Modeling Temperature Requirements for Growth and Toxin Production of Alternaria spp. Associated with Tomato




Abstract
1. Introduction
2. Results
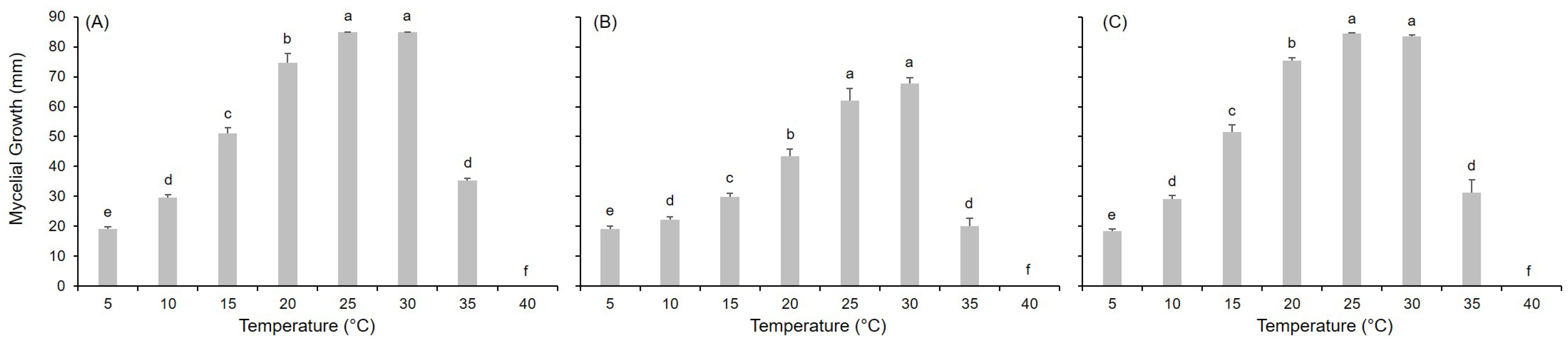
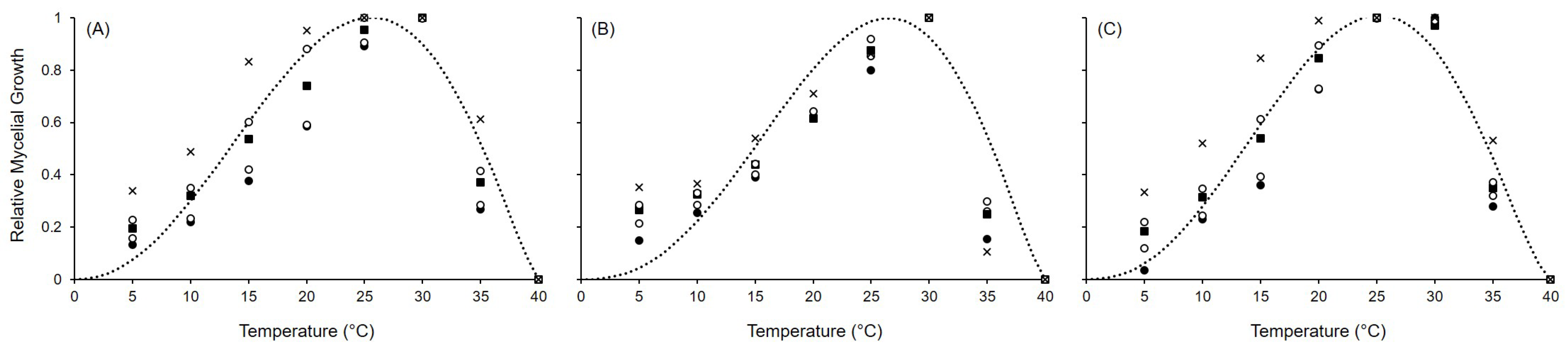
2.1. Mycelial Growth
2.2. Mycotoxin Production
3. Discussion
4. Materials and Methods
4.1. Fungal Isolates and Culture Conditions
4.2. Mycelial Growth
4.3. Mycotoxin Analysis
4.3.1. Reagents and Chemicals
4.3.2. Mycotoxin Extraction
4.3.3. UHPLC-TWIMS-QTOF Screening of Mycotoxins
4.4. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pandey, K.K.; Pandey, P.K.; Kalloo, G.; Banerjee, M.K. Resistance to early blight of tomato with respect to various parameters of disease epidemics. J. Gen. Plant Pathol. 2003, 69, 364–371. [Google Scholar] [CrossRef]
- Chaerani, R.; Voorrips, R.E. Tomato early blight (Alternaria solani): The pathogen, genetics, and breeding for resistance. J. Gen. Plant Pathol. 2006, 72, 335–347. [Google Scholar] [CrossRef]
- Troncoso-Rojas, R.; Tiznado-Hernandez, M.E. Alternaria alternata (black rot, black spot). In Postharvest Decay; Bautista-Baños, S., Ed.; Academic Press: London, UK, 2014; pp. 147–187. [Google Scholar]
- Lou, J.; Fu, L.; Peng, Y.; Zhou, L. Metabolites from Alternaria fungi and their bioactivities. Molecules 2013, 18, 5891–5935. [Google Scholar] [CrossRef] [PubMed]
- Patriarca, A. Alternaria in food products. Curr. Opin. Food Sci. 2016, 11, 1–9. [Google Scholar] [CrossRef]
- Li, J.F.; Jiang, H.B.; Jeewon, R.; Hongsanan, S.; Bhat, D.J.; Tang, S.M.; Lumyong, S.; Mortimer, P.E.; Xu, J.C.; Camporesi, E.; et al. Alternaria: Update on species limits, evolution, multi-locus phylogeny, and classification. Studi. Fungi 2023, 8, 1–61. [Google Scholar] [CrossRef]
- Bertuzzi, T.; Rastelli, S.; Pietri, A.; Giorni, P. Alternaria toxins in tomato products in Northern Italy in the period 2017–2019. Food Addit. Contam. Part B 2021, 14, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Sanzani, S.M.; Gallone, T.; Garganese, F.; Caruso, A.G.; Amenduni, M.; Ippolito, A. Contamination of fresh and dried tomato by Alternaria toxins in southern Italy. Food Addit. Contam. Part A 2019, 36, 789–799. [Google Scholar] [CrossRef] [PubMed]
- Woudenberg, J.H.C.; Truter, M.; Groenewald, J.Z.; Crous, P.W. Large-spored Alternaria pathogens in section Porri disentangled. Stud. Mycol. 2014, 79, 1–47. [Google Scholar] [CrossRef] [PubMed]
- Dalinova, A.A.; Salimova, D.R.; Berestetskiy, A.O. Fungi of the genera Alternaria as producers of biological active compounds and mycoherbicides. Appl. Biochem. Microbiol. 2020, 56, 256–272. [Google Scholar] [CrossRef]
- Pinto, V.E.F.; Patriarca, A. Alternaria species and their associated mycotoxins. In Mycotoxigenic Fungi: Methods and Protocols; Moretti, A., Susca, A., Eds.; Humana: New York, NY, USA, 2016; pp. 13–32. [Google Scholar]
