The Role of Mycotoxins in Interactions between Fusarium graminearum and F. verticillioides Growing in Saprophytic Cultures and Co-Infecting Maize Plants



Abstract
:1. Introduction
1.1. Maize pathogens Fusarium graminearum and F. verticillioides
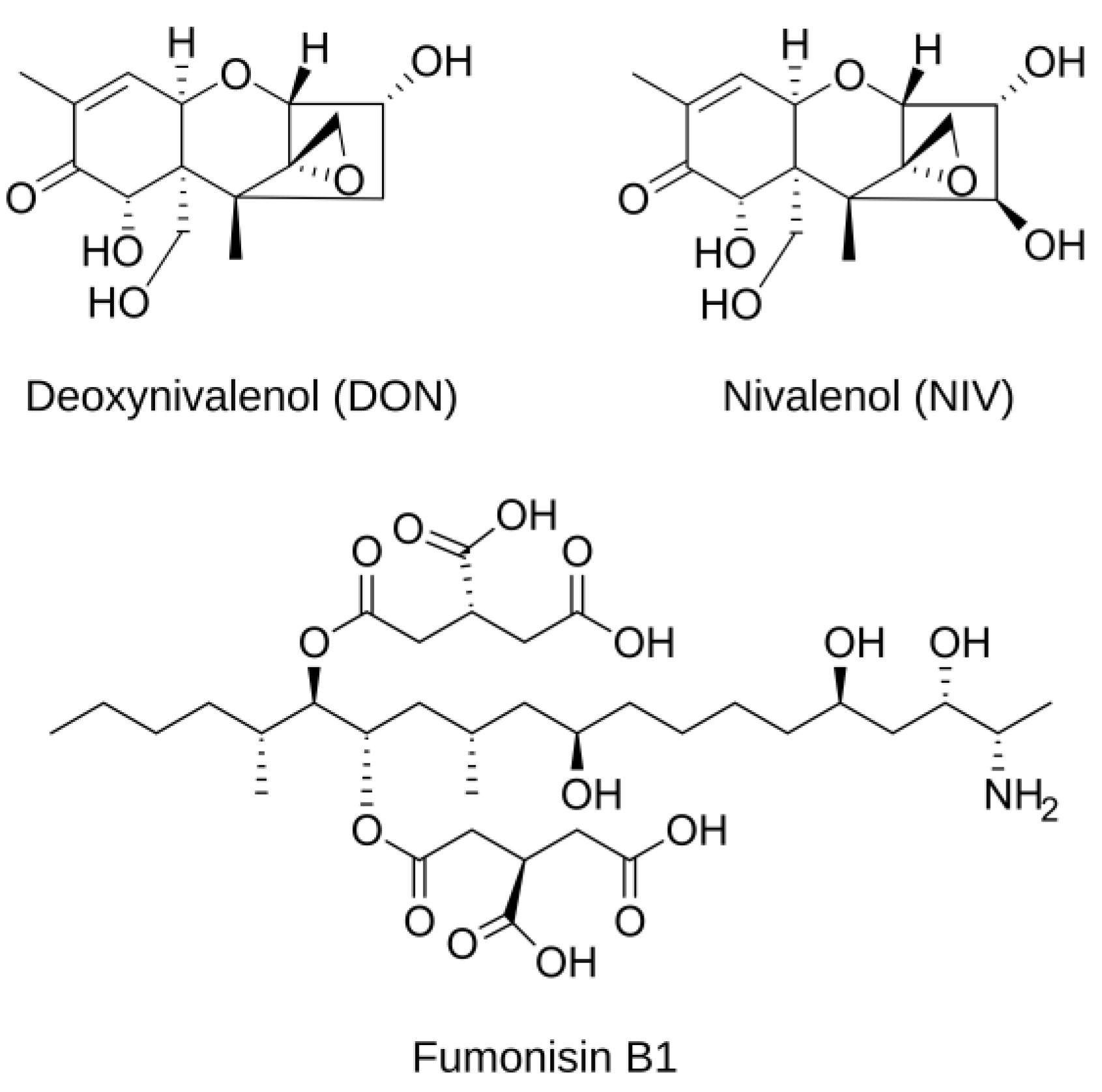
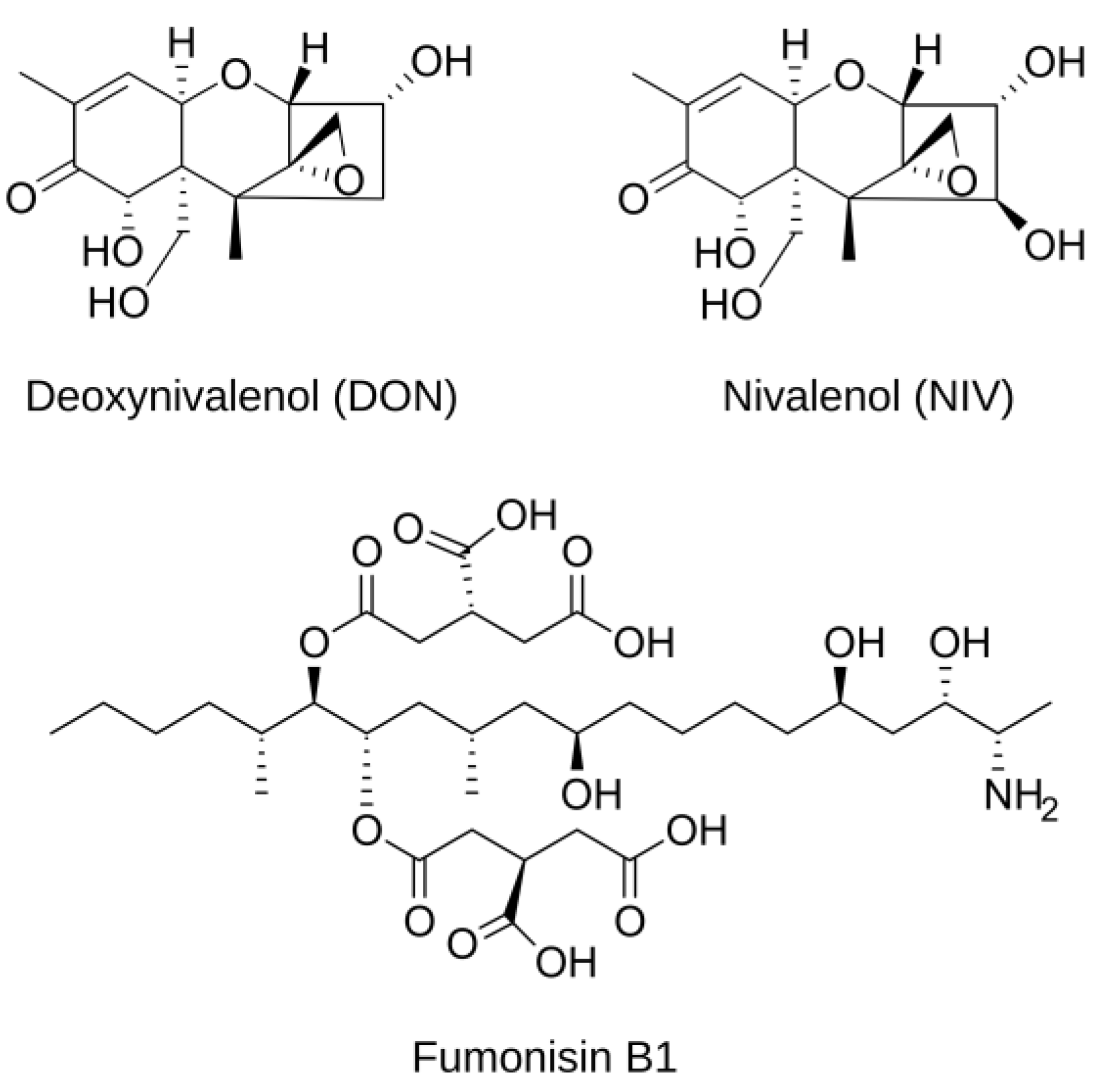
1.2. Trichothecenes of Fusarium graminearum and Fumonisins of F. verticillioides and Their Role in Plant Disease
