Combination of Rhamnetin and RXP03 Mitigates Venom-Induced Toxicity in Murine Models: Preclinical Insights into Dual-Target Antivenom Therapy


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
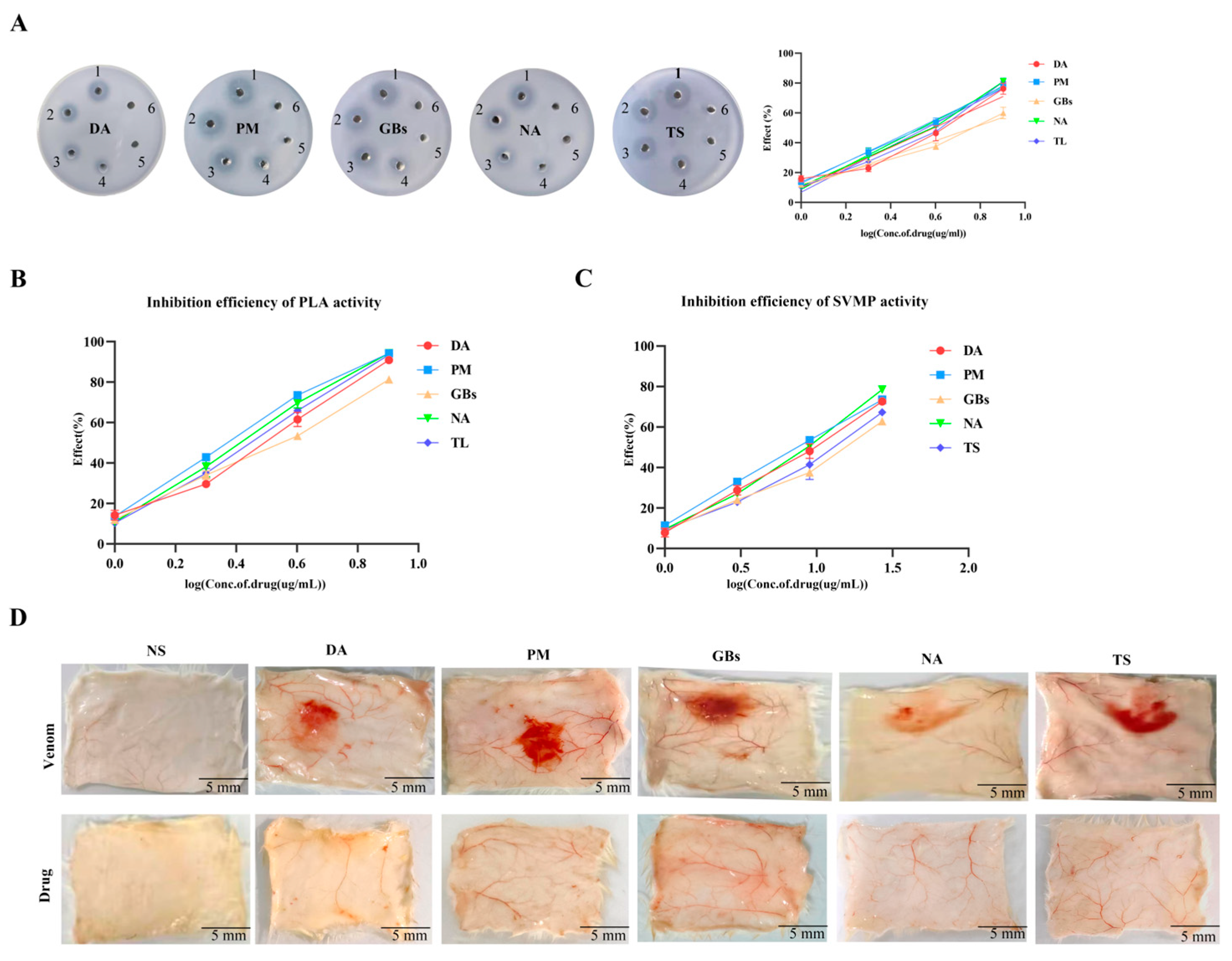
2.1. Inhibitory Effect of Venom
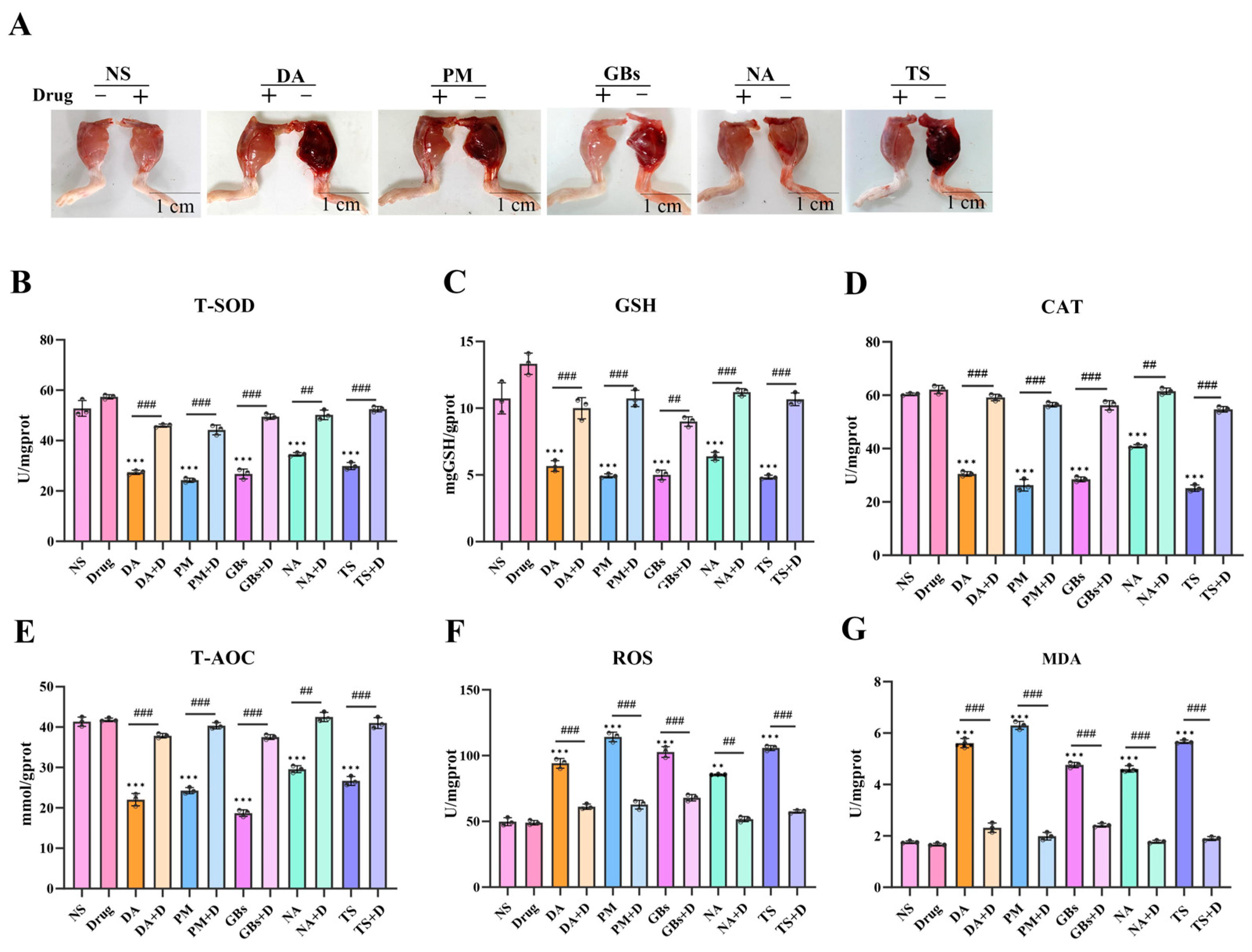
2.2. Rhamnetin and RXP03 Mitigated Venom-Induced Myotoxic Activity
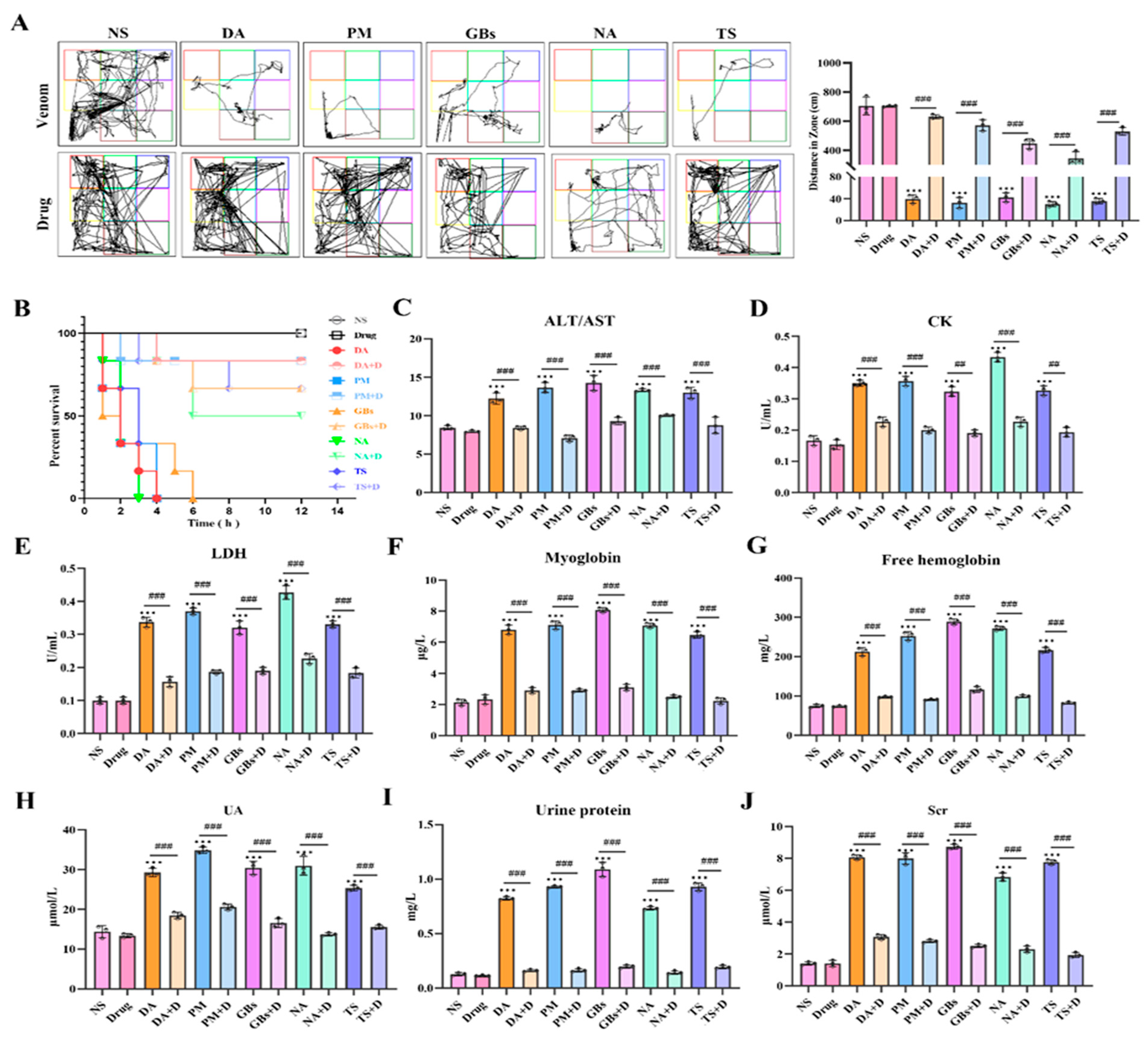
2.3. Rhamnetin and RXP03 Ablated Systemic Toxicity Induced by Venom
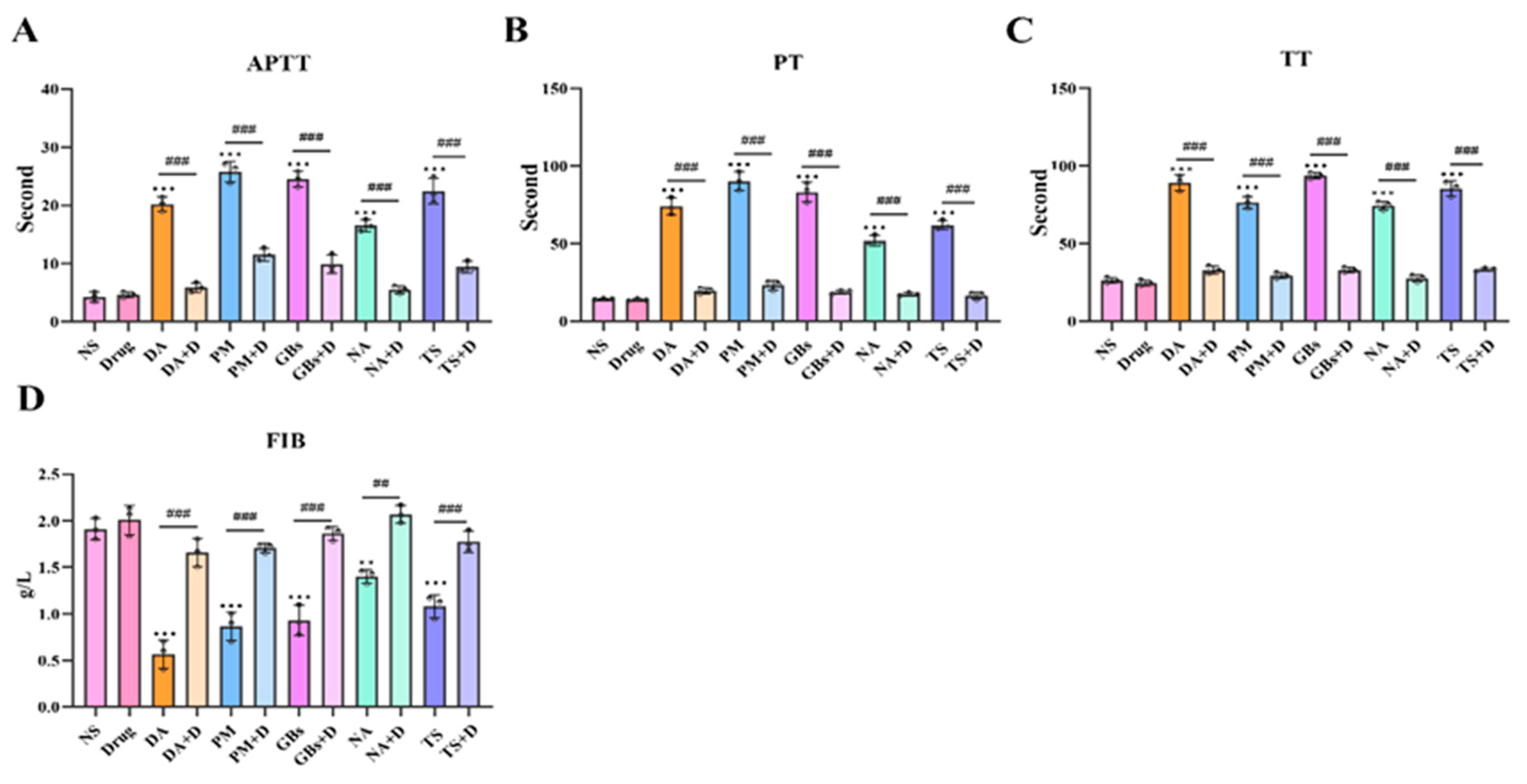
2.4. Rhamnetin and RXP03 Prevented Venom-Induced Hemostatic Dysfunction
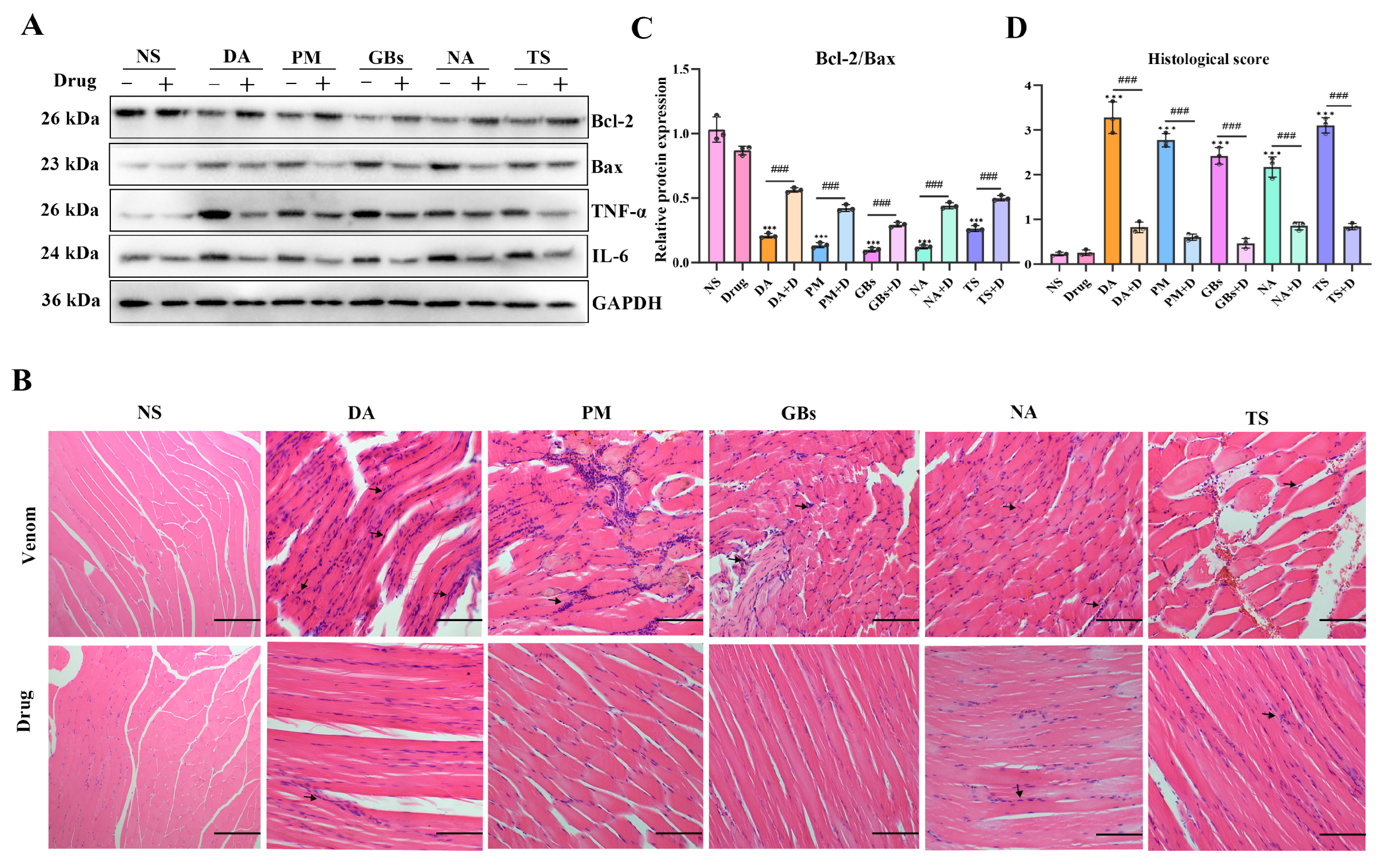
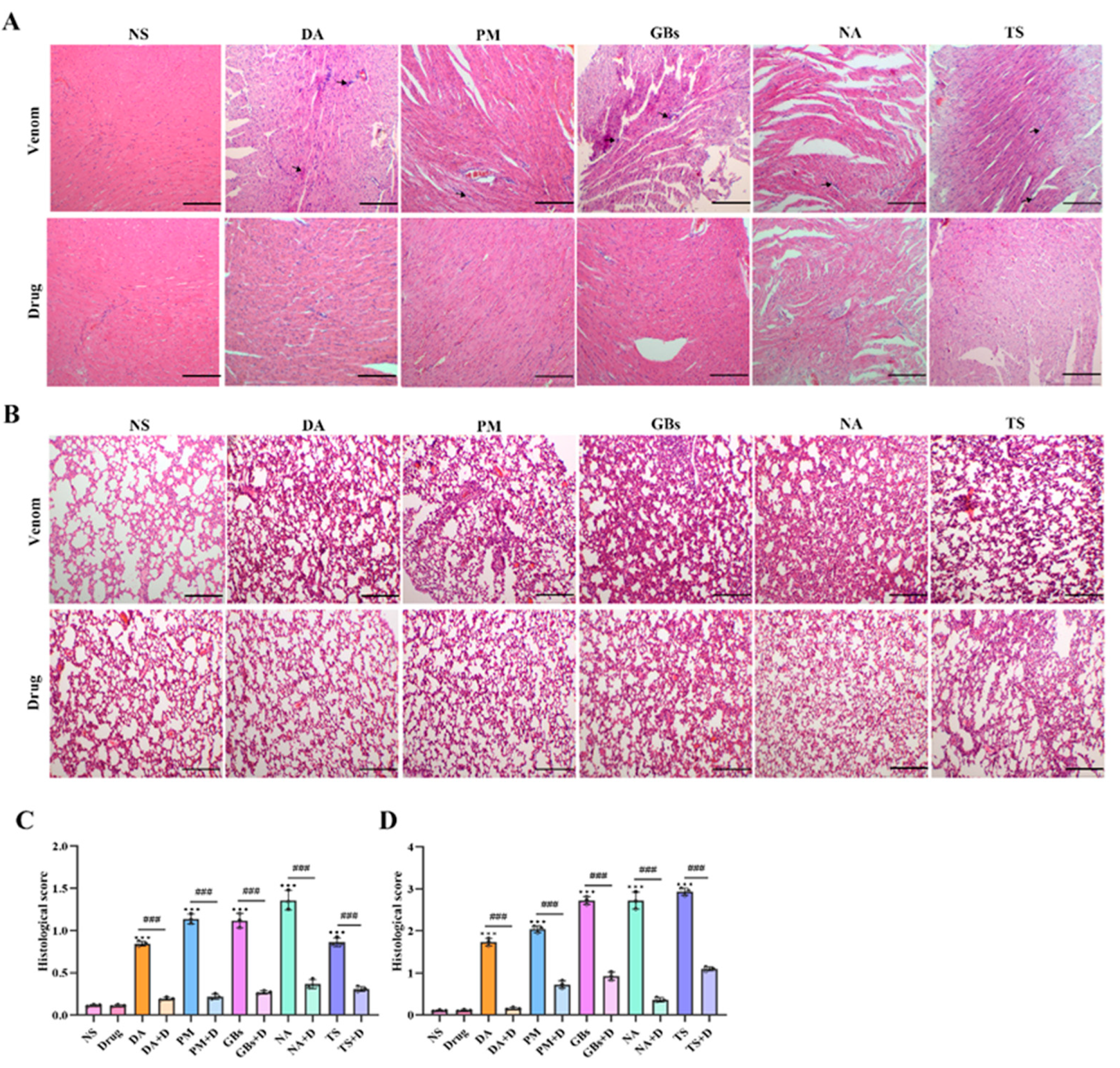
2.5. Rhamnetin and RXP03 Attenuated Histopathological Destruction Caused by Venom
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. The Snake Venom and Main Reagents
5.2. PLA2 Activity
5.3. SVMP Activity
5.4. Animals Model and Ethics
5.5. Hemorrhagic Activity
5.6. Myotoxic Activity
5.7. Venom-Induced Systemic Toxicity
5.8. Ser-Enzyme, Urine Protein, and Oxidative Stress Assays
5.9. Evaluation of Hemostatic Parameters
5.10. Western Blot
5.11. Histological Analysis
5.12. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gutiérrez, J.M.; Calvete, J.J.; Habib, A.G.; Harrison, R.A.; Williams, D.J.; Warrell, D.A. Snakebite envenoming. Nat. Rev. Dis. Prim. 2017, 3, 17063. [Google Scholar] [CrossRef]
- Kasturiratne, A.; Wickremasinghe, A.R.; de Silva, N.; Gunawardena, N.K.; Pathmeswaran, A.; Premaratna, R.; Savioli, L.; Lalloo, D.G.; de Silva, H.J. The Global Burden of Snakebite: A Literature Analysis and Modelling Based on Regional Estimates of Envenoming and Deaths. PLoS Med. 2008, 5, e218. [Google Scholar] [CrossRef] [PubMed]
- Molyneux, D.H. The London Declaration on Neglected Tropical Diseases: 5 years on. Trans. R. Soc. Trop. Med. Hyg. 2016, 110, 623–625. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.J.; Abul Faiz, M.; Abela-Ridder, B.; Ainsworth, S.; Bulfone, T.C.; Nickerson, A.D.; Habib, A.G.; Junghanss, T.; Fan, H.W.; Turner, M.; et al. Strategy for a globally coordinated response to a priority neglected tropical disease: Snakebite envenoming. PLoS Neglect. Trop. Dis. 2019, 13, e0007059. [Google Scholar] [CrossRef] [PubMed]
