Computational Modeling of Low-Abundance Proteins in Venom Gland Transcriptomes: Bothrops asper and Bothrops jararaca


Abstract
1. Introduction
2. Results and Discussion
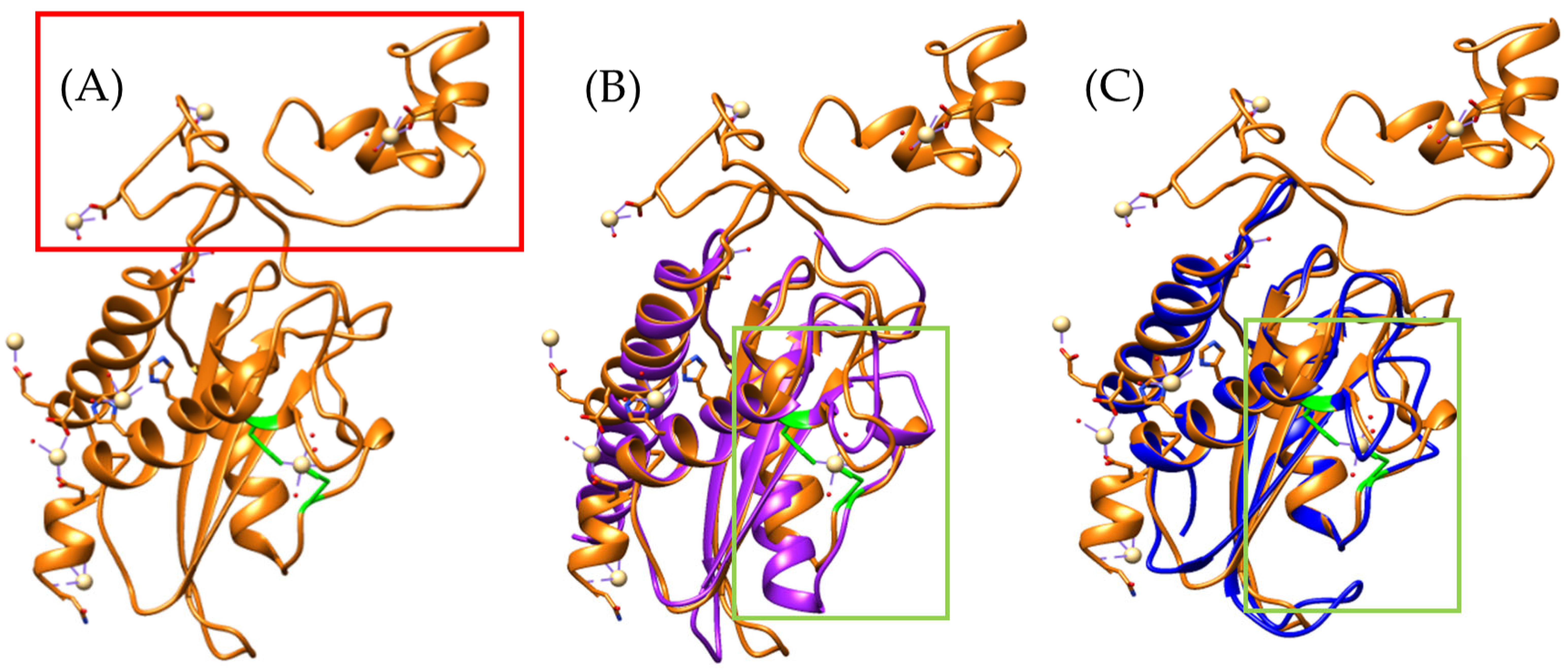
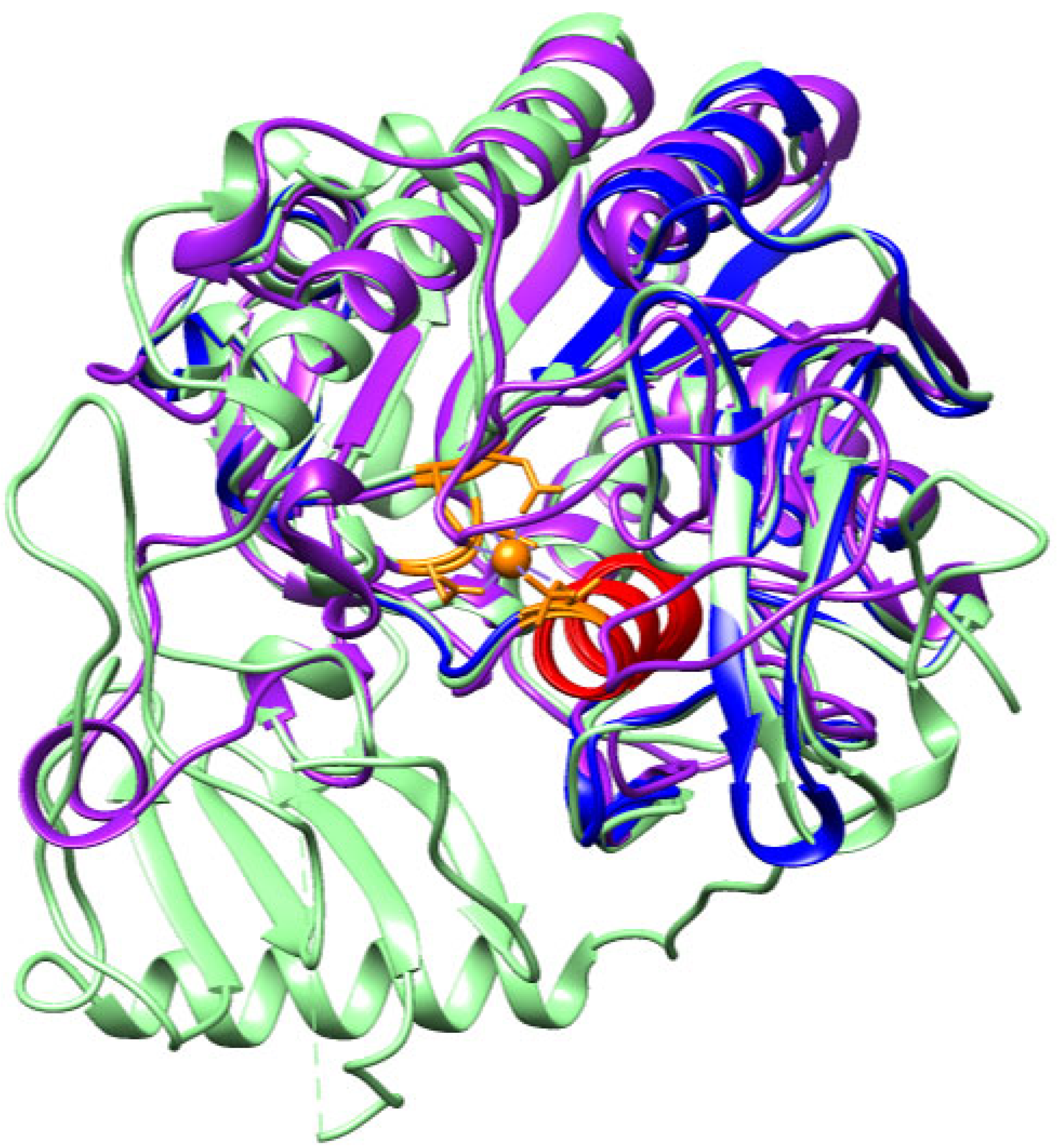
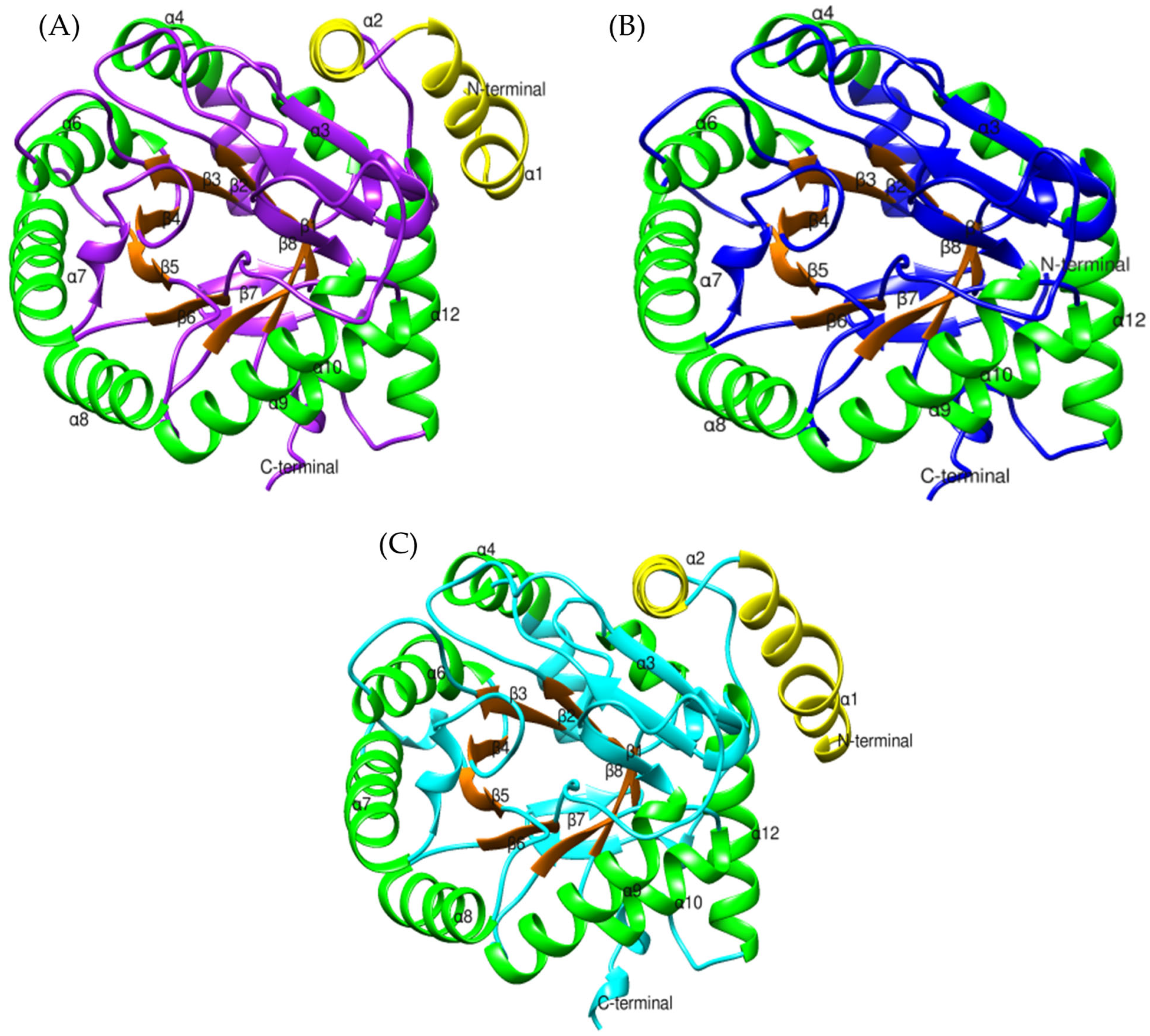
2.1. CRISP
2.2. Von Willebrand Factor Type D
2.3. Arylsulfatase
2.4. Botrocetin
2.5. Dihydroorotate Dehydrogenase
2.6. Basparin
2.7. Model Quality Analysis
3. Conclusions
4. Materials and Methods
4.1. Data Collection
4.2. Identification of Domains and Active Sites
4.3. Protein Modeling
4.4. Model Validation
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Post, Y.; Puschhof, J.; Beumer, J.; Kerkkamp, H.M.; de Bakker, M.A.; Slagboom, J.; de Barbanson, B.; Wevers, N.R.; Spijkers, X.M.; Olivier, T.; et al. Snake Venom Gland Organoids. Cell 2020, 180, 233-247.E21. [Google Scholar] [CrossRef] [PubMed]
- Jackson, T.N.; Young, B.; Underwood, G.; McCarthy, C.J.; Kochva, E.; Vidal, N.; van der Weerd, L.; Nabuurs, R.; Dobson, J.; Whitehead, D.; et al. Endless forms most beautiful: The evolution of ophidian oral glands, including the venom system, and the use of appropriate terminology for homologous structures. Zoomorphology 2017, 136, 107–130. [Google Scholar] [CrossRef]
- Casewell, N.R.; Jackson, T.N.; Laustsen, A.H.; Sunagar, K. Causes and Consequences of Snake Venom Variation. Trends Pharmacol. Sci. 2020, 41, 570–581. [Google Scholar] [CrossRef]
- Meier, J.; Stocker, K. Effects of Snake Venoms on Hemostasis. Crit. Rev. Toxicol. 1991, 21, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Jackson, K. The evolution of venom-delivery systems in snakes. Zool. J. Linn. Soc. 2003, 137, 337–354. [Google Scholar] [CrossRef]
- Vonk, F.J.; Casewell, N.R.; Henkel, C.V.; Heimberg, A.M.; Jansen, H.J.; McCleary, R.J.R.; Kerkkamp, H.M.E.; Vos, R.A.; Guerreiro, I.; Calvete, J.J.; et al. The king cobra genome reveals dynamic gene evolution and adaptation in the snake venom system. Proc. Natl. Acad. Sci. USA 2013, 110, 20651–20656. [Google Scholar] [CrossRef]
- De Mattos Pereira, L.; Messias, E.A.; Sorroche, B.P.; das Neves Oliveira, A.; Arantes, L.M.R.B.; de Carvalho, A.C.; Tanaka-Azevedo, A.M.; Grego, K.F.; Carvalho, A.L.; Melendez, M.E. In-depth transcriptome reveals the potential biotechnological application of Bothrops jararaca venom gland. J. Venom. Anim. Toxins Incl. Trop. Dis. 2020, 26, e20190058. [Google Scholar] [CrossRef]
- Casewell, N.R.; Harrison, R.A.; Wüster, W.; Wagstaff, S.C. Comparative venom gland transcriptome surveys of the saw-scaled vipers (Viperidae: Echis) reveal substantial intra-family gene diversity and novel venom transcripts. BMC Genom. 2009, 10, 564. [Google Scholar] [CrossRef]
- Tan, C.H.; Tan, K.Y.; Fung, S.Y.; Tan, N.H. Venom-gland transcriptome and venom proteome of the Malaysian king cobra (Ophiophagus hannah). BMC Genom. 2015, 16, 687. [Google Scholar] [CrossRef]
- Amorim, F.G.; Morandi-Filho, R.; Fujimura, P.T.; Ueira-Vieira, C.; Sampaio, S.V. New findings from the first transcriptome of the Bothrops moojeni snake venom gland. Toxicon 2017, 140, 105–117. [Google Scholar] [CrossRef]
- Babenko, V.V.; Ziganshin, R.H.; Weise, C.; Dyachenko, I.; Shaykhutdinova, E.; Murashev, A.N.; Zhmak, M.; Starkov, V.; Hoang, A.N.; Tsetlin, V.; et al. Novel Bradykinin-Potentiating Peptides and Three-Finger Toxins from Viper Venom: Combined NGS Venom Gland Transcriptomics and Quantitative Venom Proteomics of the Azemiops feae Viper. Biomedicines 2020, 8, 249. [Google Scholar] [CrossRef] [PubMed]
- Modahl, C.M.; Brahma, R.K. Snake venom gland. In Handbook of Venoms and Toxins of Reptiles; Mackessy, S.P., Ed.; CRC Press: Boca Raton, FL, USA, 2021; Volume 2, pp. 43–57. [Google Scholar]
- Uribe-Arjona, A.; De Patiño, H.A.; Martínez-Cortés, V.; Correa-Ceballos, D.; Rodríguez, A.; Gómez-Leija, L.; Vega, N.; Gutiérrez, J.M.; Otero-Patiño, R. Toxicological, enzymatic, and immunochemical characterization of Bothrops asper (Serpentes: Viperidae) reference venom from Panama. Rev. Biol. Trop. 2020, 69. [Google Scholar] [CrossRef]