1.3. Interaction between F. graminearum and F. verticillioides during Saprophytic Growth and the Role of Mycotoxins Produced by These Fungi
1.4. Mixed Infection of Maize Ears with F. graminearum and F. verticillioides
1.5. Objectives
2. Results
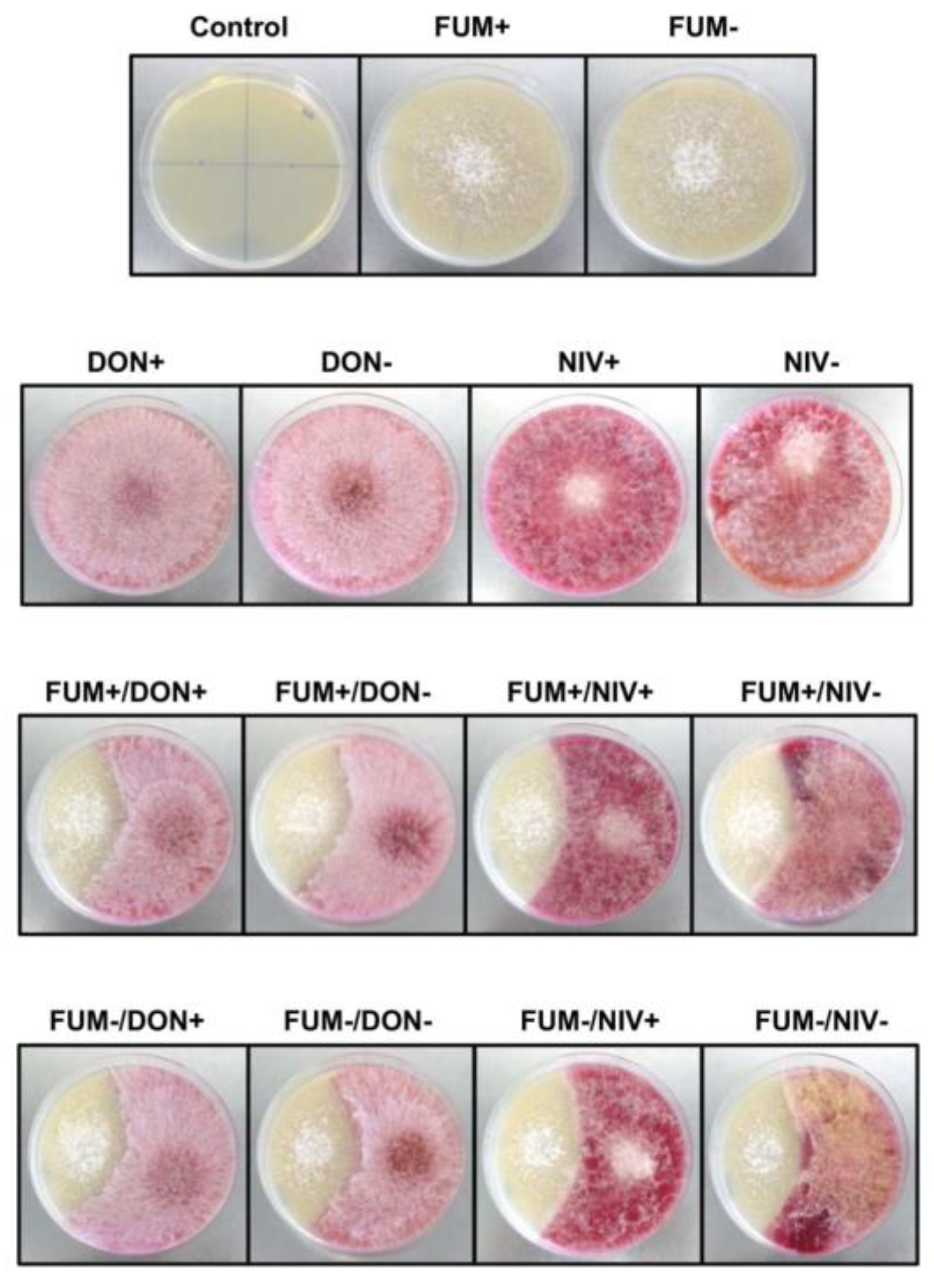
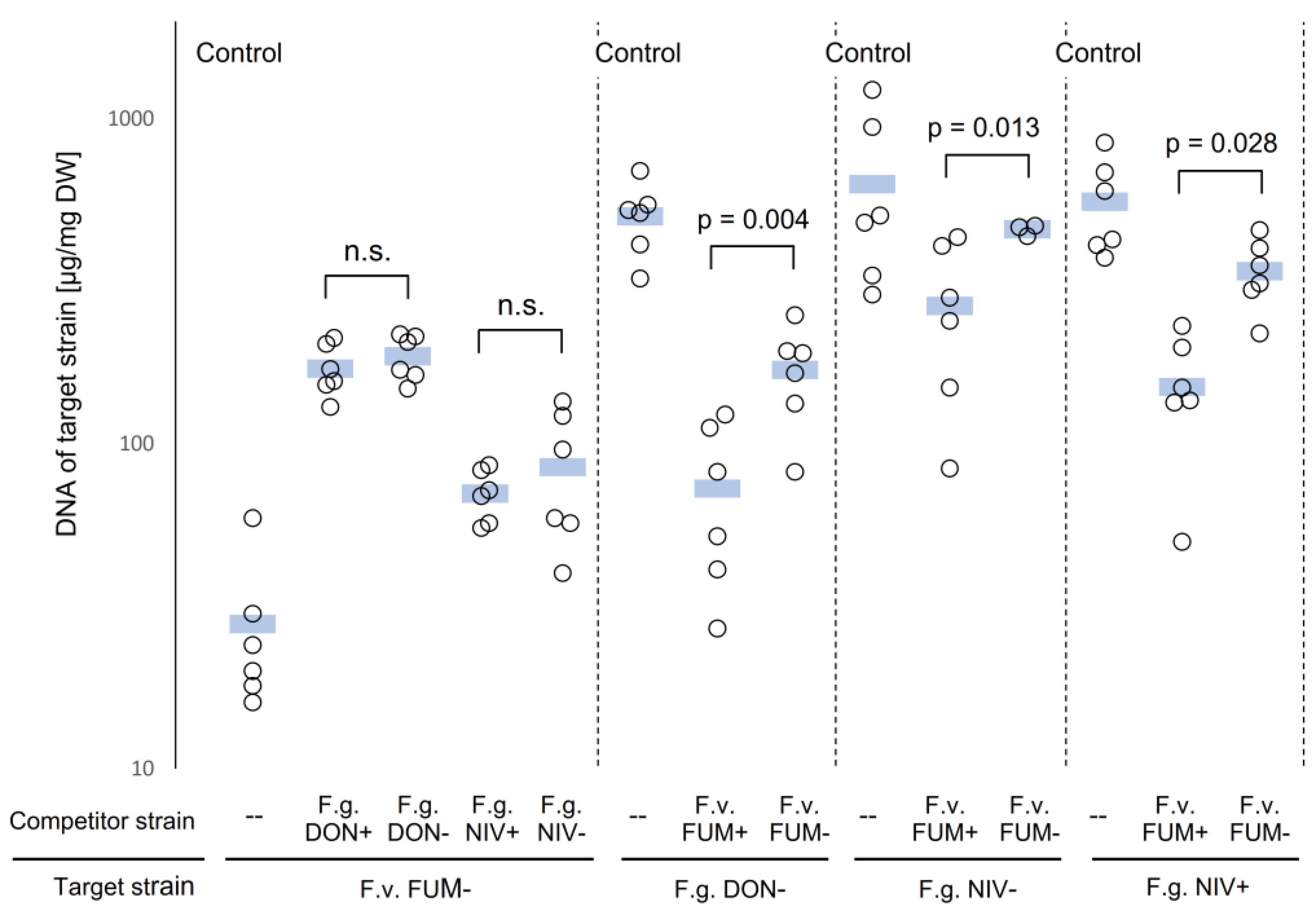
2.1. Competition between F. graminearum and F. verticillioides in Saprophytic Growth
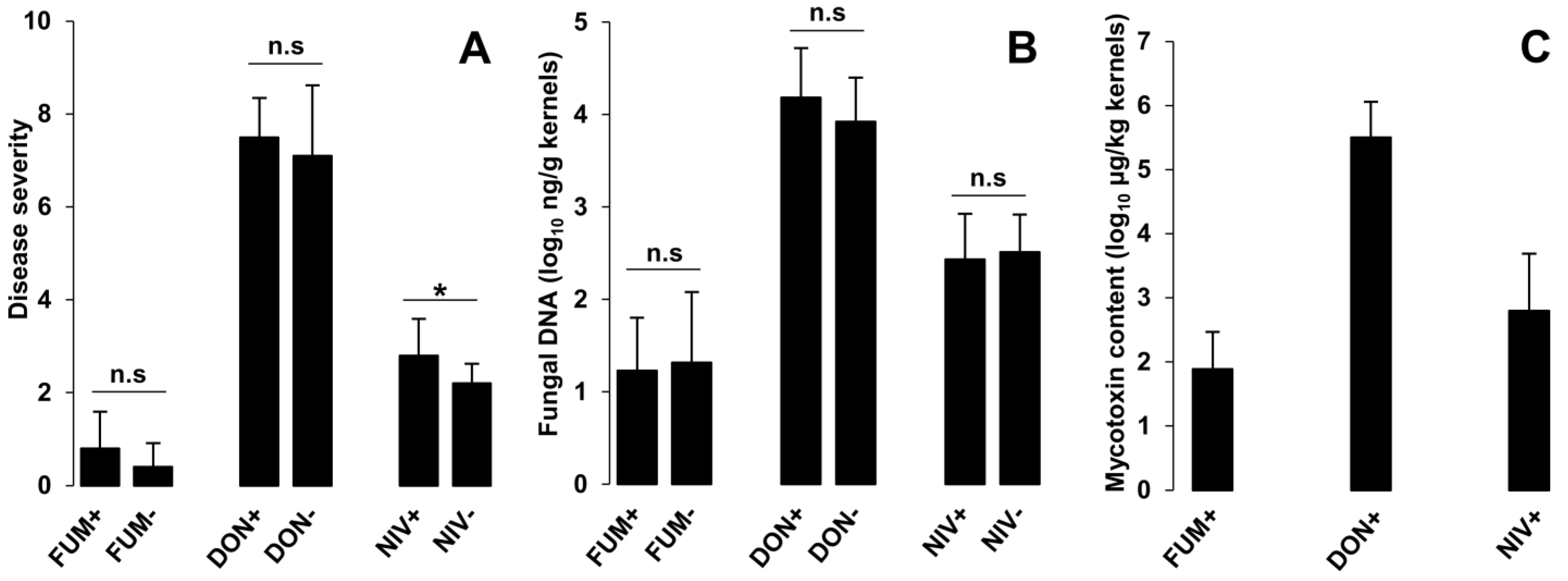
2.2. Disease Severity, Fungal Biomass, and Mycotoxin Accumulation in Maize Ears Inoculated with Single Strains of F. graminearum or F. verticillioides
