Impact of Low-Level Ergot Alkaloids and Endophyte Presence in Tall Fescue Grass on the Metabolome and Microbiome of Fall-Grazing Steers

 , , ,
, , ,
Abstract
1. Introduction
2. Results
2.1. Physiological Parameters
2.2. Endophyte and Total Plant Ergot Alkaloids
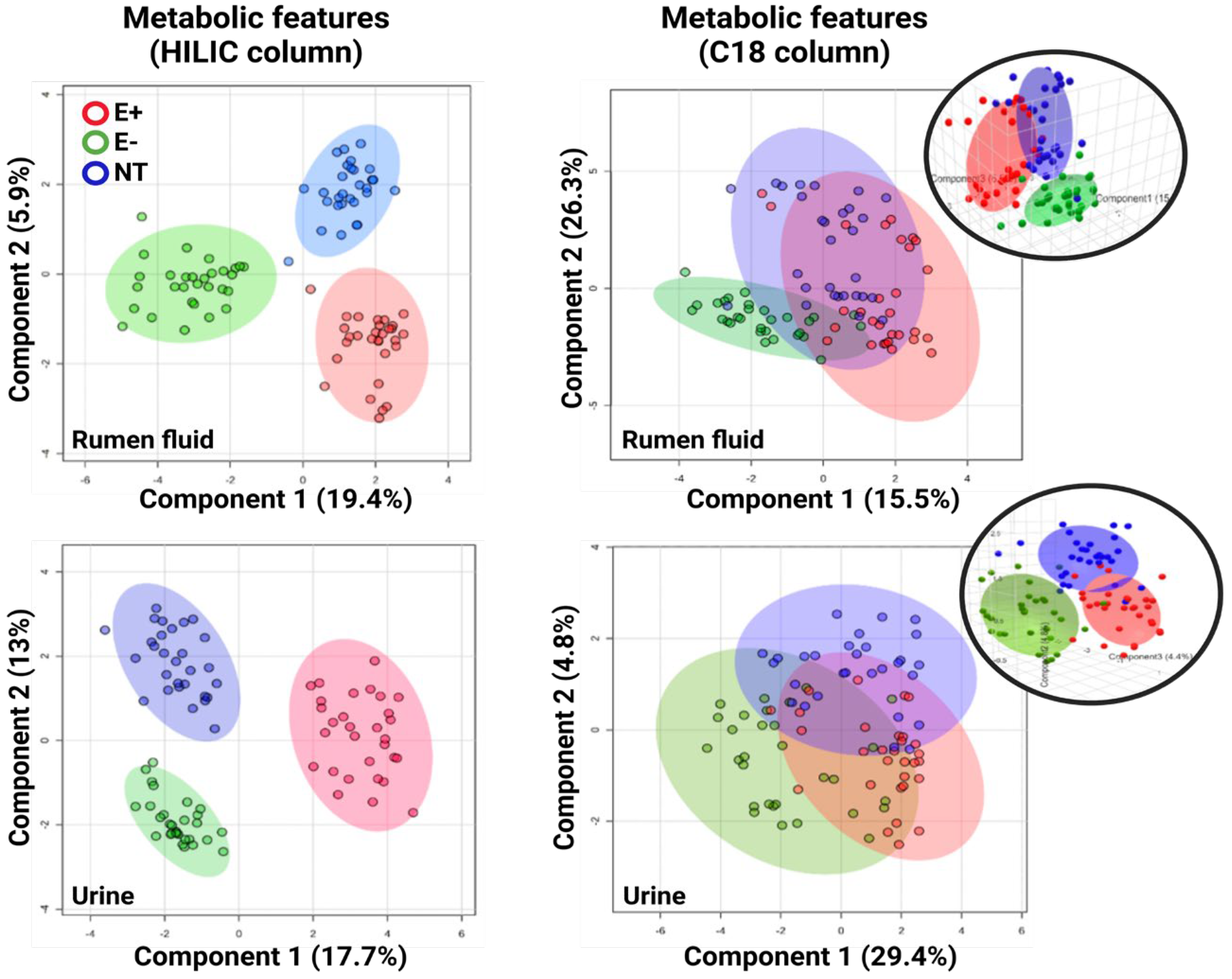
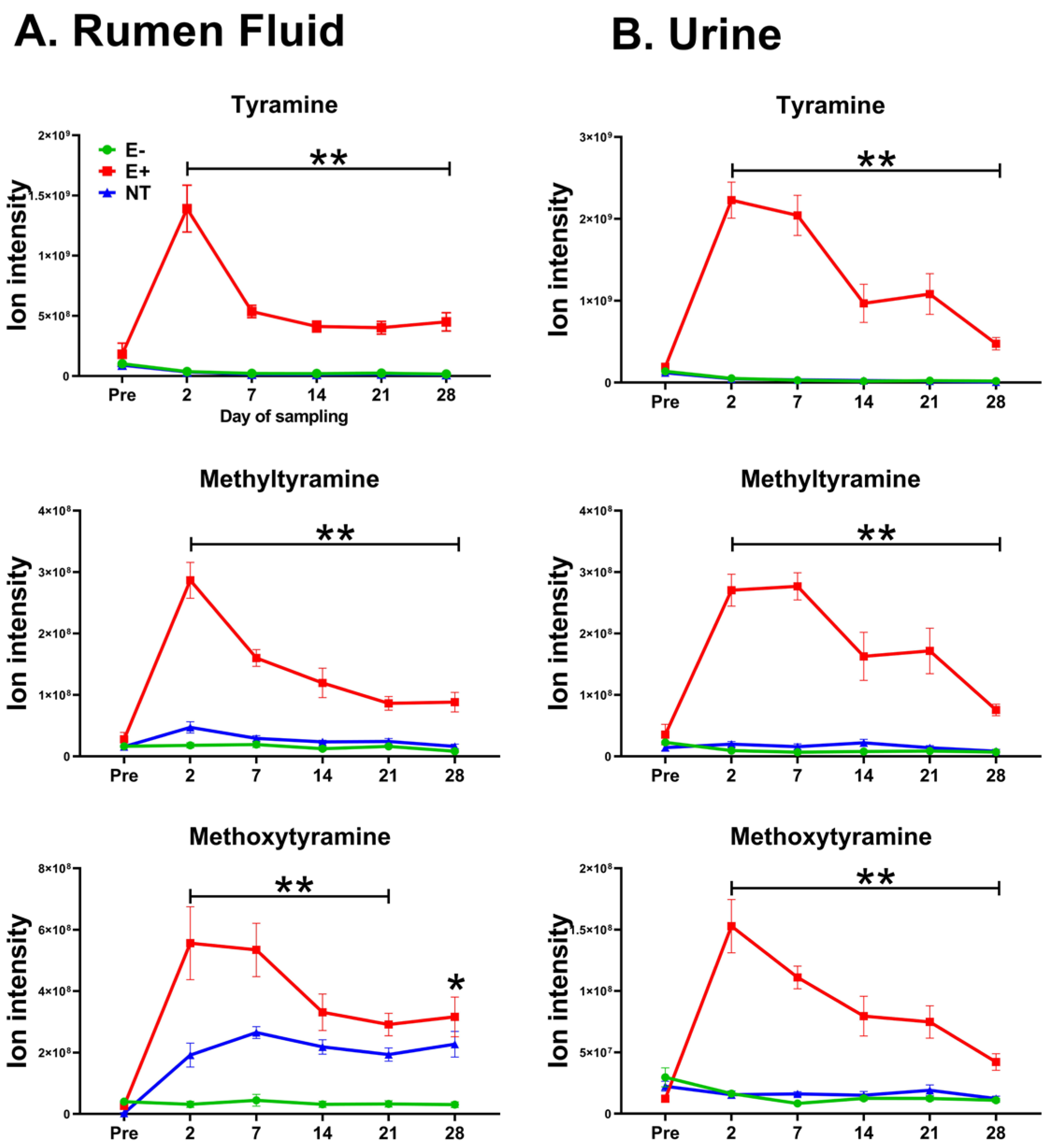
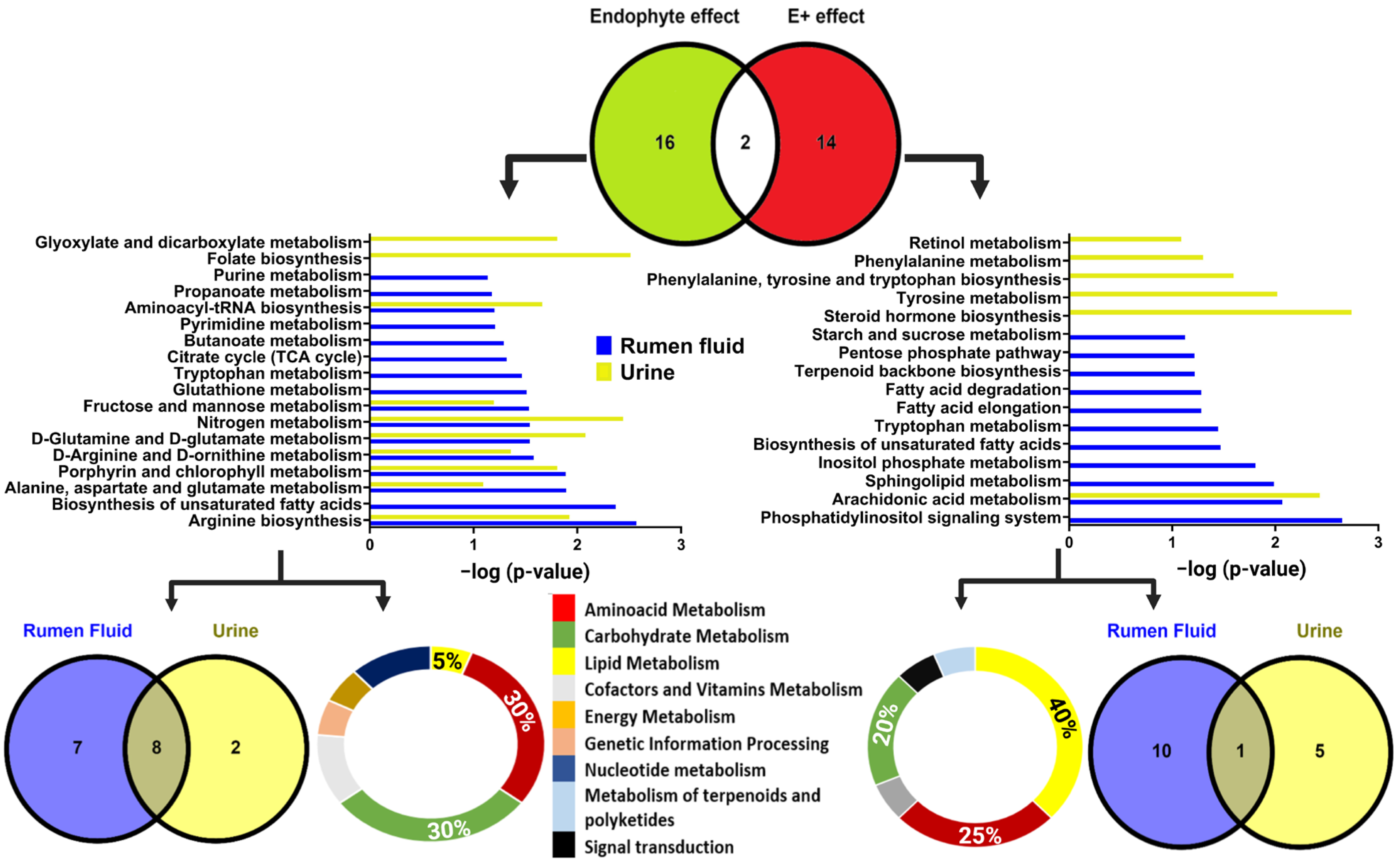
2.3. Metabolomics Results
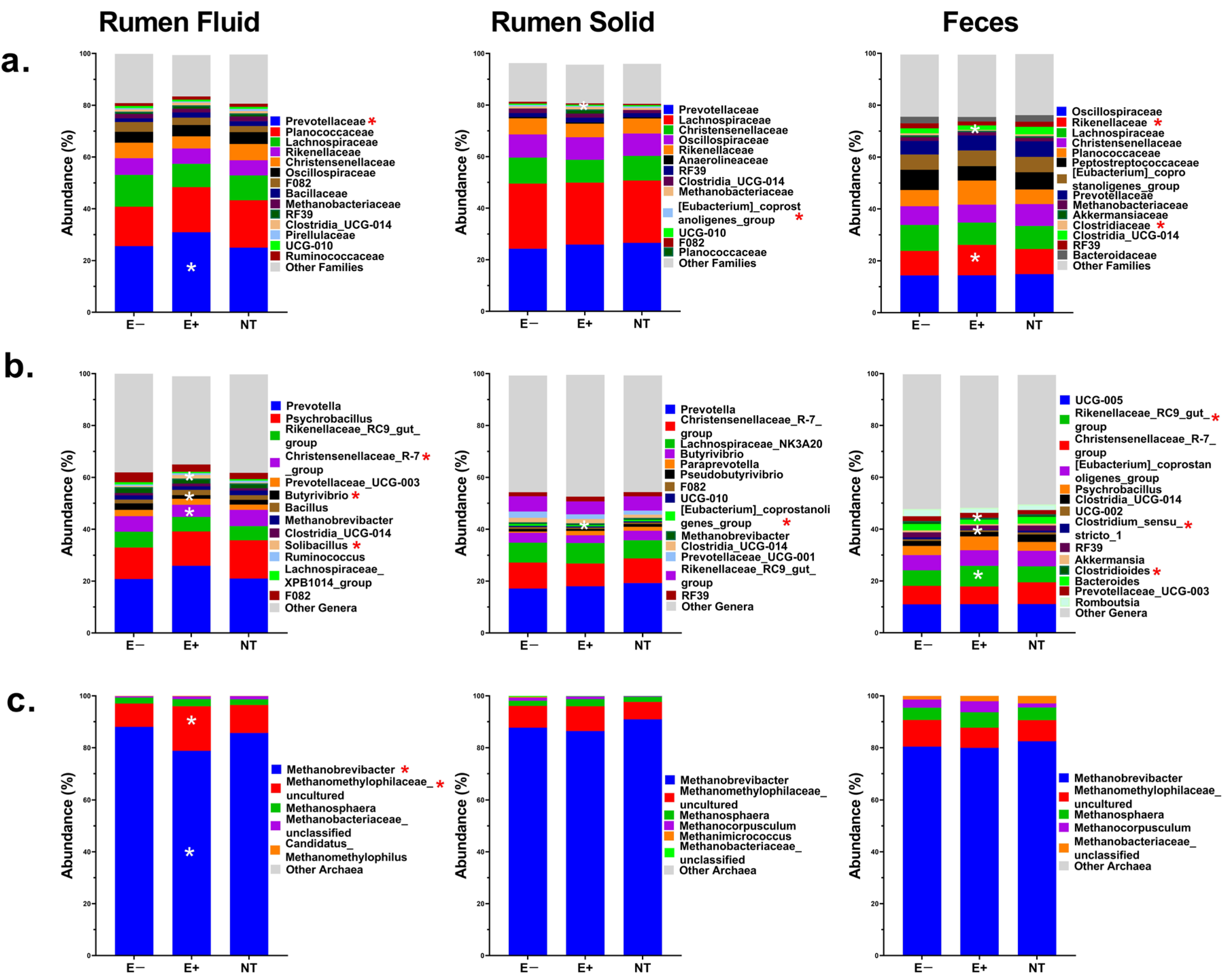
2.4. Microbiome Results
2.4.1. 16S rRNA Gene Sequencing
2.4.2. Alpha Diversity
2.4.3. Beta Diversity
2.4.4. Specific Microbial Taxa
3. Discussion
4. Conclusions
5. Materials and Methods
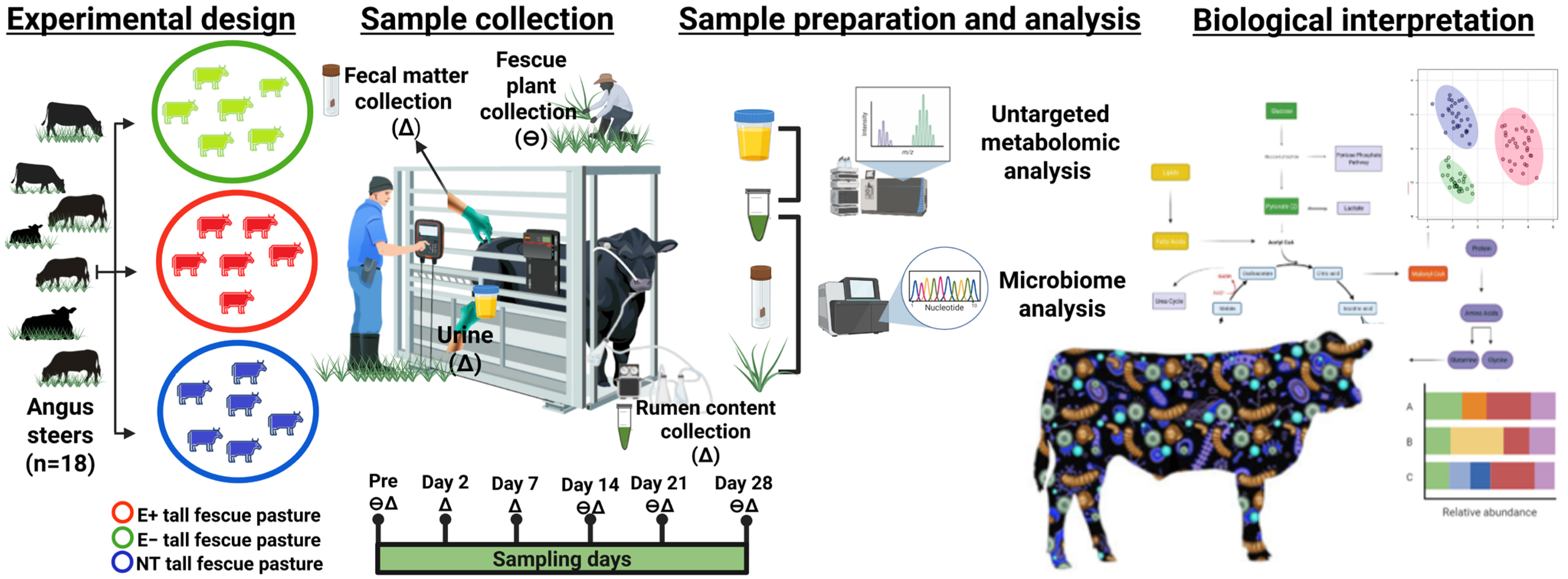
5.1. Animals, Treatments, and Experimental Design
5.2. Sample Collection and Processing
Endophyte Detection and Total Plant Ergot Alkaloids Analysis
5.3. Metabolomics Sample Processing and Data Analysis
5.3.1. Metabolomics Sample Processing
5.3.2. Urine and Rumen Fluid Metabolite Annotations
5.3.3. Metabolomics Data Processing and Statistical Analysis
5.4. Microbiome Sample Processing and Data Analysis
5.4.1. DNA Extraction
5.4.2. DNA Amplification and Sequencing
5.4.3. 16S rRNA Gene Sequence Processing and Bioinformatics Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gallo, A.; Giuberti, G.; Frisvad, J.C.; Bertuzzi, T.; Nielsen, K.F. Review on mycotoxin issues in ruminants: Occurrence in forages, effects of mycotoxin ingestion on health status and animal performance and practical strategies to counteract their negative effects. Toxins 2015, 7, 3057–3111. [Google Scholar] [CrossRef] [PubMed]
- Clay, K.; Schardl, C. Evolutionary origins and ecological consequences of endophyte symbiosis with grasses. Am. Nat. 2002, 160, S99–S127. [Google Scholar] [CrossRef] [PubMed]
- Oliver, J.W. Pathophysiologic response to endophyte toxins. In Neotyphodium in Cool-Season Grasses; Craig, A.R., West, C.P., Spiers, D.E., Eds.; Blackwell Publishing: Ames, IA, USA, 2005; Volume 1, pp. 291–304. [Google Scholar]
- Bacon, C.; Porter, J.; Robbins, J.; Luttrell, E. Epichloe typhina from toxic tall fescue grasses. Appl. Environ. Microbiol. 1977, 34, 576–581. [Google Scholar] [CrossRef]
- Hill, N.; Thompson, F.; Stuedemann, J.; Rottinghaus, G.; Ju, H.; Dawe, D.; Hiatt, E., III. Ergot alkaloid transport across ruminant gastric tissues. J. Anim. Sci. 2001, 79, 542–549. [Google Scholar] [CrossRef]
- Lyons, P.C.; Plattner, R.D.; Bacon, C.W. Occurrence of peptide and clavine ergot alkaloids in tall fescue grass. Science 1986, 232, 487–489. [Google Scholar] [CrossRef]
- Hovermale, J.T.; Craig, A.M. Correlation of ergovaline and lolitrem B levels in endophyte-infected perennial ryegrass (Lolium perenne). J. Vet. Diagn. Investig. 2001, 13, 323–327. [Google Scholar] [CrossRef]
- Rhodes, M.; Paterson, J.; Kerley, M.; Garner, H.; Laughlin, M. Reduced blood flow to peripheral and core body tissues in sheep and cattle induced by endophyte-infected tall fescue. J. Anim. Sci. 1991, 69, 2033–2043. [Google Scholar] [CrossRef]
- Aiken, G.; Kirch, B.; Strickland, J.; Bush, L.; Looper, M.; Schrick, F. Hemodynamic responses of the caudal artery to toxic tall fescue in beef heifers. J. Anim. Sci. 2007, 85, 2337–2345. [Google Scholar] [CrossRef]
- Spiers, D.E.; Evans, T.J.; Rottinghaus, G.E. Interaction between thermal stress and fescue toxicosis: Animal models and new perspectives. In Neotyphodium in Cool-Season Grasses; Craig, A.R., West, C.P., Spiers, D.E., Eds.; Blackwell Publishing: Ames, IA, USA, 2005; Volume 1, pp. 243–270. [Google Scholar]
