Micrurus nigrocinctus in Colombia: Integrating Venomics Research, Citizen Science, and Community Empowerment

 , , ,
, , ,  , and
, and
Abstract

1. Introduction
2. Results
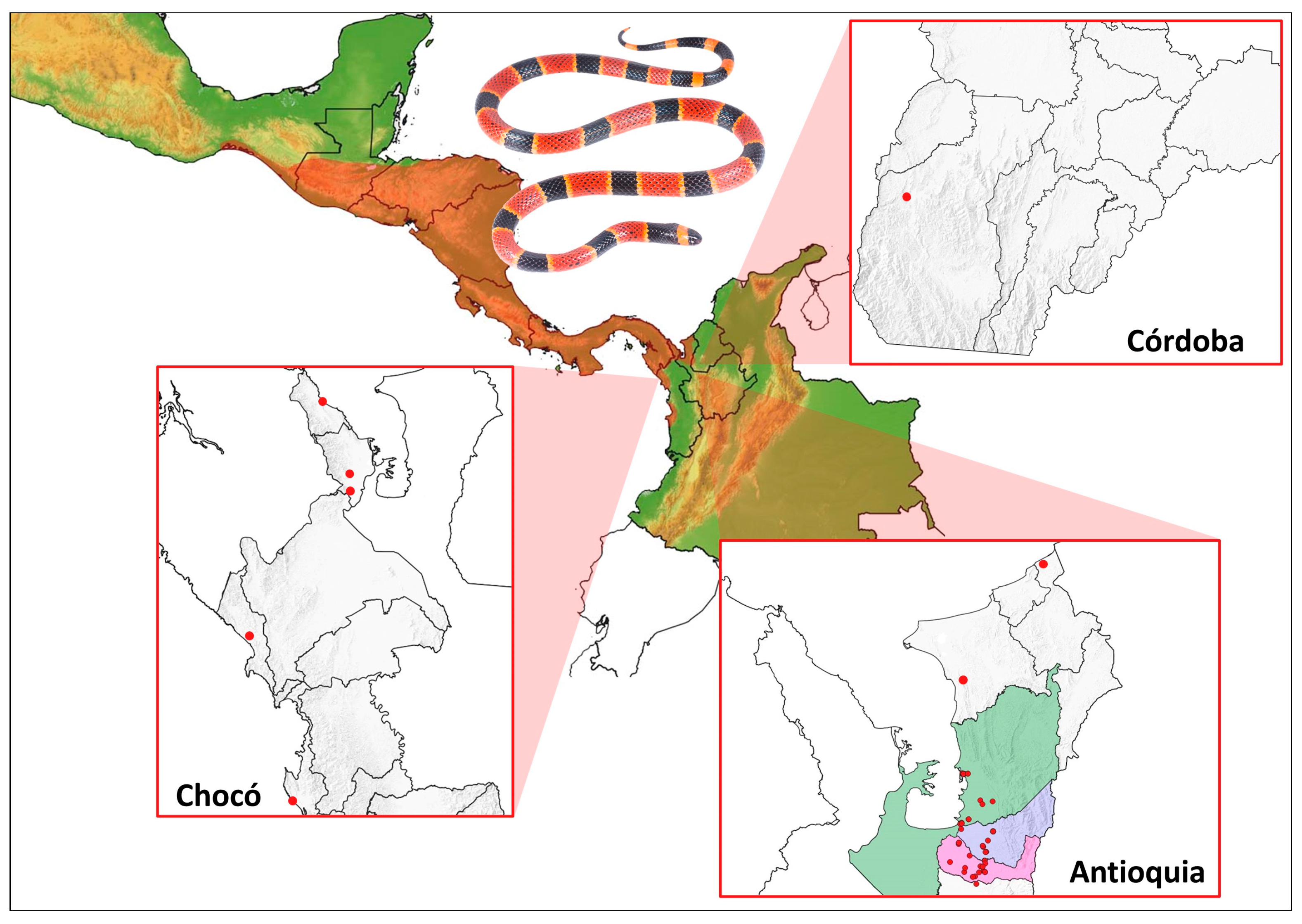
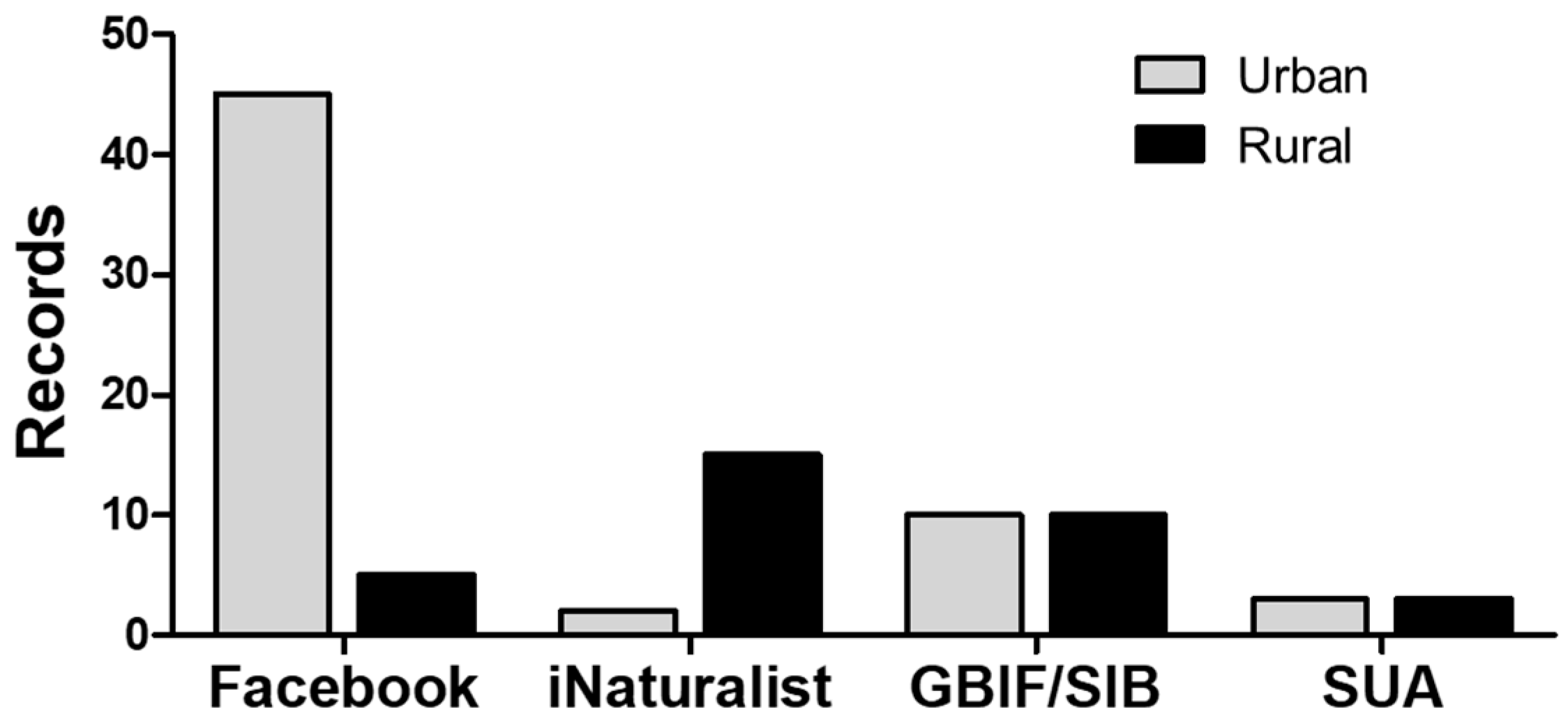
2.1. Distribution of M. nigrocinctus in Colombia Based on ‘Citizen Science’ and Scientific Database Reports
2.2. Molecular and Phylogenetic Analysis of M. nigrocinctus from Colombia
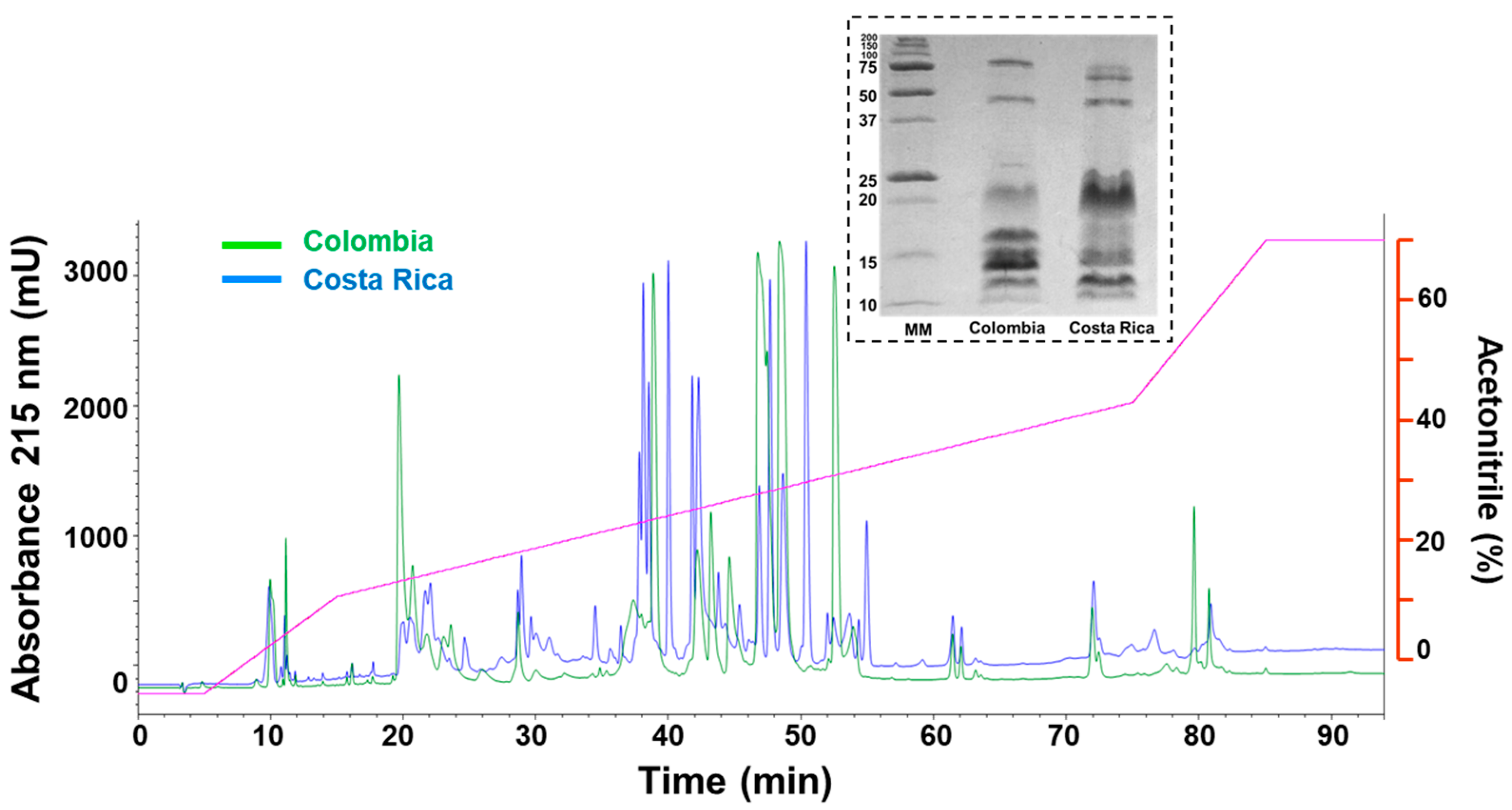
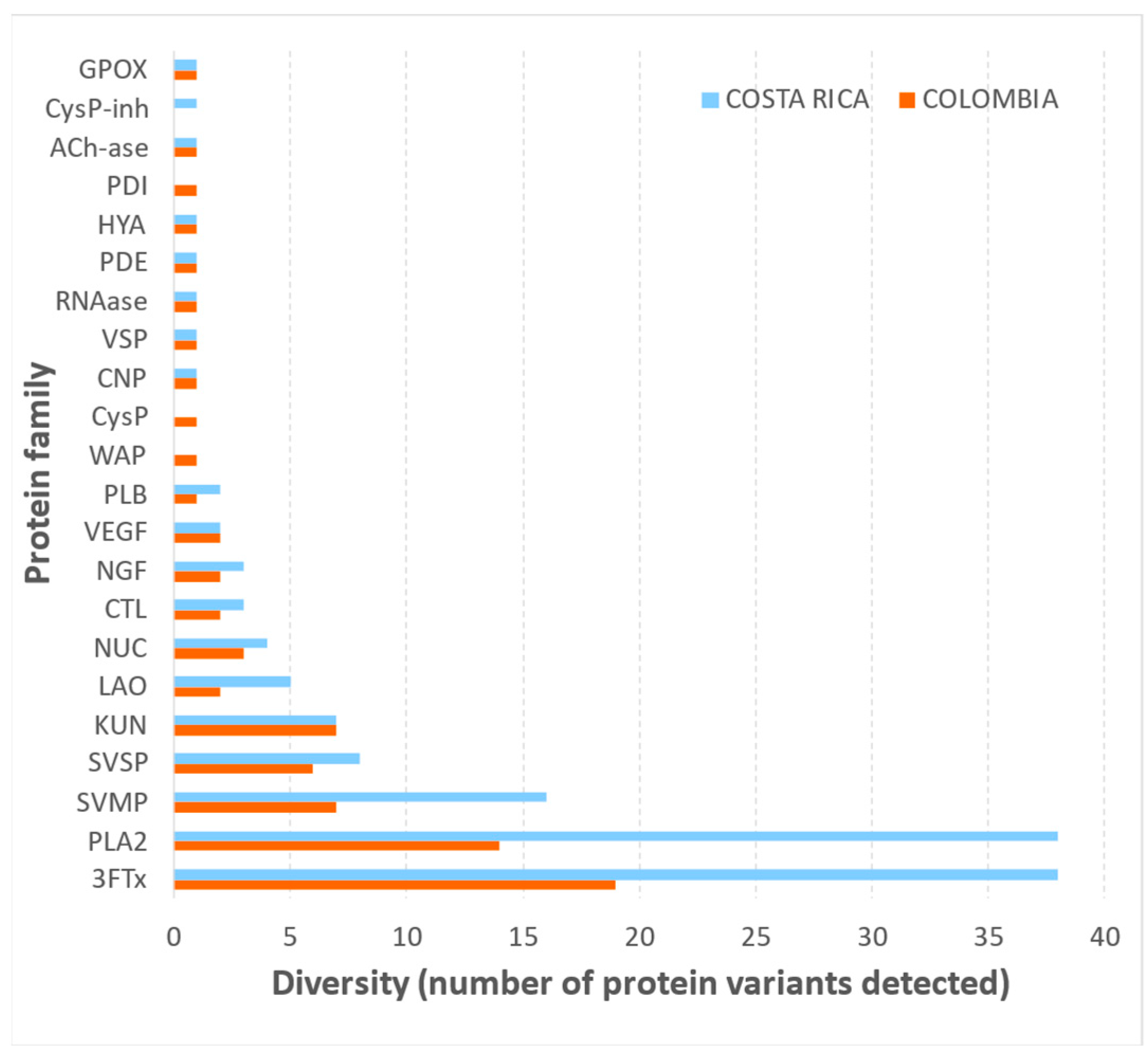
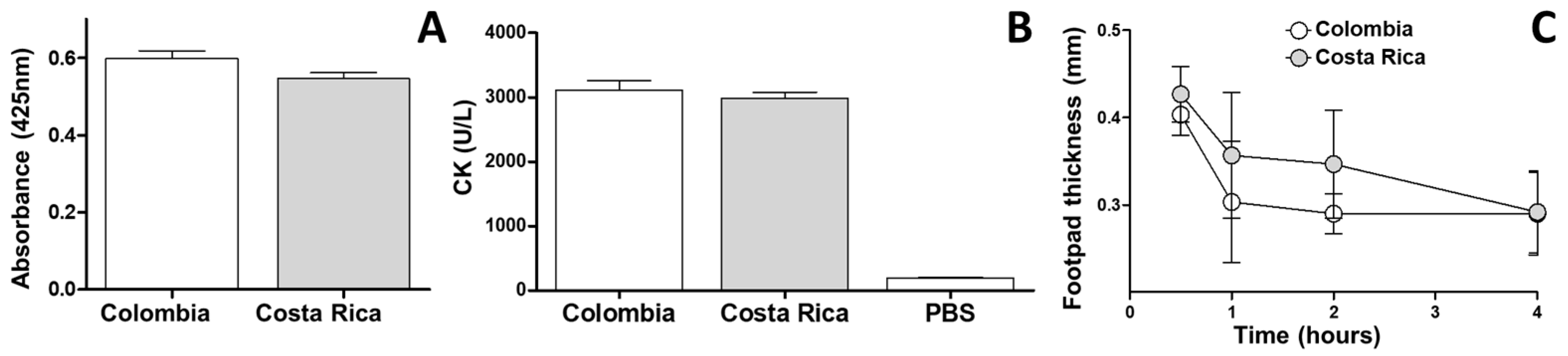
2.3. Comparative Biochemical and Biological Characterization of M. nigrocinctus Venom from Colombia and Costa Rica
2.4. Immunorecognition and Neutralization by Commercial Coral Antivenoms
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Distribution of M. nigrocinctus in Colombia Based on ‘Citizen Science’ and Scientific Database Reports
5.2. Molecular and Phylogenetic Analysis of M. nigrocinctus from Colombia
5.3. Characterization of M. nigrocinctus Venom from Colombia and Its Immunorecognition by Commercial Coral Snake Antivenoms
5.3.1. Venoms and Antivenoms
