Botulinum Toxin Type A Exerts Direct Trans-Synaptic Action at Bilateral Spinal Nociceptive Circuits

 , ,
, ,
Abstract
1. Introduction
2. Results
2.1. The Role of BoNT-A Transcytosis in the Lumbar Spinal Cord
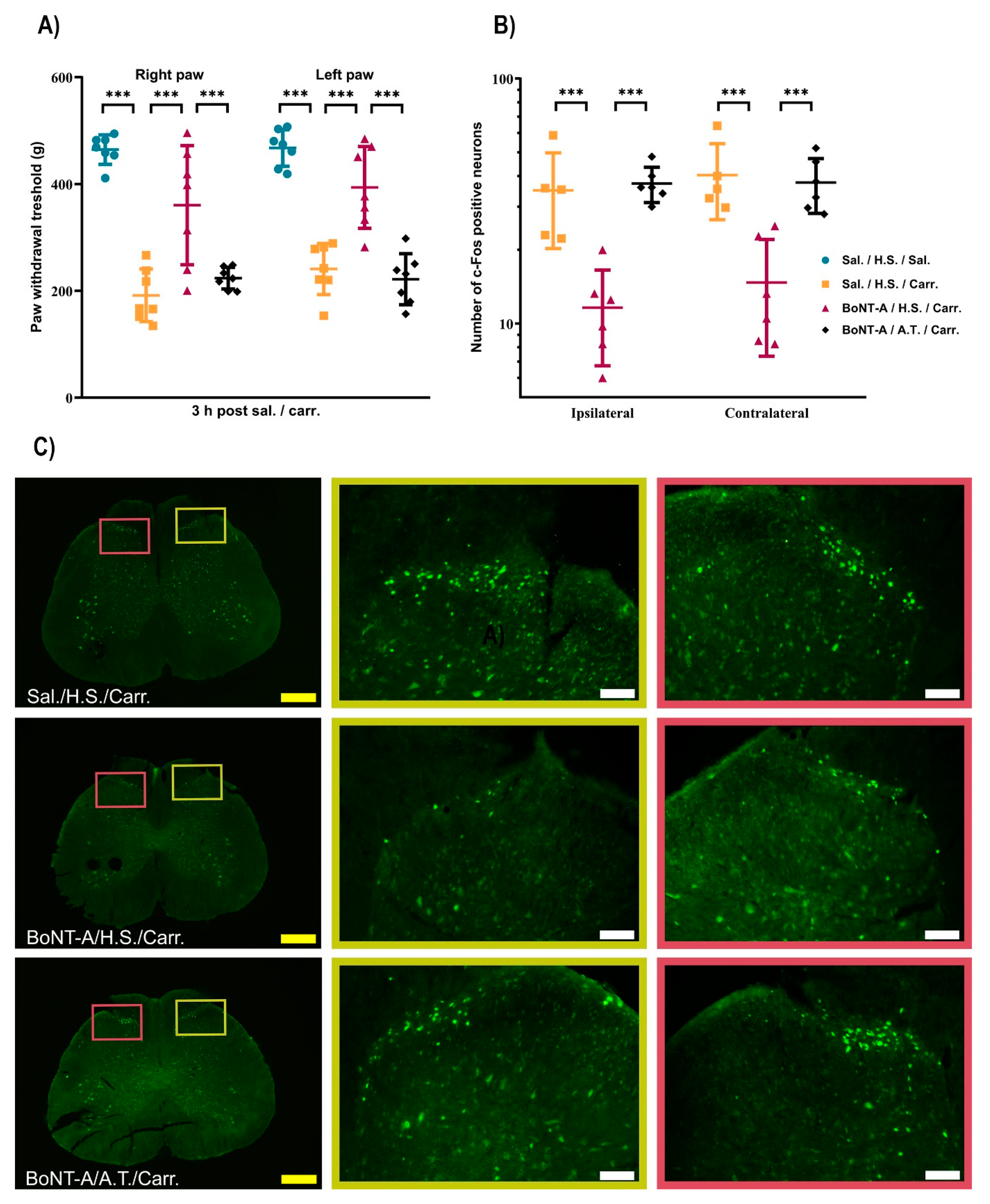
2.1.1. BoNT-A Reduces Bilateral Paw Pressure Nociception, Dependent on Its Transcytosis, with No Effect on Motor Performance
2.1.2. The BoNT-A Trans-Synaptic Effects Are Associated with the Reduced Nociceptive Neuronal Activation in the Bilateral Dorsal Horn
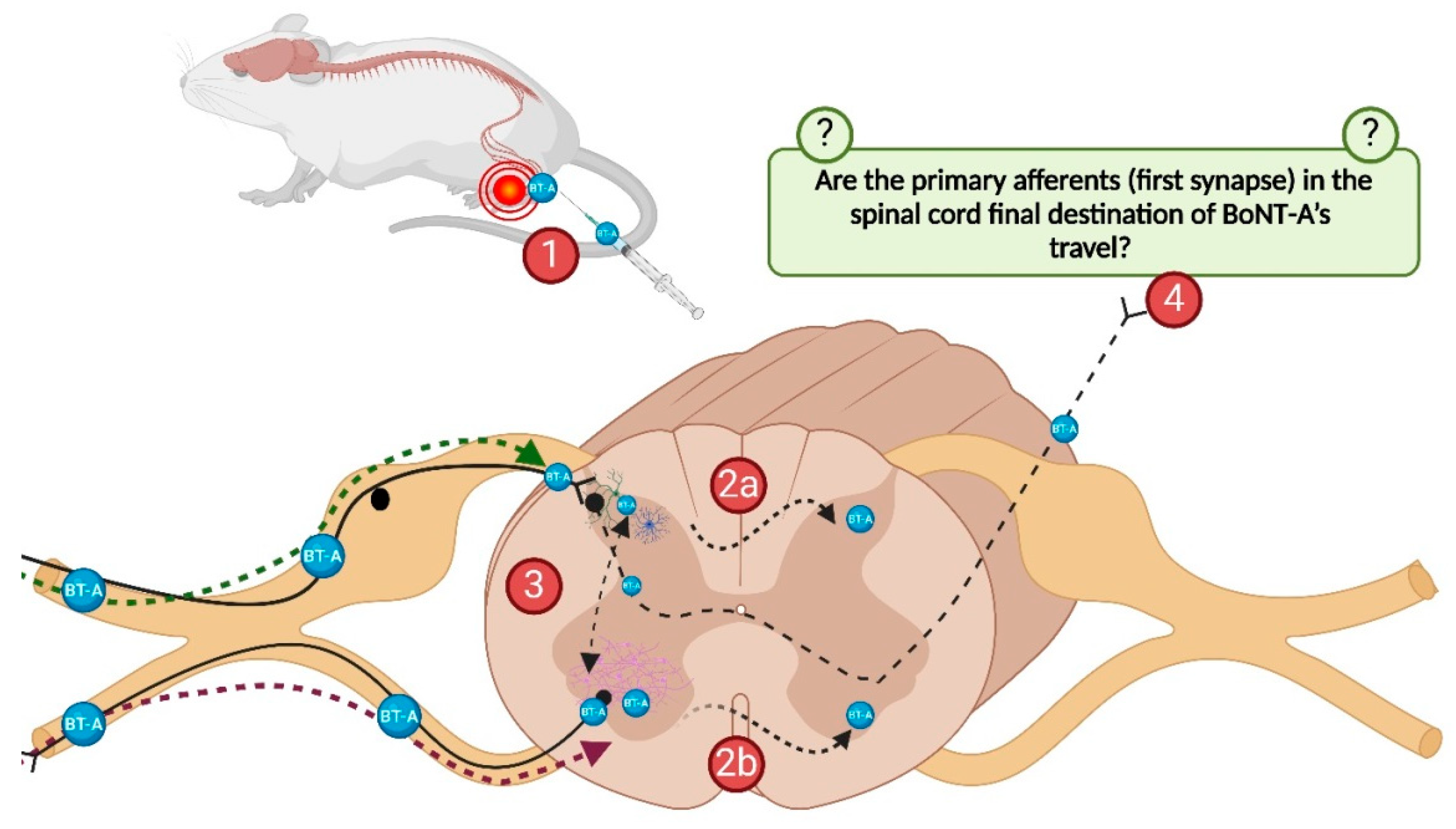
2.1.3. The BoNT-A Undergoes Trans-Synaptic Traffic in the Dorsal Horns Following Unilateral Hind-Paw Injection
2.1.4. The Preventive Effects of i.t.-Applied Antitoxin Are Not Due to Systemic Distribution After Lumbar i.t. Application
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Animals
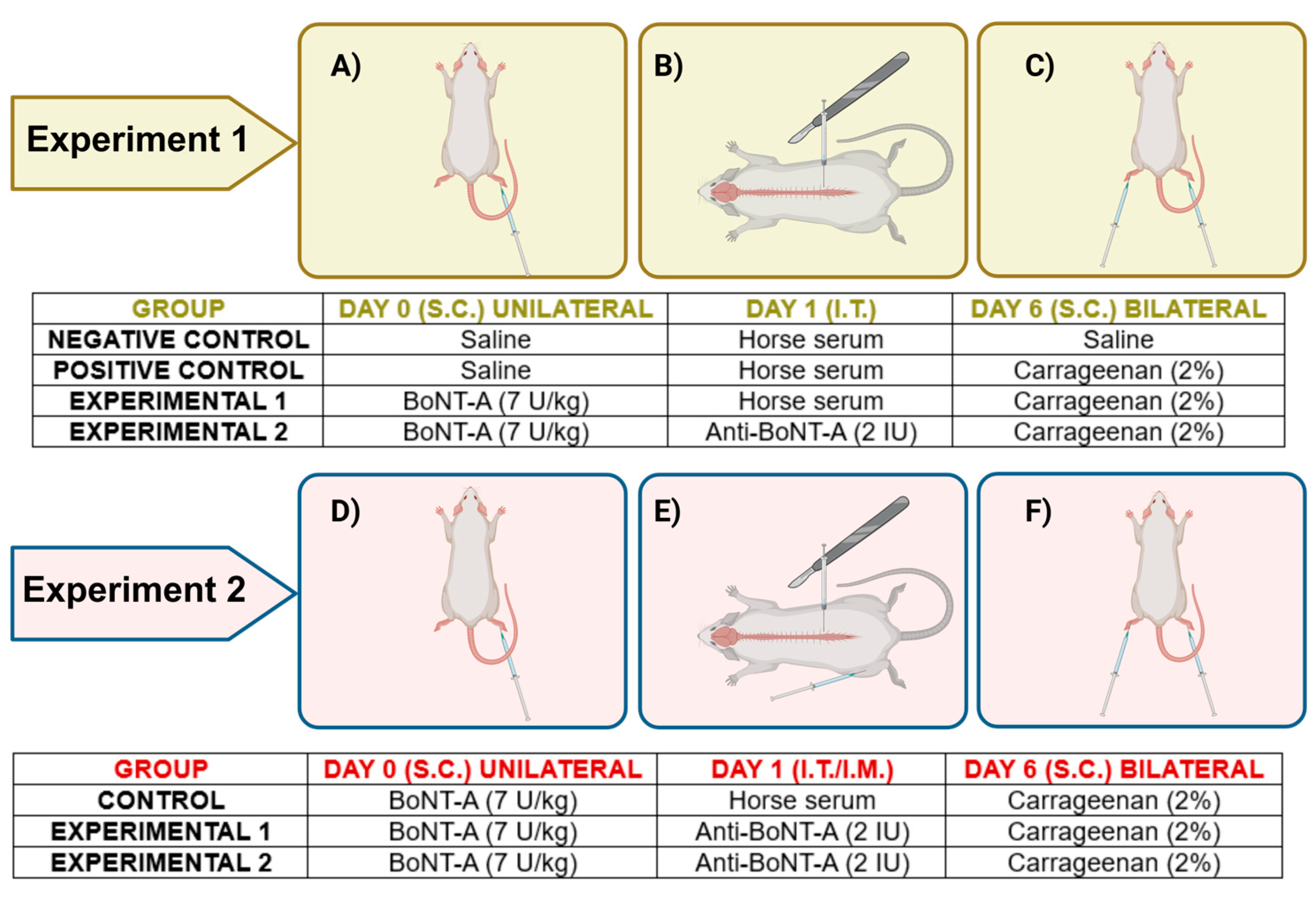
5.2. Drug Administration and Experimental Protocol
5.2.1. Intraplantar Injection
5.2.2. Intrathecal Injection
5.2.3. Intramuscular Injection
5.3. Carrageenan-Induced Inflammatory Pain Model
5.4. Behavioral Testing
5.4.1. Rota-Rod Test
5.4.2. Mechanical Hyperalgesia Measurement
5.5. Immunohistochemistry
5.5.1. C-Fos Immunohistochemistry
5.5.2. Cleaved SNAP-25 Immunohistochemistry
5.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pirazzini, M.; Montecucco, C.; Rossetto, O. Toxicology and pharmacology of botulinum and tetanus neurotoxins: An update. Arch. Toxicol. 2022, 96, 1521–1539. [Google Scholar] [CrossRef]