- Brown, N.; Landon, J. Antivenom: The Most Cost-effective Treatment in the World? Toxicon 2010, 55, 1405–1407. [Google Scholar] [CrossRef]
- Casewell, N.R.; Jackson, T.N.W.; Laustsen, A.H.; Sunagar, K. Causes and Consequences of Snake Venom Variation. Trends Pharmacol. Sci. 2020, 41, 570–581. [Google Scholar] [CrossRef]
- de Silva, H.A.; Ryan, N.M.; de Silva, H.J. Adverse Reactions to Snake Antivenom, and their Prevention and Treatment. Br. J. Clin. Pharmacol. 2016, 81, 446–452. [Google Scholar] [CrossRef]
- Harrison, R.A.; Oluoch, G.O.; Ainsworth, S.; Alsolaiss, J.; Bolton, F.; Arias, A.S.; Gutiérrez, J.M.; Rowley, P.; Kalya, S.; Ozwara, H.; et al. Preclinical Antivenom-efficacy Testing Reveals Potentially Disturbing Deficiencies of Snakebite Treatment Capability in East Africa. PLoS Neglect. Trop. Dis. 2017, 11, e0005969. [Google Scholar] [CrossRef]
- Clare, R.H.; Hall, S.R.; Patel, R.N.; Casewell, N.R. Small Molecule Drug Discovery for Neglected Tropical Snakebite. Trends Pharmacol. Sci. 2021, 42, 340–353. [Google Scholar] [CrossRef]
- Albulescu, L.O.; Xie, C.F.; Ainsworth, S.; Alsolaiss, J.; Crittenden, E.; Dawson, C.A.; Softley, R.; Bartlett, K.E.; Harrison, R.A.; Kool, J.; et al. A Therapeutic Combination of Two Small Molecule Toxin Inhibitors Provides Broad Preclinical Efficacy Against Viper Snakebite. Nat. Commun. 2021, 12, 4027. [Google Scholar] [CrossRef]
- Bulfone, T.C.; Samuel, S.P.; Bickler, P.E.; Lewin, M.R. Developing Small Molecule Therapeutics for the Initial and Adjunctive Treatment of Snakebite. J. Trop. Med. 2018, 2018, 4320175. [Google Scholar] [CrossRef] [PubMed]
- Layfield, H.J.; Williams, H.E.; Ravishankar, D.; Mehmi, A.; Sonavane, M.; Salim, A.; Vaiyapuri, R.; Lakshminarayanan, K.; Vallance, T.M.; Bicknell, A.B.; et al. Repurposing Cancer Drugs Batimastat and Marimastat to Inhibit the Activity of a Group I Metalloprotease from the Venom of the Western Diamondback Rattlesnake, Crotalus atrox. Toxins 2020, 12, 309. [Google Scholar] [CrossRef] [PubMed]
- Lewin, M.; Samuel, S.; Merkel, J.; Bickler, P. Varespladib (LY315920) Appears to Be a Potent, Broad-Spectrum, Inhibitor of Snake Venom Phospholipase A2 and a Possible Pre-Referral Treatment for Envenomation. Toxins 2016, 8, 248. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.D.; Zhang, J.; Zhang, D.H.; Xiao, H.X.; Xiong, S.W.; Huang, C.H. Exploration of the Inhibitory Potential of Varespladib for Snakebite Envenomation. Molecules 2022, 27, 5269. [Google Scholar] [CrossRef]