5.3.2. Venom Chromatographic and Electrophoretic Profiles
5.3.3. Shotgun Proteomic Profiling
5.3.4. Biological Activities of M. nigrocinctus Venom from Colombia
5.3.5. Venom Immunorecognition and Neutralization by Commercial Anticoral Antivenoms
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- GBD 2019 Snakebite Envenomation Collaborators. Global Mortality of Snakebite Envenoming between 1990 and 2019. Nat. Commun. 2022, 13, 6160. [Google Scholar] [CrossRef] [PubMed]
- Warrell, D.A. Snake bite. Lancet 2010, 375, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Chippaux, J.P. Snakebite envenomation turns again into a neglected tropical disease! J. Venom. Anim. Toxins Incl. Trop. Dis. 2017, 8, 23–38. [Google Scholar] [CrossRef] [PubMed]
- da Silva, W.R.G.B.; de Siqueira Santos, L.; Lira, D.; de Oliveira Luna, K.P.; Fook, S.M.L.; Alves, R.R.N. Who Are the Most Affected by Bothrops Snakebite Envenoming in Brazil? A Clinical-Epidemiological Profile Study among the Regions of the Country. PLoS Negl. Trop. Dis. 2023, 17, e0011634. [Google Scholar] [CrossRef] [PubMed]
