Inbred Selection for Increased Resistance to Kernel Contamination with Fumonisins





Abstract
1. Introduction
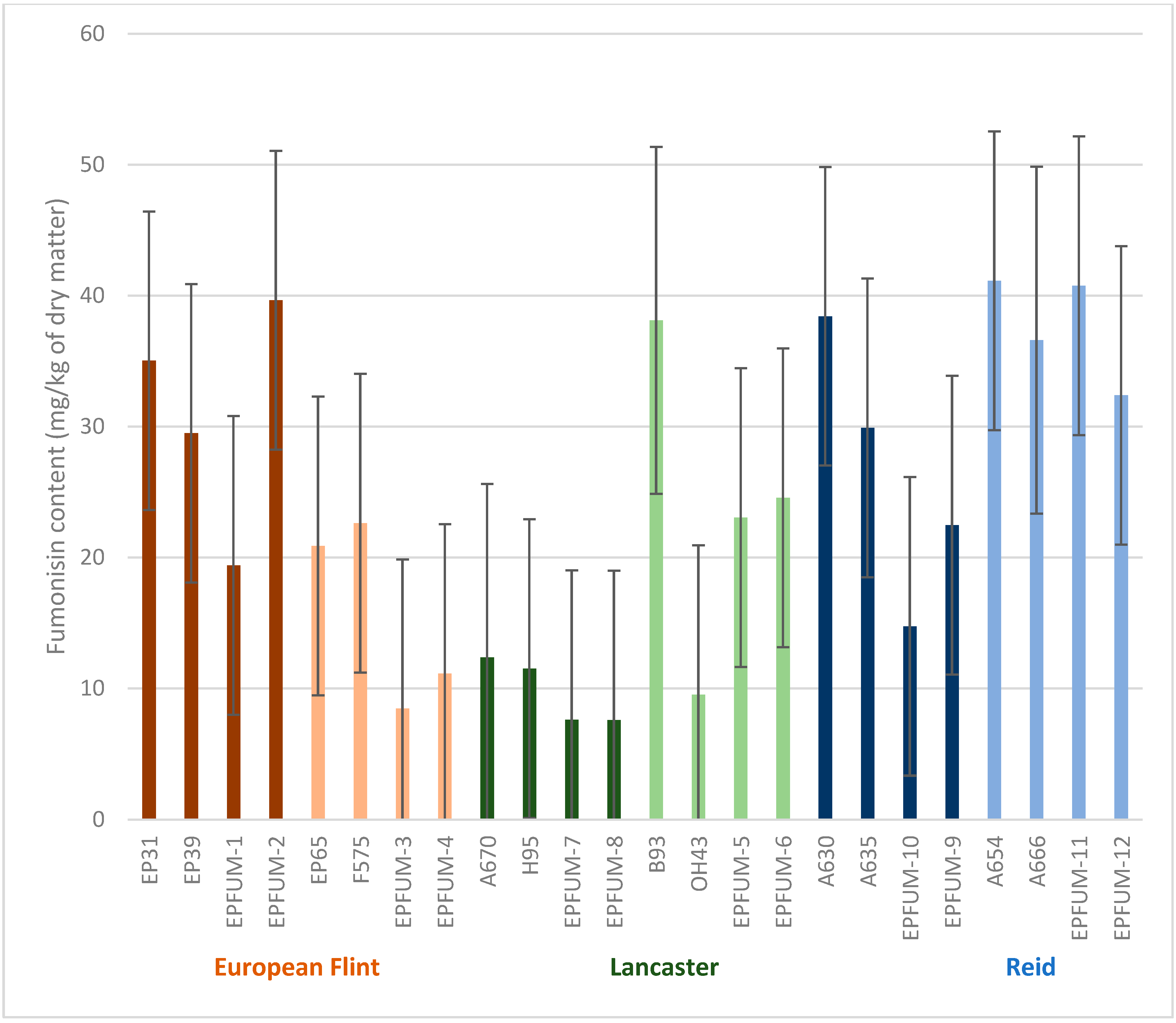
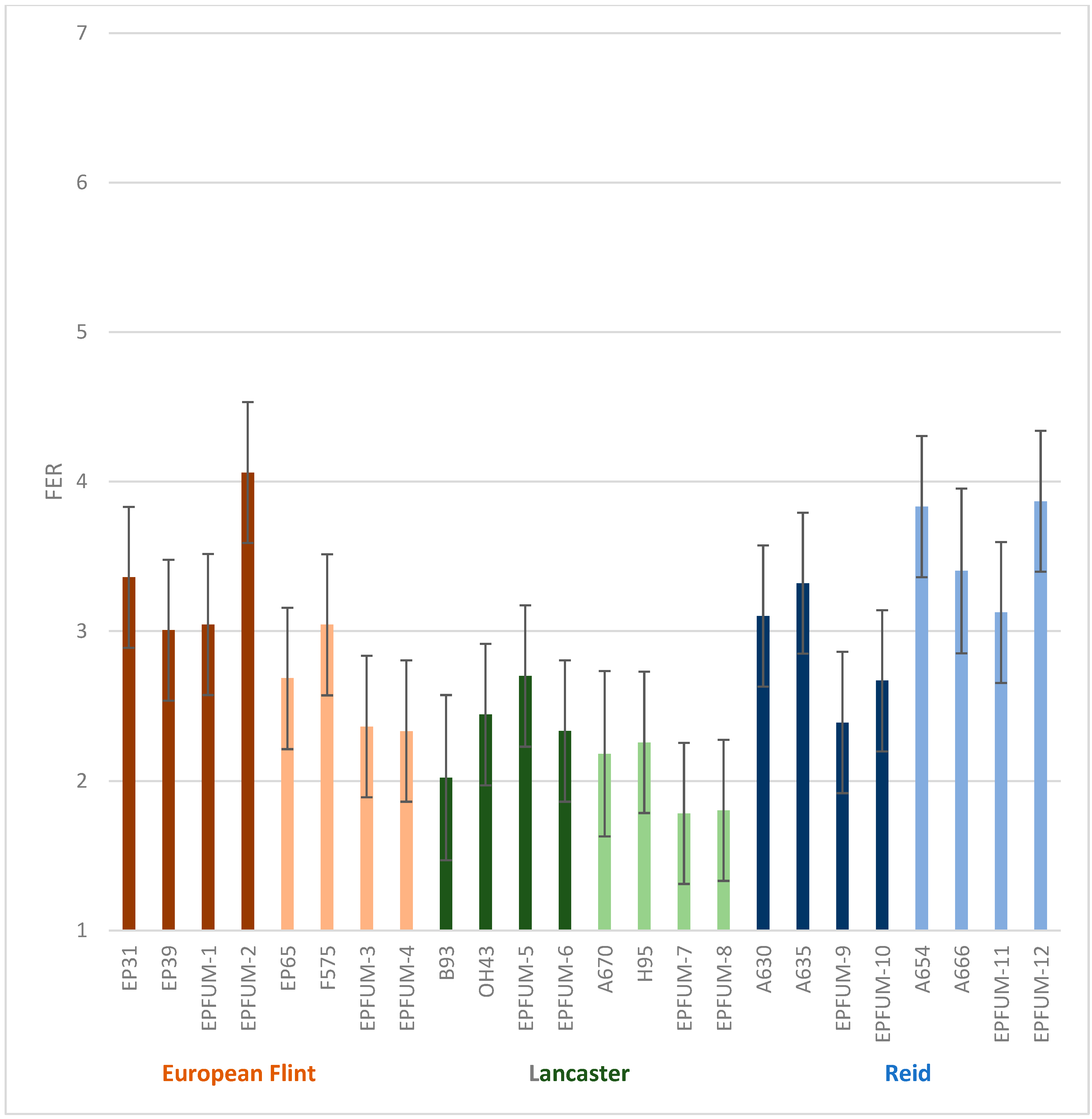
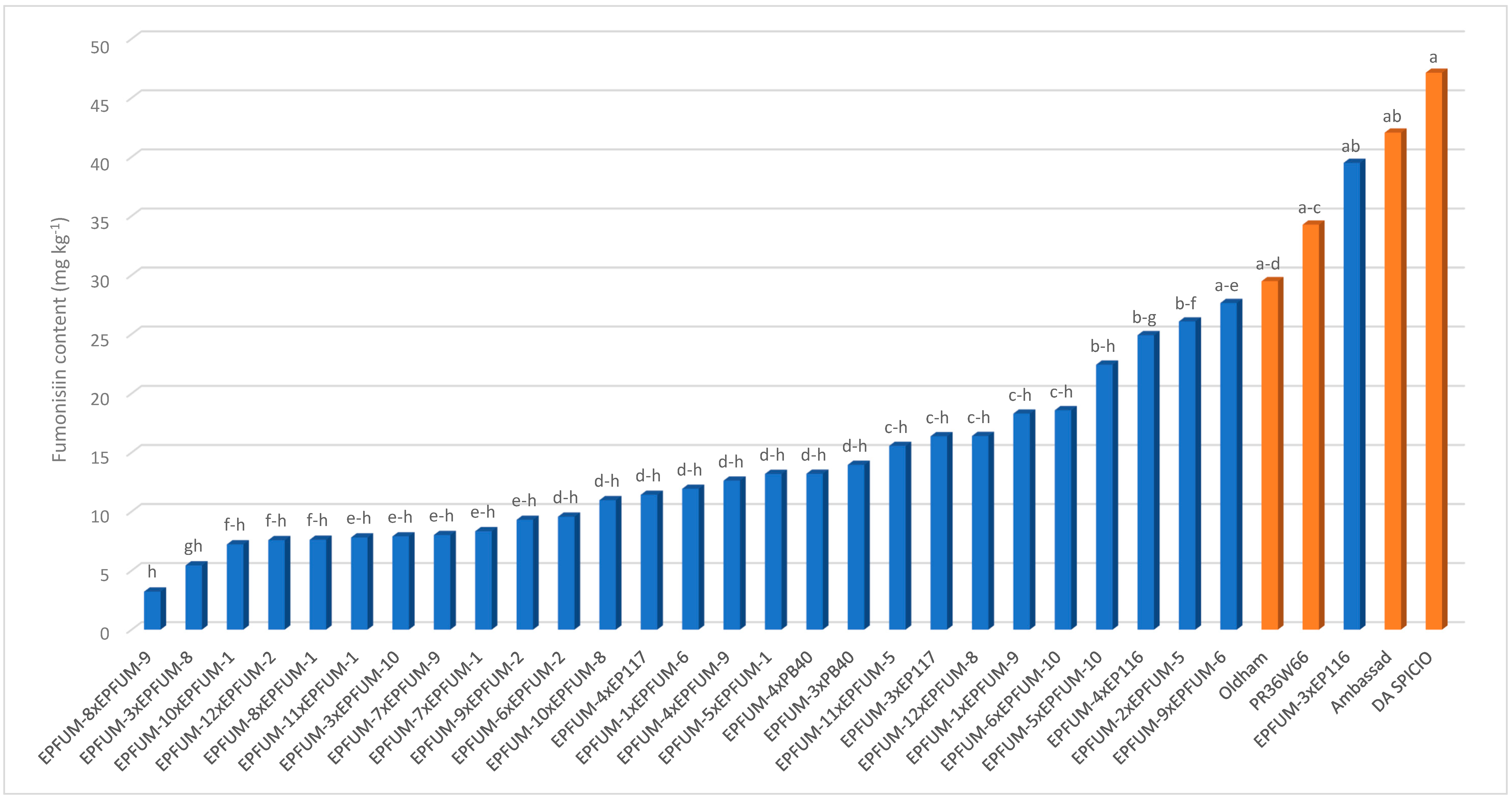
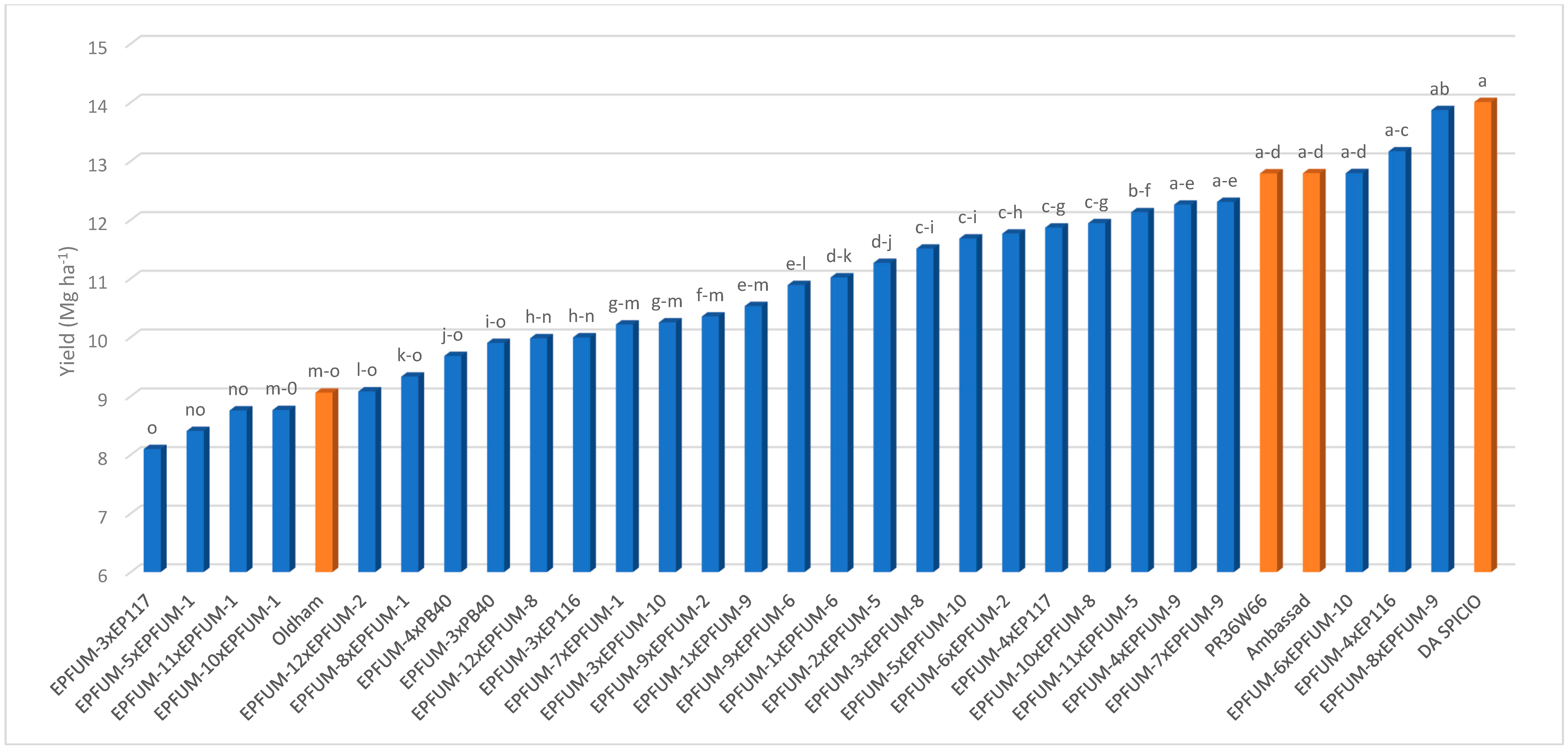
2. Results
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Inbred Selection Program
5.2. Inbred Evaluations
5.3. Hybrid Evaluations
5.4. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Furian, A.F.; Fighera, M.R.; Royes, L.F.F.; Oliveira, M.S. Recent advances in assessing the effects of mycotoxins using animal models. Curr. Opin. Food Sci. 2022, 47, 100874. [Google Scholar] [CrossRef]
- Chen, C.; Riley, R.T.; Wu, F. Dietary Fumonisin and Growth Impairment in Children and Animals: A Review. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1448–1464. [Google Scholar] [CrossRef]
- Chen, J.; Wen, J.; Tang, Y.T.; Shi, J.C.; Mu, G.D.; Yan, R.; Cai, J.; Long, M. Research Progress on Fumonisin B1 Contamination and Toxicity: A Review. Molecules 2021, 26, 5238. [Google Scholar] [CrossRef] [PubMed]
- Voss, K.A.; Riley, R.T. Fumonisin Toxicity and Mechanism of Action: Overview and Current Perspectives. Food Saf. 2013, 1, 2013006. [Google Scholar] [CrossRef]
- Deepa, N.; Achar, P.N.; Sreenivasa, M.Y. Current Perspectives of Biocontrol Agents for Management of Fusarium verticillioides and Its Fumonisin in Cereals—A Review. J. Fungi 2021, 7, 776. [Google Scholar] [CrossRef]
- Eller, M.S.; Holland, J.B.; Payne, G.A. Breeding for improved resistance to fumonisin contamination in maize. Toxin Rev. 2008, 27, 371–389. [Google Scholar] [CrossRef]
- Santiago, R.; Cao, A.; Butron, A. Genetic factors involved in fumonisin accumulation in maize kernels and their implications in maize agronomic management and breeding. Toxins 2015, 7, 3267–3296. [Google Scholar] [CrossRef] [PubMed]