- Foote, A.P.; Kristensen, N.B.; Klotz, J.L.; Kim, D.H.; Koontz, A.F.; McLeod, K.R.; Bush, L.P.; Schrick, F.N.; Harmon, D.L. Ergot alkaloids from endophyte-infected tall fescue decrease reticuloruminal epithelial blood flow and volatile fatty acid absorption from the washed reticulorumen. J. Anim. Sci. 2013, 91, 5366–5378. [Google Scholar] [CrossRef]
- Aldrich, C.; Paterson, J.; Tate, J.; Kerley, M. The effects of endophyte-infected tall fescue consumption on diet utilization and thermal regulation in cattle. J. Anim. Sci. 1993, 71, 164–170. [Google Scholar] [CrossRef]
- Seman, D.H.; Stuedemann, J.A.; Anderson, J.E. Spectral analysis of bovine grazing behavior on Neotyphodium coenophialum infested tall fescue. Appl. Anim. Behav. Sci. 1997, 54, 73–87. [Google Scholar] [CrossRef]
- Mote, R.S.; Hill, N.S.; Uppal, K.; Tran, V.T.; Jones, D.P.; Filipov, N.M. Metabolomics of fescue toxicosis in grazing beef steers. Food Chem. Toxicol. 2017, 105, 285–299. [Google Scholar] [CrossRef] [PubMed]
- Mote, R.S.; Hill, N.S.; Skarlupka, J.H.; Tran, V.T.; Walker, D.I.; Turner, Z.B.; Sanders, Z.P.; Jones, D.P.; Suen, G.; Filipov, N.M. Toxic tall fescue grazing increases susceptibility of the Angus steer fecal microbiota and plasma/urine metabolome to environmental effects. Sci. Rep. 2020, 10, 2497. [Google Scholar] [CrossRef]
- Mote, R.S.; Hill, N.S.; Skarlupka, J.H.; Carpenter, J.M.; Lourenco, J.M.; Callaway, T.R.; Tran, V.T.; Liu, K.; Smith, M.R.; Jones, D.P. Integrative interactomics applied to bovine fescue toxicosis. Sci. Rep. 2022, 12, 4899. [Google Scholar] [CrossRef]
- Hungate, R.E. The Rumen and Its Microbes; Elsevier: Amsterdam, The Netherlands, 2013. [Google Scholar]
- Russell, J.B. Rumen Microbiology and Its Role in Ruminant Nutrition; Department of Microbiology, Cornell University: Ithaca, NY, USA, 2002. [Google Scholar]
- Chai, J.; Alrashedi, S.; Coffey, K.; Burke, J.M.; Feye, K.; Ricke, S.C.; Park, S.H.; Edwards, J.L.; Zhao, J. Endophyte-infected tall fescue affects rumen microbiota in grazing ewes at gestation and lactation. Front. Vet. Sci. 2020, 7, 544707. [Google Scholar] [CrossRef]
- Roberts, C.; Andrae, J. Tall fescue toxicosis and management. Crop Manag. 2004, 3, 1–18. [Google Scholar] [CrossRef]
- Bouton, J.H.; Latch, G.C.; Hill, N.S.; Hoveland, C.S.; McCann, M.A.; Watson, R.H.; Parish, J.A.; Hawkins, L.L.; Thompson, F.N. Reinfection of tall fescue cultivars with non-ergot alkaloid–producing endophytes. Agron. J. 2002, 94, 567–574. [Google Scholar]
- Parish, J.; McCann, M.; Watson, R.; Paiva, N.; Hoveland, C.; Parks, A.; Upchurch, B.; Hill, N.; Bouton, J. Use of nonergot alkaloid-producing endophytes for alleviating tall fescue toxicosis in stocker cattle. J. Anim. Sci. 2003, 81, 2856–2868. [Google Scholar] [CrossRef]
- Llada, I.M.; Lourenco, J.M.; Dycus, M.M.; Carpenter, J.M.; Suen, G.; Hill, N.S.; Filipov, N.M. Behavioral and Physiological Alterations in Angus Steers Grazing Endophyte-Infected Toxic Fescue during Late Fall. Toxins 2023, 15, 343. [Google Scholar] [CrossRef]
- Jenkins, T. Lipid metabolism in the rumen. J. Dairy Sci. 1993, 76, 3851–3863. [Google Scholar] [CrossRef]
- Bauman, D.; Perfield, J.; De Veth, M.; Lock, A. New perspectives on lipid digestion and metabolism in ruminants. In Proceedings of the Cornell Nutrition Conference, Rochester, NY, USA, 21–23 October 2003; pp. 175–189. [Google Scholar]
