invertebr. Biol. 2013, 133, 74–98. [Google Scholar] [CrossRef]
- Kvist, S.; Sarkar, I.N.; Siddall, M.E. Genome-wide search for leech antiplatelet proteins in the non-blood-feeding leech Helobdella robusta (Rhyncobdellida: Glossiphoniidae) reveals evidence of secreted anticoagulants. Invertebr. Biol. 2011, 130, 344–350. [Google Scholar] [CrossRef]
- Orevi, M.; Eldor, A.; Giguzin, I.; Rigbi, M. Jaw anatomy of the blood-sucking leeches, Hirudinea Limnatis nilotica and Hirudo medicinalis, and its relationship to their feeding habits. J. Zool. 2000, 250, 121–127. [Google Scholar] [CrossRef]
- Hildebrandt, J.-P.; Lemke, S. Small bite, large impact—Saliva and salivary molecules in the medicinal leech, Hirudo medicinalis. Naturwissenschaften 2011, 98, 995–1008. [Google Scholar] [CrossRef] [PubMed]
- Kutschera, U.; Epshtein, V.M.; Nikolaj, A. Livanow (1876–1974) and the living relict Acanthobdella peledina (Annelida, Clitellata). Annals of the History and Philosophy of Biology 2006, 11, 85–98. [Google Scholar]
- Siddall, M. Validating Livanow: Molecular data agree that leeches, Branchiobdellidans, and Acanthobdella peledina form a monophyletic group of oligochaetes. Mol. Phylogenet. Evol. 2001, 21, 346–351. [Google Scholar] [CrossRef] [PubMed]
- Kaygorodova, I.A.; Dzyuba, E.V.; Pronin, N.M. Leech-Like parasites (Clitellata, Acanthobdellida) infecting native and endemic Eastern Siberian salmon fishes. ScientificWorldJournal 2012, 2012, 1–8. [Google Scholar] [CrossRef]
- Petersen, T.N.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods. 2011, 8, 785–786. [Google Scholar] [CrossRef] [PubMed]
- Apakupakul, K.; Siddall, M.E.; Burreson, E.M. Higher level relationships of leeches (Annelida: Clitellata: Euhirudinea) based on morphology and gene sequences. Mol. Phylogenet. Evol. 1999, 12, 350–359. [Google Scholar] [CrossRef] [PubMed]
- Salzet, M. Anticoagulants and inhibitors of platelet aggregation derived from leeches. FEBS Lett. 2001, 492, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Siddall, M.E.; Min, G.-S.; Fontanella, F.M.; Phillips, A.J.; Watson, S.C. Bacterial symbiont and salivary peptide evolution in the context of leech phylogeny. Parasitology 2011, 138, 1815–1827. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Tachikawa, Y.; Kitamura, M.; Ohno, O.; Suganuma, M.; Uemura, D. Complanine, an inflammation-inducing substance isolated from the marine fireworm Eurythoe complanata. Org. Biomol. Chem. 2008, 6, 2058. [Google Scholar] [CrossRef] [PubMed]
- Rouse, G.W.; Pleijel, F. Polychaetes; Oxford Univ Press: Oxford, UK, 2001. [Google Scholar]
- Fiege, D.; Bock, G. A new species of Archinome (Polychaeta: Archinomidae) from hydrothermal vents on the Pacific-Antarctic Ridge 37°S. J. Mar. Biol. Ass. 2009, 89, 689. [Google Scholar] [CrossRef]
- Zanol, J.; Halanych, K.M.; Struck, T.H.; Fauchald, K. Phylogeny of the bristle worm family Eunicidae (Eunicida, Annelida) and the phylogenetic utility of noncongruent 16S, COI and 18S in combined analyses. Mol. Phylogenet. Evol. 2010, 55, 660–676. [Google Scholar] [CrossRef] [PubMed]
- Wiklund, H.; Nygren, A.; Pleijel, F.; Sundberg, P. The phylogenetic relationships between Amphinomidae, Archinomidae and Euphrosinidae (Amphinomida: Aciculata: Polychaeta), inferred from molecular data. J. Mar. Biol. Ass. 2008, 88, 509–513. [Google Scholar] [CrossRef]
- Eckert, G.J. Absence of toxin-producing parapodial glands in amphinomid polychaetes (fireworms). Toxicon 1985, 23, 350–353. [Google Scholar] [CrossRef] [PubMed]
- Kem, W.R. Worm Peptides. In Handbook of Biologically Active Peptides; Academic Press: Waltham, MA, USA, 2013. [Google Scholar]
- Mazzanti, G.; Piccinelli, D. The occurrence of indole and imidazole compounds in marine worms and sea anemones from South Africa. Comp. Biochem. Physiol. C Comp. Pharmacol. Toxicol. 1979, 63, 215–219. [Google Scholar] [CrossRef]
- Nakamura, K.; Tachikawa, Y.; Uemura, D. (−)-Complanine, an inflammatory substance of marine fireworm: A synthetic study. Beilstein J. Org. Chem. 2009, 5, 12. [Google Scholar] [CrossRef]
- Nakamura, K.; Tachikawa, Y.; Ohno, O.; Kitamura, M.; Suganuma, M.; Uemura, D. Neocomplanines A and B, a Complanine Family isolated from the marine fireworm. J. Nat. Prod. 2010, 73, 303–305. [Google Scholar] [CrossRef] [PubMed]
