Ochratoxin A: Overview of Prevention, Removal, and Detoxification Methods

{kind=link}
Abstract
:1. Introduction
2. Physical Methods
2.1. Radiation
2.2. Heat
2.3. Adsorption
2.3.1. Natural Material Adsorbents
2.3.2. Inorganic Mineral Adsorbents
2.3.3. Organic Synthetic Adsorbents
2.4. Other Methods
3. Chemical Methods
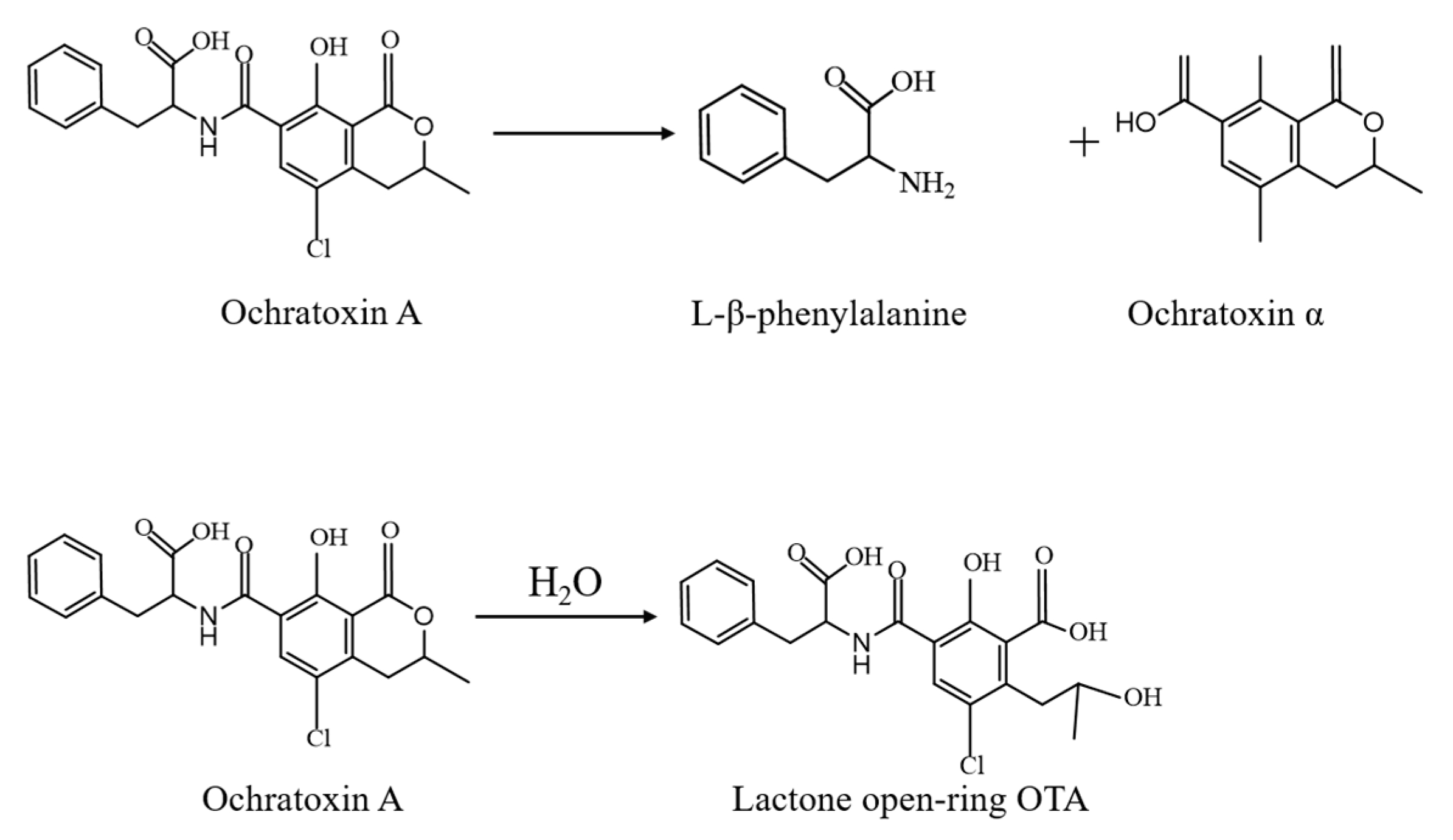
3.1. Alkalization
3.2. Oxidization
3.3. Other Chemicals
4. Biological Methods
4.1. Inhibition of OTA Biosynthesis
4.2. Microbial Bioadsorption
4.3. Enzymatic Degradation of OTA
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Dalié, D.K.D.; Deschamps, A.M.; Richard-Forget, F. Lactic acid bacteria—Potential for control of mould growth and mycotoxins: A review. Food Control 2010, 21, 370–380. [Google Scholar] [CrossRef]
- Leitão, A.L. Occurrence of Ochratoxin A in Coffee: Threads and Solutions—A Mini-Review. Beverages 2019, 5, 36. [Google Scholar] [CrossRef]
- Tao, Y.; Xie, S.; Xu, F.; Liu, A.; Wang, Y.; Chen, D.; Pan, Y.; Huang, L.; Peng, D.; Wang, X.; et al. Ochratoxin A: Toxicity, oxidative stress and metabolism. Food Chem. Toxicol. 2018, 112, 320–331. [Google Scholar] [CrossRef] [PubMed]
- Gu, K.; Ryu, D.; Lee, H.J. Ochratoxin A and its reaction products affected by sugars during heat processing. Food Chem. 2021, 348, 129038. [Google Scholar] [CrossRef] [PubMed]
- Clark, H.A.; Snedeker, S.M. Ochratoxin a: Its cancer risk and potential for exposure. J. Toxicol. Environ. Health B Crit. Rev. 2006, 9, 265–296. [Google Scholar] [CrossRef]
- Samuel, M.S.; Jeyaram, K.; Datta, S.; Chandrasekar, N.; Balaji, R.; Selvarajan, E. Detection, contamination, toxicity, and prevention methods of ochratoxins: An update review. J. Agric. Food Chem. 2021, 69, 13974–13989. [Google Scholar] [CrossRef]
- Marquardt, R.R.; Frohlich, A.A. A review of recent advances in understanding ochratoxicosis. J. Anim. Sci. 1992, 70, 3968–3988. [Google Scholar] [CrossRef]
- O’Brien, E.; Prietz, A.; Dietrich, D.R. Investigation of the teratogenic potential of ochratoxin A and B using the FETAX system. Birth Defects Res. B 2005, 74, 417–423. [Google Scholar] [CrossRef]
- Costa, S.; Utan, A.; Cervellati, R.; Speroni, E.; Guerra, M.C. Catechins: Natural free-radical scavengers against ochratoxin A-induced cell damage in a pig kidney cell line (LLC-PK1). Food Chem. Toxicol. 2007, 45, 1910–1917. [Google Scholar] [CrossRef]