- Logrieco, A.; Battilani, P.; Leggieri, M.C.; Jiang, Y.; Haesaert, G.; Lanubile, A.; Mahuku, G.; Mesterhazy, A.; Ortega-Beltran, A.; Pasti, M.; et al. Perspectives on Global Mycotoxin Issues and Management From the MycoKey Maize Working Group. Plant Dis. 2021, 105, 525–537. [Google Scholar] [CrossRef] [PubMed]
- Braun, M.S.; Wink, M. Exposure, occurrence, and chemistry of fumonisins and their cryptic derivatives. Compr. Rev. Food Sci. Food Saf. 2018, 17, 769–791. [Google Scholar] [CrossRef] [PubMed]
- Löffler, M.; Miedaner, T.; Kessel, B.; Ouzunova, M. Mycotoxin accumulation and corresponding ear rot rating in three maturity groups of European maize inoculated by two Fusarium species. Euphytica 2010, 174, 153–164. [Google Scholar] [CrossRef]
- Löffler, M.; Kessel, B.; Ouzunova, M.; Miedaner, T. Covariation between line and testcross performance for reduced mycotoxin concentrations in European maize after silk channel inoculation of two Fusarium species. Theor. Appl. Genet. 2011, 122, 925–934. [Google Scholar] [CrossRef]
- Bolduan, C.; Miedaner, T.; Schipprack, W.; Dhillon, B.S.; Melchinger, A.E. Genetic variation for resistance to ear rots and mycotoxins contamination in early European maize inbred lines. Crop Sci. 2009, 49, 2019–2028. [Google Scholar] [CrossRef]
- Samayoa, L.F.; Cao, A.; Santiago, R.; Malvar, R.A.; Butron, A. Genome-wide association analysis for fumonisin content in maize kernels. BMC Plant Biol. 2019, 19, 166. [Google Scholar] [CrossRef] [PubMed]
- Butron, A.; Reid, L.M.; Santiago, R.; Cao, A.; Malvar, R.A. Inheritance of maize resistance to gibberella and fusarium ear rots and kernel contamination with deoxynivalenol and fumonisins. Plant Pathol. 2015, 64, 1053–1060. [Google Scholar] [CrossRef]