- Wenderoth, M.; Garganese, F.; Schmidt-Heydt, M.; Soukup, S.T.; Ippolito, A.; Sanzani, S.M.; Fischer, R. Alternariol as virulence and colonization factor of Alternaria alternata during plant infection. Mol. Microbiol. 2019, 112, 131–146. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Strasser, R.J.; Qiang, S. In vivo assessment of effect of phytotoxin tenuazonic acid on PSII reaction centers. Plant Physiol. Biochem. 2014, 84, 10–21. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Contaminants in the Food Chain (Contam). Scientific opinion on the risks for animal and public health related to the presence of Alternaria toxins in feed and food. EFSA J. 2011, 9, 2407. [Google Scholar] [CrossRef]
- EFSA; Arcella, D.; Eskola, M.; Gomez Ruiz, J.A. Dietary exposure assessment to Alternaria toxins in the European population. EFSA J. 2016, 14, e04654. [Google Scholar]
- Ackermann, Y.; Curtui, V.; Dietrich, R.; Gross, M.; Latif, H.; Martlbauer, E.; Usleber, E. Widespread occurrence of low levels of alternariol in apple and tomato products, as determined by comparative immunochemical assessment using monoclonal and polyclonal antibodies. J. Agric. Food Chem. 2011, 59, 6360–6368. [Google Scholar] [CrossRef] [PubMed]
- da Motta, S.; Valente Soares, L.M. Survey of Brazilian tomato products for alternariol, alternariol monomethyl ether, tenuazonic acid and cyclopiazonic acid. J. Food Addit. Contam. 2001, 18, 630–634. [Google Scholar] [CrossRef] [PubMed]
- Noser, J.; Schneider, P.; Rother, M.; Schmutz, H. Determination of six Alternaria toxins with UPLC-MS/MS and their occurrence in tomatoes and tomato products from the Swiss market. Mycotoxin Res. 2011, 27, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Puntscher, H.; Cobankovic, I.; Marko, D.; Warth, B. Quantitation of free and modified Alternaria mycotoxins in European food products by LC-MS/MS. Food Control 2019, 102, 157–165. [Google Scholar] [CrossRef]
- Walravens, J.; Mikula, H.; Rychlik, M.; Asam, S.; Devos, T.; Njumbe Ediage, E.; Diana Di Mavungu, J.; Jacxsens, L.; Van Landschoot, A.; Vanhaecke, L.; et al. Validated UPLC-MS/MS methods to quantitate free and conjugated Alternaria toxins in commercially available tomato products and fruit and vegetable juices in Belgium. J. Agric. Food Chem. 2016, 64, 5101–5109. [Google Scholar] [CrossRef] [PubMed]
- Terminiello, L.; Patriarca, A.; Pose, G.; Fernandez Pinto, V.F. Occurrence of alternariol, alternariol monomethyl ether and tenuazonic acid in Argentinean tomato puree. Mycotoxin Res. 2006, 22, 236–240. [Google Scholar] [CrossRef] [PubMed]
- EU Commission. Recommendation (EC) no 553/2022 of 5 April 2022 related to Alternaria toxins in certain foodstuffs. Off. J. Eur. Union 2022, L107, 90–92. [Google Scholar]
- Salotti, I.; Giorni, P.; Battilani, P. Biology, ecology, and epidemiology of Alternaria species affecting tomato: Ground information for the development of a predictive model. Front. Plant Sci. 2024, 15, 1430965. [Google Scholar] [CrossRef] [PubMed]