- Vlachou, M.; Pexara, A.; Solomakos, N.; Govaris, A. Ochratoxin A in Slaughtered Pigs and Pork Products. Toxins 2022, 14, 67. [Google Scholar] [CrossRef]
- Ciarcia, R.; Damiano, S.; Squillacioti, C.; Mirabella, N.; Pagnini, U.; Florio, A.; Severino, L.; Capasso, G.; Borrelli, A.; Mancini, A.; et al. Recombinant Mitochondrial Manganese Containing Superoxide Dismutase Protects against Ochratoxin A-Induced Nephrotoxicity. J. Cell. Biochem. 2016, 117, 1352–1358. [Google Scholar] [CrossRef] [PubMed]
- Damiano, S.; Navas, L.; Lombari, P.; Montagnaro, S.; Forte, I.M.; Giordano, A.; Florio, S.; Ciarcia, R. Effects of delta-tocotrienol on ochratoxin A-induced nephrotoxicity in rats. J. Cell. Physiol. 2018, 233, 8731–8739. [Google Scholar] [CrossRef] [PubMed]
- Damiano, S.; Iovane, V.; Squillacioti, C.; Mirabella, N.; Prisco, F.; Ariano, A.; Amenta, M.; Giordano, A.; Florio, S.; Ciarcia, R. Red orange and lemon extract prevents the renal toxicity induced by ochratoxin A in rats. J. Cell. Physiol. 2020, 235, 5386–5393. [Google Scholar] [CrossRef] [PubMed]
- Pinhao, M.; Tavares, A.M.; Loureiro, S.; Louro, H.; Alvito, P.; Silva, M.J. Combined cytotoxic and genotoxic effects of ochratoxin A and fumonisin B1 in human kidney and liver cell models. Toxicol. In Vitro 2020, 68, 104949. [Google Scholar] [CrossRef]
- Zheng, Q.W.; Ding, X.F.; Cao, H.J.; Ni, Q.Z.; Zhu, B.; Ma, N.; Zhang, F.K.; Wang, Y.K.; Xu, S.; Chen, T.W.; et al. Ochratoxin A Induces Steatosis via PPARgamma-CD36 Axis. Toxins 2021, 13, 802. [Google Scholar] [CrossRef]
- Zhai, S.S.; Ruan, D.; Zhu, Y.W.; Li, M.C.; Ye, H.; Wang, W.C.; Yang, L. Protective effect of curcumin on ochratoxin A-induced liver oxidative injury in duck is mediated by modulating lipid metabolism and the intestinal microbiota. Poult. Sci. 2020, 99, 1124–1134. [Google Scholar] [CrossRef]
- Thuvander, A.; Moller, T.; Barbieri, H.E.; Jansson, A.; Salomonsson, A.C.; Olsen, M. Dietary intake of some important mycotoxins by the Swedish population. Food Addit. Contam. 2001, 18, 696–706. [Google Scholar] [CrossRef]
- Commission Regulation (EC) No 1370/2022; Commission Regulation (EC) No 1370/2022 of 5 August 2022 Amending Regulation (EC) No 1881/2006 as Regards Maximum Levels of Ochratoxin A in Certain Foodstuffs. European Union: Brussels, Belgium, 2022; L206/11-206/14.
