Endocrine Effect of Some Mycotoxins on Humans: A Clinical Review of the Ways to Mitigate the Action of Mycotoxins

 , , and
, , and 
Abstract
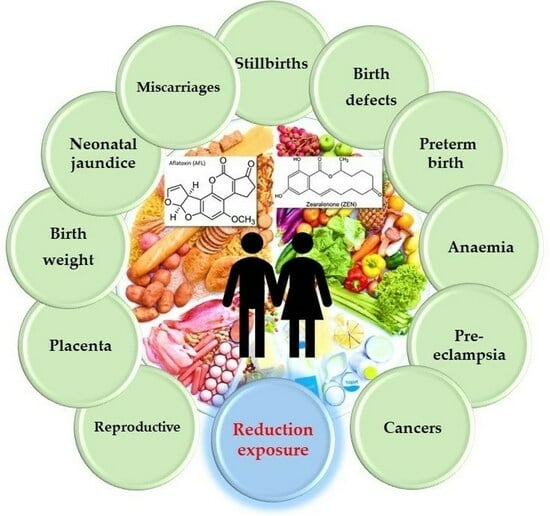
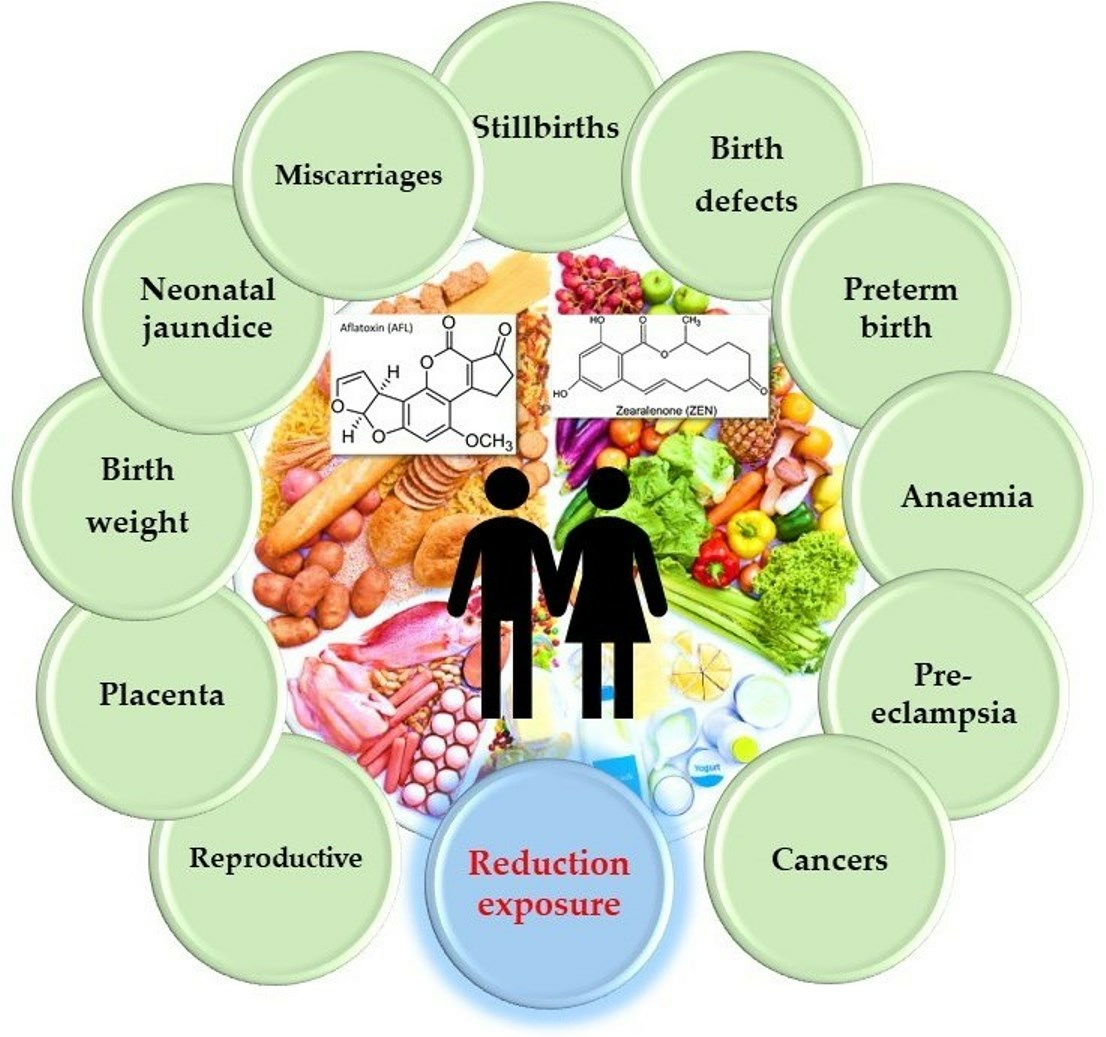{kind=link}
1. Introduction
2. Methodology
2.1. Mycotoxins and the Human Reproductive System
2.2. Mycotoxins and the Placenta
2.3. Mycotoxins and Birth Weight
2.4. Mycotoxins and Neonatal Jaundice
2.5. Mycotoxins and Miscarriages and Stillbirths
2.6. Mycotoxins and Birth Defects
2.7. Mycotoxins and Preterm Birth
2.8. Mycotoxins and Anaemia
2.9. Mycotoxins and Pre-Eclampsia
2.10. Mycotoxins and Cancer
2.11. Exposure, Prevention and Detoxification
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- UNEP; WHO. State of the Science of Endocrine Disrupting Chemicals—2012; WHO: Geneva, Switzerland, 2012. [Google Scholar]
- Kowalska, K.; Habrowska-Górczyńska, D.E.; Piastowska-Ciesielska, A.W. Zearalenone as an Endocrine Disruptor in Humans. Environ. Toxicol. Pharmacol. 2016, 48, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Council for Agricultural Science and Technology (CAST). Mycotoxins, Risks in Plants, Animal and Human System; Council Agricultural Science and Technology (CAST): Ames, IA, USA, 2003. [Google Scholar]
- Gadzała-Kopciuch, R.; Cendrowski, K.; Cesarz, A.; Kiełbasa, P.; Buszewski, B. Determination of Zearalenone and Its Metabolites in Endometrial Cancer by Coupled Separation Techniques. Anal. Bioanal. Chem. 2011, 401, 2069–2078. [Google Scholar] [CrossRef] [PubMed]
- Mostrom, M.S. Zearalenone. In Veterinary Toxicology: Basic and Clinical Principles; Gupta, R., Ed.; Academic Press: Cambridge, MA, USA, 2012; pp. 1266–1271. [Google Scholar]
- Fink-Gremmels, J.; Malekinejad, H. Clinical effects and biochemical mechanisms associated with exposure to the mycoestrogen zearalenone. Anim. Feed Sci. Technol. 2007, 137, 326–334. [Google Scholar] [CrossRef]
- Zinedine, A.; Soriano, J.M.; Moltó, J.C.; Mañes, J. Review on the toxicity, occurrence, metabolism, detoxification, regulations and intake of zearalenone: An oestrogenic mycotoxin. Food Chem. Toxicol. 2007, 45, 1–18. [Google Scholar] [CrossRef]
- Ueberschär, K.-H.; Brezina, U.; Dänicke, S. Zearalenone (ZEN) and ZEN metabolites in feed, urine and bile of sows: Analysis, determination of the metabolic profile and evaluation of the binding forms. Appl. Agric. For. Res. 2016, 1, 21–28. [Google Scholar] [CrossRef]