- Pose, G.; Patriarca, A.; Kyanko, V.; Pardo, A.; Pinto, V.F. Water activity and temperature effects on mycotoxin production by Alternaria alternata on a synthetic tomato medium. Int. J. Food Microbiol. 2010, 142, 348–353. [Google Scholar] [CrossRef] [PubMed]
- Vaquera, S.; Patriarca, A.; Pinto, V.F. Influence of environmental parameters on mycotoxin production by Alternaria arborescens. Int. J. Food Microbiol. 2016, 219, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Patriarca, A.; Medina, A.; Pinto, V.F.; Magan, N. Temperature and water stress impacts on growth and production of altertoxin-II by strains of Alternaria tenuissima from Argentinean wheat. World Mycotoxin J. 2014, 7, 329–334. [Google Scholar] [CrossRef]
- Scott, P.M. Analysis of agricultural commodities and foods for Alternaria mycotoxins. J. AOAC Int. 2001, 84, 1809–1817. [Google Scholar] [CrossRef] [PubMed]
- Bellotti, G.; Guerrieri, M.C.; Giorni, P.; Bulla, G.; Fiorini, A.; Bertuzzi, T.; Antinori, M.E.; Puglisi, E. Enhancing plant defense using rhizobacteria in processing tomatoes: A bioprospecting approach to overcoming Early Blight and Alternaria toxins. Front. Microbiol. 2023, 14, 1221633. [Google Scholar] [CrossRef] [PubMed]
- Giorni, P.; Bulla, G.; Bellotti, G.; Antinori, M.E.; Guerrieri, M.C.; Fiorini, A.; Bertuzzi, T.; Puglisi, E. In planta evaluation of different bacterial consortia for the protection of tomato plants against Alternaria spp. infection and Alternaria toxins presence in fruits. Front. Hortic. 2024, 3, 1447425. [Google Scholar] [CrossRef]
- Lee, H.B.; Patriarca, A. Alternaria in food: Ecophysiology, mycotoxin production and toxicology. Mycobiology 2015, 43, 93–106. [Google Scholar] [CrossRef] [PubMed]
- Romero Donato, C.J.; Nichea, M.J.; Cendoya, E.; Zachetti, V.G.L.; Ramirez, M.L. Interacting abiotic factors affect growth and mycotoxin production profiles of Alternaria section Alternaria strains on chickpea-based media. Pathogens 2023, 12, 565. [Google Scholar] [CrossRef] [PubMed]
- Bessadat, N.; Berruyer, R.; Hamon, B.; Bataille-Simoneau, N.; Benichou, S.; Kihal, M.; Henni, D.E.; Simoneau, P. Alternaria species associated with early blight epidemics on tomato and other Solanaceae crops in northwestern Algeria. European J. Plant Pathol. 2017, 148, 181–197. [Google Scholar] [CrossRef]
- Alhussaen, K.M. Morphological and physiological characterization of Alternaria solani isolated from tomato in Jordan valley. Res. J. Biol. Sci. 2012, 7, 316–319. [Google Scholar] [CrossRef]
- Naik, M.K.; Prasad, Y.; Bhat, K.V.; Rani, G.D. Morphological, physiological, pathogenic and molecular variability among isolates of Alternaria solani from tomato. Indian Phytopathol. 2010, 63, 168173. [Google Scholar]
- Pose, G.; Patriarca, A.; Kyanko, V.; Pardo, A.; Pinto, V.F. Effect of water activity and temperature on growth of Alternaria alternata on a synthetic tomato medium. Int. J. Food Microbiol. 2009, 135, 60–63. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, K.P.; Shakir, A.S.; Sahi, S.T. Physiological studies on the fungi causing post-harvest losses of tomato fruits. Pak. J. Phytopathol. 1999, 11, 25–29. [Google Scholar]