- Amézqueta, S.; González-Peñas, E.; Murillo-Arbizu, M.; López de Cerain, A. Ochratoxin A decontamination: A review. Food Control 2009, 20, 326–333. [Google Scholar] [CrossRef]
- Duarte, S.C.; Pena, A.; Lino, C.M. A review on ochratoxin A occurrence and effects of processing of cereal and cereal derived food products. Food Microbiol. 2010, 27, 187–198. [Google Scholar] [CrossRef]
- Massoud, R.; Cruz, A.; Darani, K.K. Ochratoxin A: From Safety Aspects to Prevention and Remediation Strategies. Curr. Nutr. Food Sci. 2018, 14, 11–16. [Google Scholar] [CrossRef]
- Magan, N.; Aldred, D. Conditions of formation of ochratoxin A in drying, transport and in different commodities. Food Addit. Contam. 2005, 22 (Suppl. S1), 10–16. [Google Scholar] [CrossRef] [PubMed]
- Di Stefano, V.; Pitonzo, R.; Cicero, N.; D’Oca, M.C. Mycotoxin contamination of animal feedingstuff: Detoxification by gamma-irradiation and reduction of aflatoxins and ochratoxin A concentrations. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2014, 31, 2034–2039. [Google Scholar] [CrossRef] [PubMed]
- Roberts, P.B. Food irradiation: Standards, regulations and world-wide trade. Radiat. Phys. Chem. 2016, 129, 30–34. [Google Scholar] [CrossRef]
- World Health Organization. High-Dose Irradiation: Wholesomeness of Food Irradiated with Doses Above 10 kGy. Report of a Joint FAO/IAEA/WHO Study Group; World Health Organization Technical Report Series; World Health Organization: Geneva, Switzerland, 1999; Volume 890, pp. 1–197. [Google Scholar]
- Zhang, H.; Wang, G.; Yang, Q.; Yang, X.; Zheng, Y.; Liu, Y.; Xing, F. Effects of Light on the Ochratoxigenic Fungi Aspergillus ochraceus and A. carbonarius. Toxins 2021, 13, 251. [Google Scholar] [CrossRef] [PubMed]
- Khalil, O.A.A.; Hammad, A.A.; Sebaei, A.S. Aspergillus flavus and Aspergillus ochraceus inhibition and reduction of aflatoxins and ochratoxin A in maize by irradiation. Toxicon 2021, 198, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Calado, T.; Fernandez-Cruz, M.L.; Cabo Verde, S.; Venancio, A.; Abrunhosa, L. Gamma irradiation effects on ochratoxin A: Degradation, cytotoxicity and application in food. Food Chem. 2018, 240, 463–471. [Google Scholar] [CrossRef]
- Shanakhat, H.; Sorrentino, A.; Raiola, A.; Reverberi, M.; Salustri, M.; Masi, P.; Cavella, S. Technological properties of durum wheat semolina treated by heating and UV irradiation for reduction of mycotoxin content. J. Food Process. Eng. 2019, 42, e13006. [Google Scholar] [CrossRef]
- Peng, C.; Ding, Y.; An, F.; Wang, L.; Li, S.; Nie, Y.; Zhou, L.; Li, Y.; Wang, C.; Li, S. Degradation of ochratoxin A in aqueous solutions by electron beam irradiation. J. Radioanal. Nucl. Chem. 2015, 306, 39–46. [Google Scholar] [CrossRef]
- Yang, K.; Li, K.; Pan, L.; Luo, X.; Xing, J.; Wang, J.; Wang, L.; Wang, R.; Zhai, Y.; Chen, Z. Effect of Ozone and Electron Beam Irradiation on Degradation of Zearalenone and Ochratoxin A. Toxins 2020, 12, 138. [Google Scholar] [CrossRef]
- Wang, L.; Liu, X.; Cai, R.; Ge, Q.; Zhao, Z.; Yue, T.; Yuan, Y.; Gao, Z.; Wang, Z. Detoxification of Ochratoxin A by pulsed light in grape juice and evaluation of its degradation products and safety. Innov. Food Sci. Emerg. Technol. 2022, 78, 103024. [Google Scholar] [CrossRef]
- Zou, D.; Ji, J.; Ye, Y.; Yang, Y.; Yu, J.; Wang, M.; Zheng, Y.; Sun, X. Degradation of Ochratoxin A by a UV-Mutated Aspergillus niger Strain. Toxins 2022, 14, 343. [Google Scholar] [CrossRef]
- Trivedi, A.B.; Doi, E.; Kitabatake, N. Detoxification of Ochratoxin A on Heating under Acidic and Alkaline Conditions. Biosci. Biotechnol. Biochem. 1992, 56, 741–745. [Google Scholar] [CrossRef] [PubMed]