- McLean, K.J.; Baldwin, R.L.; Li, C.-j.; Klotz, J.L.; Edwards, J.L.; McLeod, K.R. Synthetic alkaloid treatment influences the intestinal epithelium and mesenteric adipose transcriptome in Holstein steers. Front. Vet. Sci. 2020, 7, 615. [Google Scholar] [CrossRef]
- Oliver, J.; Schultze, A.; Rohrbach, B.; Fribourg, H.; Ingle, T.; Waller, J. Alterations in hemograms and serum biochemical analytes of steers after prolonged consumption of endophyte-infected tall fescue. J. Anim. Sci. 2000, 78, 1029–1035. [Google Scholar] [CrossRef]
- King, M.E.; Herzing, H.M.; McLeod, K.R.; Klotz, J.L.; Foote, A.P.; Edwards, J.L.; Harmon, D.L. Impact of endophyte-infected tall fescue seed consumption on endocrine changes associated with intake regulation and post-absorptive metabolism in growing steers. Domest. Anim. Endocrinol. 2024, 89, 106873. [Google Scholar] [CrossRef]
- Lafontan, M.; Berlan, M. Fat cell adrenergic receptors and the control of white and brown fat cell function. J. Lipid Res. 1993, 34, 1057–1091. [Google Scholar] [CrossRef]
- Garenc, C.; Pérusse, L.; Chagnon, Y.C.; Rankinen, T.; Gagnon, J.; Borecki, I.B.; Leon, A.S.; Skinner, J.S.; Wilmore, J.H.; Rao, D.; et al. The alpha2-adrenergic receptor gene and body fat content and distribution: The HERITAGE Family Study. Mol. Med. 2002, 8, 88–94. [Google Scholar] [CrossRef]
- Oliver, J.W.; Strickland, J.R.; Waller, J.C.; Fribourg, H.A.; Linnabary, R.D.; Abney, L.K. Endophytic fungal toxin effect on adrenergic receptors in lateral saphenous veins (cranial branch) of cattle grazing tall fescue. J. Anim. Sci. 1998, 76, 2853–2856. [Google Scholar] [CrossRef]
- Roquebert, J.; Grenie, B. Alpha 2-adrenergic agonist and alpha 1-adrenergic antagonist activity of ergotamine and dihydroergotamine in rats. Arch. Int. Pharmacodyn. Thér. 1986, 284, 30–37. [Google Scholar]
- Roquebert, J.; Grenie, B. Heart rate lowering effects of dihydroergotamine in rats. Arch. Int. Pharmacodyn. Thér. 1987, 290, 25–35. [Google Scholar]
- Pothoven, M.A.; Beitz, D.C. Effect of adipose tissue site, animal weight, and long-term fasting on lipogenesis in the bovine. J. Nutr. 1973, 103, 468–475. [Google Scholar] [CrossRef]
- Leveille, G.A. In vivo fatty acid and cholesterol synthesis in fasted and fasted-refed chicks. J. Nutr. 1969, 98, 367–372. [Google Scholar] [CrossRef]
- Baker, N.; Garfinkel, A.S.; Schotz, M.C. Hepatic triglyceride secretion in relation to lipogenesis and free fatty acid mobilization in fasted and glucose-refed rats. J. Lipid Res. 1968, 9, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Aldrich, C.; Rhodes, M.; Miner, J.; Kerley, M.; Paterson, J. The effects of endophyte-infected tall fescue consumption and use of a dopamine antagonist on intake, digestibility, body temperature, and blood constituents in sheep. J. Anim. Sci. 1993, 71, 158–163. [Google Scholar] [CrossRef]
- Goetsch, A.; Jones, A.; Stokes, S.; Beers, K.; Piper, E. Intake, digestion, passage rate and serum prolactin in growing dairy steers fed endophyte-infected fescue with noninfected fescue, clover or wheat straw. J. Anim. Sci. 1987, 64, 1759–1768. [Google Scholar] [CrossRef]
- Harlow, B.; Goodman, J.; Lynn, B.; Flythe, M.; Ji, H.; Aiken, G. Ruminal tryptophan-utilizing bacteria degrade ergovaline from tall fescue seed extract. J. Anim. Sci. 2017, 95, 980–988. [Google Scholar] [CrossRef]