- Malathrakis, N.E. Alternaria stem canker of tomato in Greece. Phytopathol. Mediterr. 1983, 22, 33–38. [Google Scholar]
- Delgado-Baquerizo, M.; Guerra, C.A.; Cano-Díaz, C.; Egidi, E.; Wang, J.T.; Eisenhauer, N.; Singh, B.K.; Maestre, F.T. The proportion of soil-borne pathogens increases with warming at the global scale. Nat. Clim. Chang. 2020, 10, 550–554. [Google Scholar] [CrossRef]
- Van de Perre, E.; Jacxsens, L.; Liu, C.; Devlieghere, F.; De Meulenaer, B. Climate impact on Alternaria moulds and their mycotoxins in fresh produce: The case of the tomato chain. Food Res. Int. 2015, 68, 41–46. [Google Scholar] [CrossRef]
- Battilani, P. Recent advances in modeling the risk of mycotoxin contamination in crops. Curr. Opin. Food Sci. 2016, 11, 10–15. [Google Scholar] [CrossRef]
- De Wolf, E.D.; Isard, S.A. Disease cycle approach to plant disease prediction. Annu. Rev. Phytopathol. 2007, 45, 203–220. [Google Scholar] [CrossRef] [PubMed]
- Rossi, V.; Caffi, T.; Salotti, I.; Fedele, G. Sharing decision-making tools for pest management may foster implementation of Integrated Pest Management. Food Secur. 2023, 15, 1459–1474. [Google Scholar] [CrossRef]
- González-Domínguez, E.; Caffi, T.; Rossi, V.; Salotti, I.; Fedele, G. Plant disease models and forecasting: Changes in principles and applications over the last 50 years. Phytopathology 2023, 113, 678–693. [Google Scholar] [CrossRef] [PubMed]
- Vandecasteele, M.; Landschoot, S.; Carrette, J.; Verwaeren, J.; Höfte, M.; Audenaert, K.; Haesaert, G. Species prevalence and disease progres sion studies demonstrate a seasonal shift in the Alternaria population composition on potato. Plant Pathol. 2018, 67, 327–336. [Google Scholar] [CrossRef]
- Salotti, I.; Camardo Leggieri, M.; Battilani, P. Engaging complexity of Alternaria-tomato pathosystem: From ecological studies to the development of a mechanistic model. In Proceedings of the 14th Conference of the European Foundation for Plant Pathology, Uppsala, Sweden, 2–5 June 2025. [Google Scholar]
- Rabaaoui, A.; Masiello, M.; Somma, S.; Crudo, F.; Dall’Asta, C.; Righetti, L.; Susca, A.; Logrieco, A.F.; Namsi, A.; Gdoura, R.; et al. Phylogeny and mycotoxin profiles of pathogenic Alternaria and Curvularia species isolated from date palm in southern Tunisia. Front. Microbiol. 2022, 13, 1034658. [Google Scholar] [CrossRef] [PubMed]
- Analytis, S. PÜber die Relation zwischen biologischer Entwicklung und Temperatur bei phytopathogenen Pilzen. Phytopathol. Z. 1977, 90, 64–76. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference. A Practical Information-Theoretic Approach; Springer: New York, NY, USA, 2002. [Google Scholar]
- Wickham, H. Modelr: Modelling Functions That Work with the Pipe, R Package, Version 0.1; 2019; p. 4. Available online: https://rdrr.io/cran/modelr/man/modelr-package.html (accessed on 16 April 2025).