- Resiere, D.; Monteiro, W.; Houcke, S.; Pujo, J.M.; Mathien, C.; Mayence, C.; Neviere, R.; Hommel, D.; Sachett, J.d.A.G.; Mehdaoui, H.; et al. Bothrops Snakebite Envenomings in the Amazon Region. Curr. Trop. Med. Rep. 2020, 7, 48–60. [Google Scholar] [CrossRef]
- Mora-Obando, D.; Salazar-Valenzuela, D.; Pla, D.; Lomonte, B.; Guerrero-Vargas, J.A.; Ayerbe, S.; Gibbs, H.L.; Calvete, J.J. Venom variation in Bothrops asper lineages from North-Western South America. J. Proteom. 2020, 229, 103945. [Google Scholar] [CrossRef] [PubMed]
- Nicolau, C.A.; Carvalho, P.C.; Junqueira-De-Azevedo, I.L.; Teixeira-Ferreira, A.; Junqueira, M.; Perales, J.; Neves-Ferreira, A.G.C.; Valente, R.H. An in-depth snake venom proteopeptidome characterization: Benchmarking Bothrops jararaca. J. Proteom. 2017, 151, 214–231. [Google Scholar] [CrossRef]
- Brahma, R.K.; McCleary, R.J.; Kini, R.M.; Doley, R. Venom gland transcriptomics for identifying, cataloging, and characterizing venom proteins in snakes. Toxicon 2015, 93, 1–10. [Google Scholar] [CrossRef]
- Boldrini-França, J.; Cologna, C.T.; Pucca, M.B.; Bordon, K.d.C.F.; Amorim, F.G.; Anjolette, F.A.P.; Cordeiro, F.A.; Wiezel, G.A.; Cerni, F.A.; Pinheiro-Junior, E.L.; et al. Minor snake venom proteins: Structure, function and potential applications. Biochim. Biophys. Acta Gen. Subj. 2017, 1861, 824–838. [Google Scholar] [CrossRef]
- Espín-Angulo, J.; Vela, D. Exploring the Venom Gland Transcriptome of Bothrops asper and Bothrops jararaca: De Novo Assembly and Analysis of Novel Toxic Proteins. Toxins 2024, 16, 511. [Google Scholar] [CrossRef]
- Van Vaerenbergh, M.; Cardoen, D.; Formesyn, E.M.; Brunain, M.; Van Driessche, G.; Blank, S.; Spillner, E.; Verleyen, P.; Wenseleers, T.; Schoofs, L.; et al. Extending the honey bee venome with the antimicrobial peptide apidaecin and a protein resembling wasp antigen Insect Mol. Biol. 2013, 22, 199–210. [Google Scholar] [CrossRef]
- Bazon, M.L.; Silveira, L.H.; Simioni, P.U.; Brochetto-Braga, M.R. Current Advances in Immunological Studies on the Vespidae Venom Antigen 5: Therapeutic and Prophylaxis to Hypersensitivity Responses. Toxins 2018, 10, 305. [Google Scholar] [CrossRef]
- Blank, S.; Bazon, M.L.; Grosch, J.; Schmidt-Weber, C.B.; Brochetto-Braga, M.R.; Bilò, M.B.; Jakob, T. Antigen 5 Allergens of Hymenoptera Venoms and Their Role in Diagnosis and Therapy of Venom Allergy. Curr. Allergy Asthma Rep. 2020, 20, 58. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, G.M.; Roelants, K.; O’Bryan, M.K. The CAP Superfamily: Cysteine-Rich Secretory Proteins, Antigen 5, and Pathogenesis-Related 1 Proteins—Roles in Reproduction, Cancer, and Immune Defense. Endocr. Rev. 2008, 29, 865–897. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, D. Allergens in Hymenoptera venom XXV: The amino acid sequences of antigen 5 molecules and the structural basis of antigenic cross-reactivity. J. Allergy Clin. Immunol. 1993, 92, 707–716. [Google Scholar] [CrossRef] [PubMed]
- Bouzid, W.; Verdenaud, M.; Klopp, C.; Ducancel, F.; Noirot, C.; Vétillard, A. De Novo sequencing and transcriptome analysis for Tetramorium bicarinatum: A comprehensive venom gland transcriptome analysis from an ant species. BMC Genom. 2014, 15, 987. [Google Scholar] [CrossRef]
- Tadokoro, T.; Modahl, C.M.; Maenaka, K.; Aoki-Shioi, N. Cysteine-Rich Secretory Proteins (CRISPs) from Venomous Snakes: An Overview of the Functional Diversity in a Large and Underappreciated Superfamily. Toxins 2020, 12, 175. [Google Scholar] [CrossRef]
- Tasoulis, T.; Isbister, G.K. A Review and Database of Snake Venom Proteomes. Toxins 2017, 9, 290. [Google Scholar] [CrossRef]