- Xie, C.F.; Albulescu, L.O.; Bittenbinder, M.A.; Somsen, G.W.; Vonk, F.J.; Casewell, N.R.; Kool, J. Neutralizing Effects of Small Molecule Inhibitors and Metal Chelators on Coagulopathic Viperinae Snake Venom Toxins. Biomedicines 2020, 8, 297. [Google Scholar] [CrossRef]
- Rucavado, A.; Escalante, T.; Franceschi, A.; Chaves, F.; León, G.; Cury, Y.; Ovadia, M.; Gutiérrez, J.M. Inhibition of Local Hemorrhage and Dermonecrosis Induced by Snake Venom: Effectiveness of Early Administration of the Peptidomimetic Metalloproteinase Inhibitor Batimastat and the Chelating Agent CaNa EDTA. Am. J. Trop. Med. Hyg. 2000, 63, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Hall, S.R.; Rasmussen, S.A.; Crittenden, E.; Dawson, C.A.; Bartlett, K.E.; Westhorpe, A.P.; Albulescu, L.O.; Kool, J.; Gutiérrez, J.M.; Casewell, N.R. Repurposed Drugs and Their Combinations Prevent Morbidity-Inducing Dermonecrosis Caused by Diverse Cytotoxic Snake Venoms. Nat. Commun. 2023, 14, 7812. [Google Scholar] [CrossRef]
- Ferraz, C.R.; Arrahman, A.; Xie, C.F.; Casewell, N.R.; Lewis, R.J.; Kool, J.; Cardoso, F.C. Multifunctional Toxins in Snake Venoms and Therapeutic Implications: From Pain to Hemorrhage and Necrosis. Front. Ecol. Evol. 2019, 7, 218. [Google Scholar] [CrossRef]
- Gutiérrez, J.M.; Escalante, T.; Rucavado, A.; Herrera, C. Hemorrhage Caused by Snake Venom Metalloproteinases: A Journey of Discovery and Understanding. Toxins 2016, 8, 93. [Google Scholar] [CrossRef]
- Li, S.T.; Wang, J.Q.; Zhang, X.M.; Ren, Y.; Wang, N.; Zhao, K.; Chen, X.S.; Zhao, C.F.; Li, X.L.; Shao, J.M.; et al. Proteomic Characterization of Two Snake Venoms: Naja Naja atra and Agkistrodon halys. Biochem. J. 2004, 384, 119–127. [Google Scholar] [CrossRef]
- Liu, C.C.; Lin, C.C.; Hsiao, Y.C.; Wang, P.J.; Yu, J.S. Proteomic Characterization of Six Taiwanese Snake Venoms: Identification of Species-Specific Proteins and Development of a SISCAPA-MRM Assay for Cobra Venom Factors. J. Proteom. 2018, 187, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.C.; Wu, C.J.; Hsiao, Y.C.; Yang, Y.H.; Liu, K.L.; Huang, G.J.; Hsieh, C.H.; Chen, C.K.; Liaw, G.W. Snake Venom Proteome of Protobothrops mucrosquamatus in Taiwan: Delaying Venom-Induced Lethality in a Rodent Model by Inhibition of Phospholipase A2 Activity with Varespladib. J. Proteom. 2021, 234, 104084. [Google Scholar] [CrossRef] [PubMed]