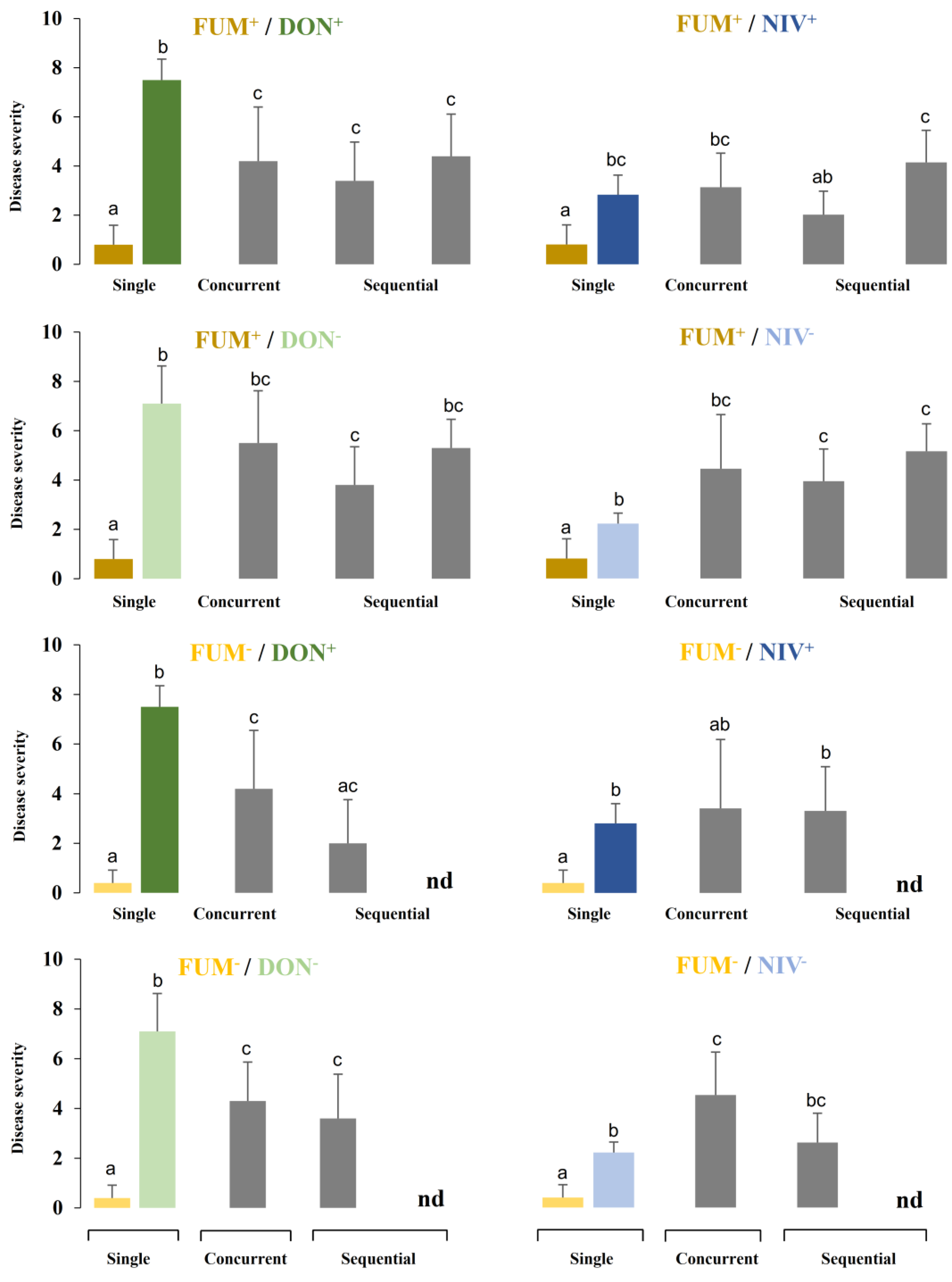
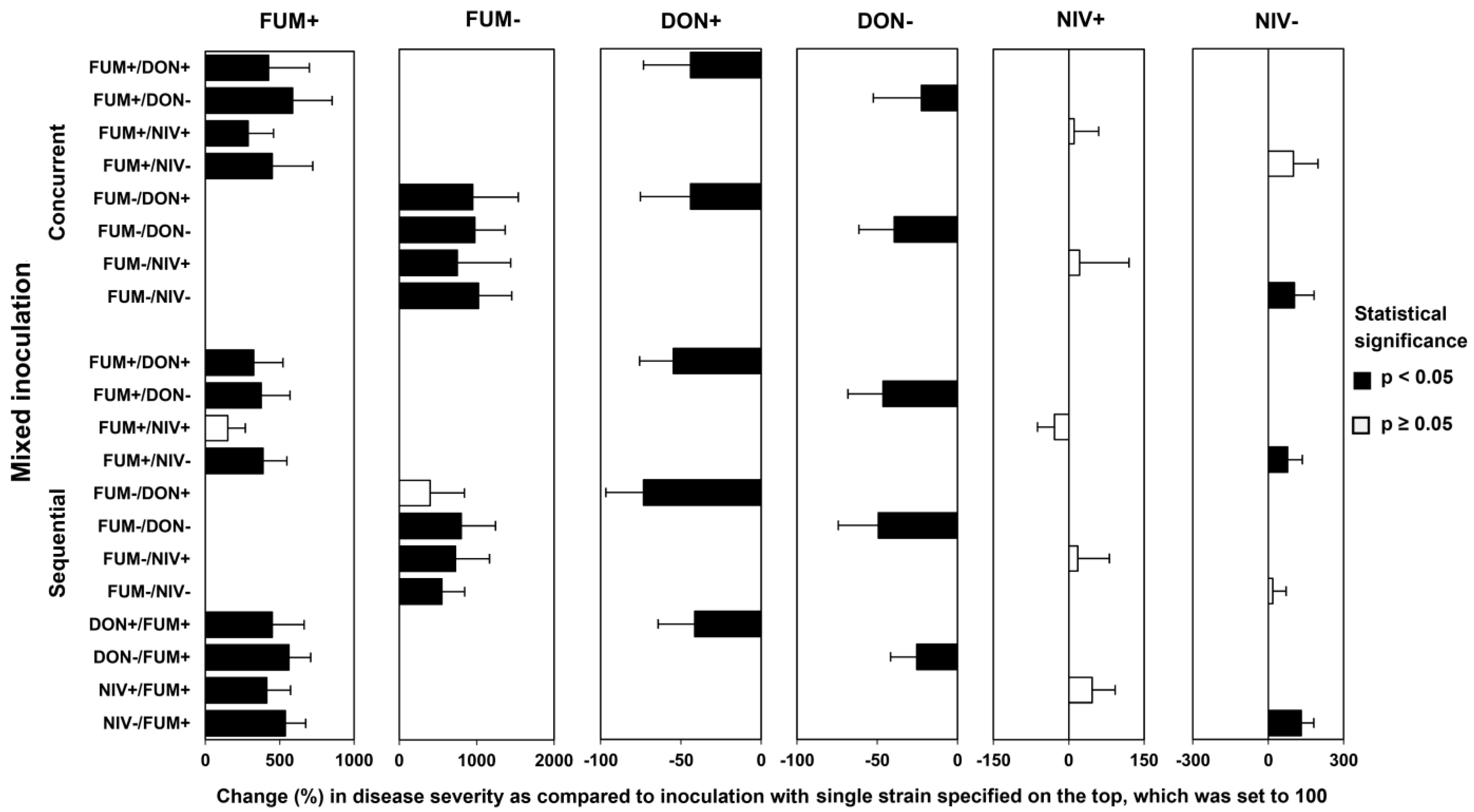
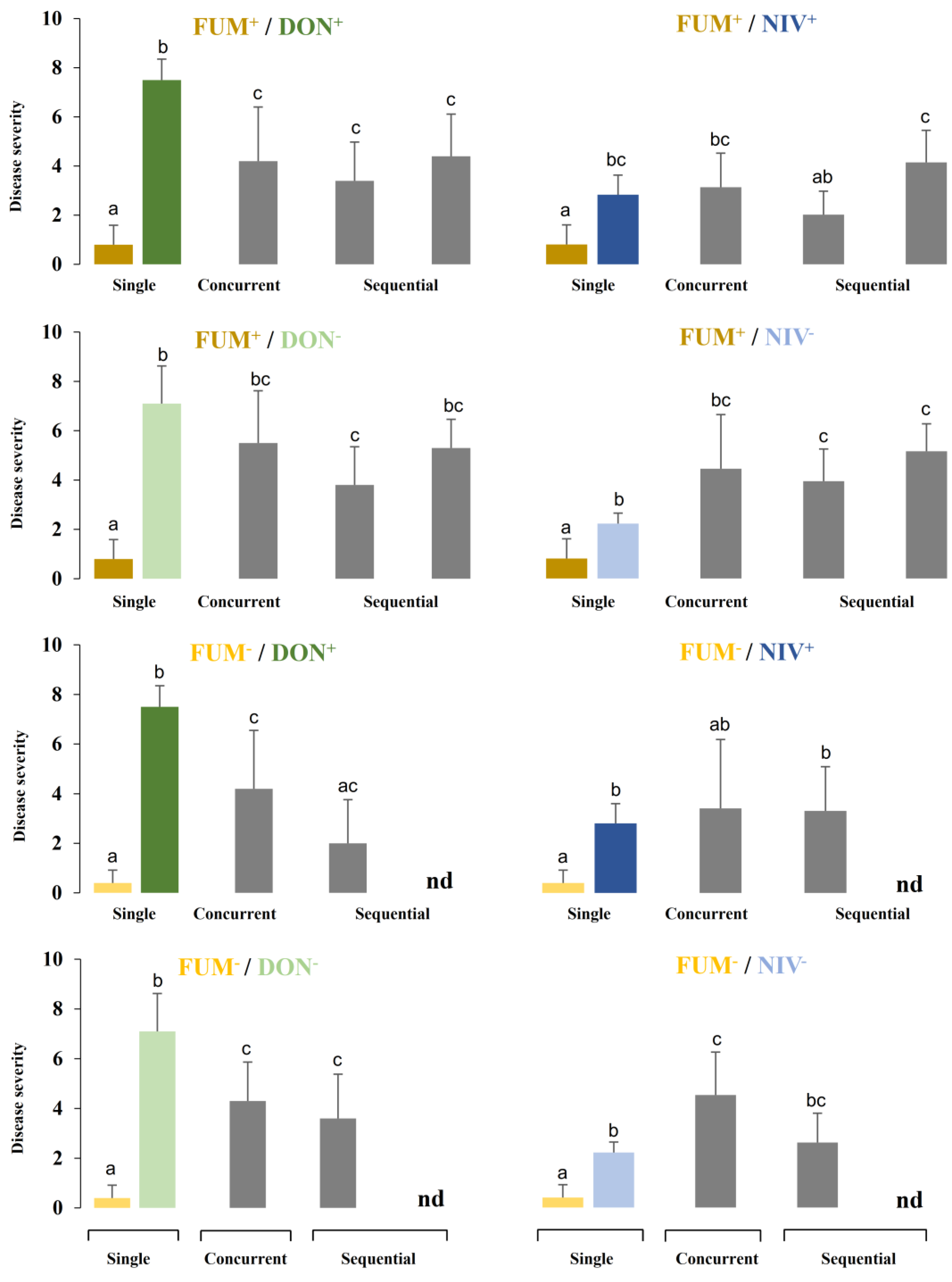
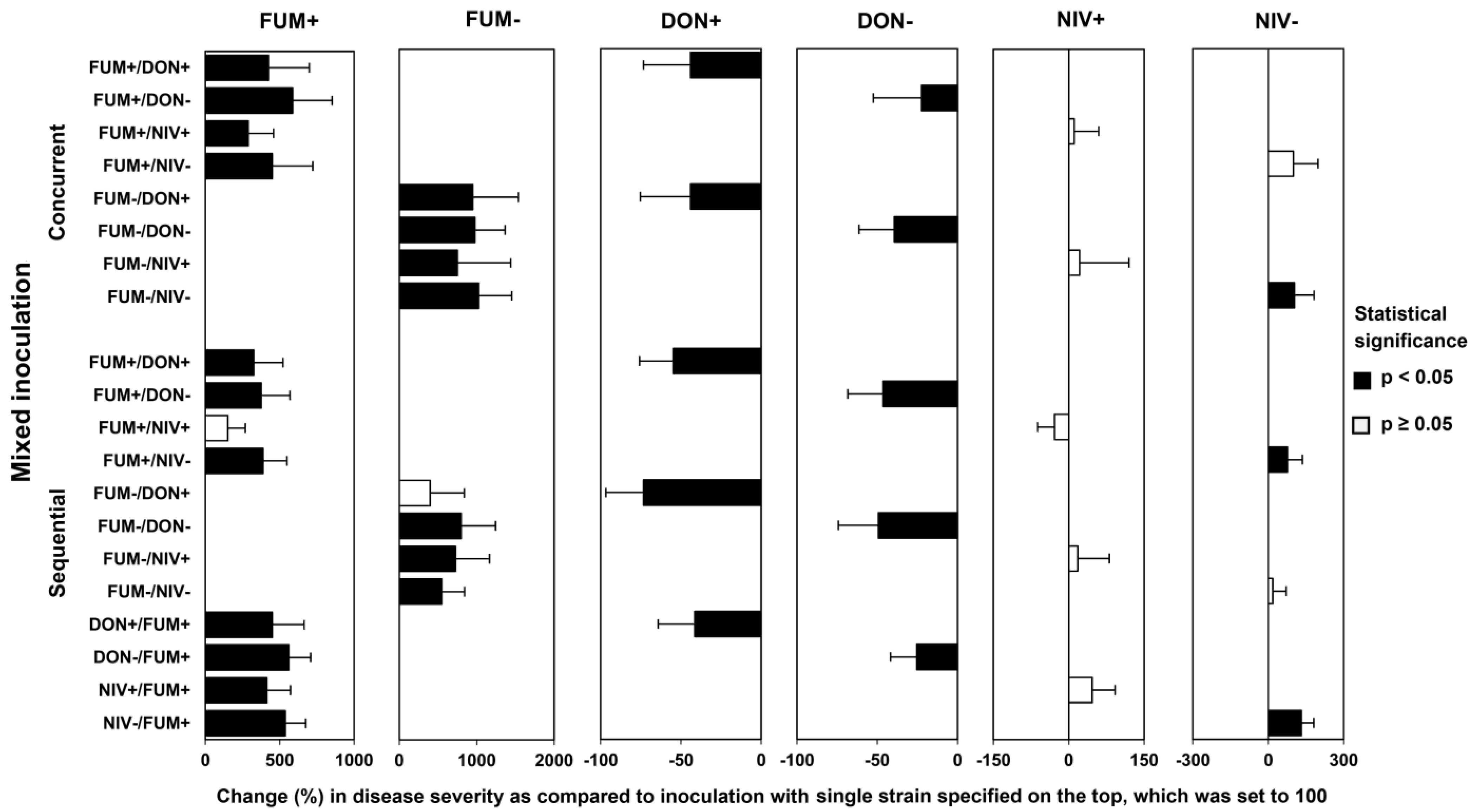
2.3. Disease Development after Co-Inoculation of Maize Ears with F. graminearum and F. verticillioides
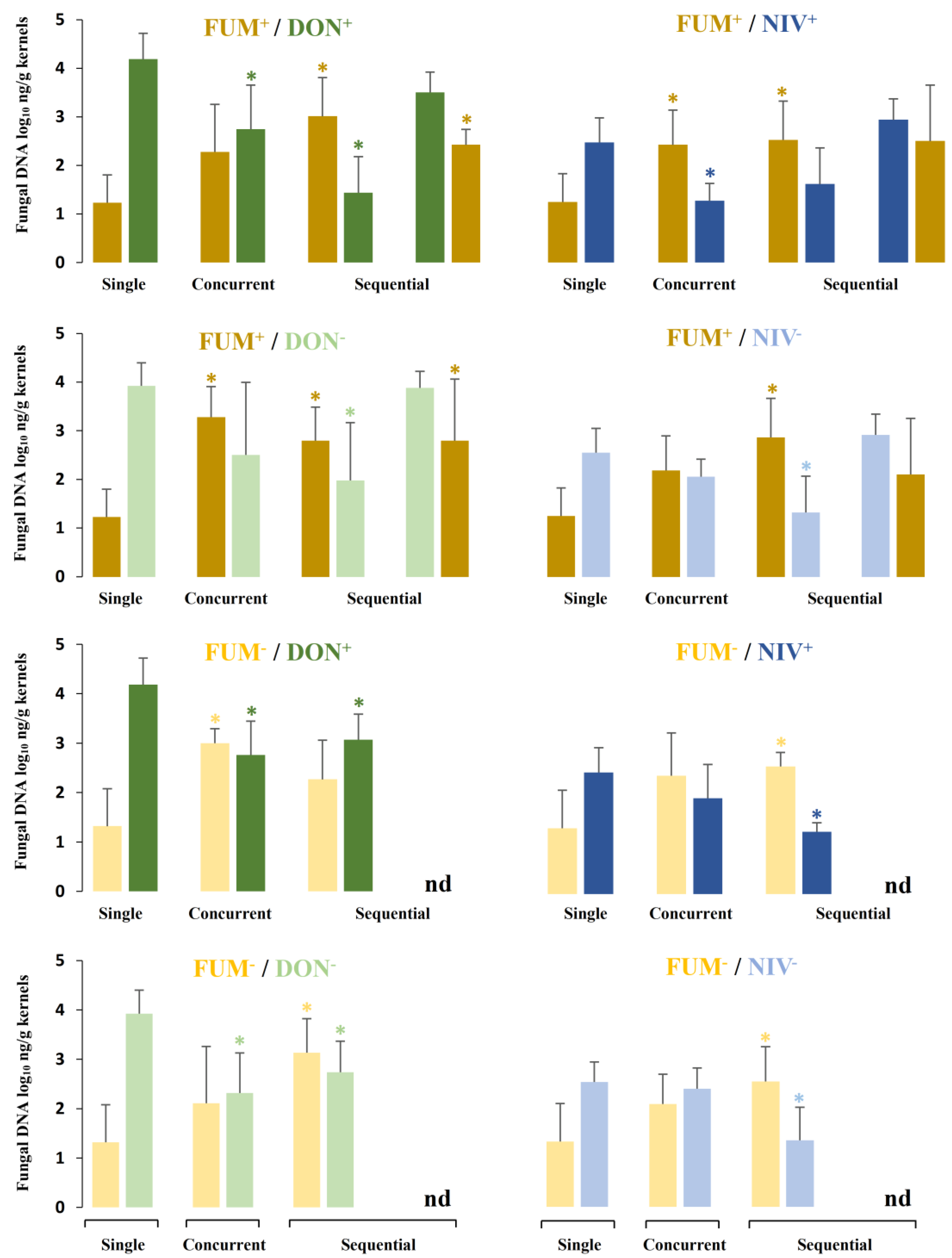
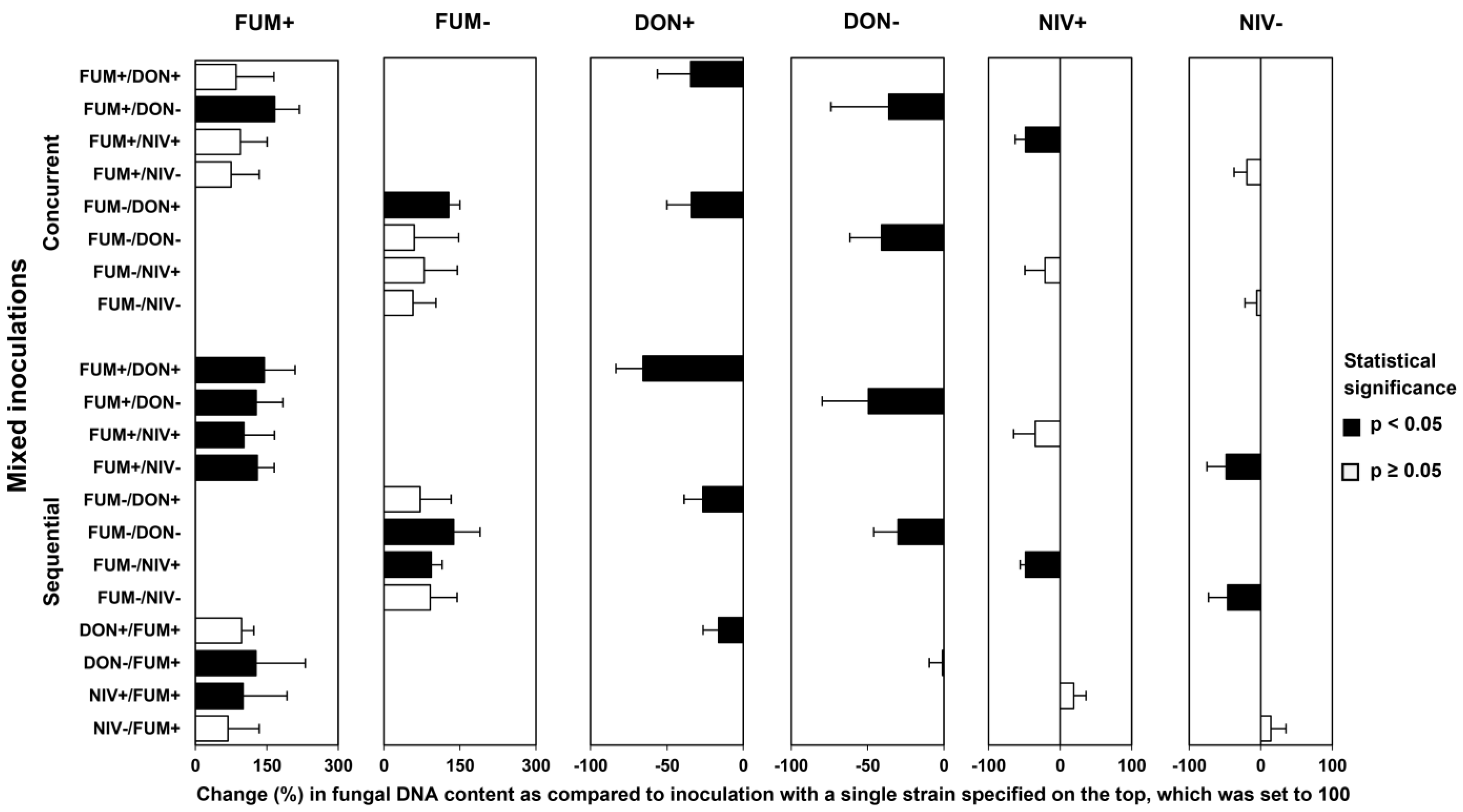
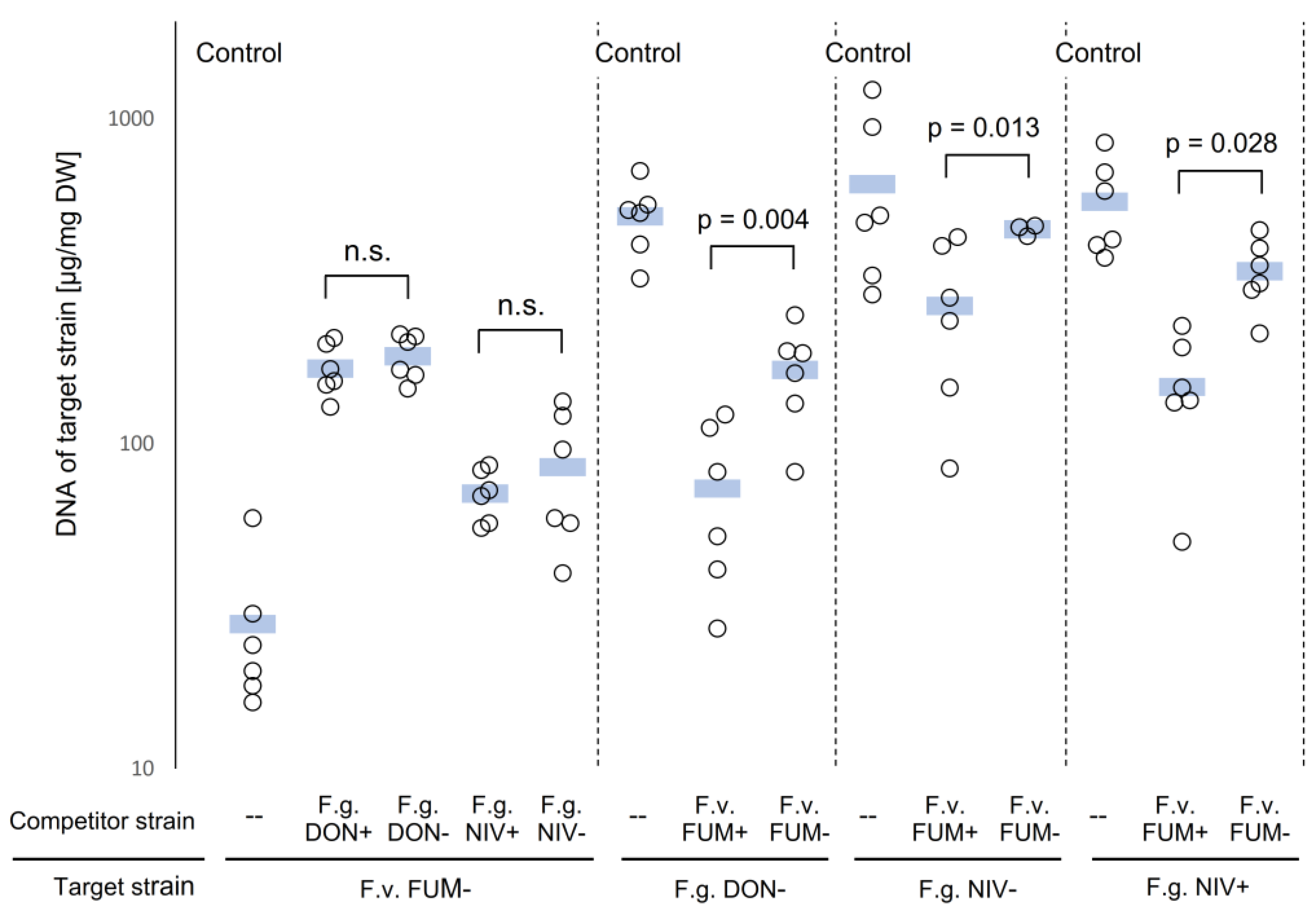
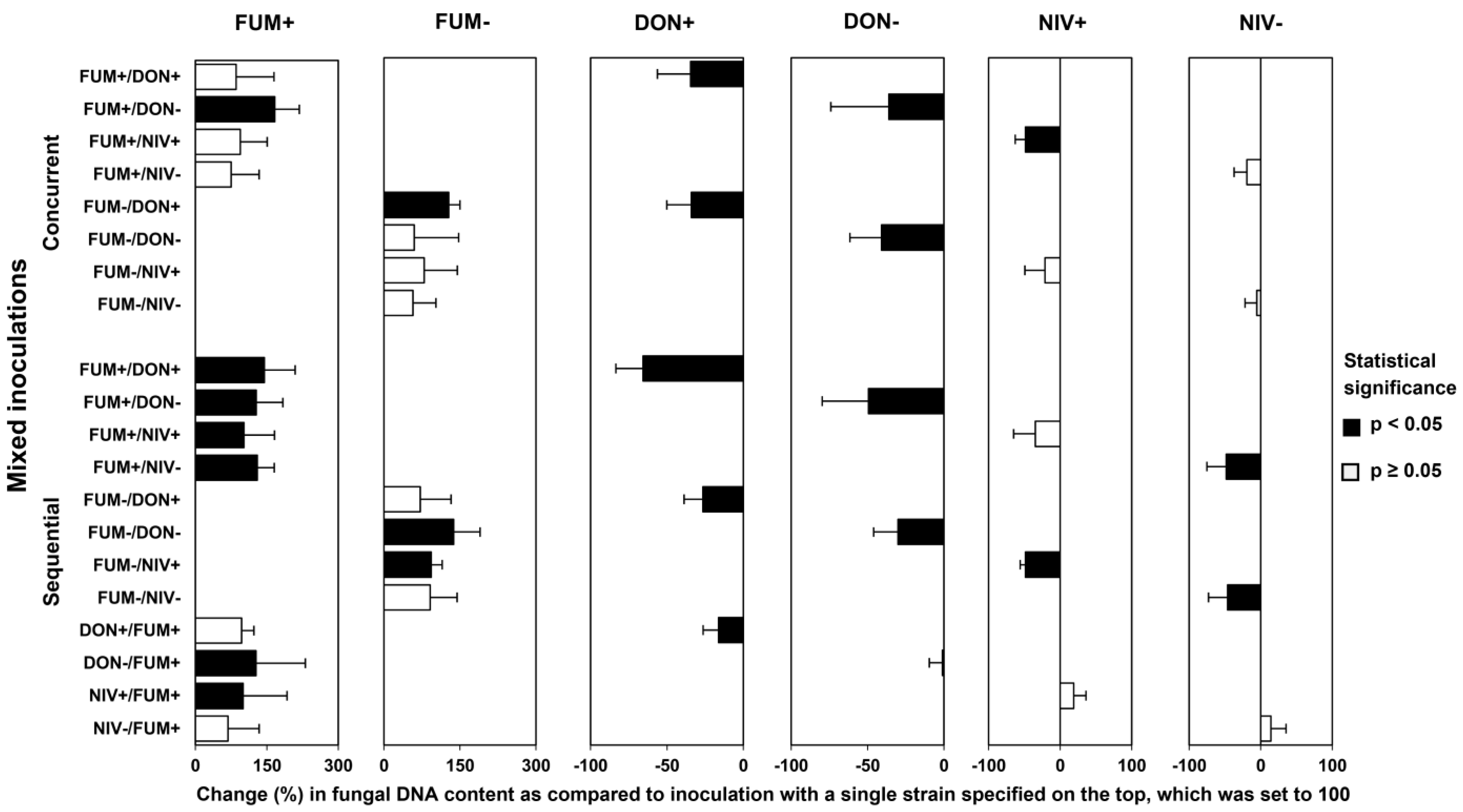