- O’Hara, E.; Neves, A.L.; Song, Y.; Guan, L.L. The role of the gut microbiome in cattle production and health: Driver or passenger? Annu. Rev. Anim. Biosci. 2020, 8, 199–220. [Google Scholar] [CrossRef]
- Griswold, K.E.; White, B.A.; Mackie, R.I. Diversity of extracellular proteolytic activities among Prevotella species from the rumen. Curr. Microbiol. 1999, 39, 187–194. [Google Scholar] [CrossRef]
- Beauchemin, K.A.; Ungerfeld, E.M.; Eckard, R.J.; Wang, M. Review: Fifty years of research on rumen methanogenesis: Lessons learned and future challenges for mitigation. Animal 2020, 14, s2–s16. [Google Scholar] [CrossRef]
- Pavao-Zuckerman, M.A.; Waller, J.C.; Ingle, T.; Fribourg, H.A. Methane Emissions of Beef Cattle Grazing Tall Fescue Pastures at Three Levels of Endophyte Infestation; 0047-2425; Wiley Online Library: New York, NY, USA, 1999. [Google Scholar]
- Llada, I.; Mote, R.; Hill, N.; Lourenco, J.; Jones, D.; Suen, G.; Ross, M.; Filipov, N. Ruminal ergovaline and volatile fatty acid dynamics: Association with poor performance and a key growth regulator in steers grazing toxic tall fescue. Environ. Toxicol. Pharmacol. 2024, 105, 104354. [Google Scholar] [CrossRef]
- Poulsen, M.; Schwab, C.; Borg Jensen, B.; Engberg, R.M.; Spang, A.; Canibe, N.; Højberg, O.; Milinovich, G.; Fragner, L.; Schleper, C.; et al. Methylotrophic methanogenic Thermoplasmata implicated in reduced methane emissions from bovine rumen. Nat. Commun. 2013, 4, 1428. [Google Scholar] [CrossRef]
- Liu, Y.; Whitman, W.B. Metabolic, phylogenetic, and ecological diversity of the methanogenic archaea. Ann. N. Y. Acad. Sci. 2008, 1125, 171–189. [Google Scholar] [CrossRef]
- Krizsan, S.J.; Ramin, M.; Chagas, J.C.; Halmemies-Beauchet-Filleau, A.; Singh, A.; Schnürer, A.; Danielsson, R. Effects on rumen microbiome and milk quality of dairy cows fed a grass silage-based diet supplemented with the macroalga Asparagopsis taxiformis. Front. Anim. Sci. 2023, 4, 1112969. [Google Scholar] [CrossRef]
- Mote, R.S.; Hill, N.S.; Skarlupka, J.H.; Turner, Z.B.; Sanders, Z.P.; Jones, D.P.; Suen, G.; Filipov, N.M. Response of beef cattle fecal microbiota to grazing on toxic tall fescue. Appl. Environ. Microbiol. 2019, 85, e00032. [Google Scholar] [CrossRef]
- Knoll, E.; Wisser, H.; Emrich, H. 3, 4-Dimethoxyphenylethylamine excretion of normals and schizophrenics, behaviour during total fasting. Clin. Chim. Acta 1978, 89, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Goto, K.; Mochizuki, H.; Hattori, T.; Nakamura, N.; Mizuno, Y. Neurotoxic effects of papaverine, tetrahydropapaverine and dimethoxyphenylethylamine on dopaminergic neurons in ventral mesencephalic-striatal co-culture. Brain Res. 1997, 754, 260–268. [Google Scholar] [CrossRef]
- Naoi, M.; Maruyama, W.; Akao, Y.; Yi, H. Dopamine-derived endogenous N-methyl-(R)-salsolinol: Its role in Parkinson’s disease. Neurotoxicol. Teratol. 2002, 24, 579–591. [Google Scholar] [CrossRef]
- Larson, B.; Harmon, D.; Piper, E.; Griffis, L.; Bush, L. Alkaloid binding and activation of D2 dopamine receptors in cell culture. J. Anim. Sci. 1999, 77, 942–947. [Google Scholar] [CrossRef]
- Dyer, D.C. Evidence that ergovaline acts on serotonin receptors. Life Sci. 1993, 53, PL223–PL228. [Google Scholar] [CrossRef]
- Lipham, L.; Thompson, F.; Stuedemann, J.; Sartin, J. Effects of metoclopramide on steers grazing endophyte-infected fescue. J. Anim. Sci. 1989, 67, 1090–1097. [Google Scholar] [CrossRef]