- Abrunhosa, L.; Paterson, R.R.; Venancio, A. Biodegradation of ochratoxin a for food and feed decontamination. Toxins 2010, 2, 1078–1099. [Google Scholar] [CrossRef] [PubMed]
- Schrenk, D.; Bodin, L.; Chipman, J.K.; del Mazo, J.; Grasl-Kraupp, B.; Hogstrand, C.; Hoogenboom, L.; Leblanc, J.C.; Nebbia, C.S.; Nielsen, E.; et al. Risk assessment of ochratoxin A in food. EFSA J. 2020, 18, e06113. [Google Scholar]
- Dahal, S.; Lee, H.J.; Gu, K.; Ryu, D. Heat Stability of Ochratoxin A in an Aqueous Buffered Model System. J. Food Prot. 2016, 79, 1748–1752. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J. Stability of ochratoxin A in oats during roasting with reducing sugars. Food Control 2020, 118, 107382. [Google Scholar] [CrossRef]
- Ameer Sumbal, G.; Hussain Shar, Z.; Hussain Sherazi, S.T.; Sirajuddin; Nizamani, S.M.; Mahesar, S.A. Decontamination of poultry feed from ochratoxin A by UV and sunlight radiations. J. Sci. Food Agric. 2016, 96, 2668–2673. [Google Scholar] [CrossRef]
- Belajová, E.; Rauová, D.; Daško, L. Retention of ochratoxin A and fumonisin B1 and B2 from beer on solid surfaces: Comparison of efficiency of adsorbents with different origin. Eur. Food Res. Technol. 2006, 224, 301–308. [Google Scholar] [CrossRef]
- Quintela, S.; Villarán, M.C.; López de Armentia, I.; Elejalde, E. Ochratoxin A removal in wine: A review. Food Control 2013, 30, 439–445. [Google Scholar] [CrossRef]
- Abdelnaby, A.; Abdelaleem, N.M.; Elshewy, E.; Mansour, A.H.; Ibrahim, S. The efficacy of clay bentonite, date pit, and chitosan nanoparticles in the detoxification of aflatoxin M1 and ochratoxin A from milk. Environ. Sci. Pollut. Res. Int. 2022, 29, 20305–20317. [Google Scholar] [CrossRef]
- Nobre, C.; Gonzalez, A.; Losoya, C.; Teixeira, J.A.; Belmares, R.; Abrunhosa, L. Detoxification of ochratoxin A and zearalenone by Pleurotus ostreatus during in vitro gastrointestinal digestion. Food Chem. 2022, 384, 132525. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, J.M.; Calado, T.; Guimaraes, A.; Rodrigues, M.A.M.; Abrunhosa, L. In vitro adsorption of aflatoxin B1, ochratoxin A, and zearalenone by micronized grape stems and olive pomace in buffer solutions. Mycotoxin Res. 2019, 35, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Loffredo, E.; Scarcia, Y.; Parlavecchia, M. Removal of ochratoxin A from liquid media using novel low-cost biosorbents. Environ. Sci. Pollut. Res. Int. 2020, 27, 34484–34494. [Google Scholar] [CrossRef] [PubMed]
- Di Gregorio, M.C.; Neeff, D.V.d.; Jager, A.V.; Corassin, C.H.; Carão, Á.C.d.P.; Albuquerque, R.d.; Azevedo, A.C.d.; Oliveira, C.A.F. Mineral adsorbents for prevention of mycotoxins in animal feeds. Toxin Rev. 2014, 33, 125–135. [Google Scholar] [CrossRef]
- Var, I.; Kabak, B.; Erginkaya, Z. Reduction in ochratoxin A levels in white wine, following treatment with activated carbon and sodium bentonite. Food Control 2008, 19, 592–598. [Google Scholar] [CrossRef]
- Cosme, F.; Inês, A.; Silva, D.; Filipe-Ribeiro, L.; Abrunhosa, L.; Nunes, F.M. Elimination of ochratoxin A from white and red wines: Critical characteristics of activated carbons and impact on wine quality. LWT 2021, 140, 110838. [Google Scholar] [CrossRef]
- Commission Regulation (EC) No 606/2009; Commission Regulation (EC) No 606/2009 of 10 July 2009 Laying down Certain Detailed Rules for Implementing Council Regulation (EC) No 479/2008 as Regards the Categories of Grapevine Products, Oenological Practices and the Applicable Restrictions. European Union: Brussels, Belgium, 2009. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A32009R0606 (accessed on 2 August 2022).