- Bernardes, C.P.; Menaldo, D.L.; Zoccal, K.F.; Boldrini-França, J.; Peigneur, S.; Arantes, E.C.; Rosa, J.C.; Faccioli, L.H.; Tytgat, J.; Sampaio, S.V. First report on BaltCRP, a cysteine-rich secretory protein (CRISP) from Bothrops alternatus venom: Effects on potassium channels and inflammatory processes. Int. J. Biol. Macromol. 2019, 140, 556–567. [Google Scholar] [CrossRef]
- Calvete, J.J.; Borges, A.; Segura, Á.; Flores-Díaz, M.; Alape-Girón, A.; Gutiérrez, J.M.; Diez, N.; De Sousa, L.; Kiriakos, D.; Sánchez, E.; et al. Snake venomics and antivenomics of Bothrops colombiensis, a medically important pitviper of the Bothrops atrox-asper complex endemic to Venezuela: Contributing to its taxonomy and snakebite management. J. Proteom. 2009, 72, 227–240. [Google Scholar] [CrossRef]
- Gonçalves-Machado, L.; Pla, D.; Sanz, L.; Jorge, R.J.B.; Leitão-De-Araújo, M.; Alves, M.L.M.; Alvares, D.J.; De Miranda, J.; Nowatzki, J.; de Morais-Zani, K.; et al. Combined venomics, venom gland transcriptomics, bioactivities, and antivenomics of two Bothrops jararaca populations from geographic isolated regions within the Brazilian Atlantic rainforest. J. Proteom. 2016, 135, 73–89. [Google Scholar] [CrossRef]
- Guo, M.; Teng, M.; Niu, L.; Liu, Q.; Huang, Q.; Hao, Q. Crystal Structure of the Cysteine-rich Secretory Protein Stecrisp Reveals That the Cysteine-rich Domain Has a K+ Channel Inhibitor-like Fold. J. Biol. Chem. 2005, 280, 12405–12412. [Google Scholar] [CrossRef]
- Shikamoto, Y.; Suto, K.; Yamazaki, Y.; Morita, T.; Mizuno, H. Crystal Structure of a CRISP Family Ca2+-channel Blocker Derived from Snake Venom. J. Mol. Biol. 2005, 350, 735–743. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Shen, B.; Guo, M.; Lou, X.; Duan, Y.; Cheng, X.P.; Teng, M.; Niu, L.; Liu, Q.; Huang, Q.; et al. Blocking effect and crystal structure of natrin toxin, a cysteine-rich secretory protein from Naja atra venom that targets the BKCa channel. Biochemistry 2005, 44, 10145–10152. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Yamazaki, Y.; Brown, R.L.; Fujimoto, Z.; Morita, T.; Mizuno, H. Structures of pseudechetoxin and pseudecin, two snake-venom cysteine-rich secretory proteins that target cyclic nucleotide-gated ion channels: Implications for movement of the C-terminal cysteine-rich domain. Acta Crystallogr. Sect. D Struct. Biol. 2008, 64, 1034–1042. [Google Scholar] [CrossRef] [PubMed]
- Carugo, O.; Pongor, S. A normalized root-mean-spuare distance for comparing protein three-dimensional structures. Protein Sci. 2001, 10, 1470–1473. [Google Scholar] [CrossRef]
- Kufareva, I.; Abagyan, R. Methods of Protein Structure Comparison. Homol. Model. Methods Protoc. 2011, 857, 231–257. [Google Scholar]
- Matsunaga, Y.; Yamazaki, Y.; Hyodo, F.; Sugiyama, Y.; Nozaki, M.; Morita, T. Structural Divergence of Cysteine-Rich Secretory Proteins in Snake Venoms†. J. Biochem. 2008, 145, 365–375. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Wang, Q.-L.; Meng, X.; Shu, Y.; Jiang, T.; Wagenknecht, T.; Yin, C.-C.; Sui, S.-F.; Liu, Z. Structural and Functional Characterization of Ryanodine Receptor-Natrin Toxin Interaction. Biophys. J. 2008, 95, 4289–4299. [Google Scholar] [CrossRef]
- Babin, P.J.; Bogerd, J.; Kooiman, F.P.; Van Marrewijk, W.J.A.; Van der Horst, D.J. Apolipophorin II/I, Apolipoprotein B, Vitellogenin, and Microsomal Triglyceride Transfer Protein Genes Are Derived from a Common Ancestor. J. Mol. Evol. 1999, 49, 150–160. [Google Scholar] [CrossRef]
- Van der Horst, D.J.; Rodenburg, K.W. Locust flight activity as a model for hormonal regulation of lipid mobilization and transport. J. Insect Physiol. 2010, 56, 844–853. [Google Scholar] [CrossRef]
- Kutty, R.K.; Kutty, G.; Kambadur, R.; Duncan, T.; Koonin, E.V.; Rodriguez, I.R.; Odenwald, W.F.; Wiggert, B. Molecular Characterization and Developmental Expression of a Retinoid- and Fatty Acid-binding Glycoprotein from Drosophila: A putative lipophorin. J. Biol. Chem. 1996, 271, 20641–20649. [Google Scholar] [CrossRef]