- Tianyi, F.L.; Oluoch, G.O.; Otundo, D.; Ofwete, R.; Ngari, C.; Trelfa, A.; Ahmed, S.; Wang, D.; Smith, M.; Meta, V.; et al. Snakebite Prevalence and Risk Factors in a Nomadic Population in Samburu County, Kenya: A Community-Based Survey. PLoS Negl. Trop. Dis. 2024, 18, e0011677. [Google Scholar] [CrossRef]
- Gupta, P.P.; Bhandari, R.; Bhandari, R.; Sarraf, D.P.; Malla, G. Presentation, Treatment Profiles, and Outcome of Snake Bite Patients Presented in Emergency Department at a Tertiary Hospital in Eastern Nepal. J. Fam. Med. Prim. Care 2024, 13, 1037–1041. [Google Scholar] [CrossRef]
- Gutiérrez, J.M.; Calvete, J.J.; Habib, A.G.; Harrison, R.A.; Williams, D.J.; Warrell, D.A. Snakebite Envenoming. Nat. Rev. Dis. Primers 2017, 3, 17063. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Snakebite Envenoming: A Strategy for Prevention and Control; World Health Organization: Geneva, Switzerland, 2019; Available online: https://www.who.int/publications/i/item/9789241515641 (accessed on 12 February 2025).
- Gutiérrez, J.M.; Borri, J.; Giles-Vernick, T.; Duda, R.; Habib, A.; Malhotra, A.; Martín, G.; Pintor, A.F.V.; Potet, J.; Scott, T.; et al. Understanding and Tackling Snakebite Envenoming with Transdisciplinary Research. PLoS Negl. Trop. Dis. 2022, 16, e0010897. [Google Scholar] [CrossRef]