- Boling, M.B.; Grogan, C.O. Gene action affecting host resistance to Fusarium ear rot of maize. Crop Sci. 1965, 5, 305–307. [Google Scholar] [CrossRef]
- Giomi, G.M.; Sampietro, D.A.; Velazco, J.G.; Iglesias, J.; Fernandez, M.; Oviedo, M.S.; Presello, D.A. Map overlapping of QTL for resistance to Fusarium ear rot and associated traits in maize. Euphytica 2021, 217, 81. [Google Scholar] [CrossRef]
- Netshifhefhe, N.E.I.; Flett, B.C.; Viljoen, A.; Rose, L.J. Inheritance and genotype by environment analyses of resistance to Fusarium verticillioides and fumonisin contamination in maize F-1 hybrids. Euphytica 2018, 214, 235. [Google Scholar] [CrossRef]
- Hung, H.-Y.; Holland, J.B. Diallel analysis of resistance to Fusarium ear rot and fumonisin contamination in maize. Crop Sci. 2012, 52, 2173–2181. [Google Scholar] [CrossRef]
- Padua, J.M.V.; Dias, K.O.D.; Pastina, M.M.; de Souza, J.C.; Queiroz, V.A.V.; da Costa, R.V.; da Silva, M.B.P.; Ribeiro, C.A.G.; Guimaraes, C.T.; Gezan, S.A.; et al. A multi-environment trials diallel analysis provides insights on the inheritance of fumonisin contamination resistance in tropical maize. Euphytica 2016, 211, 277–285. [Google Scholar] [CrossRef]
- Eller, M.S.; Payne, G.A.; Holland, J.B. Selection for reduced Fusarium ear rot and fumonisin content in advanced backcross maize lines and their topcross hybrids. Crop Sci. 2010, 50, 2249–2260. [Google Scholar] [CrossRef]
- Presello, D.A.; Pereyra, A.O.; Iglesias, J.; Fauguel, C.M.; Sampietro, D.A.; Eyherabide, G.H. Responses to selection of S-5 inbreds for broad-based resistance to ear rots and grain mycotoxin contamination caused by Fusarium spp. in maize. Euphytica 2011, 178, 23–29. [Google Scholar] [CrossRef]
- Horne, D.W.; Eller, M.S.; Holland, J.B. Responses to recurrent index selection for reduced Fusarium ear rot and lodging and for increased yield in maize. Crop Sci. 2016, 56, 85–94. [Google Scholar] [CrossRef]