- Bogerd, J.; Babin, P.J.; Kooiman, F.P.; Ballagny, C.; Van Marrewijk, W.J.; Van Der Horst, D.J. Molecular characterization and gene expression in the eye of the apolipophorin II/I precursor fromLocusta migratoria. J. Comp. Neurol. 2000, 427, 546–558. [Google Scholar] [CrossRef] [PubMed]
- Avarre, J.C.; Lubzens, E.; Babin, P.J. Apolipocrustacein, formerly vitellogenin, is the major egg yolk precursor protein in decapod crustaceans and is homologous to insect apolipophorin II/I and vertebrate apolipoprotein B. BMC Evol. Biol. 2007, 7, 3. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Leksa, N.C.; Chhabra, E.S.; Arndt, J.W.; Lu, Q.; Knockenhauer, K.E.; Peters, R.T.; Springer, T.A. The von Willebrand factor D’D3 assembly and structural principles for factor VIII binding and concatemer biogenesis. Blood 2019, 133, 1523–1533. [Google Scholar] [CrossRef]
- Shiltagh, N.; Kirkpatrick, J.; Cabrita, L.D.; McKinnon, T.A.J.; Thalassinos, K.; Tuddenham, E.G.D.; Hansen, D.F. Solution structure of the major factor VIII binding region on von Willebrand factor. Blood 2014, 123, 4143–4151. [Google Scholar] [CrossRef] [PubMed]
- Thomazini, C.M.; Sachetto, A.T.A.; de Albuquerque, C.Z.; Mattaraia, V.G.d.M.; de Oliveira, A.K.; Serrano, S.M.d.T.; Lebrun, I.; Barbaro, K.C.; Santoro, M.L. Involvement of von Willebrand factor and botrocetin in the thrombocytopenia induced by Bothrops jararaca snake venom. PLoS Negl. Trop. Dis. 2021, 15, e0009715. [Google Scholar] [CrossRef]
- Fukuda, K.; Doggett, T.; Laurenzi, I.J.; Liddington, R.C.; Diacovo, T.G. The snake venom protein botrocetin acts as a biological brace to promote dysfunctional platelet aggregation. Nat. Struct. Mol. Biol. 2005, 12, 152–159. [Google Scholar] [CrossRef]
- Andrews, R.K.; Booth, W.J.; Gorman, J.J.; Castaldi, P.A.; Berndt, M.C. Purification of Botrocetin from Bothrops jararaca Venom. Analysis of the Botrocetin-Mediated Interaction between von Willebrand Factor and the Human Platelet Membrane Glycoprotein Ib–IX Complex. Biochemistry 1989, 28, 8317–8326. [Google Scholar] [CrossRef]
- Read, M.S.; Smith, S.; Lamb, M.; Brinkhous, K. Role of botrocetin in platelet agglutination: Formation of an activated complex of botrocetin and von Willebrand factor. Blood 1989, 74, 1031–1035. [Google Scholar] [CrossRef]
- Howard, M.A.; Perkin, J.; Salem, H.H.; Firkin, B.G. The agglutination of human platelets by botrocetin: Evidence that botrocetin and ristocetin act at different sites on the factor VIII molecule and platelet membrane. Br. J. Haematol. 1984, 57, 25–35. [Google Scholar] [CrossRef]
- Pinto, A.F.; Terra, R.M.; Guimaraes, J.A.; Fox, J.W. Mapping von Willebrand factor A domain binding sites on a snake venom metalloproteinase cysteine-rich domain. Arch. Biochem. Biophys. 2007, 457, 41–46. [Google Scholar] [CrossRef]
- Matsui, T.; Hamako, J. Structure and function of snake venom toxins interacting with human von Willebrand factor. Toxicon 2005, 45, 1075–1087. [Google Scholar] [CrossRef] [PubMed]
- Allen, S.; Abuzenadah, A.M.; Blagg, J.L.; Hinks, J.; Nesbitt, I.M.; Goodeve, A.C.; Gursel, T.; Ingerslev, J.; Peake, I.R.; Daly, M.E. Two novel type 2N von Willebrand disease–causing mutations that result in defective factor VIII binding, multimerization, and secretion of von Willebrand factor. Blood 2000, 95, 2000–2007. [Google Scholar] [CrossRef]
- Maita, N.; Nishio, K.; Nishimoto, E.; Matsui, T.; Shikamoto, Y.; Morita, T.; Sadler, J.E.; Mizuno, H. Crystal structure of von Willebrand factor A1 domain complexed with snake venom, bitiscetin: Insight into glycoprotein Ibα binding mechanism induced by snake venom proteins. J. Biol. Chem. 2003, 278, 37777–37781. [Google Scholar] [CrossRef]
- Hanson, S.R.; Best, M.D.; Wong, C. Sulfatases: Structure, Mechanism, Biological Activity, Inhibition, and Synthetic Utility. Angew. Chem. Int. Ed. Engl. 2004, 43, 5736–5763. [Google Scholar] [CrossRef] [PubMed]
- Kovacs, Z.; Jung, I.; Gurzu, S. Arylsulfatases A and B: From normal tissues to malignant tumors. Pathol. Res. Pract. 2019, 215, 152516. [Google Scholar] [CrossRef]