- Nash, J.E.; Sutcliffe, J.V. River Flow Forecasting Through Conceptual Models Part 1: A Discussion of Principles. J. Hydrol. 1970, 10, 282–290. [Google Scholar] [CrossRef]
- Lin, L.I. A Concordance Correlation Coefficient to Evaluate Reproducibility. Biometrics 1989, 45, 255–268. [Google Scholar] [CrossRef] [PubMed]
- Signorell, A. DescTools: Tools for Descriptive Statistics. R Package, Version 0.99; 2020; p. 38. Available online: https://cran.r-project.org/package=DescTools (accessed on 16 April 2025).


{kind=link}
{kind=link}
{kind=link}
| Species | Tmin | Tmax | Topt | a | b | c | R2 | CCC | RMSE | CRM |
|---|---|---|---|---|---|---|---|---|---|---|
| A. alternata | 0 | 40 | 25.6 | 6.131 (0.342) | 1.771 (0.108) | 1.297 (0.183) | 0.857 | 0.927 | 0.132 | 0.023 |
| A. solani | 0 | 40 | 26.5 | 6.655 (0.353) | 1.969 (0.115) | 1.342 (0.262) | 0.807 | 0.898 | 0.147 | 0.053 |
| A. tenuissima | 0 | 40 | 25.2 | 5.975 (0.270) | 1.703 (0.082) | 1.485 (0.167) | 0.897 | 0.948 | 0.117 | 0.021 |
| Temperature | A. alternata | A. solani | A. tenuissima |
|---|---|---|---|
| Alternariol (AOH) | |||
| 5 °C | 2.4 ± 1.1 e | n.d. | 1.3 ± 0.7 e |
| 10 °C | 56.8 ± 4.5 d | 74.0 ± 5.9 cd | 26.0 ± 3.6 d |
| 15 °C | 124.7 ± 13.8 bcd | 221.7 ± 18.2 abcd | 528.0 ± 34.2 abc |
| 20 °C | 289.3 ± 28.1 abcd | 719.5 ± 47.8 ab | 712.5 ± 36.1 ab |
| 25 °C | 914.8 ± 44.0 a | 1251.3 ± 74.8 a | 510.3 ± 35.9 abc |
| 30 °C | 1067.7 ± 44.1 a | 857.5 ± 34.2 ab | 79.0 ± 8.2 cd |
| 35 °C | 755.2 ± 41.3 ab | 303.5 ± 16.5 abcd | 32.8 ± 5.9 d |
| Alternariol monomethyl ether (AME) | |||
| 5 °C | 2.7 ± 1.5 g | n.d. | 1.3 ± 0.9 g |
| 10 °C | 27.3 ± 2.9 ef | 36.0 ± 4.8 def | 14.8 ± 1.8 f |
| 15 °C | 81.8 ± 4.9 bcdef | 111.0 ± 8.5 abcde | 301.0 ± 58.9 abcd |
| 20 °C | 215.3 ± 38.9 abcd | 376.2 ± 33.1 ab | 412.0 ± 25.9 ab |
| 25 °C | 562.5 ± 25.9 a | 639.3 ± 38.0 a | 317.7 ± 16.3 abc |
| 30 °C | 646.8 ± 24.4 a | 704.5 ± 13.9 a | 63.0 ± 9.2 cdef |
| 35 °C | 530.0 ± 26.1 a | 316.0 ± 49.0 abcd | 49.5 ± 7.1 def |
| Tenuazonic acid (TeA) | |||
| 5 °C | n.d. | n.d. | n.d. |
| 10 °C | 14.8 ± 9.5 f | 45.2 ± 3.6 ef | 54.2 ± 6.5 ef |
| 15 °C | 80.8 ± 6.5 cdef | 91.8 ± 8.0 bcdef | 510.0 ± 24.7 abc |
| 20 °C | 184.7 ± 20.2 abcdef | 192.3 ± 18.6 abcdef | 636.7 ± 38.7 ab |
| 25 °C | 435.7 ± 31.9 abcd | 402.2 ± 29.9 abcd | 721.2 ± 71.6 a |
| 30 °C | 562.5 ± 52.5 abc | 366.5 ± 90.4 abcdef | 137.0 ± 18.2 abcdef |
| 35 °C | 403.0 ± 87.7 abcde | 268.5 ± 169.5 abcdef | 69.2 ± 10.4 def |
| Species | Tmin | Tmax | Topt | a | b | c | R2 | CCC | RMSE | CRM |