- Pirooznia, M.; Gong, P.; Guan, X.; Inouye, L.S.; Yang, K.; Perkins, E.J.; Deng, Y. Cloning, analysis and functional annotation of expressed sequence tags from the Earthworm Eisenia fetida. BMC Bioinformatics 2007, 8, S7. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, H.; Ohta, N.; Umeda, M. Biology of lysenin, a protein in the coelomic fluid of the earthworm Eisenia foetida. Int. Rev. Cytol. 2004, 236, 45–99. [Google Scholar] [PubMed]
- Cooper, E.L.; Balamurugan, M. Unearthing a source of medicinal molecules. Drug Discov. Today 2010, 15, 966–972. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Kwon, J.; Aksoy, S. Characterization of genes expressed in the salivary glands of the tsetse fly, Glossina morsitans morsitans. Insect Mol. Biol. 2001, 10, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Wang, C.; Sun, Z. Vermipharmaceuticals and active proteins isolated from earthworms. Pedobiologia 2011, 54, S49–S56. [Google Scholar] [CrossRef]
- Domínguez, J. State-of-the-art and new perspectives on vermicomposting research. In Earthworm Ecology; CRC Press: Boca Raton, FL, USA, 2004; pp. 401–424. [Google Scholar]
- Lee, M.S.; Cho, S.J.; Tak, E.S.; Lee, J.A.; Cho, H.J.; Park, B.J.; Shin, C.; Kim, D.K.; Park, S.C. Transcriptome analysis in the midgut of the earthworm (Eisenia andrei) using expressed sequence tags. Biochem. Biophys. Res. Commun. 2005, 328, 1196–1204. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Losada, M.; Eiroa, J.; Mato, S.; Domínguez, J. Phylogenetic species delimitation of the earthworms Eisenia fetida (Savigny, 1826) and Eisenia andrei Bouché, 1972 (Oligochaeta, Lumbricidae) based on mitochondrial and nuclear DNA sequences. Pedobiologia 2005, 49, 317–324. [Google Scholar] [CrossRef]
- Bruhn, H.; Winkelmann, J.; Andersen, C.; Andrä, J.; Leippe, M. Dissection of the mechanisms of cytolytic and antibacterial activity of lysenin, a defense protein of the annelid Eisenia fetida. Dev. Comp. Immunol. 2006, 30, 597–606. [Google Scholar] [CrossRef] [PubMed]
- Kauschke, E.; Mohrig, W.; Cooper, E.L. Coelomic fluid proteins as basic components of innate immunity in earthworms. Eur. J. Soil Biol. 2007, 43, S110–S115. [Google Scholar] [CrossRef]
- Procházková, P.; Silerová, M.; Felsberg, J.; Josková, R.; Beschin, A.; de Baetselier, P.; Bilej, M. Relationship between hemolytic molecules in Eisenia fetida earthworms. Dev. Comp. Immunol. 2006, 30, 381–392. [Google Scholar] [CrossRef] [PubMed]
- Lassalle, F.; Lassègues, M.; Roch, P. Protein analysis of earthworm coelomic fluid IV. Evidence, activity induction and purification of Eisenia fetida andrei lysozyme (Annelidae). Comp. Biochem. Physiol., B 1988, 91, 187–192. [Google Scholar]
- Sekizawa, Y.; Kubo, T.; Kobayashi, H.; Nakajima, T.; Natori, S. Molecular cloning of cDNA for lysenin, a novel protein in the earthworm Eisenia foetida that causes contraction of rat vascular smooth muscle. Gene 1997, 191, 97–102. [Google Scholar] [CrossRef] [PubMed]
- De Colibus, L.; Sonnen, A.F.P.; Morris, K.J.; Siebert, C.A.; Abrusci, P.; Plitzko, J.; Hodnik, V.; Leippe, M.; Volpi, E.; Anderluh, G.; Gilbert, R.J.C. Structures of lysenin reveal a shared evolutionary origin for pore-forming proteins and its mode of sphingomyelin recognition. Structure 2012, 20, 1498–1507. [Google Scholar] [CrossRef] [PubMed]
- Sukumwang, N.; Umezawa, K. Earthworm-derived pore-forming toxin Lysenin and screening of its inhibitors. Toxins 2013, 5, 1392–1401. [Google Scholar] [CrossRef] [PubMed]
- Szczesny, P.; Iacovache, I.; Muszewska, A.; Ginalski, K.; van der Goot, F.G.; Grynberg, M. Extending the Aerolysin family: From bacteria to vertebrates. PLoS One 2011, 6, e20349. [Google Scholar] [CrossRef] [PubMed]
- Boggemann, M. Revision of the glyceridae grube 1850 (Annelida: Polychaeta). Abh. Senckenb. Naturforch. Ges. 2002, 555, 1–249. [Google Scholar]
- Meunier, F.A.; Feng, Z.-P.; Molgo, J.; Zamponi, G.W.; Schiavo, G. Glycerotoxin from Glycera convoluta stimulates neurosecretion by up-regulating N-type Ca2+ channel activity. EMBO J. 2002, 21, 6733–6743. [Google Scholar] [CrossRef] [PubMed]
- Kagan, B.L.; Pollard, H.B.; Hanna, R.B. Induction of ion-permeable channels by the venom of the fanged bloodworm Glycera dibranchiata. Toxicon 1982, 20, 887–893. [Google Scholar] [CrossRef] [PubMed]