- Bhatti, S.A.; Khan, M.Z.; Saleemi, M.K.; Hassan, Z.U.; Khan, A. Ameliorative role of dietary activated carbon against ochratoxin-A induced oxidative damage, suppressed performance and toxicological effects. Toxin Rev. 2020, 41, 108–118. [Google Scholar] [CrossRef]
- El Khoury, R.; Choque, E.; El Khoury, A.; Snini, S.P.; Cairns, R.; Andriantsiferana, C.; Mathieu, F. OTA Prevention and Detoxification by Actinobacterial Strains and Activated Carbon Fibers: Preliminary Results. Toxins 2018, 10, 137. [Google Scholar] [CrossRef]
- Hamad, G.M.; El-Makarem, H.A.; Elaziz, A.I.A.; Amer, A.A.; El-Nogoumy, B.A.; Abou-Alella, S.A. Adsorption efficiency of sodium & calcium bentonite for ochratoxin A in some Egyptian cheeses: An innovative fortification model, in vitro and in vivo experiments. World Mycotoxin J. 2022, 15, 285–300. [Google Scholar]
- Kurtbay, H.M.; Bekci, Z.; Merdivan, M.; Yurdakoc, K. Reduction of ochratoxin a levels in red wine by bentonite, modified bentonites, and chitosan. J. Agric. Food Chem. 2008, 56, 2541–2545. [Google Scholar] [CrossRef]
- Vila-Donat, P.; Marín, S.; Sanchis, V.; Ramos, A.J. New mycotoxin adsorbents based on tri-octahedral bentonites for animal feed. Anim. Feed Sci. Technol. 2019, 255, 114228. [Google Scholar] [CrossRef]
- Rasheed, U.; Ain, Q.U.; Yaseen, M.; Fan, X.; Yao, X.; Tong, Z.; Liu, B. Modification of bentonite with orange peels extract and its application as mycotoxins’ binder in buffered solutions and simulated gastrointestinal fluids. J. Clean. Prod. 2020, 267, 122105. [Google Scholar] [CrossRef]
- Singh, R.; Mandal, A.B. Efficacy of hydrated sodium calcium aluminosilicate in ameliorating ochratoxicosis in broiler chickens. IJPS 2018, 53, 181–187. [Google Scholar] [CrossRef]
- Kannewischer, I.; Arvide, M.G.T.; White, G.N.; Dixon, J.B. Smectite clays as adsorbents of aflatoxin B1: Initial steps. Clay Sci. 2006, 12, 199–204. [Google Scholar]
- Solis-Cruz, B.; Hernandez-Patlan, D.; Beyssac, E.; Latorre, J.D.; Hernandez-Velasco, X.; Merino-Guzman, R.; Tellez, G.; Lopez-Arellano, R. Evaluation of Chitosan and Cellulosic Polymers as Binding Adsorbent Materials to Prevent Aflatoxin B1, Fumonisin B1, Ochratoxin, Trichothecene, Deoxynivalenol, and Zearalenone Mycotoxicoses Through an In Vitro Gastrointestinal Model for Poultry. Polymers 2017, 9, 529. [Google Scholar] [CrossRef]
- Zhao, Z.; Liu, N.; Yang, L.; Wang, J.; Song, S.; Nie, D.; Yang, X.; Hou, J.; Wu, A. Cross-linked chitosan polymers as generic adsorbents for simultaneous adsorption of multiple mycotoxins. Food Control 2015, 57, 362–369. [Google Scholar] [CrossRef]
- Appell, M.; Jackson, M.A. Sorption of ochratoxin A from aqueous solutions using beta-cyclodextrin-polyurethane polymer. Toxins 2012, 4, 98–109. [Google Scholar] [CrossRef]
- Song, C.; Qin, J. High-performance fabricated nano-adsorbents as emerging approach for removal of mycotoxins: A review. Int. J. Food Microbiol. 2022, 57, 5781–5789. [Google Scholar] [CrossRef]
- Abbasi Pirouz, A.; Selamat, J.; Sukor, R.; Noorahya Jambari, N. Effective Detoxification of Aflatoxin B1 and Ochratoxin A Using Magnetic Graphene Oxide Nanocomposite: Isotherm and Kinetic Study. Coatings 2021, 11, 1346. [Google Scholar] [CrossRef]
- Zahoor, M.; Gulfam, N.; Khisroon, M.; Khan, F.A. Removal of Ochratoxin A from water by novel adsorbent; magnetic carbon nanocomposites prepared from sugar beet wastes. Desalin. Water Treat. 2019, 152, 234–241. [Google Scholar] [CrossRef]
- Alford, R.; Mishael, Y.G. Bifunctional clay based sorbent for ‘Ochratoxin A’ removal and wine fining. Food Chem. 2023, 416, 135827. [Google Scholar] [CrossRef] [PubMed]
- Selcuk, M.; Oksuz, L.; Basaran, P. Decontamination of grains and legumes infected with Aspergillus spp. and Penicillum spp. by cold plasma treatment. Bioresour. Technol. 2008, 99, 5104–5109. [Google Scholar] [CrossRef] [PubMed]