- Butoto, E.N.; Brewer, J.C.; Holland, J.B. Empirical comparison of genomic and phenotypic selection for resistance to Fusarium ear rot and fumonisin contamination in maize. Theor. Appl. Genet. 2022, 135, 2799–2816. [Google Scholar] [CrossRef] [PubMed]
- Butoto, E.N.; Marino, T.P.; Holland, J.B. Effects of artificial inoculation on trait correlations with resistance to Fusarium ear rot and fumonisin contamination in maize. Crop Sci. 2021, 61, 2522–2533. [Google Scholar] [CrossRef]
- Santiago, R.; Cao, A.; Malvar, R.A.; Reid, L.M.; Butron, A. Assessment of corn resistance to fumonisin accumulation in a broad collection of inbred lines. Field Crop. Res. 2013, 149, 193–202. [Google Scholar] [CrossRef]
- Robertson, L.A.; Kleinschmidt, C.E.; White, D.G.; Payne, G.A.; Maragos, C.M.; Holland, J.B. Heritabilities and correlations of fusarium ear rot resistance and fumonisin contamination resistance in two maize populations. Crop Sci. 2006, 46, 353–361. [Google Scholar] [CrossRef]
- Revilla, P.; Soengas, P.; Malvar, R.A.; Cartea, M.E.; Ordas, A. Isozyme variation and historical relationships among the maize races of Spain. Maydica 1998, 43, 175–182. [Google Scholar]
- Revilla, P.; Malvar, R.A.; Cartea, M.E.; Soengas, P.; Ordas, A. Heterotic relationships among European maize inbreds. Euphytica 2002, 126, 259–264. [Google Scholar] [CrossRef]
- Revilla, P.; Boyat, A.; Alvarez, A.; Gouesnard, B.; Soengas, P.; Ordas, A.; Malvar, R.A. Heterotic patterns among French and Spanish maize populations. Maydica 2006, 51, 525–535. [Google Scholar]
- Malvar, R.A.; Revilla, P.; Moreno-Gonzalez, J.; Butron, A.; Sotelo, J.; Ordas, A. White maize: Genetics of quality and agronomic performance. Crop Sci. 2008, 48, 1373–1381. [Google Scholar] [CrossRef]