- Russell, J.; Wallace, R. Energy-yielding and energy-consuming reactions. In The Rumen Microbial Ecosystem; Springer: Berlin/Heidelberg, Germany, 1997; pp. 246–282. [Google Scholar]
- Voet, D.; Voet, J.G. Biochemistry; John Wiley & Sons: Hoboken, NJ, USA, 1995; pp. 180–198. [Google Scholar]
- Lourenço, M.; Ramos-Morales, E.; Wallace, R. The role of microbes in rumen lipolysis and biohydrogenation and their manipulation. Animal 2010, 4, 1008–1023. [Google Scholar] [CrossRef]
- Ricciotti, E.; FitzGerald, G.A. Prostaglandins and inflammation. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 986–1000. [Google Scholar] [CrossRef]
- Rouzer, C.A.; Marnett, L.J. Mechanism of free radical oxygenation of polyunsaturated fatty acids by cyclooxygenases. Chem. Rev. 2003, 103, 2239–2304. [Google Scholar] [CrossRef] [PubMed]
- Bhusari, S.; Liu, Z.; Hearne, L.B.; Spiers, D.E.; Lamberson, W.R.; Antoniou, E. Expression profiling of heat stress effects on mice fed ergot alkaloids. Toxicol. Sci. 2007, 95, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Kudo, I.; Murakami, M. Phospholipase A2 enzymes. Prostaglandins Other Lipid Mediat. 2002, 68, 3–58. [Google Scholar] [CrossRef]
- Gao, J.-M.; Wang, M.; Liu, L.-P.; Wei, G.-H.; Zhang, A.-L.; Draghici, C.; Konishi, Y. Ergosterol peroxides as phospholipase A2 inhibitors from the fungus Lactarius hatsudake. Phytomedicine 2007, 14, 821–824. [Google Scholar] [CrossRef]
- Parthasarathy, A.; Cross, P.J.; Dobson, R.C.; Adams, L.E.; Savka, M.A.; Hudson, A.O. A three-ring circus: Metabolism of the three proteogenic aromatic amino acids and their role in the health of plants and animals. Front. Mol. Biosci. 2018, 5, 29. [Google Scholar] [CrossRef]
- Kopin, I.J.; Fischer, J.E.; Musacchio, J.; Horst, W.D. Evidence for a false neurochemical transmitter as a mechanism for the hypotensive effect of monoamine oxidase inhibitors. Proc. Natl. Acad. Sci. USA 1964, 52, 716–721. [Google Scholar] [CrossRef]
- Broadley, K.J. The vascular effects of trace amines and amphetamines. Pharmacol. Ther. 2010, 125, 363–375. [Google Scholar] [CrossRef]
- Berry, M.D. Mammalian central nervous system trace amines. Pharmacologic amphetamines, physiologic neuromodulators. J. Neurochem. 2004, 90, 257–271. [Google Scholar] [CrossRef]
- Bunzow, J.R.; Sonders, M.S.; Arttamangkul, S.; Harrison, L.M.; Zhang, G.; Quigley, D.I.; Darland, T.; Suchland, K.L.; Pasumamula, S.; Kennedy, J.L.; et al. Amphetamine, 3,4-methylenedioxymethamphetamine, lysergic acid diethylamide, and metabolites of the catecholamine neurotransmitters are agonists of a rat trace amine receptor. Mol. Pharmacol. 2001, 60, 1181–1188. [Google Scholar] [CrossRef]
- Frewin, D.B.; Whelan, R.F. The mechanism of action of tyramine on the blood vessels of the forearm in man. Br. J. Pharmacol. Chemother. 1968, 33, 105. [Google Scholar] [CrossRef]
- Fehler, M.; Broadley, K.J.; Ford, W.R.; Kidd, E.J. Identification of trace-amine-associated receptors (TAAR) in the rat aorta and their role in vasoconstriction by β-phenylethylamine. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2010, 382, 385–398. [Google Scholar] [CrossRef] [PubMed]
- Herbert, A.A.; Kidd, E.J.; Broadley, K.J. Dietary trace amine-dependent vasoconstriction in porcine coronary artery. Br. J. Pharmacol. 2008, 155, 525–534. [Google Scholar] [CrossRef] [PubMed]
- Toda, N.; Hayashi, S.; Hattori, K. Analysis of the effect of tyramine and norepinephrine in isolated canine cerebral and mesenteric arteries. J. Pharmacol. Exp. Ther. 1978, 205, 382–391. [Google Scholar] [CrossRef]