- Gutiérrez, J.M.; Bolon, I.; Borri, J.; de Castañeda, R.R. Broadening the Research Landscape in the Field of Snakebite Envenoming: Towards a Holistic Perspective. Toxicon 2023, 233, 107279. [Google Scholar] [CrossRef]
- Minghui, R.; Malecela, M.N.; Cooke, E.; Abela-Ridder, B. WHO’s Snakebite Envenoming Strategy for Prevention and Control. Lancet Glob. Health 2019, 7, e837–e838. [Google Scholar] [CrossRef]
- Malhotra, A.; Wüster, W.; Owens, J.B.; Hodges, C.W.; Jesudasan, A.; Ch, G.; Kartik, A.; Christopher, P.; Louies, J.; Naik, H.; et al. Promoting Co-existence between Humans and Venomous Snakes through Increasing the Herpetological Knowledge Base. Toxicon X 2021, 12, 100081. [Google Scholar] [CrossRef] [PubMed]
- Munshi, H.; Gajbhiye, R.K. National Action Plan for the prevention and control of snakebite envenoming in India: Strategies and challenges. Trans. R. Soc. Trop. Med. Hyg. 2025, 119, 560–562. [Google Scholar] [CrossRef]
- The Reptile Database. Available online: http://www.reptile-database.org (accessed on 10 March 2025).