2.4. Fungal Biomass in Plants Co-Inoculated with Both Fusarium Species
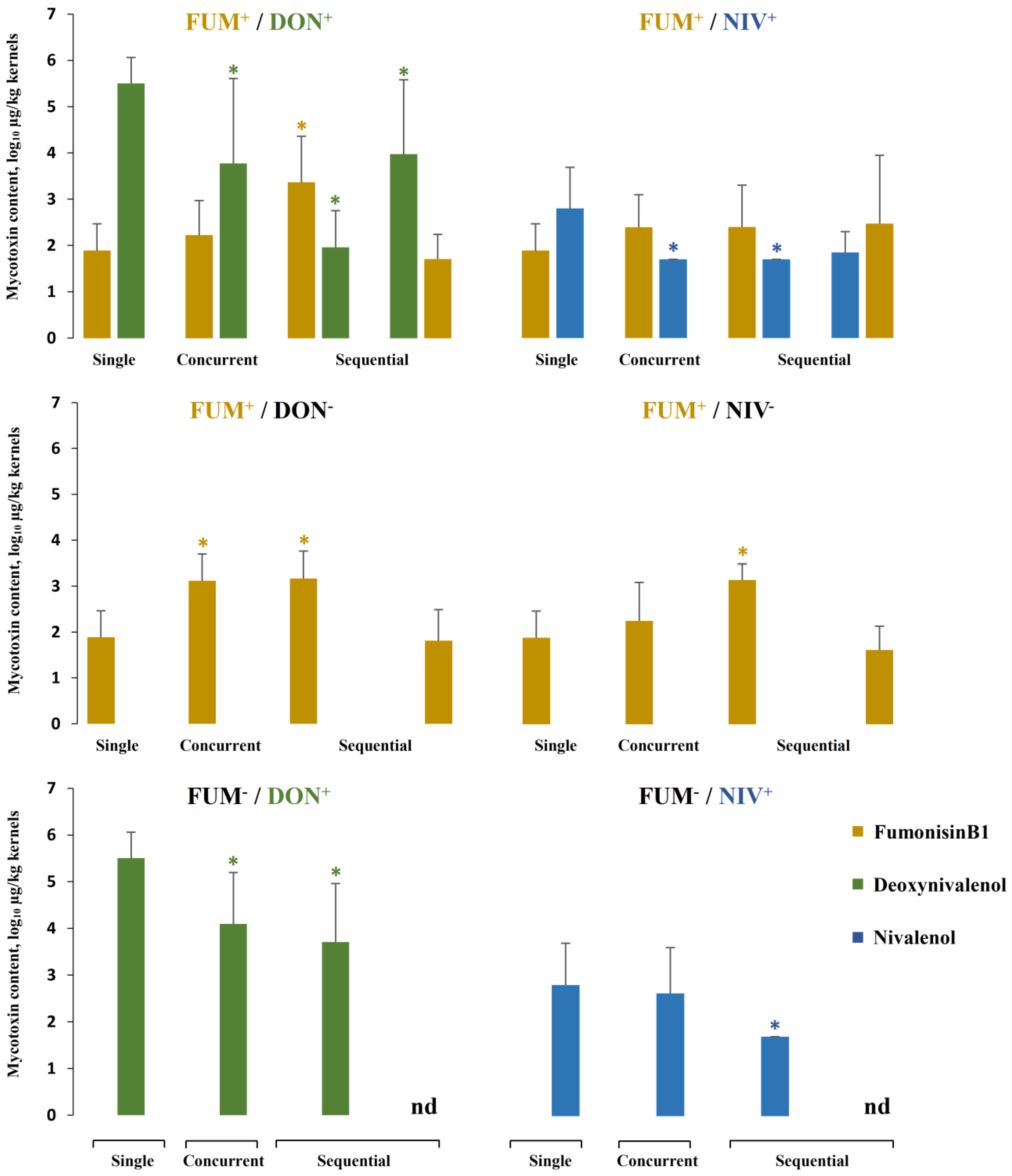
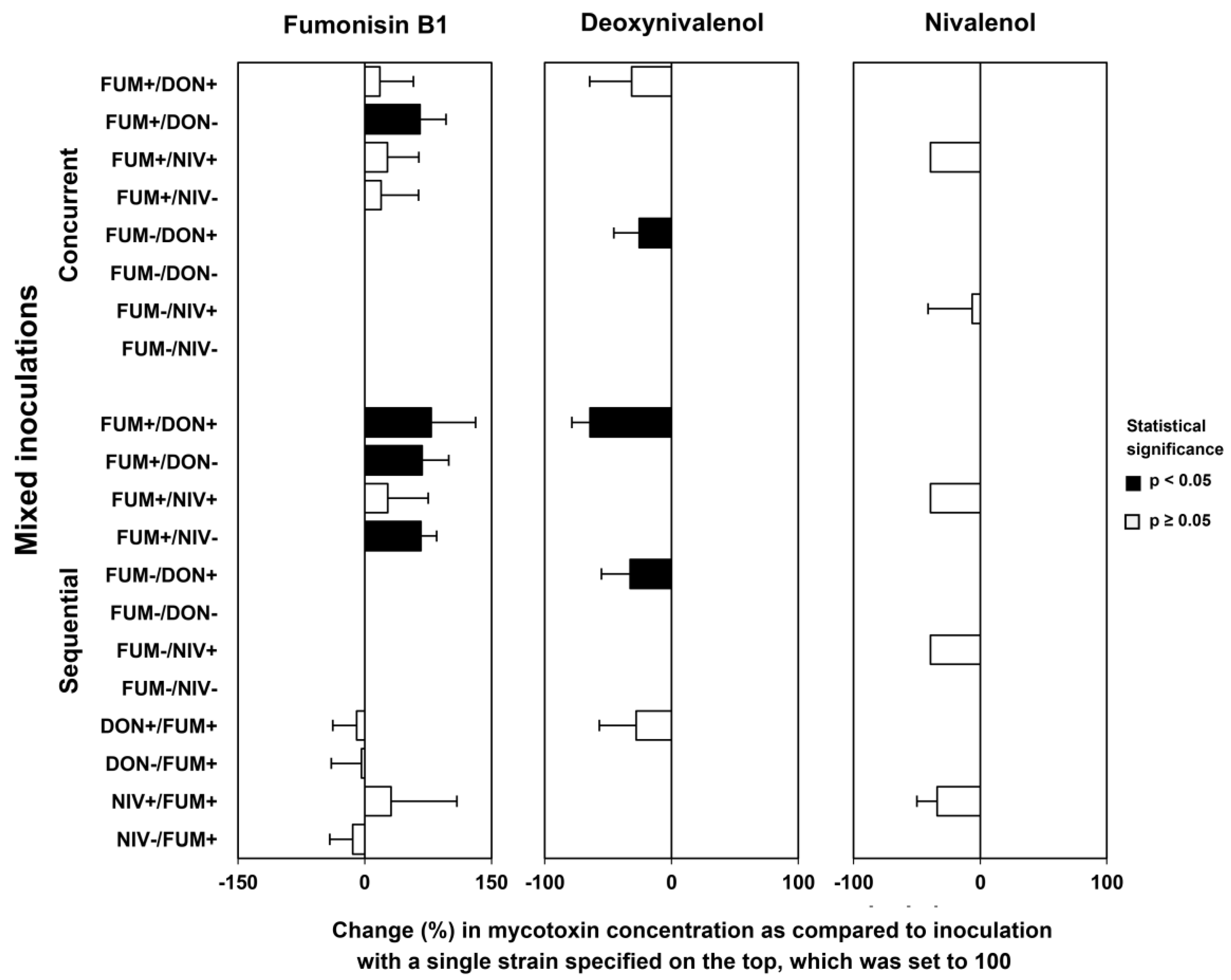
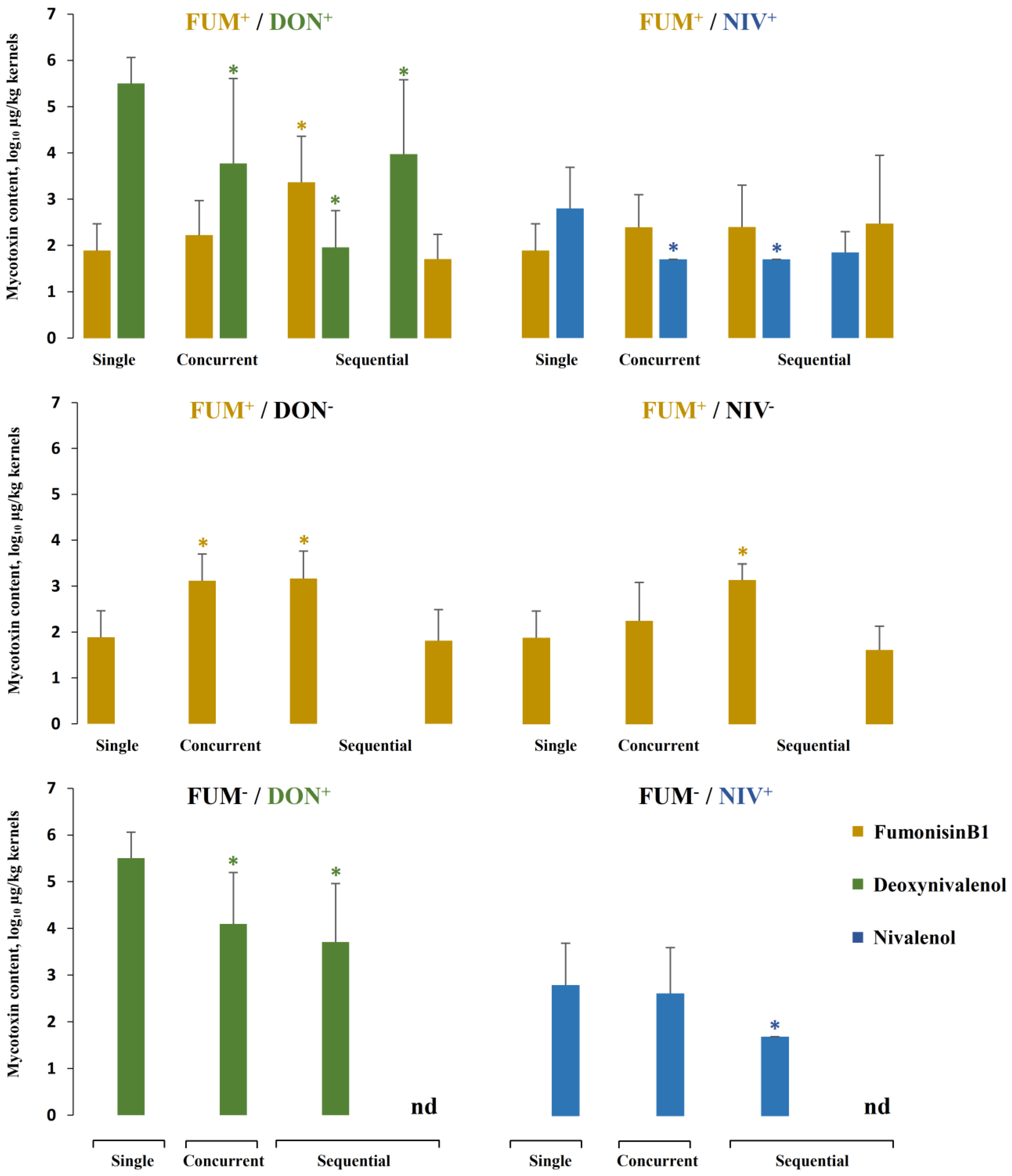
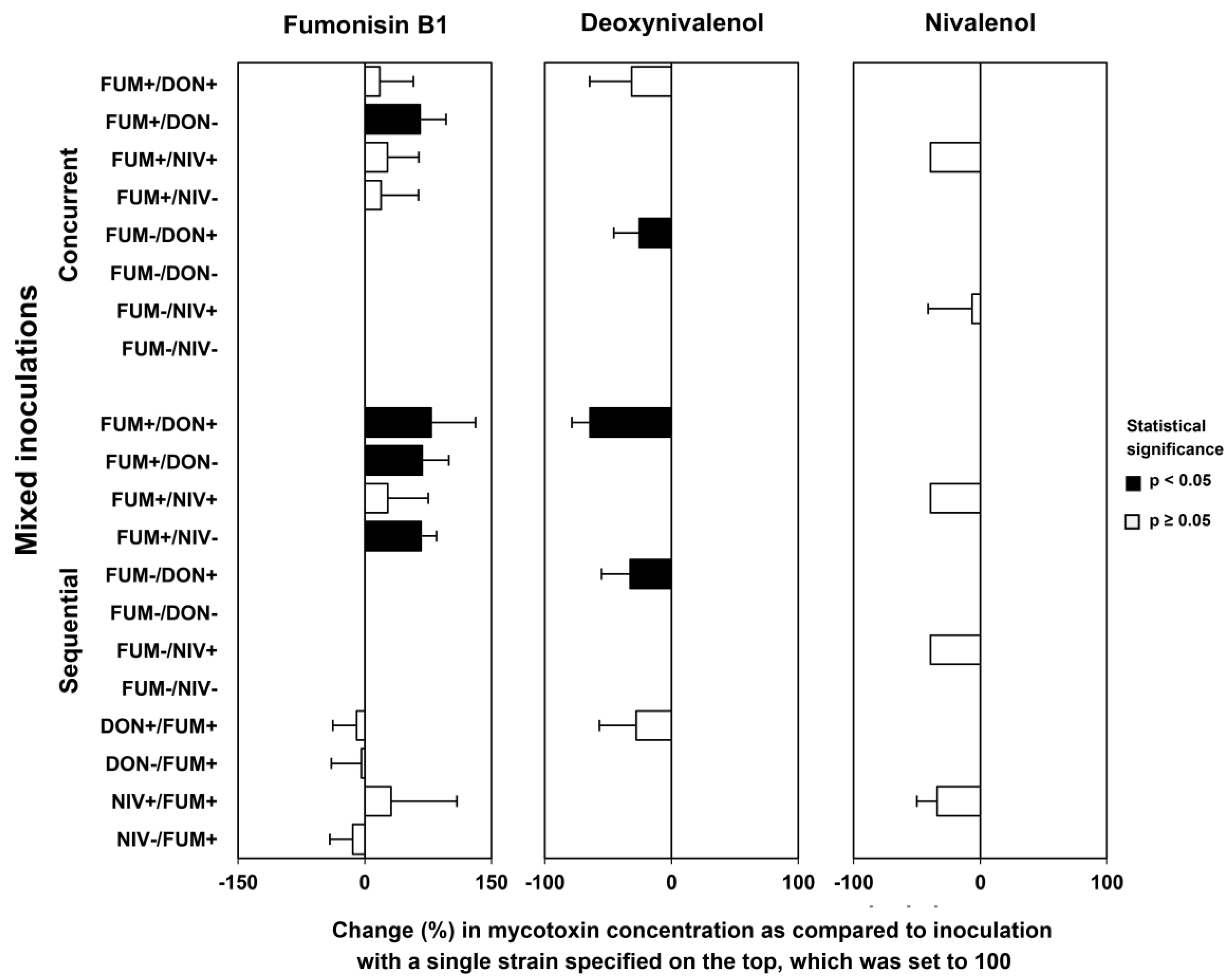
2.5. Mycotoxin Accumulation in Maize Ears Co-Inoculated with Both Fusarium Species
3. Discussion
3.1. Saprophytic Growth of F. verticillioides Is Stimulated by F. graminearum
3.2. Fumonisins Appar to Contribute to the Suppression of F. graminearum by F. verticillioides in Saprophytic Cultures, whereas Trichothecenes Are Not Involved in the Interaction
3.3. The Role of NIV as a Virulence Factor of F. graminearum in Maize Ears Was Corroborated
3.4. Trichothecenes Are Not Involved in the Stimulation of F. verticillioides Growth in Co-Infected Maize Ears
3.5. Why Has Fumonisin-Producing F. verticillioides Strain Suppressed F. graminearum More Strongly Than Its Fumonisin-Nonproducing Mutant In Vitro but Not In Planta?