- Simpson, D.M.; Hallett, M.; Ashman, E.J.; Comella, C.L.; Green, M.W.; Gronseth, G.S.; Armstrong, M.J.; Gloss, D.; Potrebic, S.; Jankovic, J.; et al. Practice guideline update summary: Botulinum neurotoxin for the treatment of blepharospasm, cervical dystonia, adult spasticity, and headache: Report of the Guideline Development Subcommittee of the American Academy of Neurology. Neurology 2016, 86, 1818–1826. [Google Scholar] [CrossRef]
- Rempel, L.; Malik, R.N.; Shackleton, C.; Calderón-Juárez, M.; Sachdeva, R.; Krassioukov, A.V. From Toxin to Treatment: A Narrative Review on the Use of Botulinum Toxin for Autonomic Dysfunction. Toxins 2024, 16, 96. [Google Scholar] [CrossRef] [PubMed]
- Matak, I.; Bölcskei, K.; Bach-Rojecky, L.; Helyes, Z. Mechanisms of Botulinum Toxin Type A Action on Pain. Toxins 2019, 11, 459. [Google Scholar] [CrossRef] [PubMed]
- Pozo-Rosich, P.; Alpuente, A.; Silberstein, S.D.; Burstein, R. Insights from 25 years of onabotulinumtoxinA in migraine-mechanisms and management. Nature reviews. Neurology 2024, 20, 555–568. [Google Scholar] [CrossRef] [PubMed]
- Safarpour, Y.; Jabbari, B. Botulinum toxin treatment of pain syndromes—an evidence based review. Toxicon 2018, 147, 120–128. [Google Scholar] [CrossRef]
- Cui, M.; Khanijou, S.; Rubino, J.; Aoki, K.R. Subcutaneous administration of botulinum toxin A reduces formalin-induced pain. Pain 2004, 107, 125–133. [Google Scholar] [CrossRef]
- Vlah, V.D.; Bach-Rojecky, L. What have we learned about antinociceptive effect of botulinum toxin type A from mirror-image pain models? Toxicon 2020, 185, 164–173. [Google Scholar] [CrossRef]
- Nemanić, D.; Mustapić, M.; Matak, I.; Bach-Rojecky, L. Botulinum toxin type a antinociceptive activity in trigeminal regions involves central transcytosis. Eur. J. Pharmacol. 2024, 963, 176279. [Google Scholar] [CrossRef]