- Wang, Y.; Zheng, W.; Bian, X.; Yuan, Y.; Gu, J.; Liu, X.; Liu, Z.; Bian, J. Zearalenone Induces Apoptosis and Cytoprotective Autophagy in Primary Leydig Cells. Toxicol. Lett. 2014, 226, 182–191. [Google Scholar] [CrossRef]
- Frizzell, C.; Ndossi, D.; Verhaegen, S.; Dahl, E.; Eriksen, G.; Sørlie, M.; Ropstad, E.; Muller, M.; Elliott, C.T.; Connolly, L. Endocrine Disrupting Effects of Zearalenone, Alpha- and Beta-Zearalenol at the Level of Nuclear Receptor Binding and Steroidogenesis. Toxicol. Lett. 2011, 206, 210–217. [Google Scholar] [CrossRef]
- Pfeiffer, E.; Kommer, A.; Dempe, J.S.; Hildebrand, A.A.; Metzler, M. Absorption and Metabolism of the Mycotoxin Zearalenone and the Growth Promotor Zeranol in Caco-2 Cells in Vitro. Mol. Nutr. Food Res. 2010, 55, 560–567. [Google Scholar] [CrossRef]
- So, M.Y.; Tian, Z.; Phoon, Y.S.; Sha, S.; Antoniou, M.N.; Zhang, J.; Wu, R.S.S.; Tan-Un, K.C. Gene Expression Profile and Toxic Effects in Human Bronchial Epithelial Cells Exposed to Zearalenone. PLoS ONE 2014, 9, e96404. [Google Scholar] [CrossRef]
- Molina-Molina, J.-M.; Real, M.; Jimenez-Diaz, I.; Belhassen, H.; Hedhili, A.; Torné, P.; Fernández, M.F.; Olea, N. Assessment of Estrogenic and Anti-Androgenic Activities of the Mycotoxin Zearalenone and Its Metabolites Using In Vitro Receptor-Specific Bioassays. Food Chem. Toxicol. 2014, 74, 233–239. [Google Scholar] [CrossRef]
- Groopman, J.D.; Egner, P.A.; Schulze, K.J.; Wu, L.S.-F.; Merrill, R.; Mehra, S.; Shamim, A.A.; Ali, H.; Shaikh, S.; Gernand, A.; et al. Aflatoxin Exposure during the First 1000 Days of Life in Rural South Asia Assessed by Aflatoxin B1-Lysine Albumin Biomarkers. Food Chem. Toxicol. 2014, 74, 184–189. [Google Scholar] [CrossRef]
- Sansen, S.; Yano, J.K.; Reynald, R.L.; Schoch, G.A.; Griffin, K.J.; Stout, C.D.; Johnson, E.F. Adaptations for the Oxidation of Polycyclic Aromatic Hydrocarbons Exhibited by the Structure of Human P450 1A2. J. Biol. Chem. 2007, 282, 14348–14355. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Wu, J.; Zhang, F.; Wen, J.; Jiang, J.; Deng, Y. The Critical Role of Porcine Cytochrome P450 3A46 in the Bioactivation of Aflatoxin B1. Biochem. Pharmacol. 2018, 156, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Kolawole, O.; Meneely, J.P.; Meneely, J.P.; Petchkongkaew, A.; Elliott, C. A review of myco-toxin biosynthetic pathways: Associated genes and their expressions under the influence of climatic factors. Fungal Biol. Rev. 2021, 37, 8–26. [Google Scholar] [CrossRef]