- Lukatela, G.; Krauss, N.; Theis, K.; Selmer, T.; Gieselmann, V.; von Figura, K.; Saenger, W. Crystal Structure of Human Arylsulfatase A: The Aldehyde Function and the Metal Ion at the Active Site Suggest a Novel Mechanism for Sulfate Ester Hydrolysis, Biochemistry 1998, 37, 3654–3664. [Google Scholar] [CrossRef] [PubMed]
- Pospisil, P.; Kassis, A.I. Computational and Biological Evaluation of Radioiodinated Quinazolinone Prodrug for Targeting Pancreatic Cancer. Mol. Diagn. Treat. Pancreat. 2014, 1, 385–403. [Google Scholar]
- Linhardt, R.J.; Toida, T. Role of Glycosaminoglycans in Cellular Communication. Accounts Chem. Res. 2004, 37, 431–438. [Google Scholar] [CrossRef]
- Makita, A.; Taniguchi, N. Glycosphingolipids. New Compr. Biochem. 1985, 10, 1–99. [Google Scholar] [CrossRef]
- Abe, A.; Shayman, J. Sphingolipid Catabolism. In Encyclopedia of Biological Chemistry, 2nd ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2013; pp. 287–292. [Google Scholar] [CrossRef]
- Frati-Munari, A.C. Glicosaminoglicanos en las enfermedades vasculares. Rev. Mex. Angiol. 2012, 40, 89–99. [Google Scholar]
- Bose, U.; Wang, T.; Zhao, M.; Motti, C.A.; Hall, M.R.; Cummins, S.F. Multiomics analysis of the giant triton snail salivary gland, a crown-of-thorns starfish predator. Sci. Rep. 2017, 7, 6000. [Google Scholar] [CrossRef] [PubMed]
- Nok, A.J.; Abubakar, M.S.; Adaudi, A.; Balogun, E. Aryl sulfatase from Naja nigricolis venom: Characterization and possible contribution in the pathology of snake poisoning. J. Biochem. Mol. Toxicol. 2003, 17, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Scieuzo, C.; Salvia, R.; Franco, A.; Pezzi, M.; Cozzolino, F.; Chicca, M.; Scapoli, C.; Vogel, H.; Monti, M.; Ferracini, C.; et al. An integrated transcriptomic and proteomic approach to identify the main Torymus sinensis venom components. Sci. Rep. 2021, 11, 5032. [Google Scholar] [CrossRef] [PubMed]
- Nystrom, G.S.; Ellsworth, S.A.; Rokyta, D.R. The remarkably enzyme-rich venom of the Big Bend Scorpion (Diplocentrus whitei). Toxicon 2023, 226, 107080. [Google Scholar] [CrossRef]
- Lane, A.N.; Nash, P.D.; Ellsworth, S.A.; Nystrom, G.S.; Rokyta, D.R. The arylsulfatase- and phospholipase-rich venom of the plutoniumid centipede Theatops posticus. Toxicon 2023, 233, 107231. [Google Scholar] [CrossRef]
- Yu, M.; Wu, M.; Secundo, F.; Liu, Z. Detection, production, modification, and application of arylsulfatases. Biotechnol. Adv. 2023, 67, 108207. [Google Scholar] [CrossRef]
- Kertesz, M.A. Riding the sulfur cycle—Metabolism of sulfonates and sulfate esters in Gram-negative bacteria. FEMS Microbiol. Rev. 2000, 24, 135–175. [Google Scholar] [CrossRef]
- Stressler, T.; Seitl, I.; Kuhn, A.; Fischer, L. Detection, production, and application of microbial arylsulfatases. Appl. Microbiol. Biotechnol. 2016, 100, 9053–9067. [Google Scholar] [CrossRef]
- Bond, C.S.; Clements, P.R.; Ashby, S.J.; Collyer, C.A.; Harrop, S.J.; Hopwood, J.J.; Guss, J.M. Structure of a human lysosomal sulfatase. Structure 1997, 5, 277–289. [Google Scholar] [CrossRef]
- Boltes, I.; Czapinska, H.; Kahnert, A.; von Bülow, R.; Dierks, T.; Schmidt, B.; von Figura, K.; Kertesz, M.A.; Usón, I. 1.3 Å Structure of Arylsulfatase from Pseudomonas aeruginosa Establishes the Catalytic Mechanism of Sulfate Ester Cleavage in the Sulfatase Family. Structure 2001, 9, 483–491. [Google Scholar] [CrossRef]
- Usami, Y.; Fujimura, Y.; Suzuki, M.; Ozeki, Y.; Nishio, K.; Fukui, H.; Titani, K. Primary structure of two-chain botrocetin, a von Willebrand factor modulator purified from the venom of Bothrops jararaca. Proc. Natl. Acad. Sci. USA 1993, 90, 928–932. [Google Scholar] [CrossRef] [PubMed]
- L. and B. I. (NIH) National Heart, Trastornos Plaquetarios—Trombocitopenia. Available online: https://www.nhlbi.nih.gov/es/salud/trombocitopenia (accessed on 3 April 2025).