- Tasoulis, T.; Isbister, G.K. A Current Perspective on Snake Venom Composition and Constituent Protein Families. Arch. Toxicol. 2023, 97, 133–153. [Google Scholar] [CrossRef] [PubMed]
- Jones, B.K.; Saviola, A.J.; Reilly, S.B.; Stubbs, A.L.; Arida, E.; Iskandar, D.T.; McGuire, J.A.; Yates, J.R.; Mackessy, S.P. Venom Composition in a Phenotypically Variable Pit Viper (Trimeresurus insularis) Across the Lesser Sunda Archipelago. J. Proteome Res. 2019, 18, 2206–2220. [Google Scholar] [CrossRef]
- Fu, K.P.; Cao, L.Y.; Tang, Y.T.; Zhao, J.Q.; Xiong, K.J.; Hong, C.J.; Huang, C.H. The Anti-myotoxic Effects and Mechanisms of Sinonatrix annularis Serum and a Novel Plasma Metalloproteinase Inhibitor Towards Deinagkistrodon acutus Envenomation. Toxicol. Lett. 2023, 388, 13–23. [Google Scholar] [CrossRef]
- Zhao, W.J.; Liu, J.H.; Wang, S.D.; Tao, Q.Q.; Lei, Q.Q.; Huang, C.H. Varespladib Mitigates Acute Liver Injury via Suppression of Excessive Mitophagy on Naja atra Envenomed Mice by Inhibiting PLA2. Toxicon 2024, 242, 107694. [Google Scholar] [CrossRef]
- Medeiros, D.L.; Lima, E.T.G.; Silva, J.C.; Medeiros, M.A.; Pinheiro, E.B.F. Rhamnetin: A Review of Its Pharmacology and Toxicity. J. Pharm. Pharmacol. 2022, 74, 793–799. [Google Scholar] [CrossRef]
- Belchor, M.N.; Gaeta, H.H.; Rodrigues, C.F.B.; Costa, C.R.D.; Toyama, D.D.; Passero, L.F.D.; Laurenti, M.D.; Toyama, M.H. Evaluation of Rhamnetin as an Inhibitor of the Pharmacological Effect of Secretory Phospholipase A2. Molecules 2017, 22, 1441. [Google Scholar] [CrossRef]
- Abdou, M.M.; Ötvös, F.; Dong, D.W.; Matziari, M. Novel glycosyl prodrug of RXP03 as MMP-11 Prodrug: Design, Synthesis and Virtual Screening. BMC Chem. 2023, 17, 167. [Google Scholar] [CrossRef]
- Matziari, M.; Dellis, D.; Dive, V.; Yiotakis, A.; Samios, J. Conformational and Solvation Studies via Computer Simulation of the Novel Large Scale Diastereoselectively Synthesized Phosphinic MMP Inhibitor RXP03 Diluted in Selected Solvents. J. Phys. Chem. B 2010, 114, 421–428. [Google Scholar] [CrossRef]