- CIMMYT. White Maize: A Traditional Food Grain in Developing Countries: A Joint Study / by the Basic Foodstuffs Service, FAO Commodities and Trade Division and the Economics Program, International Maize and Wheat Improvement Center; International Maize and Wheat Improvement Center; Food and Agriculture Organization of the United Nations: Rome, Italy, 1997. [Google Scholar]
- Lino, C.M.; Silva, L.J.G.; Pena, A.; Fernandez, M.; Manes, J. Occurrence of fumonisins B-1 and B-2 in broa, typical Portuguese maize bread. Int. J. Food Microbiol. 2007, 118, 79–82. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Pilu, R.; Cassani, E.; Sirizzotti, A.; Petroni, K.; Tonelli, C. Effect of flavonoid pigments on the accumulation of fumonisin B1 in the maize kernel. J. Appl. Genet. 2011, 52, 145–152. [Google Scholar] [CrossRef]
- Cao, A.; Butron, A.; Ramos, A.J.; Marin, S.; Souto, C.; Santiago, R. Assessing white maize resistance to fumonisin contamination. Eur. J. Plant Pathol. 2014, 138, 283–292. [Google Scholar] [CrossRef]
- Reid, L.M.; Hamilton, R.E.; Mather, D.E. Screening Maize for Resistance to Gibberella Ear Rot; Agriculture and Agri-Food Canada: Ottawa, ON, Canada, 1996; p. 62. [Google Scholar]
- Mesterhazy, A.; Lemmens, M.; Reid, L.M. Breeding for resistance to ear rots caused by Fusarium spp. in maize—A review. Plant Breed. 2012, 131, 1–19. [Google Scholar] [CrossRef]
- Statistical Analysis Systems Institute, SAS, version 9.1; SAS Institute Inc.: Cary, NC, USA, 2008.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group 1 | Released Inbred | Parent 1 | Parent 2 | Heterotic Group | Kernel Color |
|---|---|---|---|---|---|
| EP31 × EP39 | EPFUM-1 | EP31 | EP39 | European flint | yellow |
| EP31 × EP39 | EPFUM-2 | EP31 | EP39 | European flint | yellow |
| F575 × EP65 | EPFUM-3 | F575 | EP65 | European flint | white |