- Casas-Junco, P.P.; Solis-Pacheco, J.R.; Ragazzo-Sanchez, J.A.; Aguilar-Uscanga, B.R.; Bautista-Rosales, P.U.; Calderon-Santoyo, M. Cold Plasma Treatment as an Alternative for Ochratoxin a Detoxification and Inhibition of Mycotoxigenic Fungi in Roasted Coffee. Toxins 2019, 11, 337. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, X.; Matinfar, G.; Khaneghah, A.M.; Singh, A.; Pratap-Singh, A. Emergence of cold plasma and electron beam irradiation as novel technologies to counter mycotoxins in food products. World Mycotoxin J. 2021, 14, 75–83. [Google Scholar] [CrossRef]
- Misra, N.N.; Yadav, B.; Roopesh, M.S.; Jo, C. Cold Plasma for Effective Fungal and Mycotoxin Control in Foods: Mechanisms, Inactivation Effects, and Applications. Compr. Rev. Food Sci. Food Saf. 2019, 18, 106–120. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Smith, I.N.; Mikiashvili, N. Reducing Ochratoxin A Content in Grape Pomace by Different Methods. Toxins 2020, 12, 424. [Google Scholar] [CrossRef]
- Özcan, S.; Gökmen, V. Alkali-based pre-treatment may prevent ochratoxin A in grapes. World Mycotoxin J. 2016, 9, 517–523. [Google Scholar] [CrossRef]
- Fouler, S.G.; Trivedi, A.B.; Kitabatake, N. Detoxification of citrinin and ochratoxin A by hydrogen peroxide. J. AOAC Int. 1994, 77, 631–637. [Google Scholar] [CrossRef]
- Jalili, M.; Jinap, S. Role of sodium hydrosulphite and pressure on the reduction of aflatoxins and ochratoxin A in black pepper. Food Control 2012, 27, 11–15. [Google Scholar] [CrossRef]
- Scott, P.M. Effects of processing and detoxification treatments on ochratoxin A: Introduction. Food Addit. Contam. 1996, 13, 19–21. [Google Scholar]
- Chelkowski, J.; Golinski, P.; Godlewska, B.; Radomyska, W.; Szebiotko, K.; Wiewiorowska, M. Mycotoxins in cereal grain. Part IV. Inactivation of ochratoxin A and other mycotoxins during ammoniation. Nahrung 1981, 25, 631–637. [Google Scholar] [CrossRef] [PubMed]
- Amezqueta, S.; Gonzalez-Penas, E.; Lizarraga, T.; Murillo-Arbizu, M.; Lopez de Cerain, A. A simple chemical method reduces ochratoxin A in contaminated cocoa shells. J. Food Prot. 2008, 71, 1422–1426. [Google Scholar] [CrossRef] [PubMed]
- Jalili, M.; Jinap, S.; Son, R. The effect of chemical treatment on reduction of aflatoxins and ochratoxin A in black and white pepper during washing. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2011, 28, 485–493. [Google Scholar] [CrossRef] [PubMed]
- Peraica, M.; Domijan, A.-M.; Jurjević, Ž.; Cvjetković, B. Prevention of exposure to mycotoxins from food and feed. Arh. Hig. Rada Toksikol. 2002, 53, 229–237. [Google Scholar] [PubMed]
- Qi, L.; Li, Y.; Luo, X.; Wang, R.; Zheng, R.; Wang, L.; Li, Y.; Yang, D.; Fang, W.; Chen, Z. Detoxification of zearalenone and ochratoxin A by ozone and quality evaluation of ozonised corn. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2016, 33, 1700–1710. [Google Scholar] [CrossRef]
- Sujayasree, O.J.; Chaitanya, A.K.; Bhoite, R.; Pandiselvam, R.; Kothakota, A.; Gavahian, M.; Mousavi Khaneghah, A. Ozone: An Advanced Oxidation Technology to Enhance Sustainable Food Consumption through Mycotoxin Degradation. Ozone Sci. Eng. 2021, 44, 17–37. [Google Scholar] [CrossRef]
- Akbar, A.; Medina, A.; Magan, N. Potential Control of Mycotoxigenic Fungi and Ochratoxin A in Stored Coffee Using Gaseous Ozone Treatment. Microorganisms 2020, 8, 1462. [Google Scholar] [CrossRef]
- van der Merwe, K.; Steyn, P.; Fourie, L. Mycotoxins. Part II. The constitution of ochratoxins A, B, and C, metabolites of Aspergillus ochraceus wilh. J. Chem. Soc. 1965, 7083–7088. [Google Scholar] [CrossRef]
- Schmidt-Heydt, M.; Stoll, D.; Geisen, R. Fungicides effectively used for growth inhibition of several fungi could induce mycotoxin biosynthesis in toxigenic species. Int. J. Food Microbiol. 2013, 166, 407–412. [Google Scholar] [CrossRef]
- Magista, D.; Cozzi, G.; Gambacorta, L.; Logrieco, A.F.; Solfrizzo, M.; Perrone, G. Studies on the efficacy of electrolysed oxidising water to control Aspergillus carbonarius and ochratoxin A contamination on grape. Int. J. Food Microbiol. 2021, 338, 108996. [Google Scholar] [CrossRef]