- Favre-Guilmard, C.; Auguet, M.; Chabrier, P.E. Different antinociceptive effects of botulinum toxin type A in inflammatory and peripheral polyneuropathic rat models. Eur. J. Pharmacol. 2009, 617, 48–53. [Google Scholar] [CrossRef]
- Luvisetto, S. Botulinum Neurotoxins in Central Nervous System: An Overview from Animal Models to Human Therapy. Toxins 2021, 13, 751. [Google Scholar] [CrossRef] [PubMed]
- Antonucci, F.; Rossi, C.; Gianfranceschi, L.; Rossetto, O.; Caleo, M. Long-distance retrograde effects of botulinum neurotoxin A. J. Neurosci. 2008, 28, 3689–3696. [Google Scholar] [CrossRef] [PubMed]
- Cai, B.B.; Francis, J.; Brin, M.F.; Broide, R.S. Botulinum neurotoxin type A-cleaved SNAP25 is confined to primary motor neurons and localized on the plasma membrane following intramuscular toxin injection. Neuroscience 2017, 352, 155–169. [Google Scholar] [CrossRef]
- Caleo, M.; Spinelli, M.; Colosimo, F.; Matak, I.; Rossetto, O.; Lackovic, Z.; Restani, L. Transynaptic Action of Botulinum Neurotoxin Type A at Central Cholinergic Boutons. J. Neurosci. 2018, 38, 10329–10337. [Google Scholar] [CrossRef]
- Matak, I. Evidence for central antispastic effect of botulinum toxin type A. Br. J. Pharmacol. 2020, 177, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Matak, I.; Bach-Rojecky, L.; Filipović, B.; Lacković, Z. Behavioral and immunohistochemical evidence for central antinociceptive activity of botulinum toxin A. Neuroscience 2011, 186, 201–207. [Google Scholar] [CrossRef]