- Moses, D.N.; Slack, N.L.; Waitre, J.H.; Zok, F.W. Role of melanin in mechanical properties of Glycera jaws. Acta Biomater. 2006, 2, 521–530. [Google Scholar] [CrossRef] [PubMed]
- Lichtenegger, H.C. High abrasion resistance with sparse mineralization: Copper biomineral in worm jaws. Science 2002, 298, 389–392. [Google Scholar] [CrossRef]
- Ambrose, W. Influences of predatory polychaetes and epibenthic predators on the structure of a soft-bottom community in a Maine estuary. J. Exp. Mar. Biol. Ecol. 1984, 81, 115–145. [Google Scholar] [CrossRef]
- Ungerger, N.K. Characterization of the venom of Glycera dibranchiata. Department of Biology, Sweet Briar College: Virginia, VA, USA, 2002. [Google Scholar]
- Giray, C.; King, M.G. Predator deterrence and 2,4-dibromophenol conservation by the enteropneusts Saccoglossus bromophenolosus and Protoglassus graveolensis. Mar. Ecol. Prog. Ser. 1997, 159, 229–238. [Google Scholar] [CrossRef]
- Michel, C.; Keil, B. Biologically active proteins in the venomous glands of the polychaetous annelid, Glycera convoluta Keferstein. Comp. Biochem. Physiol., B 1975, 50, 29–33. [Google Scholar] [PubMed]
- Michel, C.; Imhoff, J.-M. Localization and biochemical characterization of pharyngeal protease in the polychaetous annelid Glycera convoluta. Mar. Biol. 1975, 32, 343–347. [Google Scholar] [CrossRef]
- Bon, C.; Saliou, B.; Thieffry, M.; Manaranche, R. Partial purification of α-glycerotoxin, a presynaptic neurotoxin from the venom glands of the polychaete annelid Glycera convoluta. Neurochem. Int. 1985, 7, 63–75. [Google Scholar] [CrossRef] [PubMed]
- Meunier, F.A.; Nguyen, T.H.; Colasante, C.; Luo, F.; Sullivan, R.K.P.; Lavidis, N.A.; Molgo, J.; Meriney, S.D.; Schiavo, G. Sustained synaptic-vesicle recycling by bulk endocytosis contributes to the maintenance of high-rate neurotransmitter release stimulated by glycerotoxin. J. Cell. Sci. 2010, 123, 1131–1140. [Google Scholar] [CrossRef] [PubMed]
- Schenning, M.; Proctor, D.T.; Ragnarsson, L.; Barbier, J.; Lavidis, N.A.; Molgo, J.J.; Zamponi, G.W.; Schiavo, G.; Meunier, F.A. Glycerotoxin stimulates neurotransmitter release from N-type Ca2+ channel expressing neurons. J. Neurochem. 2006, 98, 894–904. [Google Scholar] [CrossRef] [PubMed]
- Casewell, N.R.; Harrison, R.A.; Wüster, W.; Wagstaff, S.C. Comparative venom gland transcriptome surveys of the saw-scaled vipers (Viperidae: Echis) reveal substantial intra-family gene diversity and novel venom transcripts. BMC Genomics 2009, 10, 564. [Google Scholar] [CrossRef] [PubMed]
- Low, D.H.W.; Sunagar, K.; Undheim, E.A.B.; Ali, S.A.; Alagon, A.C.; Ruder, T.; Jackson, T.N.W.; Pineda Gonzalez, S.; King, G.F.; Jones, A.; et al. Dracula’s children: Molecular evolution of vampire bat venom. J. Proteomics 2013, 89, 95–111. [Google Scholar] [CrossRef] [PubMed]
- Wong, E.S.W.; Nicol, S.; Warren, W.C.; Belov, K. Echidna venom gland transcriptome provides insights into the evolution of monotreme venom. PloS One 2013, 8, e79092. [Google Scholar] [CrossRef] [PubMed]
- Norlinder, E.; Nygren, A.; Wiklund, H.; Pleijel, F. Phylogeny of scale-worms (Aphroditiformia, Annelida), assessed from 18SrRNA, 28SrRNA, 16SrRNA, mitochondrial cytochrome c oxidase subunit I (COI), and morphology. Mol. Phylogenet. Evol. 2012, 65, 490–500. [Google Scholar] [CrossRef] [PubMed]
- Struck, T.H.; Purschke, G.; Halanych, K.M. A scaleless scale worm: Molecular evidence for the phylogenetic placement of Pisione remota (Pisionidae, Annelida). Mar. Biol. Res. 2005, 1, 243–253. [Google Scholar] [CrossRef]
- Wiklund, H.; Nygren, A.; Pleijel, F.; Sundberg, P. Phylogeny of Aphroditiformia (Polychaeta) based on molecular and morphological data. Mol. Phylogenet. Evol. 2005, 37, 494–502. [Google Scholar] [CrossRef] [PubMed]
- Pettibone, M.H. Additions to the family Eulepethidae Chamberlin (Polychaeta: Aphroditacea); Smithsonian Institution Press: Washington, DC, USA, 1986. [Google Scholar]
- Miyazawa, K.; Jeon, J.; Noguchi, T.; Ito, K.; Hashimoto, K. Distribution of tetrodotoxin in the tissues of the flatworm Planocera multitentaculata (Platyhelminthes). Toxicon 1987, 25, 975–980. [Google Scholar] [CrossRef] [PubMed]