- Gutiérrez, J.M. Envenenamientos por mordeduras de serpientes en América Latina y el Caribe: Una visión integral de carácter regional. Bol. Mal. Salud 2011, 51, 1–16. [Google Scholar]
- Bucaretchi, F.; Capitani, E.M.; Vieira, R.J.; Rodrigues, C.K.; Zannin, M.; Da Silva, N.J., Jr.; Casais-e-Silva, L.L.; Hyslop, S. Coral Snake Bites (Micrurus spp.) in Brazil: A Review of Literature Reports. Clin. Toxicol. 2016, 54, 222–234. [Google Scholar] [CrossRef] [PubMed]
- Bucaretchi, F.; Trindade da Silva, Y.; Mitsunaga, T.M.; De Capitani, E.M.; Jorge da Silva, N., Jr.; Hyslop, S. Snakebites by Venomous Snakes in Brazilian Serpentaria and Zoos over a 10-Year Period (2012–2021). Toxicon 2024, 249, 108082. [Google Scholar] [CrossRef]
- Campbell, J.A.; Lamar, W.W. The Venomous Reptiles of the Western Hemisphere; Comstock Publishing Associates: Ithaca, NY, USA, 2004. [Google Scholar]
- Marques, O.A.V.; Sazima, I. The Natural History of New World Coralsnakes. In Advances in Coralsnake Biology: With an Emphasis in South America; Eagle Mountain Publishing: Eagle Mountain, UT, USA, 2021; pp. 275–314. [Google Scholar]
- Lynch, J.D.; Sierra, T.A.; Gómez, F.J.R. Programa Nacional para la Conservación de las Serpientes Presentes en Colombia. Ministerio de Ambiente y Desarrollo Sostenible: Bogota, Colombia, 2014. [Google Scholar]
- Estrada-Gómez, S.; Vargas-Muñoz, L.J.; Higuita-Gutiérrez, L.F. Epidemiology of Snake Bites Linked with the Antivenoms Production in Colombia 2008–2020: Produced Vials Do Not Meet the Needs. Drug Healthc. Patient Saf. 2022, 14, 171–184. [Google Scholar] [CrossRef]
- Chacón, D.; Rodríguez, S.; Arias, J.; Solano, G.; Bonilla, F.; Gómez, A. Maintaining Coral Snakes (Micrurus nigrocinctus, Serpentes: Elapidae) for Venom Production on an Alternative Fish-Based Diet. Toxicon 2012, 60, 249–253. [Google Scholar] [CrossRef]
- Neri-Castro, E.; Zarzosa, V.; Benard-Valle, M.; Rodríguez-Solís, A.M.; Hernández-Orihuela, L.; Ortiz-Medina, J.A.; Alagón, A. Quantifying Venom Production: A Study on Micrurus Snakes in Mexico. Toxicon 2024, 240, 107658. [Google Scholar] [CrossRef]
- León, G.; Stiles, B.; Alape, A.; Rojas, G.; Gutiérrez, J.M. Comparative Study on the Ability of IgG and F(ab’)2 Antivenoms to Neutralize Lethal and Myotoxic Effects Induced by Micrurus nigrocinctus (Coral Snake) Venom. Am. J. Trop. Med. Hyg. 1999, 61, 266–271. [Google Scholar] [CrossRef]
- Gutiérrez, J.M. Snakebite Envenomation in Central America. Epidemiology, Pathophysiology, and Treatment. In Handbook of Venoms and Toxins of Reptiles, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2021; pp. 543–558. [Google Scholar]
- Goularte, F.C.; da Cruz-Höfling, M.A.; Corrado, A.P.; Rodrigues-Simioni, L. Electrophysiological and Ultrastructural Analysis of the Neuromuscular Blockade and Myotoxicity Induced by the Micrurus nigrocinctus Snake Venom. Acta Physiol. Pharmacol. Ther. Latinoam. 1999, 49, 290–296. [Google Scholar]
- Rosso, J.P.; Vargas-Rosso, O.; Gutiérrez, J.M.; Rochat, H.; Bougis, P.E. Characterization of Alpha-Neurotoxin and Phospholipase A2 Activities from Micrurus Venoms. Determination of the Amino Acid Sequence and Receptor-Binding Ability of the Major Alpha-Neurotoxin from Micrurus nigrocinctus nigrocinctus. Eur. J. Biochem. 1996, 238, 231–239. [Google Scholar] [CrossRef]
- Gutiérrez, J.M.; Chaves, F.; Lomonte, B. Pharmacological activities of crude venom from the coral snake Micrurus nigrocinctus (Elapidae) from Costa Rica. Toxicon 1986, 24, 841–850. [Google Scholar]
- Fernández, J.; Alape-Girón, A.; Angulo, Y.; Sanz, L.; Gutiérrez, J.M.; Calvete, J.J.; Lomonte, B. Venomic and Antivenomic Analyses of the Central American Coral Snake, Micrurus nigrocinctus (Elapidae). J. Proteome Res. 2011, 10, 1816–1827. [Google Scholar] [CrossRef] [PubMed]
- Fernández, J.; Chaves, W.; Vargas-Diaz, D.; Petras, D.; Lomonte, B. Top-Down Proteomics of Venoms from Five Micrurus Species from Costa Rica: Comparative Composition of Phospholipase A2-Rich vs Three-Finger Toxin-Rich Phenotypes. Toxicon 2024, 252, 108187. [Google Scholar] [CrossRef] [PubMed]
- Jowers, M.J.; Smart, U.; Sánchez-Ramírez, S.; Murphy, J.C.; Gómez, A.; Bosque, R.J.; Sarker, G.C.; Noonan, B.P.; Faria, J.F.; Harris, D.J.; et al. Unveiling Underestimated Species Diversity within the Central American Coralsnake, a Medically Important Complex of Venomous Taxa. Sci. Rep. 2023, 13, 11674. [Google Scholar] [CrossRef]
- Dorcas, M.E.; Willson, J.D. Innovative Methods for Studies of Snake Ecology and Conservation. Snakes Ecol. Conserv. 2009, 1, 5–37. [Google Scholar]
- Reynolds, R.P.; McDiarmid, R.W. Voucher Specimens. Reptile Biodivers. Stand. Methods Inventory Monit. 2012, 89–94. [Google Scholar]
- Pyke, G.H.; Ehrlich, P.R. Biological Collections and Ecological/Environmental Research: A Review, Some Observations, and a Look to the Future. Biol. Rev. Camb. Philos. Soc. 2010, 85, 247–266. [Google Scholar] [CrossRef]