- Martin, J.F.; van den Berg, M.A.; Ver Loren van Themaat, E.; Liras, P. Sensing and transduction of nutritional and chemical signals in filamentous fungi: Impact on cell development and secondary metabolites bio-synthesis. Biotechnol. Adv. 2019, 37, 107392. [Google Scholar] [CrossRef]
- Loi, M.; Logrieco, A.F.; Pusztahelyi, T.; Leiter, É.; Hornok, L.; Pócsi, I. Advanced myco-toxin control and decontamination techniques in view of an increased aflatoxin risk in Europe due to climate change. Front. Microbiol. 2023, 13, 1085891. [Google Scholar] [CrossRef]
- Williams, J.H.; Phillips, T.D.; Jolly, P.E.; Stiles, J.K.; Jolly, C.M.; Aggarwal, D. Human Aflatoxicosis in Developing Countries: A Review of Toxicology, Exposure, Potential Health Consequences, and Interventions. Am. J. Clin. Nutr. 2004, 80, 1106–1122. [Google Scholar] [CrossRef]
- Gore, A.C.; Chappell, V.A.; Fenton, S.E.; Flaws, J.A.; Nadal, A.; Prins, G.S.; Toppari, J.; Zoeller, R.T. EDC-2: The Endocrine Society’s Second Scientific Statement on Endocrine-Disrupting Chemicals. Endocr. Rev. 2015, 36, E1–E150. [Google Scholar] [CrossRef]
- Meeker, J.D. Exposure to Environmental Endocrine Disruptors and Child Development. Arch. Pediatr. Adolesc. Med. 2012, 166, 952–958. [Google Scholar] [CrossRef]
- Lucchese, T.A.; Grunow, N.; Ian Werner, I.; de Jesus, A.L.; Arbex, A.K. Endocrine disruptors and fetal programming. OJEMD 2017, 7, 59–76. [Google Scholar] [CrossRef][Green Version]
- Li, L.; Wu, X.; Guan, H.; Mao, B.; Wang, H.; Yuan, X.; Chu, Y.; Sun, J.; Ge, R.-S. Zearalenone Inhibits Rat and Human 11β-Hydroxysteroid Dehydrogenase Type 2. BioMed Res. Int. 2015, 2015, 283530. [Google Scholar] [CrossRef][Green Version]
- Obremski, K.; Gonkowski, S.; Wojtacha, P. Zearalenone-Induced Changes in the Lymphoid Tissue and Mucosal Nerve Fibers in the Porcine Ileum. Pol. J. Vet. Sci. 2015, 18, 357–365. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Endocrine Disorders and Children, Children’s Health and the Environment. Available online: http://www.portal.pmnch.org/ceh/capacity/endocrine.pdf (accessed on 20 March 2023).
- Yum, T.; Lee, S.; Kim, Y. Association between Precocious Puberty and Some Endocrine Disruptors in Human Plasma. J. Environ. Sci. Health A 2013, 48, 912–917. [Google Scholar] [CrossRef]