- Tanu, M.; Mahmud, Y.; Arakawa, O.; Takatani, T.; Kajihara, H.; Kawatsu, K.; Hamano, Y.; Asakawa, M.; Miyazawa, K.; Noguchi, T. Immunoenzymatic visualization of tetrodotoxin (TTX) in Cephalothrix species (Nemertea: Anopla: Palaeonemertea: Cephalotrichidae) and Planocera reticulata (Platyhelminthes: Turbellaria: Polycladida: Planoceridae). Toxicon 2004, 44, 515–520. [Google Scholar] [CrossRef] [PubMed]
- Ritson-Williams, R.; Yotsu-Yamashita, M.; Paul, V.J. Ecological functions of tetrodotoxin in a deadly polyclad flatworm. Proc. Natl. Acad. Sci. U.S.A. 2006, 103, 3176–3179. [Google Scholar] [CrossRef] [PubMed]
- Williams, B.L. Behavioral and chemical ecology of marine organisms with respect to tetrodotoxin. Mar. Drugs 2010, 8, 381–398. [Google Scholar] [CrossRef] [PubMed]
- Dumont, H.; Carels, I. Flatworm predator (Mesostoma cf. lingua) releases a toxin to catch planktonic prey (Daphnia magna). Limnol. Oceanogr. 1987, 32, 699–702. [Google Scholar]
- Blaustein, L.; Dumont, H. Typhloplanid flatworms (Mesostoma and related genera): Mechanisms of predation and evidence that they structure aquatic invertebrate communities. Hydrobiologia 1990, 198, 61–77. [Google Scholar] [CrossRef]
- Nandini, S.; Sarma, S.S.S.; Dumont, H.J. Predatory and toxic effects of the turbellarian (Stenostomum cf leucops) on the population dynamics of Euchlanis dilatata, Plationus patulus (Rotifera) and Moina macrocopa (Cladocera). Hydrobiologia 2011, 662, 171–177. [Google Scholar] [CrossRef]
- Wrona, F.; Koopowitz, H. Behavior of the rhabdocoel flatworm Mesostoma ehrenbergiiin prey capture and feeding. Hydrobiologia 1998, 383, 35–40. [Google Scholar] [CrossRef]
- Kem, W.R. Worm venom peptides. In Handbook of Biologically Active Peptides; Kastion, A.J., Ed.; Academic Press: Waltham, MA, USA, 2006; pp. 397–401. [Google Scholar]
- Chalmers, I.W.; Hoffmann, K.F. Platyhelminth Venom Allergen-Like (VAL) proteins: Revealing structural diversity, class-specific features and biological associations across the phylum. Parasitology 2012, 139, 1231–1245. [Google Scholar] [CrossRef] [PubMed]
- McDermott, J. Observations on the food and feeding behavior of estuarine nemertean worms belonging to the order Hoplonemertea. Biol. Bull. 1976, 150, 57–68. [Google Scholar] [CrossRef]
- Stricker, S.; Cloney, R. The stylet apparatus of the nemertean Paranemertes peregrina: Its ultrastructure and role in prey capture. Zoomorphology 1981, 97, 205–223. [Google Scholar] [CrossRef]
- Thollesson, M.; Norenburg, J.L. Ribbon worm relationships: A phylogeny of the phylum Nemertea. Proc. Biol. Sci. 2003, 270, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Kvist, S.; Laumer, C.E.; Junoy, J.; Giribet, G. New insights into the phylogeny, systematics and DNA barcoding of Nemertea. Invertebr. Syst. 2014, 28, 287–308. [Google Scholar] [CrossRef]
- Andrade, S.; Montenegro, H.; Strand, M. A transcriptomic approach to ribbon worm systematics (Nemertea): Resolving the Pilidiophora problem. Mol. Biol. Evol. 2014. [Google Scholar] [CrossRef]
- McDermott, J.; Roe, P. Food, feeding behavior and feeding ecology of nemerteans. Am. Zool. 1985, 25, 113–125. [Google Scholar]
- Stricker, S.A.; Cloney, R.A. The ultrastructure of venom-producing cells in Paranemertes peregrina (Nemertea, Hoplonemertea). J. Morphol. 1983, 177, 89–107. [Google Scholar] [CrossRef]
- Kem, W. Structure and action of nemertine toxins. Am. Zool. 1985, 25, 99–111. [Google Scholar]
- Jennings, J.B.; Gibson, R. Observations on the nutrition of seven species of rhynchocoelan worms. Biol. Bull. 1969, 136, 405–433. [Google Scholar] [CrossRef]
- Carroll, S.; McEvoy, E.; Gibson, R. The production of tetrodotoxin-like substances by nemertean worms in conjunction with bacteria. J. Exp. Mar. Biol. Ecol. 2003, 288, 51–63. [Google Scholar] [CrossRef]
- Asakawa, M.; Ito, K.; Kajihara, H. Highly toxic ribbon worm Cephalothrix simula containing tetrodotoxin in Hiroshima Bay, Hiroshima Prefecture, Japan. Toxins 2013, 5, 376–395. [Google Scholar] [CrossRef] [PubMed]
- Beleneva, I.A.; Magarlamov, T.Y.; Kukhlevsky, A.D. Characterization, identification, and screening for tetrodotoxin production by bacteria associated with the ribbon worm (Nemertea) Cephalotrix simula (Ivata, 1952). Microbiology 2014, 83, 220–226. [Google Scholar] [CrossRef]