- Marinelli, S.; Vacca, V.; Ricordy, R.; Uggenti, C.; Tata, A.M.; Luvisetto, S.; Pavone, F. The analgesic effect on neuropathic pain of retrogradely transported botulinum neurotoxin A involves Schwann cells and astrocytes. PLoS ONE 2012, 7, e47977. [Google Scholar] [CrossRef]
- Vlah, V.D.; Bach-Rojecky, L.; Lacković, Z. Antinociceptive action of botulinum toxin type A in carrageenan-induced mirror pain. J. Neural Transm. 2016, 123, 1403–1413. [Google Scholar] [CrossRef]
- Koizumi, H.; Goto, S.; Okita, S.; Morigaki, R.; Akaike, N.; Torii, Y.; Harakawa, T.; Ginnaga, A.; Kaji, R. Spinal Central Effects of Peripherally Applied Botulinum Neurotoxin A in Comparison between Its Subtypes A1 and A2. Front. Neurol. 2014, 5, 98. [Google Scholar] [CrossRef]
- Luz, L.L.; Lima, S.; Fernandes, E.C.; Kokai, E.; Gomori, L.; Szucs, P.; Safronov, B.V. Contralateral Afferent Input to Lumbar Lamina I Neurons as a Neural Substrate for Mirror-Image Pain. J. Neurosci. 2023, 43, 3245–3258. [Google Scholar] [CrossRef]
- Favre-Guilmard, C.; Chabrier, P.E.; Kalinichev, M. Bilateral analgesic effects of abobotulinumtoxinA (Dysport®) following unilateral administration in the rat. Eur. J. Pain 2017, 21, 927–937. [Google Scholar] [CrossRef] [PubMed]
- Matak, I.; Rossetto, O.; Lacković, Z. Botulinum toxin type A selectivity for certain types of pain is associated with capsaicin-sensitive neurons. Pain 2014, 155, 1516–1526. [Google Scholar] [CrossRef] [PubMed]
- Montecucco, C.; Schiavo, G.; Pantano, S. SNARE complexes and neuroexocytosis: How many, how close? Trends Biochem. Sci. 2005, 30, 367–372. [Google Scholar] [CrossRef]
- Oehler, B.; Périer, C.; Martin, V.; Fisher, A.; Lezmi, S.; Kalinichev, M.; McMahon, S.B. Evaluation of Recombinant Botulinum Neurotoxin Type A1 Efficacy in Peripheral Inflammatory Pain in Mice. Front. Mol. Neurosci. 2022, 15, 909835. [Google Scholar] [CrossRef] [PubMed]