- Ruane, S. New Data from Old Specimens. Ichthyol. Herpetol. 2021, 109, 392–396. [Google Scholar] [CrossRef]
- Durso, A.M.; Ruiz de Castañeda, R.; Montalcini, C.; Mondardini, M.R.; Fernandez-Marques, J.L.; Grey, F.; Müller, M.M.; Uetz, P.; Marshall, B.M.; Gray, R.J.; et al. Citizen Science and Online Data: Opportunities and Challenges for Snake Ecology and Action Against Snakebite. Toxicon X 2021, 9-10, 100071. [Google Scholar] [CrossRef]
- Martínez, R.; Machado, K. Presencia, Distribución y Ataques de los Ofidios Venenosos del Chocó Biogeográfico: Una Ventana a la Bioprospección de los Recursos Genéticos de la Región. Rev. Bioetnia 2009, 6, 40–52. [Google Scholar] [CrossRef]
- Chacón, J.; Humanez-López, E.; De Ávila, O.B. Vertebrados Asociados con Sistemas de Ganadería Extensiva en el Departamento de Córdoba, Colombia. Rev. Biodivers. Neotrop. 2012, 2, 45–52. [Google Scholar] [CrossRef]
- Ruíz, O. Ofidios del Corregimiento de San Rafael de Pirú, Valencia, Córdoba-Colombia. Rev. Colomb. Cienc. Anim. 2014, 6, 3–13. [Google Scholar] [CrossRef]
- Rojas Murcia, L.E.; Carvajal Cogollo, J.E.; Cabrejo Bello, J.A. Reptiles del Bosque Seco Estacional en el Caribe Colombiano: Distribución de los Hábitats y del Recurso Alimentario. Acta Biol. Colomb. 2016, 21, 365–377. [Google Scholar] [CrossRef]
- Savage, J.W. The Amphibians and Reptiles of Costa Rica; The University of Chicago Press: Chicago, IL, USA, 2002; 934p. [Google Scholar]
- Urdaneta, A.H.; Bolaños, F.; Gutiérrez, J.M. Feeding Behavior and Venom Toxicity of Coral Snake Micrurus nigrocinctus (Serpentes: Elapidae) on Its Natural Prey in Captivity. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2004, 138, 485–492. [Google Scholar] [CrossRef]
- Bisneto, P.F.; Alcântara, J.A.; Mendonça da Silva, I.; de Almeida Gonçalves Sachett, J.; Bernarde, P.S.; Monteiro, W.M.; Kaefer, I.L. Coral Snake Bites in Brazilian Amazonia: Perpetrating Species, Epidemiology, and Clinical Aspects. Toxicon 2020, 175, 7–18. [Google Scholar] [CrossRef]
- Marques, O.A.V.; Pereira, D.N.; Barbo, F.E.; Germano, V.J.; Sawaya, R.J. Os Répteis do Município de São Paulo: Diversidade e Ecologia da Fauna Pretérita e Atual. Biota Neotrop. 2009, 9, 139–150. [Google Scholar] [CrossRef]
- Silva, N.J., Jr.; Feitosa, D.T.; Pires, M.G.; Prudente, A.L.C. Coralsnake Diversity in Brazil. In Advances in Coralsnake Biology: With an Emphasis on South America; Prudente, A.L.C., Ed.; Eagle Mountain Publishing: Eagle Mountain, UT, USA, 2021; pp. 141–252. [Google Scholar]
- Zaher, H.; Grazziotin, F.G.; Prudente, A.L.C.; Quadros, A.B.A.; Trevine, V.C.; Silva, N.J., Jr. Origin and Evolution of Elapids and New World Coralsnakes. In Advances in Coralsnake Biology: With an Emphasis on South America; Silva, N.J., Jr., Porras, L.W., Aird, S.D., Prudente, A.L.C., Eds.; Eagle Mountain Publishing: Eagle Mountain, UT, USA, 2021. [Google Scholar]
- Streicher, J.W.; McEntee, J.P.; Drzich, L.C.; Card, D.C.; Schield, D.R.; Smart, U.; Parkinson, L.P.; Jezkova, T.; Smith, E.N.; Castoe, T. A Genetic Surfing, Not Allopatric Divergence, Explains Spatial Sorting of Mitochondrial Haplotypes in Venomous Coralsnakes. Evolution 2016, 70, 1435–1449. [Google Scholar] [CrossRef] [PubMed]
- Slowinski, J.B.; Keogh, J.S. Phylogenetic Relationships of Elapid Snakes Based on Cytochrome B mtDNA Sequences. Mol. Phylogenet. Evol. 2000, 15, 157–164. [Google Scholar] [CrossRef]
- Castoe, T.A.; Poole, A.W.; de Koning, A.P.; Jones, K.L.; Tomback, D.F.; Oyler-McCance, S.J.; Fike, J.A.; Lance, S.L.; Streicher, J.W.; Smith, E.N.; et al. Rapid Microsatellite Identification from Illumina Paired-End Genomic Sequencing in Two Birds and a Snake. PLoS ONE 2012, 7, e30953. [Google Scholar] [CrossRef]
- Nascimento, L.R.S.; Silva, N.J.J.; Feitosa, D.T.; Prudente, A.L.C. Taxonomy of the Micrurus spixii Species Complex (Serpentes, Elapidae). Zootaxa 2019, 4668, 424–436. [Google Scholar] [CrossRef]
- Reyes-Velasco, J.; Adams, R.H.; Boissinot, S.; Parkinson, C.L.; Campbell, J.A.; Castoe, T.A.; Smith, E.N. Genome-Wide SNPs Clarify Lineage Diversity Confused by Coloration in Coralsnakes of the Micrurus diastema Species Complex (Serpentes: Elapidae). Mol. Phylogenet. Evol. 2020, 147, 106770. [Google Scholar] [CrossRef] [PubMed]
- Jowers, M.J.; Garcia Mudarra, J.L.; Charles, S.P.; Murphy, J.C. Phylogeography of West Indies Coral Snakes (Micrurus): Island Colonization and Banding Patterns. Zool. Scr. 2019, 48, 263–276. [Google Scholar] [CrossRef]
- Gutiérrez, J.M.; Lomonte, B.; Portilla, E.; Cerdas, L.; Rojas, E. Local Effects Induced by Coral Snake Venoms: Evidence of Myonecrosis After Experimental Inoculations of Venoms from Five Species. Toxicon 1983, 21, 777–783. [Google Scholar] [CrossRef]