|---|---|---|---|---|---|---|---|---|---|---|
| Alternariol (AOH) | ||||||||||
| A. alternata | 5 | 40 | 30.1 | 8.237 (0.501) | 2.544 (0.155) | 1.803 (0.312) | 0.979 | 0.991 | 0.052 | 0.019 |
| A. solani | 5 | 40 | 25.9 | 5.339 (0.187) | 1.482 (0.062) | 3.231 (0.408) | 0.985 | 0.993 | 0.041 | 0.024 |
| A. tenuissima | 5 | 40 | 19.8 | 3.252 (0.091) | 0.728 (0.030) | 5.832 (0.811) | 0.982 | 0.992 | 0.049 | −0.010 |
| Alternariol monomethyl ether (AME) | ||||||||||
| A. alternata | 5 | 40 | 30.7 | 8.833 (0.592) | 2.748 (0.183) | 1.386 (0.246) | 0.984 | 0.993 | 0.046 | 0.008 |
| A. solani | 5 | 40 | 28.0 | 6.522 (0.131) | 1.917 (0.041) | 2.180 (0.126) | 0.997 | 0.999 | 0.020 | 0.011 |
| A. tenuissima | 5 | 40 | 20.1 | 3.323 (0.131) | 0.755 (0.043) | 5.288 (0.972) | 0.966 | 0.985 | 0.066 | 0.013 |
| Tenuazonic acid (TeA) | ||||||||||
| A. alternata | 5 | 40 | 30.0 | 8.075 (0.228) | 2.490 (0.074) | 1.640 (0.133) | 0.996 | 0.998 | 0.023 | 0.013 |
| A. solani | 5 | 40 | 28.4 | 6.626 (0.603) | 2.016 (0.194) | 1.526 (0.373) | 0.951 | 0.979 | 0.072 | 0.019 |
| A. tenuissima | 5 | 40 | 20.9 | 3.529 (0.233) | 0.829 (0.077) | 4.322 (1.196) | 0.922 | 0.966 | 0.102 | −0.002 |
| Code | Species | Country of Origin | Isolation Substrate |
|---|---|---|---|
| CBS 118814 | A. alternata | United States of America, Florida, Quincy | Solanum lycopersicum |
| CBS 109157 | A. solani | United States of America, Washington, Douglas | Solanum tuberosum |
| CBS 117.44 | A. tenuissima | Denmark, Sjaelland, Clausdal | Clarkia spp. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salotti, I.; Giorni, P.; Dall’Asta, C.; Battilani, P. Modeling Temperature Requirements for Growth and Toxin Production of Alternaria spp. Associated with Tomato. Toxins 2025, 17, 361. https://doi.org/10.3390/toxins17080361
Salotti I, Giorni P, Dall’Asta C, Battilani P. Modeling Temperature Requirements for Growth and Toxin Production of Alternaria spp. Associated with Tomato. Toxins. 2025; 17(8):361. https://doi.org/10.3390/toxins17080361
Chicago/Turabian StyleSalotti, Irene, Paola Giorni, Chiara Dall’Asta, and Paola Battilani. 2025. "Modeling Temperature Requirements for Growth and Toxin Production of Alternaria spp. Associated with Tomato" Toxins 17, no. 8: 361. https://doi.org/10.3390/toxins17080361
APA StyleSalotti, I., Giorni, P., Dall’Asta, C., & Battilani, P. (2025). Modeling Temperature Requirements for Growth and Toxin Production of Alternaria spp. Associated with Tomato. Toxins, 17(8), 361. https://doi.org/10.3390/toxins17080361