- Muramatsu, I. The effect of reserpine on sympathetic, purinergic neurotransmission in the isolated mesenteric artery of the dog: A pharmacological study. Br. J. Pharmacol. 1987, 91, 467. [Google Scholar] [CrossRef]
- Koh, A.H.W.; Chess-Williams, R.; Lohning, A.E. Differential mechanisms of action of the trace amines octopamine, synephrine and tyramine on the porcine coronary and mesenteric artery. Sci. Rep. 2019, 9, 10925. [Google Scholar] [CrossRef]
- Elliott, J.; Berhane, Y.; Bailey, S.R. Effects of monoamines formed in the cecum of horses on equine digital blood vessels and platelets. Am. J. Vet. Res. 2003, 64, 1124–1131. [Google Scholar] [CrossRef]
- Pretorius, L.; Smith, C. Tyramine-induced gastrointestinal dysregulation is attenuated via estradiol associated mechanisms in a zebrafish larval model. Toxicol. Appl. Pharmacol. 2023, 461, 116399. [Google Scholar] [CrossRef]
- Bhattarai, Y.; Williams, B.B.; Battaglioli, E.J.; Whitaker, W.R.; Till, L.; Grover, M.; Linden, D.R.; Akiba, Y.; Kandimalla, K.K.; Zachos, N.C.; et al. Gut microbiota-produced tryptamine activates an epithelial G-protein-coupled receptor to increase colonic secretion. Cell Host Microbe 2018, 23, 775–785.e5. [Google Scholar] [CrossRef]
- Latch, G.C. Physiological interactions of endophytic fungi and their hosts. Biotic stress tolerance imparted to grasses by endophytes. Agric. Ecosyst. Environ. 1993, 44, 143–156. [Google Scholar] [CrossRef]
- Rowan, D.D. Lolitrems, peramine and paxilline: Mycotoxins of the ryegrass/endophyte interaction. Agric. Ecosyst. Environ. 1993, 44, 103–122. [Google Scholar] [CrossRef]
- Pownall, D.; Familton, A.; Field, R.; Fletcher, L.; Lane, G. The effect of peramine ingestation in pen-fed lambs. Proc. N. Z. Soc. Anim. Prod. 1995, 55, 186. [Google Scholar]
- Stumpf, D.A.; Parks, J.K. Urea cycle regulation: I. Coupling of ornithine metabolism to mitochondrial oxidative phosphorylation. Neurology 1980, 30, 178. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Black, S.M. Carnitine homeostasis, mitochondrial function and cardiovascular disease. Drug Discov. Today Dis. Mech. 2009, 6, e31–e39. [Google Scholar] [CrossRef]
- Hamiel, C.R.; Pinto, S.; Hau, A.; Wischmeyer, P.E. Glutamine enhances heat shock protein 70 expression via increased hexosamine biosynthetic pathway activity. Am. J. Physiol.-Cell Physiol. 2009, 297, C1509–C1519. [Google Scholar] [CrossRef]
- Zhang, Y.-H.; Yanase-Fujiwara, M.; Hosono, T.; Kanosue, K. Warm and cold signals from the preoptic area: Which contribute more to the control of shivering in rats? J. Physiol. 1995, 485, 195–202. [Google Scholar] [CrossRef]
- Liu, K.H.; Nellis, M.; Uppal, K.; Ma, C.; Tran, V.; Liang, Y.; Walker, D.I.; Jones, D.P. Reference standardization for quantification and harmonization of large-scale metabolomics. Anal. Chem. 2020, 92, 8836–8844. [Google Scholar] [CrossRef]
- Yu, T.; Park, Y.; Johnson, J.M.; Jones, D.P. apLCMS—Adaptive processing of high-resolution LC/MS data. Bioinformatics 2009, 25, 1930–1936. [Google Scholar] [CrossRef]
- Uppal, K.; Soltow, Q.A.; Strobel, F.H.; Pittard, W.S.; Gernert, K.M.; Yu, T.; Jones, D.P. xMSanalyzer: Automated pipeline for improved feature detection and downstream analysis of large-scale, non-targeted metabolomics data. BMC Bioinform. 2013, 14, 15. [Google Scholar] [CrossRef]