- Huang, F.; Zhao, S.Q.; Tong, F.; Liang, Y.; Le Grange, J.M.; Kuang, W.H.; Zhou, Y.W. Unexpected Death in a Young Man Associated with a Unilateral Swollen Leg: Pathological and Toxicological Findings in a Fatal Snakebite from Deinagkistrodon acutus (Chinese moccasin). J. Forensic Sci. 2021, 66, 786–792. [Google Scholar] [CrossRef]
- Kini, R.M.; Sidhu, S.S.; Laustsen, A.H. Biosynthetic Oligoclonal Antivenom (BOA) for Snakebite and Next-Generation Treatments for Snakebite Victims. Toxins 2018, 10, 534. [Google Scholar] [CrossRef] [PubMed]
- Sharma, N.; Chauhan, S.; Faruqi, S.; Bhat, P.; Varma, S. Snake Envenomation in a North Indian hospital. Emerg. Med. J. 2005, 22, 118–120. [Google Scholar] [CrossRef] [PubMed]
- Abouyannis, M.; Boga, M.; Amadi, D.; Ouma, N.; Nyaguara, A.; Mturi, N.; Berkley, J.A.; Adetifa, I.M.; Casewell, N.R.; Lalloo, D.G.; et al. A Long-Term Observational Study of Paediatric Snakebite in Kilifi County, South-east Kenya. PLoS Neglect. Trop. Dis. 2023, 17, e0010987. [Google Scholar] [CrossRef]
- Carregari, V.C.F.R.; Rodrigues-Simioni, L.; Winck, F.V.; Baldasso, P.A.; Ponce-Soto, L.A.; Marangoni, S. Biochemical, Pharmacological, and Structural Characterization of New Basic PLA2 Bbil-TX from Bothriopsis bilineata Snake Venom. Biomed. Res. Int. 2013, 2013, 612649. [Google Scholar] [CrossRef]
- Oyama, E.; Takahashi, H. Structures and Functions of Snake Venom Metalloproteinases (SVMP) from Protobothrops Venom Collected in Japan. Molecules 2017, 22, 1305. [Google Scholar] [CrossRef]
- Qin, W.G.; Zhuo, Z.P.; Hu, H.; Lay, M.M.; Li, Q.Q.; Huang, J.T.; Zeng, L.B.; Liang, Z.J.; Long, F.; Liang, Q. Proteomic Characteristics of Six Snake Venoms from the Viperidae and Elapidae Families in China and Their Relation to Local Tissue Necrosis. Toxicon 2023, 235, 107317. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Chen, F.; Wu, S.K. The Related Risk Factors Analysis of Snake-Bite Induced Acute Kidney Injury. Med. Sci. Monit. 2016, 22, 2335–2339. [Google Scholar] [CrossRef]
- Zhang, M.L.; Cheng, J.J.; Sun, Z.W.; Kong, H.; Zhang, Y.; Wang, S.N.; Wang, X.K.; Zhao, Y.; Qu, H.H. Protective Effects of Carbon Dots Derived from Phellodendri Chinensis Cortex Carbonisata Against Deinagkistrodon acutus Venom-Induced Acute Kidney Injury. Nanoscale Res. Lett. 2019, 14, 377. [Google Scholar] [CrossRef]