- Massart, F.; Meucci, V.; Saggese, G.; Soldani, G. High Growth Rate of Girls with Precocious Puberty Exposed to Estrogenic Mycotoxins. J. Pediatr. 2008, 152, 690–695.e1. [Google Scholar] [CrossRef]
- Massart, F.; Saggese, G. Oestrogenic Mycotoxin Exposures and Precocious Pubertal Development. Int. J. Androl. 2010, 33, 369–376. [Google Scholar] [CrossRef] [PubMed]
- Bandera, E.V.; Chandran, U.; Buckley, B.; Lin, Y.; Isukapalli, S.; Marshall, I.; King, M.; Zarbl, H. Urinary mycoestrogens, body size and breast development in New Jersey girls. Sci. Total Environ. 2011, 409, 5221–5227. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Núñez, Z.; Barrett, E.S.; Szamreta, E.A.; Shapses, S.A.; Qin, B.; Lin, Y.; Zarbl, H.; Buckley, B.; Bandera, E.V. Urinary Mycoestrogens and Age and Height at Menarche in New Jersey Girls. Environ. Health 2019, 18, 24. [Google Scholar] [CrossRef]
- Asci, A.; Durmaz, E.; Erkekoglu, P.; Pasli, D.; Bircan, I.; Kocer-Gumusel, B. Urinary Zearalenone Levels in Girls with Premature Thelarche and Idiopathic Central Precocious Puberty. Minerva Pediatr. 2014, 66, 571–578. [Google Scholar]
- Szuets, P.; Mesterházy, Á.; Falkay, G.; Bartók, T. Early Telarche Symptoms in Children and Their Relations to Zearalenon Contamination in Foodstuffs. Cereal Res. Commun. 1997, 25, 429–436. [Google Scholar] [CrossRef]
- Zheng, W.; Feng, N.; Wang, Y.; Noll, L.; Xu, S.; Liu, X.; Lu, N.; Zou, H.; Gu, J.; Yuan, Y.; et al. Effects of Zearalenone and Its Derivatives on the Synthesis and Secretion of Mammalian Sex Steroid Hormones: A Review. Food Chem. Toxicol Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2019, 126, 262–276. [Google Scholar] [CrossRef]
- Zwierzchowski, W.; Przybyłowicz, M.; Obremski, K.; Zielonka, L.; Skorska-Wyszyńska, E.; Gajecka, M.; Polak, M.; Jakimiuk, E.; Jana, B.; Rybarczyk, L.; et al. Level of Zearalenone in Blood Serum and Lesions in Ovarian Follicles of Sexually Immature Gilts in the Course of Zearalenone Micotoxicosis. Pol. J. Vet. Sci. 2005, 8, 209–218. [Google Scholar]
- Jakimiuk, E.; Rybarczyk, L.; Zwierzchowski, W.; Obremski, K.; Gajęcka, M.; Zielonka, Ł.; Gajęcki, M. Effect of experimental long-term exposure to low-dose zearalenone mycotoxicosis on selected morphometric parameters of the reproductive tract in sexually-immature gilts. Bull. Vet. Inst. Pulawy 2010, 54, 25–28. [Google Scholar]