- Shen, C.; Liu, M.; Mackeigan, D.T.; Chen, Z.Y.; Chen, P.; Karakas, D.; Li, J.; Norris, P.A.A.; Li, J.; Deng, Y.; et al. Viper venoms drive the macrophages and hepatocytes to sequester and clear platelets: Novel mechanism and therapeutic strategy for venom-induced thrombocytopenia. Arch. Toxicol. 2021, 95, 3589–3599. [Google Scholar] [CrossRef] [PubMed]
- Arlinghaus, F.T.; Eble, J.A. C-type lectin-like proteins from snake venoms. Toxicon 2012, 60, 512–519. [Google Scholar] [CrossRef]
- Sen, U.; Vasudevan, S.; Subbarao, G.; McClintock, R.A.; Celikel, R.; Ruggeri, Z.M.; Varughese, K.I. Crystal Structure of the von Willebrand Factor Modulator Botrocetin. Biochemistry 2000, 40, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Dutta, R.; Yoshida, T.; Inouye, M. The critical role of the conserved Thr247 residue in the functioning of the osmosensor EnvZ, a histidine kinase/phosphatase, in Escherichia coli. J. Biol. Chem. 2000, 275, 38645–38653. [Google Scholar] [CrossRef]
- Soto, M.; Kamiguti, A.S.; Theakston, D.G.; Fox, J.W.; Escalante, T.; Gutiérrez, J.M.; Rucavado, A. Characterization of Aspercetin, a Platelet Aggregating Component from the Venom of the Snake Bothrops asper which Induces Thrombocytopenia and Potentiates Metalloproteinase-induced Hemorrhage. Thromb. Haemost. 2001, 85, 710–715. [Google Scholar] [CrossRef]
- Rucavado Romero, A.; Soto Morera, M.; Escalante Muñoz, T.; Loría Masís, G.D.; Arni, R.K.; Gutiérrez, J.M. Thrombocytopenia and platelet hypoaggregation induced by Bothrops asper snake venom. Toxins involved and their contribution to metalloproteinase-induced pulmonary hemorrhage. Thromb Haemost 2005, 94, 123–131. [Google Scholar] [CrossRef]
- Markland, F.S. Snake venoms and the hemostatic system. Toxicon 1998, 36, 1749–1800. [Google Scholar] [CrossRef]
- Angulo, Y.; Lomonte, B. Biochemistry and toxicology of toxins purified from the venom of the snake Bothrops asper. Toxicon 2009, 54, 949–957. [Google Scholar] [CrossRef]
- Kubota, T.; Tani, O.; Yamaguchi, T.; Namatame, I.; Sakashita, H.; Furukawa, K.; Yamasaki, K. Crystal structures of FMN-bound and FMN-free forms of dihydroorotate dehydrogenase from Trypanosoma Brucei. FEBS Open Bio. 2018, 8, 680–691. [Google Scholar] [CrossRef]
- Liu, S.; Neidhardt, E.A.; Grossman, T.H.; Ocain, T.; Clardy, J. Structures of human dihydroorotate dehydrogenase in complex with antiproliferative agents. Structure 2000, 8, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Rowland, P.; Larsen, S.; Björnberg, O.; Nielsen, F.S.; Jensen, K.F. The crystal structure of lactococcus lactis dihydroorotate dehydrogenase A complexed with the enzyme reaction product throws light on its enzymatic function. Protein Sci. 1998, 7, 1269–1279. [Google Scholar] [CrossRef] [PubMed]
- Madak, J.T.; Bankhead, A.; Cuthbertson, C.R.; Showalter, H.D.; Neamati, N. Revisiting the role of dihydroorotate dehydrogenase as a therapeutic target for cancer. Pharmacol. Ther. 2019, 195, 111–131. [Google Scholar] [CrossRef] [PubMed]
- Reis, R.A.; Calil, F.A.; Feliciano, P.R.; Pinheiro, M.P.; Nonato, M.C. The dihydroorotate dehydrogenases: Past and present. Arch. Biochem. Biophys. 2017, 632, 175–191. [Google Scholar] [CrossRef]
- Ulanet, D.; Chubukov, V.; Coco, J.; McDonald, G.; Steadman, M.; Narayanaswamy, R.; Ronseaux, S.; Choe, S.; Erdmann, T.; Truskowski, K.; et al. Hematologic Malignancies Exhibit Selective Vulnerability to Inhibition of De Novo Pyrimidine Biosynthesis By AG-636, a Novel Inhibitor of Dihydroorotate Dehydrogenase in Phase 1 Clinical Trials. Blood 2019, 134, 1570. [Google Scholar] [CrossRef]
- Kurth, I.; Yamaguchi, N.; Andreu-Agullo, C.; Tian, H.S.; Sridhar, S.; Takeda, S.; Gonsalves, F.C.; Loo, J.M.; Barlas, A.; Manova-Todorova, K.; et al. Therapeutic targeting of SLC6A8 creatine transporter suppresses colon cancer progression and modulates human creatine levels. Sci. Adv. 2021, 7, eabi7511. [Google Scholar] [CrossRef]
- Dobritzsch, D.; Schneider, G.; Schnackerz, K.D.; Lindqvist, Y. Crystal structure of dihydropyrimidine dehydrogenase, a major determinant of the pharmacokinetics of the anti-cancer drug 5-fluorouracil. EMBO J. 2001, 20, 650–660. [Google Scholar] [CrossRef]
- Smith, M.M.; Forouzesh, D.C.; Kaley, N.E.; Liu, D.; Moran, G.R. Mammalian dihydropyrimidine dehydrogenase: Added mechanistic details from transient-state analysis of charge transfer complexes. Arch. Biochem. Biophys. 2023, 736, 109517. [Google Scholar] [CrossRef]
- Giannotti, K.C.; Sesso, A.; Grego, K.F.; Fernandes, W.; Cardoso, R.P.; Camargo, G.G.; Carneiro, S.M. Viperid venom glands with defective venom production. Morphological study. Toxicon 2013, 70, 32–43. [Google Scholar] [CrossRef]
- Mackessy, S.P. Morphology and ultrastructure of the venom glands of the northern pacific rattlesnake Crotalus viridis oreganus. J. Morphol. 1991, 208, 109–128. [Google Scholar] [CrossRef]
- Weinstein, S.A. Reptile venom glands: Form, function, future, concepts and controversies. In Handbook of Venoms and Toxins of Reptiles; Mackessy, S.P., Ed.; CRC Press: Boca Raton, FL, USA, 2021; Volume 2, pp. 99–115. [Google Scholar]
- Mackessy, S.P.; Baxter, L.M. Bioweapons synthesis and storage: The venom gland of front-fanged snakes. Zool. Anz. A J. Comp. Zool. 2006, 245, 147–159. [Google Scholar] [CrossRef]
- Shambaugh, G. Pyrimidine biosynthesis. Am. J. Clin. Nutr. 1979, 32, 1290–1297. [Google Scholar] [CrossRef] [PubMed]
- Copeland, R.A.; Davis, J.P.; Dowling, R.L.; Lombardo, D.; Murphy, K.B.; Patterson, T.A. Recombinant Human Dihydroorotate Dehydrogenase: Expression, Purification, and Characterization of a Catalytically Functional Truncated Enzyme. Arch. Biochem. Biophys. 1995, 323, 79–86. [Google Scholar] [CrossRef]
- Jones, M.E. Pyrimidine nucleotide biosynthesis in animals: Genes, enzymes, and regulation of UMP biosynthesis. Annu Rev. Biochem. 1980, 49, 253–279. [Google Scholar] [CrossRef] [PubMed]