- Magarlamov, T.Y.; Beleneva, I.A.; Chernyshev, A.V.; Kuhlevsky, A.D. Tetrodotoxin-producing Bacillus sp. from the ribbon worm (Nemertea) Cephalothrix simula (Iwata, 1952). Toxicon 2014, 85, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Berne, S.; Sepčić, K.; Križaj, I.; Kem, W.R.; Mcclintock, J.B.; Turk, T. Isolation and characterisation of a cytolytic protein from mucus secretions of the Antarctic heteronemertine Parborlasia corrugatus. Toxicon 2003, 41, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Kem, W.R.; Mahnir, V.M.; Papke, R.L.; Lingle, C.J. Anabaseine is a potent agonist on muscle and neuronal alpha-bungarotoxin-sensitive nicotinic receptors. J. Pharmacol. Exp. Ther. 1997, 283, 979–992. [Google Scholar] [PubMed]
- McDermott, J. Status of the Nemertea as prey in marine ecosystems. Hydrobiologia 2001, 456, 7–20. [Google Scholar] [CrossRef]
- Stricker, S. The stylet apparatus of monostiliferous hoplonemerteans. Am. Zool. 1985, 25, 87–97. [Google Scholar]
- Olivera, B.M.; Showers-Corneli, P.; Watkins, M.; Fedosov, A. Biodiversity of cone snails and other venomous marine gastropods: Evolutionary success through neuropharmacology. Annu. Rev. Anim. Biosci. 2013, 2, 487–513. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.D. Understanding biodiversity: Adaptive radiations of predatory marine gastropods. In The Marine Biology of the South China Sea; Morton, B., Ed.; Hong Kong University Press: Hong Kong, 1998; pp. 187–206. [Google Scholar]
- Ruder, T.; Sunagar, K.; Undheim, E.A.B.; Ali, S.A.; Wai, T.-C.; Low, D.H.W.; Jackson, T.N.W.; King, G.F.; Antunes, A.; Fry, B.G. Molecular phylogeny and evolution of the proteins encoded by coleoid (cuttlefish, octopus, and squid) posterior venom glands. J. Mol. Evol. 2013, 76, 192–204. [Google Scholar] [CrossRef] [PubMed]
- Bouchet, P.; Lozouet, P.; Sysoev, A. An inordinate fondness for turrids. Deep Sea Res. II Top. Stud. Oceanogr. 2009, 56, 1724–1731. [Google Scholar] [CrossRef]
- Puillandre, N.; Kantor, Y.I.; Sysoev, A.; Couloux, A.; Meyer, C.; Rawlings, T.; Todd, J.A.; Bouchet, P. The dragon tamed? A molecular phylogeny of the Conoidea (Gastropoda). J. Molluscan Stud. 2011, 77, 259–272. [Google Scholar] [CrossRef]
- López-Vera, E.; Heimer de la Cotera, E.; Maillo, M.; Riesgo-Escovar, J.; Olivera, B.; Aguilar, M. A novel structural class of toxins: The methionine-rich peptides from the venoms of turrid marine snails (Mollusca, Conoidea). Toxicon 2004, 43, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Arumugam, M.; Giji, S.; Tamilmozhi, S.; Kumar, S.; Balasubramanian, T. Studies on biochemical and biological properties of turrids venom (Turricula javana and Lophiotoma indica). Indian J. Geomar. Sci. 2013, 42, 800–806. [Google Scholar]
- Watkins, M.; Hillyard, D.R.; Olivera, B.M. Genes expressed in a turrid venom duct: Divergence and similarity to conotoxins. J. Mol. Evol. 2006, 62, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Heralde, F.M., III; Imperial, J.; Bandyopadhyay, P.K.; Olivera, B.M.; Concepcion, G.P.; Santos, A.D. A rapidly diverging superfamily of peptide toxins in venomous Gemmula species. Toxicon 2008, 51, 890–897. [Google Scholar] [CrossRef] [PubMed]
- Imperial, J.S.; Cabang, A.B.; Song, J.; Raghuraman, S.; Gajewiak, J.; Watkins, M.; Showers-Corneli, P.; Fedosov, A.; Concepcion, G.P.; Terlau, H.; et al. A family of excitatory peptide toxins from venomous crassispirine snails: Using Constellation Pharmacology to assess bioactivity. Toxicon 2014, 89, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Cabang, A.B.; Imperial, J.S.; Gajewiak, J.; Watkins, M.; Corneli, P.S.; Olivera, B.M.; Concepcion, G.P. Characterization of a venom peptide from a crassispirid gastropod. Toxicon 2011, 58, 672–680. [Google Scholar] [CrossRef] [PubMed]
- Holford, M.; Puillandre, N.; Modica, M.V.; Watkins, M.; Collin, R.; Bermingham, E.; Olivera, B.M. Correlating molecular phylogeny with venom apparatus occurrence in Panamic auger snails (Terebridae). PLoS One 2009, 4, e7667. [Google Scholar] [CrossRef] [PubMed]