- Arroyo, O.; Rosso, J.P.; Vargas, O.; Gutiérrez, J.M.; Cerdas, L. Skeletal Muscle Necrosis Induced by a Phospholipase A2 Isolated from the Venom of the Coral Snake Micrurus nigrocinctus nigrocinctus. Comp. Biochem. Physiol. B 1987, 87, 949–952. [Google Scholar] [CrossRef]
- Fernández, J.; Vargas-Vargas, N.; Pla, D.; Sasa, M.; Rey-Suárez, P.; Sanz, L.; Gutiérrez, J.M.; Calvete, J.J.; Lomonte, B. Snake Venomics of Micrurus alleni and Micrurus mosquitensis from the Caribbean Region of Costa Rica Reveals Two Divergent Compositional Patterns in New World Elapids. Toxicon 2015, 107 (Pt B), 217–233. [Google Scholar] [CrossRef]
- Lippa, E.; Török, F.; Gómez, A.; Corrales, G.; Chacón, D.; Sasa, M.; Gutiérrez, J.M.; Lomonte, B.; Fernández, J. First Look into the Venom of Roatan Island’s Critically Endangered Coral Snake Micrurus ruatanus: Proteomic Characterization, Toxicity, Immunorecognition, and Neutralization by an Antivenom. J. Proteom. 2019, 198, 177–185. [Google Scholar] [CrossRef]
- Castillo-Beltrán, M.C.; Hurtado-Gómez, J.P.; Corredor-Espinel, V.; Ruiz-Gómez, F.J. A Polyvalent Coral Snake Antivenom with Broad Neutralization Capacity. PLoS Negl. Trop. Dis. 2019, 13, e0007250. [Google Scholar] [CrossRef]
- Piedrahita, J.D.; Cardona-Ruda, A.; Pereañez, J.A.; Rey-Suárez, P. In-Depth Immunorecognition and Neutralization Analyses of Micrurus mipartitus and M. dumerilii Venoms and Toxins by a Commercial Antivenom. Biochimie 2024, 216, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Rey-Suárez, P.; Núñez, V.; Gutiérrez, J.M.; Lomonte, B. Proteomic and Biological Characterization of the Venom of the Redtail Coral Snake, Micrurus mipartitus (Elapidae), from Colombia and Costa Rica. J. Proteom. 2011, 75, 655–667. [Google Scholar] [CrossRef]
- Lomonte, B.; Rey-Suárez, P.; Fernández, J.; Sasa, M.; Pla, D.; Vargas, N.; Bénard-Valle, M.; Sanz, L.; Corrêa-Netto, C.; Núñez, V.; et al. Venoms of Micrurus Coral Snakes: Evolutionary Trends in Compositional Patterns Emerging from Proteomic Analyses. Toxicon 2016, 122, 7–25. [Google Scholar] [CrossRef]
- Mena, G.; Chaves-Araya, S.; Chacón, J.; Török, E.; Török, F.; Bonilla, F.; Sasa, M.; Gutiérrez, J.M.; Lomonte, B.; Fernández, J. Proteomic and Toxicological Analysis of the Venom of Micrurus yatesi and Its Neutralization by an Antivenom. Toxicon X 2022, 13, 100097. [Google Scholar] [CrossRef] [PubMed]
- Roze, J.A. Coral Snakes of the Americas: Biology, Identification, and Venoms; Krieger Publishing Company: Malabar, FL, USA, 1996. [Google Scholar]
- Sasaki, J.; Khalil, P.A.; Chegondi, M.; Raszynski, A.; Meyer, K.G.; Totapally, B.R. Coral Snake Bites and Envenomation in Children: A Case Series. Pediatr. Emerg. Care 2014, 30, 262–265. [Google Scholar] [CrossRef]
- Oliveira, I.S.; Pucca, M.B.; Cerni, F.A.; Vieira, S.; Sachett, J.; Seabra de Farias, A.; Lacerda, M.; Murta, F.; Baia-da-Silva, D.; Rocha, T.A.H.; et al. Snakebite Envenoming in Brazilian Children: Clinical Aspects, Management, and Outcomes. J. Trop. Pediatr. 2023, 69, fmad010. [Google Scholar] [CrossRef]
- da Silva, A.M.; Bernarde, P.S.; de Abreu, L.C. Accidents with Poisonous Animals in Brazil by Age and Sex. J. Hum. Growth Dev. 2015, 25, 54–62. [Google Scholar] [CrossRef]
- Velagapudi, M.E.; Navarro, J.J.; Hill, A.E.; Darracq, M.A.; Thornton, S.L. Epidemiology and Characteristics of Coral Snake Bites Reported to the National Poison Data System (2006–2022). J. Am. Coll. Emerg. Physicians Open 2024, 5, e13296. [Google Scholar] [CrossRef] [PubMed]
- Afroz, A.; Siddiquea, B.N.; Shetty, A.N.; Jackson, T.N.W.; Watt, A.D. Assessing Knowledge and Awareness Regarding Snakebite and Management of Snakebite Envenoming in Healthcare Workers and the General Population: A Systematic Review and Meta-Analysis. PLoS Negl. Trop. Dis. 2023, 17, e0011048. [Google Scholar] [CrossRef]
- Bala, A.A.; Jatau, A.I.; Yunusa, I.; Mohammed, M.; Mohammed, A.H.; Isa, A.M.; Wada, A.S.; Gulma, K.A.; Bello, I.; Malami, S.; et al. Knowledge Assessment of Anti-Snake Venom among Healthcare Practitioners in Northern Nigeria. Ther. Adv. Infect. Dis. 2021, 8, 20499361211039379. [Google Scholar] [CrossRef]
- Strauch, M.A.; Souza, G.J.; Pereira, J.N.; Ramos, T.D.S.; Cesar, M.O.; Tomaz, M.A.; Monteiro-Machado, M.; Patrão-Neto, F.C.; Melo, P.A. True or False Coral Snake: Is It Worth the Risk? A Micrurus corallinus Case Report. J. Venom. Anim. Toxins Incl. Trop. Dis. 2018, 24, 10. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An Integrated and Extendable Desktop Software Platform for the Organization and Analysis of Sequence Data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Nicholas, K.B. Genedoc: A Tool for Editing and Annotating Multiple Sequence Alignments. 1997. Available online: http://www.psc.edu/biomed/genedoc (accessed on 1 April 2024).