- Pang, Z.; Chong, J.; Zhou, G.; de Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.-É.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the gap between raw spectra and functional insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef]
- Chong, J.; Wishart, D.S.; Xia, J. Using MetaboAnalyst 4.0 for comprehensive and integrative metabolomics data analysis. Curr. Protoc. Bioinform. 2019, 68, e86. [Google Scholar] [CrossRef]
- Li, S.; Park, Y.; Duraisingham, S.; Strobel, F.H.; Khan, N.; Soltow, Q.A.; Jones, D.P.; Pulendran, B. Predicting network activity from high throughput metabolomics. PLoS Comput. Biol. 2013, 9, e1003123. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, D.M.; Weimer, P.J. Dominance of Prevotella and low abundance of classical ruminal bacterial species in the bovine rumen revealed by relative quantification real-time PCR. Appl. Microbiol. Biotechnol. 2007, 75, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Dill-McFarland, K.A.; Breaker, J.D.; Suen, G. Microbial succession in the gastrointestinal tract of dairy cows from 2 weeks to first lactation. Sci. Rep. 2017, 7, 40864. [Google Scholar] [CrossRef]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the MiSeq Illumina sequencing platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef]
- Kittelmann, S.; Seedorf, H.; Walters, W.A.; Clemente, J.C.; Knight, R.; Gordon, J.I.; Janssen, P.H. Simultaneous amplicon sequencing to explore co-occurrence patterns of bacterial, archaeal and eukaryotic microorganisms in rumen microbial communities. PLoS ONE 2013, 8, e47879. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]
- Vázquez-Baeza, Y.; Pirrung, M.; Gonzalez, A.; Knight, R. EMPeror: A tool for visualizing high-throughput microbial community data. Gigascience 2013, 2, 16. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef]
- Good, I.J. The population frequencies of species and the estimation of population parameters. Biometrika 1953, 40, 237–264. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RUMEN FLUID | ||||
|---|---|---|---|---|
| m/z | Retention Time (s) | Annotation | Adducts | Database |
| 206.1286 | 34 | Kynuramine | M + ACN + H | BMDB0012246 |
| 185.1285 | 43 | Methoxytyramine | M + NH4 | BMDB0012162 |
| 128.107 | 39 | Iso-Valeraldehyde | M + ACN + H | BMDB0063681 |
| 169.1335 | 42 | Methyltyramine | M + NH4 | BMDB0003633 |
| 183.1129 | 47 | Phenylalanine | M + NH4 | BMDB0000159 |
| 143.1180 | 69 | Isopropyl alcohol | M + 2ACN + H | HMDB00863 |
| 213.1597 | 36 | 14-Bipiperidine-1-carboxylic acid | M + H | HMDB60336 |
| 249.1346 | 35 | Histidylproline diketopiperazine | M + H | HMDB02053 |
| 155.1179 | 39 | Tyramine | M + NH4 | BMDB0000306 |
| 197.1285 | 40 | Salsolinol | M + NH4 | BMDB0096156 |
| 112.0757 | 38 | 3,4-Dimethoxyphenylethylamine (DMPEA) | M + ACN + H | BMDB0096131 |
| 313.1069 | 31 | Not identified | ||
| 251.0713 | 187 | 7-hydroxy-2-methylisoflavone | [M-H]- | HMDB33979 |
| 316.2572 | 224 | Monoacylglycerol (MG-0:015:00) | [M-H]- | HMDB11532 |
| 357.2015 | 251 | 5-HETE | [M + Cl]- | C04805 |
| URINE | ||||
| 155.1179 | 37 | Tyramine | M + NH4 | BMDB0000306 |
| 169.1336 | 35 | Methyltyramine | M + NH4 | BMDB0003633 |
| 171.1128 | 67 | Dopamine | M + NH4 | BMDB0000073 |
| 185.1285 | 89 | Methoxytyramine | M + NH4 | BMDB0012162 |
| 197.1285 | 40 | Salsolinol | M + NH4 | BMDB0096156 |
| 209.0688 | 62 | 1H-Indole-3-carboxaldehyde (metabolite of L-tryptophan) | M + ACN + Na | BMDB0063668 and LMDB00661 |
| 213.1233 | 64 | Acetyldopamine | M + NH4 | BMDB0096146 |