- Farbo, M.G.; Urgeghe, P.P.; Fiori, S.; Marcello, A.; Oggiano, S.; Balmas, V.; Hassan, Z.U.; Jaoua, S.; Migheli, Q. Effect of yeast volatile organic compounds on ochratoxin A-producing Aspergillus carbonarius and A. ochraceus. Int. J. Food Microbiol. 2018, 284, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Chen, X.; Lei, S.; Zhao, H.; Liu, Y.; Shi, J. Lipopeptides from Bacillus subtilis have potential application in the winemaking process: Inhibiting fungal and ochratoxin A contamination and enhancing esters and acids biosynthesis. Aust. J. Grape Wine Res. 2017, 23, 350–358. [Google Scholar] [CrossRef]
- Jiang, C.; Li, Z.; Shi, Y.; Guo, D.; Pang, B.; Chen, X.; Shao, D.; Liu, Y.; Shi, J. Bacillus subtilis inhibits Aspergillus carbonarius by producing iturin A, which disturbs the transport, energy metabolism, and osmotic pressure of fungal cells as revealed by transcriptomics analysis. Int. J. Food Microbiol. 2020, 330, 108783. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Yang, B.; Humza, M.; Geng, H.; Wang, G.; Zhang, C.; Gao, S.; Xing, F.; Liu, Y. A novel strain Lactobacillus brevis 8-2B inhibiting Aspergillus carbonarius growth and ochratoxin a production. LWT 2021, 136, 110308. [Google Scholar] [CrossRef]
- Liu, J.; Sui, Y.; Wisniewski, M.; Droby, S.; Liu, Y. Review: Utilization of antagonistic yeasts to manage postharvest fungal diseases of fruit. Int. J. Food Microbiol. 2013, 167, 153–160. [Google Scholar] [CrossRef]
- Tryfinopoulou, P.; Chourdaki, A.; Nychas, G.E.; Panagou, E.Z. Competitive yeast action against Aspergillus carbonarius growth and ochratoxin A production. Int. J. Food Microbiol. 2020, 317, 108460. [Google Scholar] [CrossRef]
- Pulvirenti, A.; De Vero, L.; Blaiotta, G.; Sidari, R.; Iosca, G.; Gullo, M.; Caridi, A. Selection of Wine Saccharomyces cerevisiae Strains and Their Screening for the Adsorption Activity of Pigments, Phenolics and Ochratoxin A. Fermentation 2020, 6, 80. [Google Scholar] [CrossRef]
- Wang, L.; Hua, X.; Jing, N.; Ji, T.; Zhou, C.; Liu, W.; Lv, B.; Liu, L.; Chen, Y. Isolation and characterization of Bacillus amyloliquefaciens YL-1 with ochratoxin A degradation ability and biocontrol activity against Aspergillus westerdijkiae. Biol. Control 2022, 175, 105052. [Google Scholar] [CrossRef]
- Wei, W.; Qian, Y.; Wu, Y.; Chen, Y.; Peng, C.; Luo, M.; Xu, J.; Zhou, Y. Detoxification of ochratoxin A by Lysobacter sp. CW239 and characteristics of a novel degrading gene carboxypeptidase cp4. Environ. Pollut. 2020, 258, 113677. [Google Scholar] [CrossRef]
- Xiong, K.; Zhi, H.W.; Liu, J.Y.; Wang, X.Y.; Zhao, Z.Y.; Pei, P.G.; Deng, L.; Xiong, S.Y. Detoxification of Ochratoxin A by a novel Aspergillus oryzae strain and optimization of its biodegradation. Rev. Argent. Microbiol. 2021, 53, 48–58. [Google Scholar] [CrossRef]
- Gil-Serna, J.; Patino, B.; Cortes, L.; Gonzalez-Jaen, M.T.; Vazquez, C. Mechanisms involved in reduction of ochratoxin A produced by Aspergillus westerdijkiae using Debaryomyces hansenii CYC 1244. Int. J. Food Microbiol. 2011, 151, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Caridi, A.; Sidari, R.; Pulvirenti, A.; Blaiotta, G. Genetic Improvement of wine yeasts for opposite adsorption activity of phenolics and ochratoxin A during red winemaking. Food Biotechnol. 2020, 34, 352–370. [Google Scholar] [CrossRef]
- Ringot, D.; Lerzy, B.; Bonhoure, J.P.; Auclair, E.; Oriol, E.; Larondelle, Y. Effect of temperature on in vitro ochratoxin A biosorption onto yeast cell wall derivatives. Process Biochem. 2005, 40, 3008–3016. [Google Scholar] [CrossRef]
- Piotrowska, M. The adsorption of ochratoxin a by lactobacillus species. Toxins 2014, 6, 2826–2839. [Google Scholar] [CrossRef] [PubMed]
- Wei, M.; Dhanasekaran, S.; Ji, Q.; Yang, Q.; Zhang, H. Sustainable and efficient method utilizing N-acetyl-L-cysteine for complete and enhanced ochratoxin A clearance by antagonistic yeast. J. Hazard Mater. 2023, 448, 130975. [Google Scholar] [CrossRef]
- Wei, M.; Dhanasekaran, S.; Legrand Ngolong Ngea, G.; Abiso Godana, E.; Zhang, X.; Yang, Q.; Zheng, X.; Zhang, H. Cryptococcus podzolicus Y3 degrades ochratoxin A by intracellular enzymes and simultaneously eliminates citrinin. Biol. Control 2022, 168, 104857. [Google Scholar] [CrossRef]