- Zhang, D.; Gao, F.; Jakovlić, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An Integrated and Scalable Desktop Platform for Streamlined Molecular Sequence Data Management and Evolutionary Phylogenetics Studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef]
- Ranwez, V.; Douzery, E.J.P.; Cambon, C.; Chantret, N.; Delsuc, F. MACSE v2: Toolkit for the Alignment of Coding Sequences Accounting for Frameshifts and Stop Codons. Mol. Biol. Evol. 2018, 35, 2582–2584. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior Summarization in Bayesian Phylogenetics Using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef]
- Lomonte, B.; Calvete, J.J. Strategies in ‘snake venomics’ aiming at an integrative view of compositional, functional, and immunological characteristics of venoms. J. Venom. Anim. Toxins Incl. Trop. Dis. 2017, 23, 26. [Google Scholar] [CrossRef] [PubMed]
- Mora-Obando, D.; Fernández, J.; Montecucco, C.; Gutiérrez, J.M.; Lomonte, B. Synergism between basic Asp49 and Lys49 phospholipase A2 myotoxins of viperid snake venom in vitro and in vivo. PLoS ONE 2014, 9, e109846. [Google Scholar] [CrossRef]
- Lomonte, B.; Tarkowski, A.; Hanson, L.A. Host response to Bothrops asper snake venom. Analysis of edema formation, inflammatory cells, and cytokine release in a mouse model. Inflammation 1993, 17, 93–105. [Google Scholar] [CrossRef]
- Gutiérrez, J.M.; Lomonte, B.; Cerdas, L. Isolation and partial characterization of a myotoxin from the venom of the snake Bothrops nummifer. Toxicon 1986, 24, 885–894. [Google Scholar] [CrossRef]
- Trevors, J.T. A BASIC program for estimating LD50 values using the IBM-PC. Bull. Environ. Contam. Toxicol. 1986, 37, 18–26. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Venom/Antivenom | Dead/Inject 1 |
|---|---|---|
| M. nigrocinctus venom | 0 | 4/4 |
| M. nigrocinctus venom + anticoral-ICP | 0.2 mg/mL | 0/4 |
| M. nigrocinctus venom + anticoral-INS | 0.2 mg/mL | 1/4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rey-Suárez, P.; Rojo, L.P.; Gómez-Robles, J.; Parra-Moreno, S.; Pachon-Camelo, E.; Fuentes-Florez, Y.; Lomonte, B.; Fernández, J.; Sasa, M.; Núñez, V.; et al. Micrurus nigrocinctus in Colombia: Integrating Venomics Research, Citizen Science, and Community Empowerment. Toxins 2025, 17, 268. https://doi.org/10.3390/toxins17060268
Rey-Suárez P, Rojo LP, Gómez-Robles J, Parra-Moreno S, Pachon-Camelo E, Fuentes-Florez Y, Lomonte B, Fernández J, Sasa M, Núñez V, et al. Micrurus nigrocinctus in Colombia: Integrating Venomics Research, Citizen Science, and Community Empowerment. Toxins. 2025; 17(6):268. https://doi.org/10.3390/toxins17060268
Chicago/Turabian StyleRey-Suárez, Paola, Lina Preciado Rojo, Jeisson Gómez-Robles, Sanin Parra-Moreno, Erica Pachon-Camelo, Yirlys Fuentes-Florez, Bruno Lomonte, Julián Fernández, Mahmood Sasa, Vitelbina Núñez, and et al. 2025. "Micrurus nigrocinctus in Colombia: Integrating Venomics Research, Citizen Science, and Community Empowerment" Toxins 17, no. 6: 268. https://doi.org/10.3390/toxins17060268
APA StyleRey-Suárez, P., Rojo, L. P., Gómez-Robles, J., Parra-Moreno, S., Pachon-Camelo, E., Fuentes-Florez, Y., Lomonte, B., Fernández, J., Sasa, M., Núñez, V., & Saldarriaga-Cordoba, M. (2025). Micrurus nigrocinctus in Colombia: Integrating Venomics Research, Citizen Science, and Community Empowerment. Toxins, 17(6), 268. https://doi.org/10.3390/toxins17060268