| F575 × EP65 | EPFUM-4 | F575 | EP65 | European flint | white |
| B93 × Oh43 | EPFUM-5 | B93 | Oh43 | Lancaster | yellow |
| B93 × Oh43 | EPFUM-6 | B93 | Oh43 | Lancaster | yellow |
| A670 × H95 | EPFUM-7 | A670 | H95 | Lancaster | yellow |
| A670 × H95 | EPFUM-8 | A670 | H95 | Lancaster | yellow |
| A630 × A635 | EPFUM-9 | A630 | A635 | Reid | yellow |
| A630 × A635 | EPFUM-10 | A630 | A635 | Reid | yellow |
| A654 × A666 | EPFUM-11 | A654 | A666 | Reid | yellow |
| A654 × A666 | EPFUM-12 | A654 | A666 | Reid | yellow |
| Source of Variation | Days to Shedding Pollen | Days to Silking | Kernel Moisture | Yield | FER | Fumonisin Content |
|---|---|---|---|---|---|---|
| Random (Z-value) | ||||||
| Year | 0.68 | 0.69 | 0.68 | 0.68 | 0.18 | |
| Replication (year) | 0.95 | 0.94 | 0.75 | 0.88 | 0.40 | |
| Year × Hybrid | 1.37 | 1.88 * | 0.50 | 0.53 | 2.81 ** | |
| Residual | 5.57 ** | 5.57 ** | 5.62 ** | 5.51 ** | 5.53 ** | 6.82 ** |
| Fi×ed (F-value) | ||||||
| Hybrid | 18.40 ** | 16.95 ** | 2.56 ** | 6.56 ** | 1.44 | 2.68 ** |
| Hybrid | FER (1–7) | Kernel Moisture (%) | Days to Shedding Pollen |
|---|---|---|---|
| Ambassad | 2.9 a | 17.1 e–g | 73.2 bc |
| Da SPICIO | 3.5 a | 17.3 e–g | 72.5 c–e |
| PR36W66 | 3.2 a | 18.0 d–g | 75.8 a |
| Oldham | 3.3 a | 15.9 fg | 63.3 l |
| EPFUM-1 × EPFUM-6 | 2.5 a | 23.0 ab | 69.3 g–i |
| EPFUM-1 × EPFUM-9 | 2.7 a | 19.8 c–e | 70.8 d–h |
| EPFUM-2 × EPFUM-5 | 2.2 a | 19.9 c–e | 65.8 kj |
| EPFUM-3 × EP116 | 3.4 a | 19.1 c–f | 71.5 c–f |
| EPFUM-3 × EP117 | 2.9 a | 19.2 c–f | 69.3 g–i |
| EPFUM-3 × EPFUM-10 | 2.4 a | 18.1 d–g | 72.0 c–e |
| EPFUM-3 × EPFUM-8 | 1.8 a | 19.0 d–f | 72.8 cd |
| EPFUM-3 × PB40 | 3.0 a | 19.1 c–f | 70.8 d–h |
| EPFUM-4 × EP116 | 3.2 a | 19.1 d–f | 71.0 d–g |
| EPFUM-4 × EP117 | 2.9 a | 19.4 c–e | 69.5 f–i |
| EPFUM-4 × EPFUM-9 | 2.6 a | 17.8 d–g | 71.8 c–e |