- Holford, M.; Puillandre, N.; Terryn, Y.; Cruaud, C.; Olivera, B.; Bouchet, P. Evolution of the Toxoglossa venom apparatus as inferred by molecular phylogeny of the Terebridae. Mol. Biol. Evol. 2009, 26, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Puillandre, N.; Holford, M. The Terebridae and teretoxins: Combining phylogeny and anatomy for concerted discovery of bioactive compounds. BMC Chem. Biol. 2010, 10, 7. [Google Scholar] [CrossRef] [PubMed]
- Castelin, M.; Puillandre, N.; Kantor, Y.I.; Modica, M.V.; Terryn, Y.; Cruaud, C.; Bouchet, P.; Holford, M. Macroevolution of venom apparatus innovations in auger snails (Gastropoda; Conoidea; Terebridae). Mol. Phylogenet. Evol. 2012, 64, 21–44. [Google Scholar] [CrossRef] [PubMed]
- Imperial, J.S.; Kantor, Y.; Watkins, M.; Heralde, F.M.; Stevenson, B.; Chen, P.; Hansson, K.; Stenflo, J.; Ownby, J.-P.; Bouchet, P.; et al. Venomous auger snail Hastula (Impages) hectica (Linnaeus, 1758): Molecular phylogeny, foregut anatomy and comparative toxinology. J. Exp. Zool. 2007, 308B, 744–756. [Google Scholar] [CrossRef]
- Kendel, Y.; Melaun, C.; Kurz, A.; Nicke, A.; Peigneur, S.; Tytgat, J.; Wunder, C.; Mebs, D.; Kauferstein, S. Venomous secretions from marine snails of the Terebridae family target acetylcholine receptors. Toxins 2013, 5, 1043–1050. [Google Scholar] [CrossRef] [PubMed]
- Shiomi, K.; Kawashima, Y.; Mizukami, M.; Nagashima, Y. Properties of proteinaceous toxins in the salivary gland of the marine gastropod (Monoplex echo). Toxicon 2002, 40, 563–571. [Google Scholar] [CrossRef] [PubMed]
- Kawashima, Y.; Nagai, H.; Ishida, M.; Nagashima, Y.; Shiomi, K. Primary structure of echotoxin 2, an actinoporin-like hemolytic toxin from the salivary gland of the marine gastropod Monoplex echo. Toxicon 2003, 42, 491–497. [Google Scholar] [CrossRef] [PubMed]
- Shiomi, K.; Ishii, M.; Shimakura, K.; Nagashima, Y.; Chino, M. Tigloylcholine: A new choline ester toxin from the hypobranchial gland of two species of muricid gastropods (Thais clavigera and Thais bronni). Toxicon 1998, 36, 795–798. [Google Scholar] [CrossRef] [PubMed]
- Undheim, E.A.B.; Georgieva, D.N.; Thoen, H.H.; Norman, J.A.; Mork, J.; Betzel, C.; Fry, B.G. Venom on ice: First insights into Antarctic octopus venoms. Toxicon 2010, 56, 897–913. [Google Scholar] [CrossRef] [PubMed]
- Williams, B.L.; Stark, M.R.; Caldwell, R.L. Microdistribution of tetrodotoxin in two species of blue-ringed octopuses (Hapalochlaena lunulata and Hapalochlaena fasciata) detected by fluorescent immunolabeling. Toxicon 2012, 60, 1307–1313. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, I.; Valladolid, G.; Varon, J.; Sternbach, G. Encounters with venomous sea-life. J. Emerg. Med. 2011, 40, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Townsend, K.A.; Altvater, J.; Thomas, M.C.; Schuyler, Q.A.; Nette, G.W. Death in the octopus’ garden: Fatal blue-lined octopus envenomations of adult green sea turtles. Mar. Biol. 2012, 159, 689–695. [Google Scholar] [CrossRef] [PubMed]
- Halstead, W.B. Venomous echinoderms and annelids: Starfishes, sea Urchins, sea cucumbers, and segmented worms. In Venomous Animals and Their Venoms. Voll III Venomous Invertebrates; Bucherl, W., Buckley, E.E., Eds.; Academic Press, Inc.: Waltham, MA, USA, 1971. [Google Scholar]
- Chanley, J.D.; Mezzetti, T.; Sobotka, H. The holothurinogenins. Tetrahedron 1966, 22, 1857–1884. [Google Scholar] [CrossRef]
- Chanley, J.D.; Rossi, C. The holothurinogenins—II: Methoxylated neo-holothurinogenins. Tetrahedron 1969, 25, 1897–1909. [Google Scholar] [CrossRef] [PubMed]
- Habermehl, G.G. Echinodermata (Echinoderms). In Venomous Animals and Their Toxins; Habermehl, G.G., Ed.; Springer-Verlag: New York, NY, USA, 1981. [Google Scholar]
- Lee, C.C.; Hsieh, H.J.; Hsieh, C.H.; Hwang, D.F. Antioxidative and anticancer activities of various ethanolic extract fractions from crown-of-thorns starfish (Acanthaster planci). Environ. Toxicol. Pharmacol. 2014, 38, 761–773. [Google Scholar] [PubMed]