- Bagues, A.; Hu, J.; Alshanqiti, I.; Chung, M.K. Neurobiological mechanisms of botulinum neurotoxin-induced analgesia for neuropathic pain. Pharmacol. Ther. 2024, 259, 108668. [Google Scholar] [CrossRef]
- Restani, L.; Novelli, E.; Bottari, D.; Leone, P.; Barone, I.; Galli-Resta, L.; Strettoi, E.; Caleo, M. Botulinum neurotoxin A impairs neurotransmission following retrograde transynaptic transport. Traffic 2012, 13, 1083–1089. [Google Scholar] [CrossRef]
- Bartels, F.; Bergel, H.; Bigalke, H.; Frevert, J.; Halpern, J.; Middlebrook, J. Specific antibodies against the Zn(2+)-binding domain of clostridial neurotoxins restore exocytosis in chromaffin cells treated with tetanus or botulinum A neurotoxin. J. Biol. Chem. 1994, 269, 8122–8127. [Google Scholar] [CrossRef] [PubMed]
- Silos-Santiago, I.; Snider, W.D. Development of commissural neurons in the embryonic rat spinal cord. J. Comp. Neurol. 1992, 325, 514–526. [Google Scholar] [CrossRef]
- du Sert, N.P.; Hurst, V.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; et al. The ARRIVE guidelines 2.0: Updated guidelines for reporting animal research. PLoS Biol. 2020, 18, e3000410. [Google Scholar] [CrossRef]
- Wu, C.; Xie, N.; Lian, Y.; Xu, H.; Chen, C.; Zheng, Y.; Chen, Y.; Zhang, H. Central antinociceptive activity of peripherally applied botulinum toxin type A in lab rat model of trigeminal neuralgia. SpringerPlus 2016, 5, 431. [Google Scholar] [CrossRef]