- Schreuder, M.; Reitsma, P.H.; Bos, M.H. Blood coagulation factor Va’s key interactive residues and regions for prothrombinase assembly and prothrombin binding. J. Thromb. Haemost. 2019, 17, 1229–1239. [Google Scholar] [CrossRef]
- Nishida, S.; Fujita, T.; Kohno, N.; Atoda, H.; Morita, T.; Takeya, H.; Kido, I.; Paine, M.J.I.; Kawabata, S.-I.; Iwanaga, S. cDNA cloning and deduced amino acid sequence of prothrombin activator (ecarin) from Kenyan Echis carinatus venom. Biochemistry 1995, 34, 1771–1778. [Google Scholar] [CrossRef]
- Loría, G.D.; Rucavado, A.; Kamiguti, A.S.; Theakston, R.G.; Fox, J.W.; Alape, A.; Gutiérrez, J.M. Characterization of ‘basparin A’, a prothrombin-activating metalloproteinase, from the venom of the snake Bothrops asper that inhibits platelet aggregation and induces defibrination and thrombosis. Arch. Biochem. Biophys. 2003, 418, 13–24. [Google Scholar] [CrossRef]
- Paine, M.; Desmond, H.; Theakston, R.; Crampton, J. Purification, cloning, and molecular characterization of a high molecular weight hemorrhagic metalloprotease, jararhagin, from Bothrops jararaca venom. Insights into the disintegrin gene family. J. Biol. Chem. 1992, 267, 22869–22876. [Google Scholar] [CrossRef]
- Zhou, Q.; Smith, J.B.; Grossman, M.H. Molecular cloning and expression of catrocollastatin, a snake-venom protein from Crotalus atrox (western diamondback rattlesnake) which inhibits platelet adhesion to collagen. Biochem. J. 1995, 307, 411–417. [Google Scholar] [CrossRef]
- Olaoba, O.T.; dos Santos, P.K.; Selistre-De-Araujo, H.S.; de Souza, D.H.F. Snake Venom Metalloproteinases (SVMPs): A structure-function update. Toxicon X 2020, 7, 100052. [Google Scholar] [CrossRef]
- Bell, J.; Markland, J. Defibrinogenating enzymes. Drugs 1997, 54 (Suppl. S3), 18–31. [Google Scholar] [CrossRef] [PubMed]
- Herrera, C.; Escalante, T.; Rucavado, A.; Gutiérrez, J.M. Hemorrhagic and procoagulant P-III snake venom metalloproteinases differ in their binding to the microvasculature of mouse cremaster muscle. Toxicon 2020, 178, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Dawson, C.A.; Ainsworth, S.; Albulescu, L.O.; Casewell, N.R. Snake venom metalloproteinases. In Handbook of Venoms and Toxins of Reptiles; Mackessy, S.P., Ed.; CRC press: Boca Raton, FL, USA, 2021; Volume 2, pp. 363–380. [Google Scholar]
- Brust, A.; Sunagar, K.; Undheim, E.A.; Vetter, I.; Yang, D.C.; Casewell, N.R.; Jackson, T.N.; Koludarov, I.; Alewood, P.F.; Hodgson, W.C.; et al. Differential Evolution and Neofunctionalization of Snake Venom Metalloprotease Domains. Mol. Cell. Proteom. 2013, 12, 651–663. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Gao, Y.; Zhu, Z.; Yu, Y.; Zhang, X.; Zang, J.; Teng, M.; Niu, L. Structural basis of the autolysis of AaHIV suggests a novel target recognizing model for ADAM/reprolysin family proteins. Biochem. Biophys. Res. Commun. 2009, 386, 159–164. [Google Scholar] [CrossRef]
- Guo, X.-X.; Zeng, L.; Lee, W.-H.; Zhang, Y.; Jin, Y. Isolation and cloning of a metalloproteinase from king cobra snake venom. Toxicon 2007, 49, 954–965. [Google Scholar] [CrossRef]
- Souza, D.; Iemma, M.; Ferreira, L.; Faria, J.; Oliva, M.; Zingali, R.; Niewiarowski, S.; Selistre-De-Araujo, H. The Disintegrin-like Domain of the Snake Venom Metalloprotease Alternagin Inhibits α2β1 Integrin-Mediated Cell Adhesion. Arch. Biochem. Biophys. 2000, 384, 341–350. [Google Scholar] [CrossRef]
- Calvete, J.J.; Moreno-Murciano, M.P.; Sanz, L.; Jurgens, M.; Schrader, M.; Raida, M.; Benjamin, D.C.; Fox, J.W. The disulfide bond pattern of catrocollastatin C, a disintegrin-like/cysteine-rich protein isolated from Crotalus atrox venom. Protein Sci. 2000, 9, 1365–1373. [Google Scholar] [CrossRef]
- Takeda, S.; Igarashi, T.; Mori, H.; Araki, S. Crystal structures of VAP1 reveal ADAMs’ MDC domain architecture and its unique C-shaped scaffold. EMBO J. 2006, 25, 2388–2396. [Google Scholar] [CrossRef]
- Sajevic, T.; Leonardi, A.; Kovačič, L.; Lang-Balija, M.; Kurtović, T.; Pungerčar, J.; Halassy, B.; Trampuš-Bakija, A.; Križaj, I. VaH3, one of the principal hemorrhagins in Vipera ammodytes ammodytes venom, is a homodimeric P-IIIc metalloproteinase. Biochimie 2013, 95, 1158–1170. [Google Scholar] [CrossRef]
- Takeda, S.; Igarashi, T.; Mori, H. Crystal structure of RVV-X: An example of evolutionary gain of specificity by ADAM proteinases. FEBS Lett. 2007, 581, 5859–5864. [Google Scholar] [CrossRef]
- Takeya, H.; Nishida, S.; Miyata, T.; Kawada, S.; Saisaka, Y.; Morita, T.; Iwanaga, S. Coagulation factor X activating enzyme from Russell’s viper venom (RVV-X). A novel metalloproteinase with disintegrin (platelet aggregation inhibitor)-like and C-type lectin-like domains. J. Biol. Chem. 1992, 267, 14109–14117. [Google Scholar] [CrossRef] [PubMed]
- Leonardi, A.; Sajevic, T.; Pungerčar, J.; Križaj, I. Comprehensive Study of the Proteome and Transcriptome of the Venom of the Most Venomous European Viper: Discovery of a New Subclass of Ancestral Snake Venom Metalloproteinase Precursor-Derived Proteins. J. Proteome Res. 2019, 18, 2287–2309. [Google Scholar] [CrossRef] [PubMed]
- Muniz, J.R.; Ambrosio, A.L.; Selistre-De-Araujo, H.S.; Cominetti, M.R.; Moura-Da-Silva, A.M.; Oliva, G.; Garratt, R.C.; Souza, D.H. The three-dimensional structure of bothropasin, the main hemorrhagic factor from Bothrops jararaca venom: Insights for a new classification of snake venom metalloprotease subgroups. Toxicon 2008, 52, 807–816. [Google Scholar] [CrossRef] [PubMed]
- Takeda, S.; Takeya, H.; Iwanaga, S. Snake venom metalloproteinases: Structure, function and relevance to the mammalian ADAM/ADAMTS family proteins. Biochim. Biophys. Acta Proteins Proteom. 2012, 1824, 164–176. [Google Scholar] [CrossRef]
- Bode, W.; Gomis-Rüth, F.-X.; Stöckler, W. Astacins, serralysins, snake venom and matrix metalloproteinases exhibit identical zinc-binding environments (HEXXHXXGXXH and Met-turn) and topologies and should be grouped into a common family, the ‘metzincins’. FEBS Lett. 1993, 331, 134–140. [Google Scholar] [CrossRef]