- Orozco-Cortes, P.C.; Flores-Ortiz, C.M.; Hernandez-Portilla, L.B.; Vazquez Medrano, J.; Rodriguez-Pena, O.N. Molecular Docking and In Vitro Studies of Ochratoxin A (OTA) Biodetoxification Testing Three Endopeptidases. Molecules 2023, 28, 2019. [Google Scholar] [CrossRef]
- Dobritzsch, D.; Wang, H.; Schneider, G.; Yu, S. Structural and functional characterization of ochratoxinase, a novel mycotoxin-degrading enzyme. Biochem. J. 2014, 462, 441–452. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, Y.; Yin, T.; Wang, J.; Zhang, X. Heterologous Expression and Characterization of A Novel Ochratoxin A Degrading Enzyme, N-acyl-L-amino Acid Amidohydrolase, from Alcaligenes faecalis. Toxins 2019, 11, 518. [Google Scholar] [CrossRef]
- Luo, H.; Wang, G.; Chen, N.; Fang, Z.; Xiao, Y.; Zhang, M.; Gerelt, K.; Qian, Y.; Lai, R.; Zhou, Y. A Superefficient Ochratoxin A Hydrolase with Promising Potential for Industrial Applications. Appl. Environ. Microbiol. 2022, 88, e0196421. [Google Scholar] [CrossRef]
- Peng, M.; Zhang, Z.; Xu, X.; Zhang, H.; Zhao, Z.; Liang, Z. Purification and characterization of the enzymes from Brevundimonas naejangsanensis that degrade ochratoxin A and B. Food Chem. 2023, 419, 135926. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Pang, M.; Liu, J.; Wang, Y.; Wu, X.; Huang, K.; Liang, Z. Genome mining reveals the genes of carboxypeptidase for OTA-detoxification in Bacillus subtilis CW14. Int. J. Biol. Macromol. 2021, 186, 800–810. [Google Scholar] [CrossRef]
- Zhao, M.; Wang, X.Y.; Xu, S.H.; Yuan, G.Q.; Shi, X.J.; Liang, Z.H. Degradation of ochratoxin A by supernatant and ochratoxinase of Aspergillus niger W-35 isolated from cereals. World Mycotoxin J. 2020, 13, 287–298. [Google Scholar] [CrossRef]
- Liuzzi, V.C.; Fanelli, F.; Tristezza, M.; Haidukowski, M.; Picardi, E.; Manzari, C.; Lionetti, C.; Grieco, F.; Logrieco, A.F.; Thon, M.R.; et al. Transcriptional Analysis of Acinetobacter sp. neg1 Capable of Degrading Ochratoxin A. Front. Microbiol. 2016, 7, 2162. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Qiu, X.; Li, Y.; Tang, S.; Shen, W.; Xing, C.; Kong, D.; Sheng, J. Carboxypeptidase A immobilization with zeolitic imidazolate framework for enhancement of ochratoxin A degradation ability. Food Agric. Immunol. 2020, 31, 587–599. [Google Scholar] [CrossRef]
- Colovic, R.; Puvaca, N.; Cheli, F.; Avantaggiato, G.; Greco, D.; Duragic, O.; Kos, J.; Pinotti, L. Decontamination of Mycotoxin-Contaminated Feedstuffs and Compound Feed. Toxins 2019, 11, 617. [Google Scholar] [CrossRef]
- Sheikh-Zeinoddin, M.; Khalesi, M. Biological detoxification of ochratoxin A in plants and plant products. Toxin Rev. 2018, 38, 187–199. [Google Scholar] [CrossRef]
- Freire, L.; Furtado, M.M.; Guerreiro, T.M.; da Graca, J.S.; da Silva, B.S.; Oliveira, D.N.; Catharino, R.R.; Sant’Ana, A.S. The presence of ochratoxin A does not influence Saccharomyces cerevisiae growth kinetics but leads to the formation of modified ochratoxins. Food Chem. Toxicol. 2019, 133, 110756. [Google Scholar] [CrossRef]
- Wang, L.; Wang, Q.; Wang, S.; Cai, R.; Yuan, Y.; Yue, T.; Wang, Z. Bio-control on the contamination of Ochratoxin A in food: Current research and future prospects. Curr. Res. Food Sci. 2022, 5, 1539–1549. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, L.; Han, M.; Wang, X.; Guo, Y. Ochratoxin A: Overview of Prevention, Removal, and Detoxification Methods. Toxins 2023, 15, 565. https://doi.org/10.3390/toxins15090565
Ding L, Han M, Wang X, Guo Y. Ochratoxin A: Overview of Prevention, Removal, and Detoxification Methods. Toxins. 2023; 15(9):565. https://doi.org/10.3390/toxins15090565
Chicago/Turabian StyleDing, Lijuan, Meihua Han, Xiangtao Wang, and Yifei Guo. 2023. "Ochratoxin A: Overview of Prevention, Removal, and Detoxification Methods" Toxins 15, no. 9: 565. https://doi.org/10.3390/toxins15090565
APA StyleDing, L., Han, M., Wang, X., & Guo, Y. (2023). Ochratoxin A: Overview of Prevention, Removal, and Detoxification Methods. Toxins, 15(9), 565. https://doi.org/10.3390/toxins15090565