| 268.1764 | 41 | Histidylleucine | M + NH4 | BMDB0063926 |
| 112.0757 | 42 | 3,4-Dimethoxyphenylethylamine (DMPEA) | M + ACN + 2H | BMDB0096131 |
| 173.1172 | 66 | 4-Hydroperoxy-2-nonenal | M + H | HMDB6027 |
| 213.0114 | 144 | Not identified | ||
| 214.1265 | 64 | 7a,12a-Dihydroxy-3-oxo-4-cholenoic acid | M + H + Na | BMDB0000447 |
| 246.1813 | 80 | Lysyl-Valine | M + H | HMDB28964 |
| 284.1716 | 38 | Histidinyl-Lysine | M + H | HMDB28890 |
| Matrices | Family/Genera | Metabolic Feature m/z | rt (s) | Annotation | Phys. Param | Kendall’s Rank | p-Value |
|---|---|---|---|---|---|---|---|
| Rumen fluid | Prevotellaceae | 185.1285 (R) | 43 | Methoxytyramine | NO | 0.54 | <0.01 |
| 169.1335 (R) | 42 | Methyltyramine | 0.45 | <0.01 | |||
| 183.1129 (R) | 47 | Phenylalanine | 0.52 | <0.01 | |||
| 197.1285 (R) | 40 | Salsolinol | 0.54 | <0.01 | |||
| Methanobrevibacter | 185.1285 (R) | 43 | Methoxytyramine | NO | −0.38 | 0.02 | |
| Methanomethylophilaceae_uncultured | 185.1285 (R) | 43 | Methoxytyramine | NO | 0.41 | 0.02 | |
| 155.1179 (R) | 39 | Tyramine | RT | −0.41 | 0.003 | ||
| 316.2572 (R) | 224 | Monoacylglycerol | RT | 0.39 | 0.004 | ||
| Rumen solid | Eubacterium_ coprostanoligenes_group | 206.1286 (R) | 34 | Kynuramine | NO | −0.38 | 0.003 |
| Feces | Rikenellaceae_RC9_gut_ group | 155.1179 (U) | 37 | Tyramine | NO | −0.32 | 0.04 |
| Clostridium_sensu_ stricto_1 | 112.0757 (U) | 42 | 3,4-Dimethoxyphenylethylamine | NO | −0.41 | 0.003 | |
| 155.1179 (U) | 37 | Tyramine | −0.39 | 0.003 | |||
| 169.1336 (U) | 35 | Methyltyramine | −0.40 | 0.002 | |||
| 185.1285 (U) | 89 | Methoxytyramine | −0.43 | 0.001 | |||
| 197.1285 (U) | 40 | Salsolinol | −0.40 | 0.006 | |||
| Urine | 112.0757 (U) | 42 | 3,4-Dimethoxyphenylethylamine | WG | −0.59 | 0.001 | |
| 197.1285 (U) | 40 | Salsolinol | WG | −0.45 | 0.009 | ||
| E− vs. E+ | E− vs. NT | E+ vs. NT | |
|---|---|---|---|
| √ | X | √ | E+ Toxin(s) effect |
| X | X | √ | |
| √ | X | X | |
| √ | √ | X | Endophyte effect |
| X | √ | X |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Llada, I.M.; Lourenco, J.M.; Dycus, M.M.; Carpenter, J.M.; Jarrell, Z.R.; Jones, D.P.; Suen, G.; Hill, N.S.; Filipov, N.M. Impact of Low-Level Ergot Alkaloids and Endophyte Presence in Tall Fescue Grass on the Metabolome and Microbiome of Fall-Grazing Steers. Toxins 2025, 17, 251. https://doi.org/10.3390/toxins17050251
Llada IM, Lourenco JM, Dycus MM, Carpenter JM, Jarrell ZR, Jones DP, Suen G, Hill NS, Filipov NM. Impact of Low-Level Ergot Alkaloids and Endophyte Presence in Tall Fescue Grass on the Metabolome and Microbiome of Fall-Grazing Steers. Toxins. 2025; 17(5):251. https://doi.org/10.3390/toxins17050251
Chicago/Turabian StyleLlada, Ignacio M., Jeferson M. Lourenco, Madison M. Dycus, Jessica M. Carpenter, Zachery R. Jarrell, Dean P. Jones, Garret Suen, Nicholas S. Hill, and Nikolay M. Filipov. 2025. "Impact of Low-Level Ergot Alkaloids and Endophyte Presence in Tall Fescue Grass on the Metabolome and Microbiome of Fall-Grazing Steers" Toxins 17, no. 5: 251. https://doi.org/10.3390/toxins17050251
APA StyleLlada, I. M., Lourenco, J. M., Dycus, M. M., Carpenter, J. M., Jarrell, Z. R., Jones, D. P., Suen, G., Hill, N. S., & Filipov, N. M. (2025). Impact of Low-Level Ergot Alkaloids and Endophyte Presence in Tall Fescue Grass on the Metabolome and Microbiome of Fall-Grazing Steers. Toxins, 17(5), 251. https://doi.org/10.3390/toxins17050251