- Rawlings, N.D.; Barrett, A.J. [13] Evolutionary families of metallopeptidases. Methods Enzymol. 1995, 248, 183–228. [Google Scholar] [CrossRef]
- Gong, W.; Zhu, X.; Liu, S.; Teng, M.; Niu, L. Crystal structures of acutolysin A, a three-disulfide hemorrhagic zinc metalloproteinase from the snake venom of Agkistrodon acutus. J. Mol. Biol. 1998, 283, 657–668. [Google Scholar] [CrossRef]
- Watanabe, L.; Shannon, J.D.; Valente, R.H.; Rucavado, A.; Alape-Girón, A.; Kamiguti, A.S.; Theakston, R.D.G.; Fox, J.W.; Gutiérrez, J.M.; Arni, R.K. Amino acid sequence and crystal structure of BaP1, a metalloproteinase from Bothrops asper snake venom that exerts multiple tissue-damaging activities. Protein Sci. 2003, 12, 2273–2281. [Google Scholar] [CrossRef]
- Arce-Estrada, V.; Azofeifa-Cordero, G.; Estrada, R.; Alape-Girón, A.; Flores-Díaz, M. Neutralization of venom-induced hemorrhage by equine antibodies raised by immunization with a plasmid encoding a novel P-II metalloproteinase from the lancehead pitviper Bothrops asper. Vaccine 2009, 27, 460–466. [Google Scholar] [CrossRef]
- Fox, J.W.; Serrano, S.M.T. Insights into and speculations about snake venom metalloproteinase (SVMP) synthesis, folding and disulfide bond formation and their contribution to venom complexity. FEBS J. 2008, 275, 3016–3030. [Google Scholar] [CrossRef]
- Van de Velde, A.C.; Gay, C.C.; Moritz, M.N.d.O.; dos Santos, P.K.; Bustillo, S.; Rodríguez, J.P.; Acosta, O.C.; Biscoglio, M.J.; Selistre-De-Araujo, H.S.; Leiva, L.C. Purification of a fragment obtained by autolysis of a PIIIb-SVMP from Bothrops alternatus venom. Int. J. Biol. Macromol. 2018, 113, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Igarashi, T.; Araki, S.; Mori, H.; Takeda, S. Crystal structures of catrocollastatin/VAP2B reveal a dynamic, modular architecture of ADAM/adamalysin/reprolysin family proteins. FEBS Lett. 2007, 581, 2416–2422. [Google Scholar] [CrossRef] [PubMed]
- Omar, S.; Tap, F.M.; Shameli, K.; Ali, R.R.; Jusoh, N.W.C.; Khairudin, N.B.A. Sequence analysis and comparative modelling of nucleocapsid protein from pseudomonas stutzeri. IOP Conf. Ser. Mater. Sci. Eng. 2018, 458, 012025. [Google Scholar] [CrossRef]
- Dym, O.; Eisenberg, D.; Yeates, T.O. Detection of errors in protein models. Int. Tables Crystallogr. 2006, 520–530. [Google Scholar] [CrossRef]
- Mirdita, M.; Schütze, K.; Moriwaki, Y.; Heo, L.; Ovchinnikov, S.; Steinegger, M. ColabFold: Making protein folding accessible to all. Nat. Methods 2022, 19, 679–682. [Google Scholar] [CrossRef]
- Yang, Z.; Zeng, X.; Zhao, Y.; Chen, R. AlphaFold2 and its applications in the fields of biology and medicine. Signal Transduct. Target. Ther. 2023, 8, 115. [Google Scholar] [CrossRef]
- Laskowski, R.A.; Rullmannn, J.A.; MacArthur, M.W.; Kaptein, R.; Thornton, J.M. AQUA and PROCHECK-NMR: Programs for checking the quality of protein structures solved by NMR. J. Biomol. NMR 1996, 8, 477–486. [Google Scholar] [CrossRef]
- Colovos, C.; Yeates, T.O. Verification of protein structures: Patterns of nonbonded atomic interactions. Protein Sci. 1993, 2, 1511–1519. [Google Scholar] [CrossRef]
- Eisenberg, D.; Lüthy, R.; Bowie, J.U. VERIFY3D: Assessment of protein models with three-dimensional profiles. Methods Enzym. 1997, 277, 396–404. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Proteins | ERRAT | RAMACHANDRAN | Verify3D | |
|---|---|---|---|---|
| Allergen 5 | B. asper | 95.74% | 92.3% | 92% |
| B. jararaca | 93.52% | 90.3% | 98.64% | |
| Von Willebrand Factor | B. asper | 84% | 81.3% | 48.45% |
| B. jararaca | 84.53% | 88.5% | 76.14% | |
| Arylsulfatase | B. asper | 86.62% | 86.7% | 73.68% |
| B. jararaca | 80.89% | 88.6% | 72% | |
| Botrocetin | B. asper | 84.34% | 87% | 56.91% |
| B. jararaca | 82.14% | 82.8% | 61.42% | |
| Dihydroorotate dehydrogenase | B. asper | 96.51% | 92.7% | 81.41% |
| B. jararaca | 99% | 92.1% | 93.13% | |
| Basparin | B. asper | 100% | 82% | 1.79% |
| Bothropasin | B. asper | 86% | 85% | 59% |
| Protein | Species | ID |
|---|---|---|
| Allergen 5 | Vespula germanica | P35784 |
| Apolipophorin | Odontomachus monticola | A0A348G622 |
| von Willebrand factor | Homo sapiens | P04275 |
| Arylsulfatase I | Ophiophagus hannah | V8PBU1 |
| Botrocetin | Bothrops jararaca | P22030 |
| Dihydroorotate dehydrogenase | Homo sapiens | Q02127 |
| Basparin | Bothrops asper | P84035 |
| Bothropasin | Bothrops jararaca | O93523 |
| Protein | Species | ID |
|---|---|---|
| Allergen 5 | Vespula germanica | UniProt P35784 |
| Triflin | Trimeresurus flavoviridis | PDB 1WVR |
| vWFD | Homo sapiens | PDB 6N29 |
| Arylsulfatase | Ophiophagus hannah | UniProt V8PBU1 |
| Arylsulfatase B | Homo sapiens | PDB 1FSU |
| Botrocetin | Bothrops jararaca | PDB 1FVU |
| Aspercetin | Bothrops asper | UniProt P0DJC9 |
| Dihydroorotate dehydrogenase | Homo sapiens | PDB 3U2O |
| Basparin A | Bothrops asper | UniProt P84035 |
| Bothropasin | Bothrops jararaca | PDB 3DSL |
| Bothrasperin | Bothrops asper | UniProt Q072L5 |
| Snake venom metalloproteinase BaP1 | Bothrops asper | PDB 1ND1 |
| Zinc metalloproteinase-disintegrin-like VAP2B | Crotalus atrox | PDB 2DW0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Espín-Angulo, J.; Vela, D. Computational Modeling of Low-Abundance Proteins in Venom Gland Transcriptomes: Bothrops asper and Bothrops jararaca. Toxins 2025, 17, 262. https://doi.org/10.3390/toxins17060262
Espín-Angulo J, Vela D. Computational Modeling of Low-Abundance Proteins in Venom Gland Transcriptomes: Bothrops asper and Bothrops jararaca. Toxins. 2025; 17(6):262. https://doi.org/10.3390/toxins17060262
Chicago/Turabian StyleEspín-Angulo, Joseph, and Doris Vela. 2025. "Computational Modeling of Low-Abundance Proteins in Venom Gland Transcriptomes: Bothrops asper and Bothrops jararaca" Toxins 17, no. 6: 262. https://doi.org/10.3390/toxins17060262
APA StyleEspín-Angulo, J., & Vela, D. (2025). Computational Modeling of Low-Abundance Proteins in Venom Gland Transcriptomes: Bothrops asper and Bothrops jararaca. Toxins, 17(6), 262. https://doi.org/10.3390/toxins17060262