3.6. Why Does F. graminearum Stimulate the Growth of F. verticilliodies?
3.7. Genetic Background of Fungal Strains Might Explain Differences between the Suppression of DON and NIV Chemotypes
3.8. What Is the Biological Function of Fumonisins in Planta?
4. Conclusions
5. Materials and Methods
5.1. Fungal Strains
5.2. Fungal Media, Fungal Inoculum, Plant Material, and Growth Conditions
5.3. Inoculation of Plants
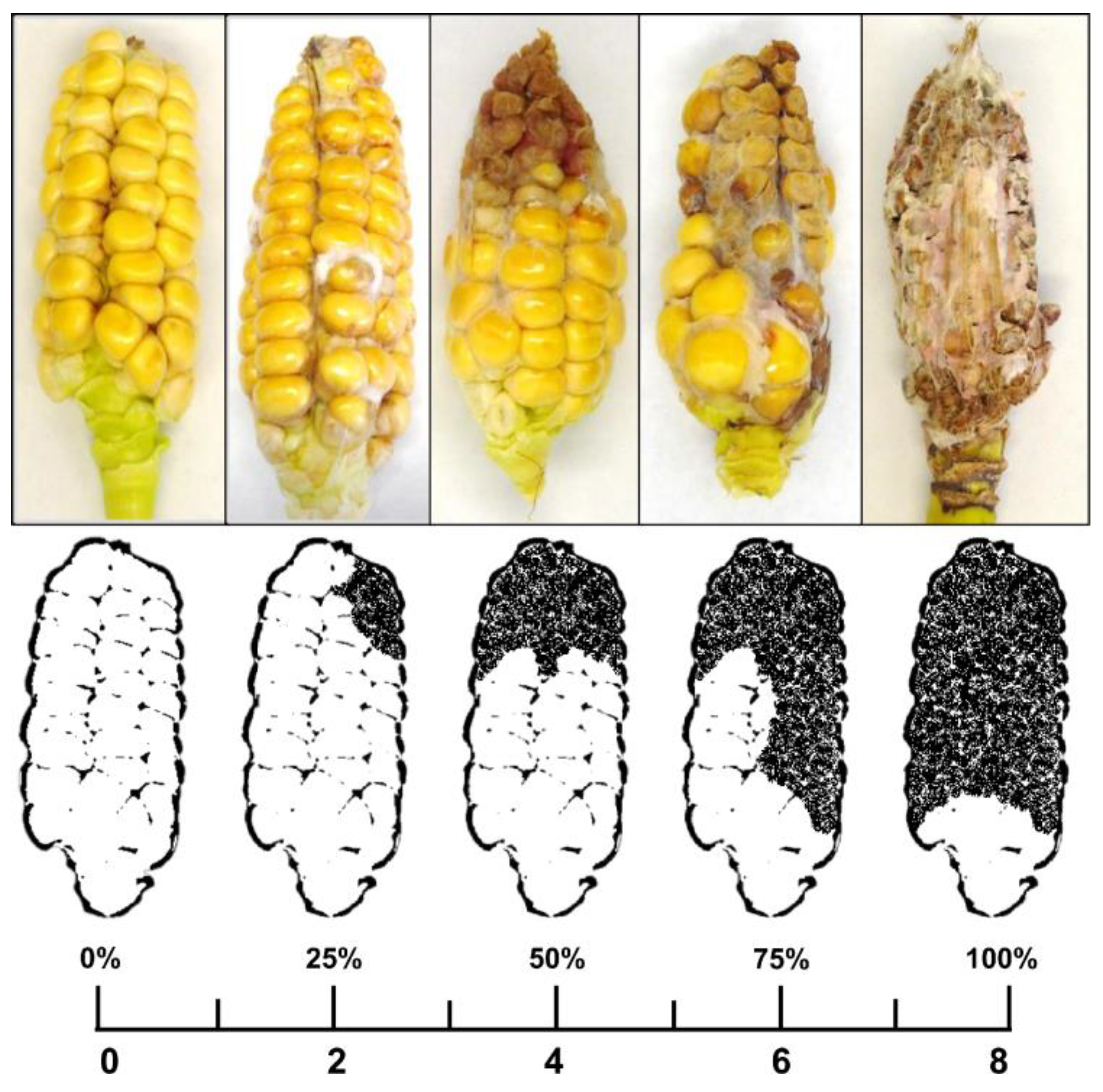
5.4. Scoring Disease Symptoms and Sampling
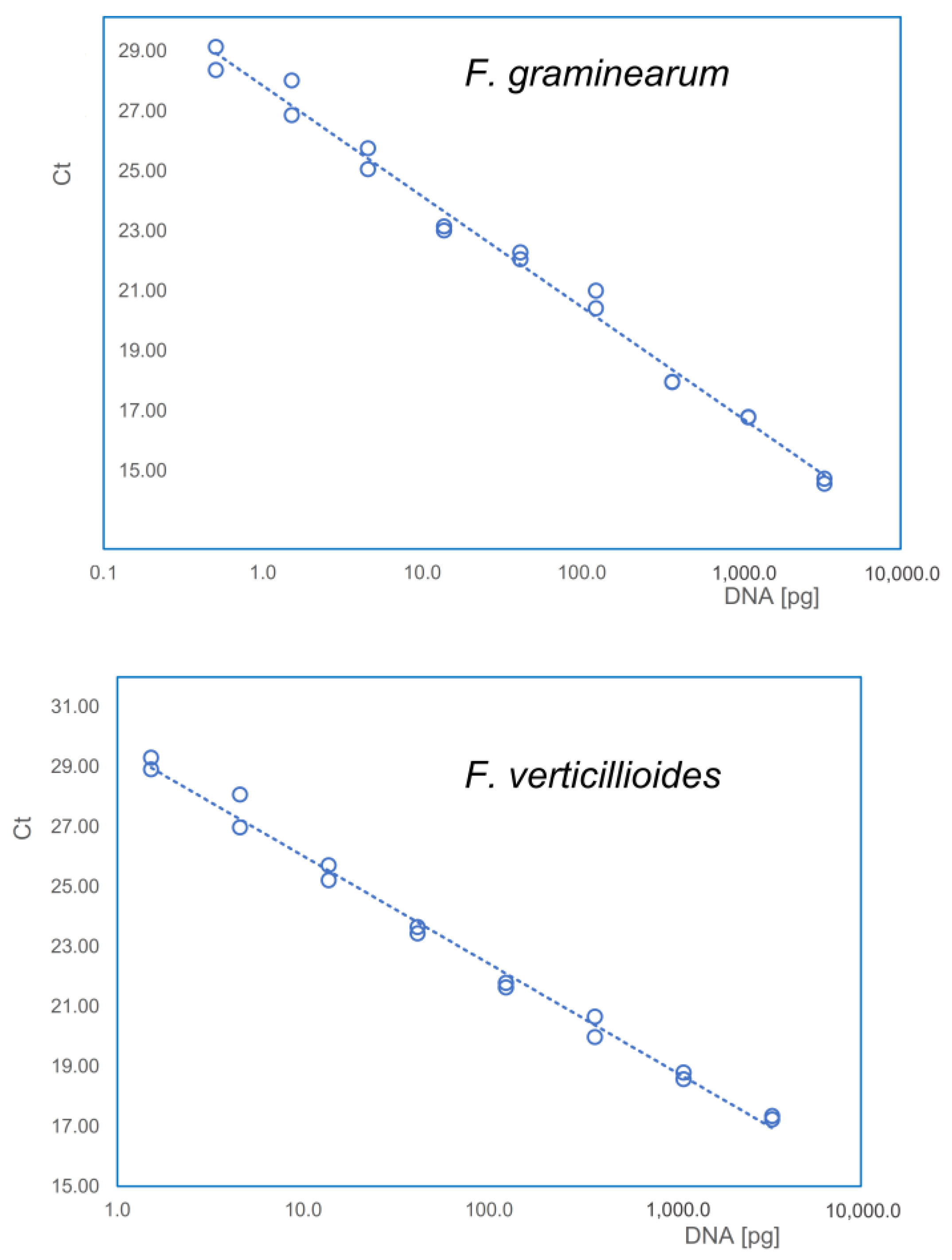
5.5. DNA Extraction and Quantification of Fungal Biomass
5.6. Extraction and Quantification of Mycotoxins
5.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain Combination a | Autoclaved Grains | Maize Ears b |
|---|---|---|
| FUM+/DON- | 3.5 ± 1.4 (6) c | 1.9 ± 2.1 (9) d |
| FUM+/NIV- | 2.8 ± 0.5 (6) | 0.6 ± 0.6 (10) |
| FUM+/NIV+ | 3.5 ± 1.6 (6) | 0.5 ± 0.4 (10) |
References
- FAOSTAT. 2021. Available online: https://www.fao.org/faostat/en/#data/QCL/visualize (accessed on 3 July 2023).