- Sitprija, V.; Sitprija, S. Renal effects and injury induced by animal toxins. Toxicon 2012, 60, 943–953. [Google Scholar] [CrossRef]
- Fu, K.P.; Zhao, J.Q.; Zhong, L.P.; Xu, H.Y.; Yu, X.H.; Bi, X.W.; Huang, C.H. Dual Therapy with Phospholipase and Metalloproteinase Inhibitors from Sinonatrix annularis Alleviated Acute Kidney and Liver Injury Caused by Multiple Snake Venoms. Biomed. Pharmacother. 2024, 177, 116967. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, S.; Sinha, R.; Chaudhury, A.R.; Maduwage, K.; Abeyagunawardena, A.; Bose, N.; Pradhan, S.; Bresolin, N.L.; Garcia, B.A.; McCulloch, M. Snake Bite Associated with Acute Kidney Injury. Pediatr. Nephrol. 2020, 36, 3829–3840. [Google Scholar] [CrossRef] [PubMed]
- Benati, R.B.; Costa, T.R.; Cacemiro, M.D.; Sampaio, S.V.; de Castro, F.A.; Burin, S.M. Cytotoxic and Pro-apoptotic Action of MjTX-I, a Phospholipase A2 Isolated from Bothrops moojeni Snake Venom, Towards Leukemic Cells. J. Venom. Anim. Toxins 2018, 24, 40. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.A.; Ilies, M.A. The Phospholipase A2 Superfamily: Structure, Isozymes, Catalysis, Physiologic and Pathologic Roles. Int. J. Mol. Sci. 2023, 24, 1353. [Google Scholar] [CrossRef]
- Chen, Y.J.; Cui, T.Q.; Xiao, S.R.; Li, T.Y.; Zhong, Y.D.; Tang, K.J.; Guo, J.Y.; Huang, S.Y.; Chen, J.B.; Li, J.Y.; et al. Hepatic ZBTB22-mediated Detoxification Ameliorates Acetaminophen-induced Liver Injury by Inhibiting Pregnane X Receptor Signaling. iScience 2023, 26, 106318. [Google Scholar] [CrossRef]
- Dutra, F.F.; Alves, L.S.; Rodrigues, D.; Fernandez, P.L.; de Oliveira, R.B.; Golenbock, D.T.; Zamboni, D.S.; Bozza, M.T. Hemolysis-induced Lethality Involves Inflammasome Activation by Heme. Proc. Natl. Acad. Sci. USA 2014, 111, E4110–E4118. [Google Scholar] [CrossRef]
- Herrera, C.; Escalante, T.; Voisin, M.B.; Rucavado, A.; Morazán, D.; Macêdo, J.K.A.; Calvete, J.J.; Sanz, L.; Nourshargh, S.; Gutiérrez, J.M.; et al. Tissue Localization and Extracellular Matrix Degradation by PI, PII and PIII Snake Venom Metalloproteinases: Clues on the Mechanisms of Venom-Induced Hemorrhage. PLoS Neglect. Trop. Dis. 2015, 9, e0003731. [Google Scholar] [CrossRef]