| EPFUM-4 × PB40 | 2.8 a | 19.6 c–e | 72.0 c–e |
| EPFUM-5 × EPFUM-1 | 2.3 a | 19.3 c–e | 64.5 kl |
| EPFUM-5 × EPFUM-10 | 3.4 a | 19.6 c–e | 70.5 e–h |
| EPFUM-6 × EPFUM-10 | 3.4 a | 19.1 d–f | 71.3 c–g |
| EPFUM-6 × EPFUM-2 | 2.1 a | 23.8 a | 68.8 ih |
| EPFUM-7 × EPFUM-1 | 2.0 a | 17.3 e–g | 67.8 ij |
| EPFUM-7 × EPFUM-9 | 1.8 a | 17.3 e–g | 75.3 ab |
| EPFUM-8 × EPFUM-1 | 1.7 a | 20.5 b–d | 68.0 i |
| EPFUM-8 × EPFUM-9 | 2.4 a | 19.4 c–e | 71.8 c–e |
| EPFUM-9 × EPFUM-2 | 2.5 a | 15.4 f | 68.3 i |
| EPFUM-9 × EPFUM-6 | 2.5 a | 22.2 a–c | 72.5 c–e |
| EPFUM-10 × EPFUM-1 | 2.0 a | 19.7 c–e | 65.3 kl |
| EPFUM-10 × EPFUM-8 | 3.1 a | 18.7 d–f | 70.5 e–h |
| EPFUM-11 × EPFUM-1 | 2.3 a | 19.5 c–e | 64.8 kl |
| EPFUM-11 × EPFUM-5 | 2.9 a | 19.8 c–e | 67.8 ij |
| EPFUM-12 × EPFUM-2 | 2.3 a | 19.9 c–e | 64.3 kl |
| EPFUM-12 × EPFUM-8 | 3.3 a | 19.3 c–e | 68.3 i |
| Hybrid | Mid-Parent 1 | Mid-Parent Heterosis | ||
|---|---|---|---|---|
| FER | Fumonisin | FER | Fumonisin | |
| EPFUM-1 × EPFUM-6 | 2.70 | 22.0 | −0.0859 | −0.4575 |
| EPFUM-1 × EPFUM-9 | 2.72 | 21.0 | −0.0035 | −0.1263 |
| EPFUM-2 × EPFUM-5 | 3.38 | 31.3 | −0.3592 | −0.1684 |
| EPFUM-3 × EPFUM-8 | 2.08 | 8.0 | −0.1203 | −0.3218 |
| EPFUM-3 × EPFUM-10 | 2.52 | 11.6 | −0.0397 | −0.3192 |
| EPFUM-4 × EPFUM-9 | 2.36 | 16.8 | 0.1113 | −0.2488 |
| EPFUM-5 × EPFUM-1 | 2.87 | 21.2 | −0.2169 | −0.3788 |
| EPFUM-5 × EPFUM-10 | 2.69 | 18.9 | 0.2567 | 0.1870 |
| EPFUM-6 × EPFUM-2 | 3.20 | 32.1 | −0.3485 | −0.7021 |
| EPFUM-6 × EPFUM-10 | 2.50 | 19.7 | 0.3490 | −0.0550 |
| EPFUM-7 × EPFUM-1 | 2.41 | 13.5 | −0.1716 | −0.3829 |
| EPFUM-7 × EPFUM-9 | 2.09 | 15.0 | −0.1416 | −0.4670 |
| EPFUM-8 × EPFUM-1 | 2.42 | 13.5 | −0.3125 | −0.4359 |
| EPFUM-8 × EPFUM-9 | 2.10 | 15.0 | 0.1322 | −0.7858 |
| EPFUM-9 × EPFUM-2 | 3.23 | 31.1 | −0.2380 | −0.7006 |