- de Lange, E.S.; Balmer, D.; Mauch-Mani, B.; Turlings, T.C.J. Insect and pathogen attack and resistance in maize and its wild ancestors, the teosintes. New Phytol. 2014, 204, 329–341. [Google Scholar] [CrossRef]
- Oldenburg, E.; Ellner, F. Distribution of disease symptoms and mycotoxins in maize ears infected by Fusarium culmorum and Fusarium graminearum. Mycotoxin Res. 2015, 31, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Munkvold, G.P. Epidemiology of Fusarium diseases and their mycotoxins in maize ears. Eur. J. Plant Pathol. 2003, 109, 705–713. [Google Scholar] [CrossRef]
- Vigier, B.; Reid, L.M.; Seifert, K.A.; Stewart, D.W.; Hamilton, R.I. Distribution and prediction of Fusarium species associated with maize ear rot in Ontario. Can. J. Plant Pathol. 1997, 19, 60–65. [Google Scholar] [CrossRef]
- Reid, L.M.; Nicol, R.W.; Ouellet, T.; Savard, M.; Miller, J.D.; Young, J.C.; Stewart, D.W.; Schaafsma, A.W. Interaction of Fusarium graminearum and F. moniliforme in maize ears: Disease progress, fungal biomass, and mycotoxin accumulation. Phytopathology 1999, 89, 1028–1037. [Google Scholar] [CrossRef] [PubMed]
- Bacon, C.W.; Hinton, D.M. Symptomless endophytic colonization of maize by Fusarium moniliforme. Can. J. Bot. 1996, 74, 1195–1202. [Google Scholar] [CrossRef]
- Munkvold, G.P.; Hellmich, R.L.; Showers, W.B. Reduced fusarium ear rot and symptomless infection in kernels of maize genetically engineered for European corn borer resistance. Phytopathology 1997, 87, 1071–1077. [Google Scholar] [CrossRef]
- Munkvold, G.P.; Mcgee, D.C.; Carlton, W.M. Importance of different pathways for maize kernel infection by Fusarium Moniliforme. Phytopathology 1997, 87, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Larran, S.; Perello, A.; Simon, M.R.; Moreno, V. The endophytic fungi from wheat (Triticum aestivum L.). World J. Microbiol. Biotechnol. 2007, 23, 565–572. [Google Scholar] [CrossRef]
- Quazi, S.A.J.; Burgess, L.W.; Smith-White, J. Colonization type of Gibberella zeae in Sorghum bicolor. J. Plant Pathol. 2010, 92, 261–265. [Google Scholar] [CrossRef]
- Doohan, F.M.; Brennan, J.; Cooke, B.M. Influence of climatic factors on Fusarium species pathogenic to cereals. Eur. J. Plant Pathol. 2003, 109, 755–768. [Google Scholar] [CrossRef]
- Miller, S.S.; Reid, L.M.; Harris, L.J. Colonization of maize silks by Fusarium graminearum, the causative organism of gibberella ear rot. Can. J. Bot. 2007, 85, 369–376. [Google Scholar] [CrossRef]
- Koehler, B. Natural mode of entrance of fungi into corn ears and some symptoms that indicate infection. J. Agric. Res. 1942, 64, 421–442. [Google Scholar]
- Thompson, M.E.H.; Raizada, M.N. Fungal pathogens of maize gaining free passage along the silk road. Pathogens 2018, 7, 81. [Google Scholar] [CrossRef] [PubMed]
- Duncan, K.E.; Howard, R.J. Biology of maize kernel infection by Fusarium verticillioides. Mol. Plant Microbe Interact. 2010, 23, 6–16. [Google Scholar] [CrossRef] [PubMed]
- Bacon, C.W.; Glenn, A.E.; Yates, I.E. Fusarium verticillioides: Managing the endophytic association with maize for reduced fumonisins accumulation. Toxin Rev. 2008, 27, 411–446. [Google Scholar] [CrossRef]
- Dastjerdi, R.; Karlovsky, P. Systemic infection of maize, sorghum, rice, and beet seedlings with fumonisin-producing and nonproducing Fusarium verticillioides strains. Plant Pathol. J. 2015, 31, 334–342. [Google Scholar] [CrossRef] [PubMed]
- Bacon, C.W.; Bennett, R.M.; Hinton, D.M.; Voss, K.A. Scanning electron microscopy of Fusarium moniliforme within asymptomatic corn kernels and kernels associated with equine leukoencephalomalacia. Plant Dis. 1992, 76, 144–148. [Google Scholar] [CrossRef]
- Sumarah, M.W. The deoxynivalenol challenge. J. Agric. Food Chem. 2022, 70, 9619–9624. [Google Scholar] [CrossRef]
- Schiwek, S.; Alhussein, M.; Rodemann, C.; Budragchaa, T.; Beule, L.; von Tiedemann, A.; Karlovsky, P. Fusarium culmorum produces NX-2 toxin simultaneously with deoxynivalenol and 3-acetyl-deoxynivalenol or nivalenol. Toxins 2022, 14, 456. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel CONTAM; Knutsen, H.K.; Alexander, J.; Barregård, L.; Bignami, M.; Brüschweiler, B.; Ceccatelli, S.; Cottrill, B.; Dinovi, M.; Grasl-Kraupp, B.; et al. Risks to human and animal health related to the presence of deoxynivalenol and its acetylated and modified forms in food and feed. EFSA J. 2017, 15, e04718. [Google Scholar] [CrossRef] [PubMed]
- Proctor, R.H.; Hohn, T.M.; McCormick, S.P. Reduced virulence of Gibberella zeae caused by disruption of a trichothecene toxin biosynthetic gene. Mol. Plant Microbe Interact. 1995, 8, 593–601. [Google Scholar] [CrossRef]
- Jansen, C.; von Wettstein, D.; Schäfer, W.; Kogel, K.-H.; Felk, A.; Maier, F.J. Infection patterns in barley and wheat spikes inoculated with wild-type and trichodiene synthase gene disrupted Fusarium Graminearum. Proc. Natl. Acad. Sci. USA 2005, 102, 16892–16897. [Google Scholar] [CrossRef] [PubMed]
- Maier, F.J.; Miedaner, T.; Hadeler, B.; Felk, A.; Salomon, S.; Lemmens, M.; Kassner, H.; Schäfer, W. Involvement of trichothecenes in fusarioses of wheat, barley and maize evaluated by gene disruption of the trichodiene synthase (Tri5) gene in three field isolates of different chemotype and virulence. Mol. Plant Pathol. 2006, 7, 449–461. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wen, J.; Tang, Y.; Shi, J.; Mu, G.; Yan, R.; Cai, J.; Long, M. Research progress on fumonisin B1 contamination and toxicity: A review. Molecules 2021, 26, 5238. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Abbas, H.K.; Duke, S.O. Structure-dependent phytotoxicity of fumonisins and related compounds in a duckweed bioassay. Phytochemistry 1993, 33, 779–785. [Google Scholar] [CrossRef]
- Renaud, J.B.; DesRochers, N.; Hoogstra, S.; Garnham, C.P.; Sumarah, M.W. Structure activity relationship for fumonisin phytotoxicity. Chem. Res. Toxicol. 2021, 34, 1604–1611. [Google Scholar] [CrossRef]
- Shim, W.-B.; Flaherty, J.E.; Woloshuk, C.P. Comparison of fumonisin B1 biosynthesis in maize germ and degermed kernels by Fusarium verticillioides. J. Food Prot. 2003, 66, 2116–2122. [Google Scholar] [CrossRef] [PubMed]
- Desjardins, A.E.; Plattner, R.D. Fumonisin B1-nonproducing strains of Fusarium verticillioides cause maize (Zea mays) ear infection and ear rot. J. Agric. Food Chem. 2000, 48, 5773–5780. [Google Scholar] [CrossRef]
- Desjardins, A.E.; Munkvold, G.P.; Plattner, R.D.; Proctor, R.H. FUM1—A gene required for fumonisin biosynthesis but not for maize ear rot and ear infection by Gibberella moniliformis in field tests. Mol. Plant Microbe Interact. 2002, 15, 1157–1164. [Google Scholar] [CrossRef]
- Proctor, R.H.; Desjardins, A.E.; McCormick, S.P.; Plattner, R.D.; Alexander, N.J.; Brown, D.W. Genetic analysis of the role of trichothecene and fumonisin mycotoxins in the virulence of Fusarium. Eur. J. Plant Pathol. 2002, 108, 691–698. [Google Scholar] [CrossRef]
- Williams, L.D.; Glenn, A.E.; Zimeri, A.M.; Bacon, C.W.; Smith, M.A.; Riley, R.T. Fumonisin disruption of ceramide biosynthesis in maize roots and the effects on plant development and Fusarium verticillioides-induced seedling disease. J. Agric. Food Chem. 2007, 55, 2937–2946. [Google Scholar] [CrossRef] [PubMed]
- Glenn, A.E.; Zitomer, N.C.; Zimeri, A.M.; Williams, L.D.; Riley, R.T.; Proctor, R.H. Transformation-mediated complementation of a FUM gene cluster deletion in Fusarium verticillioides restores both fumonisin production and pathogenicity on maize seedlings. Mol. Plant Microbe Interact. 2008, 21, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Audenaert, K.; Vanheule, A.; Höfte, M.; Haesaert, G. Deoxynivalenol: A major player in the multifaceted response of Fusarium to its environment. Toxins 2014, 6, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Marín, S.; Sanchis, V.; Ramos, A.J.; Vinas, I.; Magan, N. Environmental factors, in vitro interactions, and niche overlap between Fusarium moniliforme, F. proliferatum, and F. graminearum, Aspergillus and Penicillium species from maize grain. Mycol. Res. 1998, 102, 831–837. [Google Scholar] [CrossRef]
- Velluti, A.; Marín, S.; Gonzalez, R.; Ramos, A.J.; Sanchis, V. Fumonisin B1, zearalenone and deoxynivalenol production by Fusarium moniliforme, F proliferatum and F graminearum in mixed cultures on irradiated maize kernels. J. Sci. Food Agric. 2001, 81, 88–94. [Google Scholar] [CrossRef]
- Velluti, A.; Marín, S.; Bettucci, L.; Ramos, A.J.; Sanchis, V. the effect of fungal competition on colonization of maize grain by Fusarium moniliforme, F. proliferatum and F. graminearum and on fumonisin B1 and zearalenone formation. Int. J. Food Microbiol. 2000, 59, 59–66. [Google Scholar] [CrossRef]
- Chatterjee, S.; Kuang, Y.; Splivallo, R.; Chatterjee, P.; Karlovsky, P. Interactions among filamentous fungi Aspergillus niger, Fusarium verticillioides and Clonostachys rosea: Fungal biomass, diversity of secreted metabolites and fumonisin production. BMC Microbiol. 2016, 16, 83. [Google Scholar] [CrossRef] [PubMed]
- Leggieri, M.C.; Giorni, P.; Pietri, A.; Battilani, P. Aspergillus flavus and Fusarium verticillioides interaction: Modeling the impact on mycotoxin production. Front. Microbiol. 2019, 10, 2653. [Google Scholar] [CrossRef] [PubMed]
- Wicklow, D.T. Interference competition. In The Fungal Community, Wicklow DR and Carrol GC (edsx); Marcel Dekker Inc.: New York, NY, USA, 1981; pp. 351–375. [Google Scholar]
- Venkatesh, N.; Keller, N. Mycotoxins in conversation with bacteria and fungi. Front. Microbiol. 2019, 10, 403. [Google Scholar] [CrossRef] [PubMed]
- Keyser, Z.; Vismer, H.F.; Klaasen, J.A.; Snijman, P.W.; Marasas, W.F. The antifungal effect of fumonisin B1 on Fusarium and other fungal species. S. Afr. J. Sci. 1999, 95, 455–458. [Google Scholar]
- Dawidziuk, A.; Koczyk, G.; Popiel, D. Adaptation and response to mycotoxin presence in pathogen-pathogen interactions within the Fusarium genus. World Mycotoxin J. 2016, 9, 565–575. [Google Scholar] [CrossRef]
- Ueno, Y. The toxicology of mycotoxins. Crit. Rev. Toxicol. 1985, 14, 99–132. [Google Scholar] [CrossRef]
- Campos, F.F.; Johann, S.; Cota, B.B.; Alves, T.M.A.; Rosa, L.H.; Caligiorne, R.B.; Cisalpino, P.S.; Rosa, C.A.; Zani, C.L. Antifungal activity of trichothecenes from Fusarium sp. against clinical isolates of Paracoccidioides brasiliensis. Mycoses 2011, 54, e122–e129. [Google Scholar] [CrossRef] [PubMed]
- Popiel, D.; Dawidziuk, A.; Koczyk, G. Efflux pumps as an additional source of resistance to trichothecenes in Fusarium proliferatum and Fusarium oxysporum isolates. J. Appl. Genet. 2019, 60, 405–416. [Google Scholar] [CrossRef] [PubMed]
- Cooney, J.M.; Lauren, D.R.; die Menna, M.E. Impact of competitive fungi on trichothecene production by Fusarium graminearum. J. Agric. Food Chem. 2001, 49, 522–526. [Google Scholar] [CrossRef] [PubMed]
- Yoshizawa, T.; Morooka, N. Biological modification of trichothecene mycotoxins: Acetylation and deacetylation of deoxynivalenols by Fusarium spp. Appl. Environ. Microbiol. 1975, 29, 54–58. [Google Scholar] [CrossRef] [PubMed]