- Lee, C.-C.; Hsieh, H.J.; Hsieh, C.-H.; Hwang, D.-F. Spine venom of crown-of-thorns starfish (Acanthaster planci) induces antiproliferation and apoptosis of human melanoma cells (A375.S2). Toxicon 2014, 91, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Shiomi, K.; Midorikawa, S.; Ishida, M.; Nagashima, Y.; Nagai, H. Plancitoxins, lethal factors from the crown-of-thorns starfish Acanthaster planci, are deoxyribonucleases II. Toxicon 2004, 44, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, A.; Nagai, H.; Nagashima, Y.; Shiomi, K. Structural characterization of plancitoxin I, a deoxyribonuclease II-like lethal factor from the crown-of-thorns starfish Acanthaster planci, by expression in Chinese hamster ovary cells. Fish. Sci. 2008, 75, 225–231. [Google Scholar] [CrossRef]
- Ihama, Y.; Fukasawa, M.; Ninomiya, K.; Kawakami, Y.; Nagai, T.; Fuke, C.; Miyazaki, T. Anaphylactic shock caused by sting of crown-of-thorns starfish (Acanthaster planci). Forensic Sci. Int. 2014, 236, e5–e8. [Google Scholar] [CrossRef] [PubMed]
- Kroh, A.; Smith, A.B. The phylogeny and classification of post-Palaeozoic echinoids. J. Syst. Palaeontol. 2010, 8, 147–212. [Google Scholar] [CrossRef]
- Coppard, S.E.; Kroh, A.; Smith, A.B. The evolution of pedicellariae in echinoids: An arms race against pests and parasites. Acta Zool. 2010, 93, 125–148. [Google Scholar] [CrossRef]
- Alender, C.B.; Feigen, G.A.; Tomita, J.T. Isolation and characterization of sea urchin toxin. Toxicon 1965, 3, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Shiomi, K.; Yamamoto, S.; Yamanaka, H.; Kikuchi, T. Purification and characterization of a lethal factor in venom from the crown-of-thorns starfish (Acanthaster planci). Toxicon 1988, 26, 1077–1083. [Google Scholar] [CrossRef] [PubMed]
- Shiomi, K.A.; Kazama, A.; Shimakura, K.; Nagashima, Y. Purification and properties of phospholipases A2 from the crown-of-thorns starfish (Acanthaster planci) venom. Toxicon 1998, 36, 589–599. [Google Scholar] [CrossRef] [PubMed]
- Ota, E.; Nagai, H.; Nagashima, Y.; Shiomi, K. Molecular cloning of two toxic phospholipases A2 from the crown-of-thorns starfish Acanthaster planci venom. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2006, 143, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Karasudani, I.; Koyama, T.; Nakandakari, S.; Aniya, Y. Purification of anticoagulant factor from the spine venom of the crown-of-thorns starfish, Acanthaster planci. Toxicon 1996, 34, 871–879. [Google Scholar] [CrossRef] [PubMed]
- Ota, E.; Nagashima, Y.; Shiomi, K.; Sakurai, T.; Kojima, C.; Waalkes, M.P.; Himeno, S. Caspase-independent apoptosis induced in rat liver cells by plancitoxin I, the major lethal factor from the crown-of-thorns starfish Acanthaster planci venom. Toxicon 2006, 48, 1002–1010. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.-C.; Tsai, W.-S.; Hsieh, H.J.; Hwang, D.-F. Hemolytic activity of venom from crown-of-thorns starfish Acanthaster planci spines. J. Venom. Anim. Toxins Incl. Trop. Dis. 2013, 19, 22. [Google Scholar] [CrossRef] [PubMed]
- Sciani, J.M.; Antoniazzi, M.M.; da Costa Neves, A.; Pimenta, D.C. Cathepsin B/X is secreted by Echinometra lucunter sea urchin spines, a structure rich in granular cells and toxins. J. Venom. Anim. Toxins Incl. Trop. Dis. 2013, 19, 8. [Google Scholar] [CrossRef] [PubMed]
- Kuwabara, S. Purification and properties of peditoxin and the structure of its prosthetic group, pedoxin, from the sea urchin Toxopneustes pileolus (Lamarck). J. Biol. Chem. 1994, 269, 26734–26738. [Google Scholar] [PubMed]
- Kimura, A.; Nakagawa, H. Action of an extract from the sea urchin Toxopneustes pileolus on isolated smooth muscle. Toxicon 1980, 18, 689–693. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, H.; Tu, A.T.; Kimura, A. Purification and characterization of Contractin A from the pedicellarial venom of sea urchin, Toxopneustes pileolus. Arch. Biochem. Biophys. 1991, 284, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, H.; Hashimoto, T.; Hayashi, H.; Shinohara, M.; Ohura, K.; Tachikawa, E.; Kashimoto, T. Isolation of a novel lectin from the globiferous pedicellariae of the sea urchin Toxopneustes pileolus. Adv. Exp. Med. Biol. 1996, 391, 213–223. [Google Scholar] [PubMed]
- Zhang, Y.; Wada, T.; Nakagawa, H.; Ichida, S. A toxic extract from sea urchin (T. pileolus) inhibits 45Ca2+ uptake in P2 fraction from chick brain under physiological ionic conditions. Toxicon 1999, 37, 1639–1643. [Google Scholar] [CrossRef] [PubMed]
- FastQC. Available online: http://www.bioinformatics.bbsrc.ac.uk/projects/fastqc (accessed on 11 December 2014).