| EPFUM-9 × EPFUM-6 | 2.36 | 23.5 | 0.0582 | 0.1750 |
| EPFUM-10 × EPFUM-1 | 2.86 | 17.1 | −0.3000 | −0.5773 |
| EPFUM-10 × EPFUM-8 | 2.24 | 11.2 | 0.3635 | −0.0184 |
| EPFUM-11 × EPFUM-1 | 3.09 | 30.1 | −0.2707 | −0.7407 |
| EPFUM-11 × EPFUM-5 | 2.91 | 31.9 | −0.0133 | −0.5124 |
| EPFUM-12 × EPFUM-2 | 3.96 | 36.0 | −0.4220 | −0.7896 |
| EPFUM-12 × EPFUM-8 | 2.84 | 20.0 | 0.1487 | −0.1797 |
| Days to Silking | Kernel Moisture | Yield | FER | Fumonisin Content | |
|---|---|---|---|---|---|
| Days to shedding pollen | 0.95 ** | −0.11 | 0.64 ** | 0.22 | 0.28 |
| Days to silking | −0.06 | 0.51 ** | 0.23 | 0.23 | |
| Kernel moisture | −0.10 | −0.24 | −0.21 | ||
| Yield | 0.29 | 0.32 | |||
| FER | 0.66 ** |
| Mid-Parent 1 | Heterosis | |||
|---|---|---|---|---|
| r2 | p-Value | r2 | p-Value | |
| FER | 0.02 | 0.94 | 0.80 | <0.0001 |
| Fumonisin content | 0.29 | 0.20 | 0.76 | <0.0001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santiago, R.; Ramos, A.J.; Cao, A.; Malvar, R.A.; Butrón, A. Inbred Selection for Increased Resistance to Kernel Contamination with Fumonisins. Toxins 2023, 15, 444. https://doi.org/10.3390/toxins15070444
Santiago R, Ramos AJ, Cao A, Malvar RA, Butrón A. Inbred Selection for Increased Resistance to Kernel Contamination with Fumonisins. Toxins. 2023; 15(7):444. https://doi.org/10.3390/toxins15070444
Chicago/Turabian StyleSantiago, Rogelio, Antonio J. Ramos, Ana Cao, Rosa Ana Malvar, and Ana Butrón. 2023. "Inbred Selection for Increased Resistance to Kernel Contamination with Fumonisins" Toxins 15, no. 7: 444. https://doi.org/10.3390/toxins15070444
APA StyleSantiago, R., Ramos, A. J., Cao, A., Malvar, R. A., & Butrón, A. (2023). Inbred Selection for Increased Resistance to Kernel Contamination with Fumonisins. Toxins, 15(7), 444. https://doi.org/10.3390/toxins15070444