- Silveira, L.B.; Marchi-Salvador, D.P.; Santos, N.A.; Silva, F.P.; Marcussi, S.; Fuly, A.L.; Nomizo, A.; da Silva, S.L.; Stábeli, R.G.; Arantes, E.C.; et al. Isolation and Expression of a Hypotensive and Anti-platelet Acidic Phospholipase A2 from Bothrops moojeni Snake Venom. J. Pharm. Biomed. Anal. 2013, 73, 35–43. [Google Scholar] [CrossRef]
- Tiwari, N.; Aggarwal, G.; Jain, G.K.; Mittal, G. Multi-drug Loaded Microneedles for Emergency Treatment of Snakebite Envenomation. Med. Hypotheses 2022, 165, 110908. [Google Scholar] [CrossRef]
- Liu, C.C.; Chou, Y.S.; Chen, C.Y.; Liu, K.L.; Huang, G.J.; Yu, J.S.; Wu, C.J.; Liaw, G.W.; Hsieh, C.H.; Chen, C.K. Pathogenesis of Local Necrosis Induced by Naja atra Venom: Assessment of the Neutralization Ability of Taiwanese Freeze-dried Neurotoxic Antivenom in Animal Models. PLoS Neglect. Trop. Dis. 2020, 14, e0008054. [Google Scholar] [CrossRef]
- Bin Haidar, H.; Almeida, J.R.; Williams, J.; Guo, B.; Bigot, A.; Senthilkumaran, S.; Vaiyapuri, S.; Patel, K. Differential Effects of the Venoms of Russell’s Viper and Indian Cobra on Human Myoblasts. Sci. Rep. 2024, 14, 3184. [Google Scholar] [CrossRef] [PubMed]
- Alkilani, A.Z.; McCrudden, M.T.C.; Donnelly, R.F. Transdermal Drug Delivery: Innovative Pharmaceutical Developments Based on Disruption of the Barrier Properties of the Stratum Corneum. Pharmaceutics 2015, 7, 438–470. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, G.T.T.; O’Brien, C.; Wouters, Y.; Seneci, L.; Gallissà-Calzado, A.; Campos-Pinto, I.; Ahmadi, S.; Laustsen, A.H.; Ljungars, A. High-throughput Proteomics and Functional Characterization of the 26 Medically Most Important Elapids and Vipers from Sub-Saharan Africa. Gigascience 2022, 11, giac121. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, J.M.; Lewin, M.R.; Williams, D.J.; Lomonte, B. Varespladib (LY315920) and Methyl Varespladib (LY333013) Abrogate or Delay Lethality Induced by Presynaptically Acting Neurotoxic Snake Venoms. Toxins 2020, 12, 131. [Google Scholar] [CrossRef]
- Lay, M.; Liang, Q.; Isbister, G.K.; Hodgson, W.C. In Vitro Efficacy of Antivenom and Varespladib in Neutralising Chinese Russell’s Viper (Daboia siamensis) Venom Toxicity. Toxins 2023, 15, 62. [Google Scholar] [CrossRef]
- Lian, Q.; Zhong, L.P.; Fu, K.P.; Ji, Y.X.; Zhang, X.Y.; Liu, C.J.; Huang, C.H. Hepatic Inhibitors Expression Profiling of Venom-Challenged Sinonatrix annularis and Antidotal Activities. Biomed. Pharmacother. 2022, 156, 113900. [Google Scholar] [CrossRef]