- Tran, S.T.; Smith, T.K. Conjugation of deoxynivalenol by Alternaria alternata (54028 nrrl), Rhizopus microsporus var. rhizopodiformis (54029 nrrl) and Aspergillus oryzae (5509 nrrl). Mycotoxin Res. 2014, 30, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Tan, Y.; Liu, N.; Yan, Z.; Liao, Y.; Chen, J.; de Saeger, S.; Yang, H.; Zhang, Q.; Wu, A. Detoxification of deoxynivalenol via glycosylation represents novel insights on antagonistic activities of Trichoderma when confronted with Fusarium graminearum. Toxins 2016, 8, 335. [Google Scholar] [CrossRef] [PubMed]
- Garnham, C.P.; Butler, S.G.; Telmer, P.G.; Black, F.E.; Renaud, J.B.; Sumarah, M.W. Identification and characterization of an Aspergillus niger amine oxidase that detoxifies intact fumonisins. J. Agric. Food Chem. 2020, 68, 13779–13790. [Google Scholar] [CrossRef]
- Utermark, J.; Karlovsky, P. Role of zearalenone lactonase in protection of Gliocladium roseum from fungitoxic effects of the mycotoxin zearalenone. Appl. Environ. Microbiol. 2007, 73, 637–642. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Vinas, M.; Alsarrag, A.; Su, L.; Pfohl, K.; Rohlfs, M.; Schäfer, W.; Chen, W.; Karlovsky, P. Bis-naphthopyrone pigments protect filamentous Ascomycetes from a wide range of predators. Nat. Commun. 2019, 10, 3579. [Google Scholar] [CrossRef]
- Picot, A.; Hourcade-Marcolla, D.; Barreau, C.; Pinson-Gadais, L.; Caron, D.; Richard-Forget, F.; Lannou, C. Interactions between Fusarium verticillioides and Fusarium graminearum in maize ears and consequences for fungal development and mycotoxin accumulation. Plant Pathol. 2012, 61, 140–151. [Google Scholar] [CrossRef]
- Giorni, P.; Bertuzzi, T.; Battilani, P. Impact of fungi co-occurrence on mycotoxin contamination in maize during the growing season. Front. Microbiol. 2019, 10, 1265. [Google Scholar] [CrossRef] [PubMed]
- Sherif, M.; Becker, E.-M.; Herrfurth, C.; Feussner, I.; Karlovsky, P.; Splivallo, R. Volatiles emitted from maize ears simultaneously infected with two Fusarium species mirror the most competitive fungal pathogen. Front. Plant Sci. 2016, 7, 1460. [Google Scholar] [CrossRef] [PubMed]
- Wicklow, D.T.; Horn, B.W.; Shotwell, O.L.; Hesseltine, C.W.; Caldwell, R.W. Fungal interference with Aspergillus flavus infection and aflatoxin contamination of maize grown in a controlled environment. Phytopathology 1988, 78, 68–74. [Google Scholar] [CrossRef]
- Chen, X.; Landschoot, S.; Detavernier, C.; De Saeger, S.; Rajkovic, A.; Audenaert, K. Cross-talk between Fusarium verticillioides and Aspergillus flavus in vitro and in planta. Mycotoxin Res. 2021, 37, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Pan, J.J.; May, G. Endophytic Fusarium verticillioides reduces disease severity caused by Ustilago maydis on maize. FEMS Microbiol. Lett. 2009, 299, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, M.F.; De Boevre, M.; Landschoot, S.; De Saeger, S.; Haesaert, G.; Audenaert, K. Fungal endophytes control Fusarium graminearum and reduce trichothecenes and zearalenone in maize. Toxins 2018, 10, 493. [Google Scholar] [CrossRef]
- Sarrocco, S.; Valenti, F.; Manfredini, S.; Esteban, P.; Bernardi, R.; Puntoni, G.; Baroncelli, R.; Haidukowski, M.; Moretti, A.; Vannacci, G. Is exploitation competition involved in a multitrophic strategy for the biocontrol of Fusarium head blight? Phytopathology 2019, 109, 560–570. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Jensen, J.D.; Svensson, B.; Jørgensen, H.J.L.; Collinge, D.B.; Finnie, C. Secretomics identifies Fusarium graminearum proteins involved in the interaction with barley and wheat. Mol. Plant Pathol. 2012, 13, 445–453. [Google Scholar] [CrossRef]
- Geißinger, C.; Gastl, M.; Becker, T. Enzymes from cereal and Fusarium metabolism involved in the malting process—A review. J. Amer. Soc. Brew. Chem. 2022, 80, 1–16. [Google Scholar] [CrossRef]
- Gao, M.; Glenn, A.E.; Gu, X.; Mitchell, T.R.; Satterlee, T.; Duke, M.V.; Scheffler, B.E.; Gold, S.E. Pyrrocidine, a molecular off switch for fumonisin biosynthesis. PLoS Pathog. 2020, 16, e1008595. [Google Scholar] [CrossRef]
- Naef, A.; Senatore, M.; Défago, G. A microsatellite based method for quantification of fungi in decomposing plant material elucidates the role of Fusarium graminearum DON production in the saprophytic competition with Trichoderma atroviride in maize tissue microcosms. FEMS Microbiol. Ecol. 2006, 55, 211–220. [Google Scholar] [CrossRef]
- Müller, M.E.; Steier, I.; Köppen, R.; Siegel, D.; Proske, M.; Korn, U.; Koch, M. Cocultivation of phytopathogenic Fusarium and Alternaria strains affects fungal growth and mycotoxin production. J. Appl. Microbiol. 2012, 113, 874–887. [Google Scholar] [CrossRef] [PubMed]
- Reid, L.M.; Woldemariam, T.; Zhu, X.; Stewart, D.W.; Schaafsma, A.W. Effect of inoculation time and point of entry on disease severity in Fusarium graminearum, Fusarium verticillioides, or Fusarium subglutinans inoculated maize ears. Can. J. Plant Pathol. 2002, 24, 162–167. [Google Scholar] [CrossRef]
- Miedaner, T.; Bolduan, C.; Melchinger, A.E. Aggressiveness and mycotoxin production of eight isolates each of Fusarium graminearum and Fusarium verticillioides for ear rot on susceptible and resistant early maize inbred lines. Eur. J. Plant Pathol. 2010, 127, 113–123. [Google Scholar] [CrossRef]
- Miedaner, T.; Reinbrecht, C.; Schilling, A.G. Association among aggressiveness, fungal colonization, and mycotoxin production of 26 isolates of Fusarium graminearum in winter rye head blight. J. Plant Dis. Prot. 2000, 107, 124–134. [Google Scholar]
- Becker, E.-M.; Herrfurth, C.; Irmisch, S.; Köllner, T.G.; Feussner, I.; Karlovsky, P.; Splivallo, R. Infection of corn ears by Fusarium spp. induces the emission of volatile sesquiterpenes. J. Agric. Food Chem. 2014, 62, 5226–5236. [Google Scholar] [CrossRef] [PubMed]
- Shu, X.; Livingston, D.P.; Woloshuk, C.P.; Payne, G.A. Comparative histological and transcriptional analysis of maize kernels infected with Aspergillus flavus and Fusarium verticillioides. Front. Plant Sci. 2017, 8, 2075. [Google Scholar] [CrossRef] [PubMed]
- Siou, D.; Gélisse, S.; Laval, V.; Suffert, F.; Lannou, C. Mutual exclusion between fungal species of the Fusarium head blight complex in a wheat spike. Appl. Environ. Microbiol. 2015, 81, 4682–4689. [Google Scholar] [CrossRef] [PubMed]
- Walkowiak, S.; Bonner, C.T.; Wang, L.; Blackwell, B.; Rowland, O.; Subramaniam, R. Intraspecies interaction of Fusarium graminearum contributes to reduced toxin production and virulence. Mol. Plant Pathog. Interact. 2015, 28, 1256–1267. [Google Scholar] [CrossRef]
- Bahadoor, A.; Brauer, E.K.; Bosnich, W.; Schneiderman, D.; Johnston, A.; Aubin, Y.; Blackwell, B.; Melanson, J.E.; Harris, L.J. Gramillin A and B: Cyclic lipopeptides identified as the nonribosomal biosynthetic products of Fusarium graminearum. J. Am. Chem. Soc. 2018, 140, 16783–16791. [Google Scholar] [CrossRef] [PubMed]
- Warfield, C.Y.; Gilchrist, D.G. Influence of kernel age on fumonisin B1 production in maize by Fusarium moniliforme. Appl Environ. Microbiol. 1999, 65, 2853–2856. [Google Scholar] [CrossRef] [PubMed]
- Proctor, R.H.; Desjardins, A.E.; Plattner, R.D.; Hohn, T.M. A polyketide synthase gene required for biosynthesis of fumonisin mycotoxins in Gibberella fujikuroi mating population A. Fungal Genet. Biol. 1999, 27, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Hanway, J.J.; Ritchie, S.W. How a Corn Plant Develops. Iowa State University Publications Online. 1986, Report No. 48. Available online: http://publications.iowa.gov/id/eprint/18027 (accessed on 3 June 2022).
- Brandfass, C.; Karlovsky, P. Upscaled CTAB-based DNA extraction and real-time PCR assays for Fusarium culmorum and F. graminearum DNA in plant material with reduced sampling error. Int. J. Mol. Sci. 2008, 9, 2306–2321. [Google Scholar] [CrossRef] [PubMed]
- Nutz, S.; Döll, K.; Karlovsky, P. Determination of the LOQ in real-time PCR by receiver operating characteristic curve analysis: Application to qPCR assays for Fusarium verticillioides and F. Proliferatum. Anal. Bioanal. Chem. 2011, 401, 717–726. [Google Scholar] [CrossRef]
- Ten Bosch, L.; Pfohl, K.; Avramidis, G.; Wieneke, S.; Viöl, W.; Karlovsky, P. Plasma-based degradation of mycotoxins produced by Fusarium, Aspergillus and Alternaria species. Toxins 2017, 9, 97. [Google Scholar] [CrossRef] [PubMed]
- Adejumo, T.O.; Hettwer, U.; Karlovsky, P. Survey of maize from South-Western Nigeria for zearalenone, alpha- and beta-zearalenols, fumonisin B-1 and enniatins produced by Fusarium species. Food Addit. Contam. 2007, 24, 993–1000. [Google Scholar] [CrossRef] [PubMed]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Wenzl, T.; Haedrich, J.; Schaechtele, A.; Robouch, P.; Stroka, J. Guidance Document on the Estimation of LOD and LOQ for Measurements in the Field of Contaminants in Feed and Food; EUR 28099; Publications Office of the European Union: Luxembourg, 2016; ISBN 978-92-79-61768-3. [Google Scholar] [CrossRef]




| Strain | Code | Source |
|---|---|---|
| F. graminearum 2311 | DON+ | |
| F. graminearum 2311#2899 a | DON− | Wilhelm Schäfer, Hamburg University, Hamburg, Germany |
| F. graminearum 06 | NIV+ | |
| F. graminearum 06#7 b | NIV− | |
| F. verticillioides M-3125 | FUM+ | Robert Proctor, National Center for Agricultural Utilization Research/U. S. Department of Agriculture Peoria, IL, USA |
| F. verticillioides GfA2364 c | FUM− |
| Inoculation Type | Fungal Strains a |
|---|---|
| Single | DON+, DON−, NIV+, NIV−, FUM+, FUM- |
| Concurrent | FUM+/DON+, FUM+/DON−, FUM+/NIV+, FUM+/NIV−, FUM−/DON+, FUM-/DON−, FUM−/NIV+, FUM−/NIV− |
| Sequential b | FUM+/DON+, FUM+/DON−, FUM+/NIV+, FUM+/NIV−, FUM−/DON+, FUM−/DON−, FUM−/NIV+, FUM−/NIV−, DON+/FUM+, DON−/FUM+, NIV+/FUM+, NIV−/FUM+ |
| Control | Mock inoculation with sterile tap water |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sherif, M.; Kirsch, N.; Splivallo, R.; Pfohl, K.; Karlovsky, P. The Role of Mycotoxins in Interactions between Fusarium graminearum and F. verticillioides Growing in Saprophytic Cultures and Co-Infecting Maize Plants. Toxins 2023, 15, 575. https://doi.org/10.3390/toxins15090575
Sherif M, Kirsch N, Splivallo R, Pfohl K, Karlovsky P. The Role of Mycotoxins in Interactions between Fusarium graminearum and F. verticillioides Growing in Saprophytic Cultures and Co-Infecting Maize Plants. Toxins. 2023; 15(9):575. https://doi.org/10.3390/toxins15090575
Chicago/Turabian StyleSherif, Mohammed, Nadine Kirsch, Richard Splivallo, Katharina Pfohl, and Petr Karlovsky. 2023. "The Role of Mycotoxins in Interactions between Fusarium graminearum and F. verticillioides Growing in Saprophytic Cultures and Co-Infecting Maize Plants" Toxins 15, no. 9: 575. https://doi.org/10.3390/toxins15090575
APA StyleSherif, M., Kirsch, N., Splivallo, R., Pfohl, K., & Karlovsky, P. (2023). The Role of Mycotoxins in Interactions between Fusarium graminearum and F. verticillioides Growing in Saprophytic Cultures and Co-Infecting Maize Plants. Toxins, 15(9), 575. https://doi.org/10.3390/toxins15090575