- Randall, L.O.; Selitto, J.J. A method for measurement of analgesic activity on inflamed tissue. Arch. Int. Pharmacodyn. Ther. 1957, 111, 409–419. [Google Scholar] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment/Test (Mean ± SEM) | Sal./H.S./Sal. | Sal./H.S./Carr. | BoNT-A/H.S./Carr. | BoNT-A/A.T./Carr. |
|---|---|---|---|---|
| Weight (g) pre-BoNT-A | 536.57 ± 15.6 | 536 ± 13.92 | 515.42 ± 13.35 | 523.71 ± 13.63 |
| Weight (g) 6 days post-BoNT-A | 528 ± 15.49 | 520 ± 17.59 | 498 ± 12.22 | 502 ± 15.69 |
| Rota-rod latency (s) pre-BoNT-A | 138.28 ± 14.18 | 118.14 ± 16.72 | 124.71 ± 17.73 | 125.71 ± 16.08 |
| Rota-rod latency (s) 6 days post-BoNT-A | 143 ± 10.64 | 109.14 ± 19.02 | 114.29 ± 19.41 | 120 ± 13.49 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nemanić, D.; Šoštarić, P.; Meglić, P.; Matak, I.; Bach-Rojecky, L. Botulinum Toxin Type A Exerts Direct Trans-Synaptic Action at Bilateral Spinal Nociceptive Circuits. Toxins 2025, 17, 140. https://doi.org/10.3390/toxins17030140
Nemanić D, Šoštarić P, Meglić P, Matak I, Bach-Rojecky L. Botulinum Toxin Type A Exerts Direct Trans-Synaptic Action at Bilateral Spinal Nociceptive Circuits. Toxins. 2025; 17(3):140. https://doi.org/10.3390/toxins17030140
Chicago/Turabian StyleNemanić, Dalia, Petra Šoštarić, Patrik Meglić, Ivica Matak, and Lidija Bach-Rojecky. 2025. "Botulinum Toxin Type A Exerts Direct Trans-Synaptic Action at Bilateral Spinal Nociceptive Circuits" Toxins 17, no. 3: 140. https://doi.org/10.3390/toxins17030140
APA StyleNemanić, D., Šoštarić, P., Meglić, P., Matak, I., & Bach-Rojecky, L. (2025). Botulinum Toxin Type A Exerts Direct Trans-Synaptic Action at Bilateral Spinal Nociceptive Circuits. Toxins, 17(3), 140. https://doi.org/10.3390/toxins17030140