- Zhang, M.L.; Zhang, R.; Liu, J.; Wang, H.L.; Wang, Z.; Liu, J.; Shan, Y.; Yu, H.L. The Effects of 5,6,7,8,3′,4′-Hexamethoxyflavone on Apoptosis of Cultured Human Choriocarcinoma Trophoblast Cells. Molecules 2020, 25, 946. [Google Scholar] [CrossRef]
- Da Silva, D.P.; Ferreira, S.D.; Torres-Rego, M.; Furtado, A.A.; Yamashita, F.D.; Diniz, E.A.D.; Vieira, D.S.; Ururahy, M.A.G.; Da Silva, A.A.; Luna, K.P.D.; et al. Antiophidic Potential of Chlorogenic Acid and Rosmarinic Acid Against Bothrops leucurus Snake Venom. Biomed. Pharmacother. 2022, 148, 112766. [Google Scholar] [CrossRef]
- Li, M.; Fan, Z.J.; Gao, Q.X.; He, Y.; Xu, A.Y.; Gu, Z.F.; Wang, S.X.; Bai, H.P.; Liao, Y.H.; Zhang, R.L. Dual-Atom Catalyst Au@S-rGO for Rapid and Highly Sensitive Electrochemical Detection of Fentanyl in Serum. Adv. Sci. 2025, 12, 2500430. [Google Scholar] [CrossRef]
- Ho, C.H.; Chiang, L.C.; Mao, Y.C.; Lan, K.C.; Tsai, S.H.; Shih, Y.J.; Tzeng, Y.S.; Lin, C.S.; Lin, W.L.; Fang, W.H.; et al. Analysis of the Necrosis-Inducing Components of the Venom of and Assessment of the Neutralization Ability of Freeze-Dried Antivenom. Toxins 2021, 13, 619. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, J.; Liu, G.; Shi, X.; Huang, C. Combination of Rhamnetin and RXP03 Mitigates Venom-Induced Toxicity in Murine Models: Preclinical Insights into Dual-Target Antivenom Therapy. Toxins 2025, 17, 280. https://doi.org/10.3390/toxins17060280
Zhao J, Liu G, Shi X, Huang C. Combination of Rhamnetin and RXP03 Mitigates Venom-Induced Toxicity in Murine Models: Preclinical Insights into Dual-Target Antivenom Therapy. Toxins. 2025; 17(6):280. https://doi.org/10.3390/toxins17060280
Chicago/Turabian StyleZhao, Jianqi, Guangyao Liu, Xiao Shi, and Chunhong Huang. 2025. "Combination of Rhamnetin and RXP03 Mitigates Venom-Induced Toxicity in Murine Models: Preclinical Insights into Dual-Target Antivenom Therapy" Toxins 17, no. 6: 280. https://doi.org/10.3390/toxins17060280
APA StyleZhao, J., Liu, G., Shi, X., & Huang, C. (2025). Combination of Rhamnetin and RXP03 Mitigates Venom-Induced Toxicity in Murine Models: Preclinical Insights into Dual-Target Antivenom Therapy. Toxins, 17(6), 280. https://doi.org/10.3390/toxins17060280