- Patel, R.K.; Jain, M. NGS QC Toolkit: A toolkit for quality control of next generation sequencing data. PLoS One 2012, 7, e30619. [Google Scholar] [CrossRef] [PubMed]
- Dodt, M.; Roehr, J.; Ahmed, R.; Dieterich, C. FLEXBAR—Flexible Barcode and Adapter Processing for Next-Generation Sequencing Platforms. Biology 2012, 1, 895–905. [Google Scholar] [CrossRef] [PubMed]
- Heinz, S.; Benner, C.; Spann, N.; Bertolino, E.; Lin, Y.C.; Laslo, P.; Cheng, J.X.; Murre, C.; Singh, H.; Glass, C.K. Simple combinations of lineage-determining transcription factors prime cis-regulatory elements required for macrophage and B cell identities. Mol. Cell. 2010, 38, 576–589. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Leung, H.C.M.; Yiu, S.-M.; Lv, M.-J.; Zhu, X.-G.; Chin, F.Y.L. IDBA-tran: A more robust de novo de Bruijn graph assembler for transcriptomes with uneven expression levels. Bioinformatics 2013, 29, i326–i334. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Zhu, H.; Ruan, J.; Qian, W.; Fang, X.; Shi, Z.; Li, Y.; Li, S.; Shan, G.; Kristiansen, K.; et al. De novo assembly of human genomes with massively parallel short read sequencing. Genome Res. 2010, 20, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Zhao, L.; Gao, J.; Fei, Z. iAssembler: A package for de novo assembly of Roche-454/Sanger transcriptome sequences. BMC Bioinformatics 2011, 12, 453. [Google Scholar] [CrossRef] [PubMed]
- SeqClean. Available online: ftp://occams.dfci.harvard.edu/pub/(accessed on 11 December 2014).
- VecScreen. Available online: http://www.ncbi.nlm.nih.gov/tools/vecscreen/ (accessed on 11 December 2014).
- Hoffmann, S.; Otto, C.; Kurtz, S.; Sharma, C.M.; Khaitovich, P.; Vogel, J.; Stadler, P.F.; Hackermüller, J. Fast mapping of short sequences with mismatches, insertions and deletions using index structures. PLoS Comput. Biol. 2009, 5, e1000502. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 2009, 10, R25. [Google Scholar] [CrossRef] [PubMed]
- Milne, I.; Peng, Y.; Stephen, G.; Leung, H.C.M.; Bayer, M.; Yiu, S.-M.; Cock, P.J.A.; Lv, M.-J.; Pritchard, L.; Zhu, X.-G.; et al. Using Tablet for visual exploration of second-generation sequencing data. Brief. Bioinformatics 2013, 14, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Casewell, N.R.; Huttley, G.A.; Ster, W.W.U. Dynamic evolution of venom proteins in squamate reptiles. Nat. Commun. 2012, 3, 1066:1–1066:10. [Google Scholar] [CrossRef]
- Reyes-Velasco, J.; Card, D.C.; Andrew, A.L.; Shaney, K.J.; Adams, R.H.; Schield, D.R.; Casewell, N.R.; Mackessy, S.P.; Castoe, T.A. Expression of venom gene homologs in diverse python tissues suggests a new model for the evolution of snake venom. Mol. Biol. Evol. 2014. [Google Scholar] [CrossRef]
- Alfaro, M.E. Bayes or Bootstrap? A simulation study comparing the performance of Bayesian Markov Chain Monte Carlo sampling and Bootstrapping in assessing phylogenetic confidence. Mol. Biol. Evol. 2003, 20, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Lemmon, A.R.; Brown, J.M.; Stanger-Hall, K.; Lemmon, E.M. The effect of ambiguous data on phylogenetic estimates obtained by Maximum Likelihood and Bayesian inference. Syst. Biol. 2009, 58, 130–145. [Google Scholar] [CrossRef] [PubMed]
- i5K. Available online: http://www.arthropodgenomes.org/wiki/i5K (accessed on 11 December 2014).
- Wong, E.S.W.; Papenfuss, A.T.; Whittington, C.M.; Warren, W.C.; Belov, K. A limited role for gene duplications in the evolution of platypus venom. Mol. Biol. Evol. 2012, 29, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Casewell, N.R.; Wagstaff, S.C.; Wüster, W.; Cook, D.A.N.; Bolton, F.M.S.; King, S.I.; Pla, D.; Sanz, L.; Calvete, J.J.; Harrison, R.A. Medically important differences in snake venom composition are dictated by distinct postgenomic mechanisms. Proc. Natl. Acad. Sci. USA 2014, 111, 9205–9210. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Von Reumont, B.M.; Campbell, L.I.; Jenner, R.A. Quo Vadis Venomics? A Roadmap to Neglected Venomous Invertebrates. Toxins 2014, 6, 3488-3551. https://doi.org/10.3390/toxins6123488
Von Reumont BM, Campbell LI, Jenner RA. Quo Vadis Venomics? A Roadmap to Neglected Venomous Invertebrates. Toxins. 2014; 6(12):3488-3551. https://doi.org/10.3390/toxins6123488
Chicago/Turabian StyleVon Reumont, Bjoern Marcus, Lahcen I. Campbell, and Ronald A. Jenner. 2014. "Quo Vadis Venomics? A Roadmap to Neglected Venomous Invertebrates" Toxins 6, no. 12: 3488-3551. https://doi.org/10.3390/toxins6123488
APA StyleVon Reumont, B. M., Campbell, L. I., & Jenner, R. A. (2014). Quo Vadis Venomics? A Roadmap to Neglected Venomous Invertebrates. Toxins, 6(12), 3488-3551. https://doi.org/10.3390/toxins6123488