- Caserta, D.; Mantovani, A.; Marci, R.; Fazi, A.; Ciardo, F.; La Rocca, C.; Maranghi, F.; Moscarini, M. Environment and Women’s Reproductive Health. Hum. Reprod. Update 2011, 17, 418–433. [Google Scholar] [CrossRef]
- Balló, A.; Busznyákné Székvári, K.; Czétány, P.; Márk, L.; Török, A.; Szántó, Á.; Máté, G. Estrogenic and Non-Estrogenic Disruptor Effect of Zearalenone on Male Reproduction: A Review. Int. J. Mol. Sci. 2023, 24, 1578. [Google Scholar] [CrossRef] [PubMed]
- Ibeh, I.N.; Uraih, N.; Ogonor, J.I. Dietary exposure to aflatoxin in Benin City, Nigeria: A possible public health concern. Int. J. Food Microbiol. 1991, 14, 171–174. [Google Scholar] [CrossRef] [PubMed]
- Uriah, N.; Ibeh, I.N.; Oluwafemi, F. A Study on the Impact of Aflatoxin on Human Reproduction. AJRH 2001, 5, 106–110. [Google Scholar] [CrossRef]
- Kinkade, C.W.; Rivera-Núñez, Z.; Gorcyzca, L.; Aleksunes, L.M.; Barrett, E.S. Impact of Fusarium-Derived Mycoestrogens on Female Reproduction: A Systematic Review. Toxins 2021, 13, 373. [Google Scholar] [CrossRef]
- Prouillac, C.; Videmann, B.; Mazallon, M.; Lecoeur, S. Induction of Cells Differentiation and ABC Transporters Expression by a Myco-Estrogen, Zearalenone, in Human Choriocarcinoma Cell Line (BeWo). Toxicology 2009, 263, 100–107. [Google Scholar] [CrossRef]
- Prouillac, C.; Koraichi, F.; Videmann, B.; Mazallon, M.; Rodriguez, F.; Baltas, M.; Lecoeur, S. In Vitro Toxicological Effects of Estrogenic Mycotoxins on Human Placental Cells: Structure Activity Relationships. Toxicol. Appl. Pharmacol. 2012, 259, 366–375. [Google Scholar] [CrossRef]
- Partanen, H.A.; El-Nezami, H.S.; Leppänen, J.M.; Myllynen, P.K.; Woodhouse, H.J.; Vähäkangas, K.H. Aflatoxin B1 Transfer and Metabolism in Human Placenta. Toxicol. Sci. 2010, 113, 216–225. [Google Scholar] [CrossRef]
- Warth, B.; Preindl, K.; Manser, P.; Wick, P.; Marko, D.; Buerki-Thurnherr, T. Transfer and Metabolism of the Xenoestrogen Zearalenone in Human Perfused Placenta. Environ. Health Perspect. 2019, 127, 107004. [Google Scholar] [CrossRef] [PubMed]
- Maxwell, S.M.; Familusi, J.B.; Sodeinde, O.; Chan, M.C.; Hendrickse, R.G. Detection of Naphthols and Aflatoxins in Nigerian Cord Blood. Ann. Trop. Paediatr. 1994, 14, 3–5. [Google Scholar] [CrossRef] [PubMed]
- Abulu, E.O.; Uriah, N.; Aigbefo, H.S.; Oboh, P.A.; Agbonlahor, D.E. Preliminary Investigation on Aflatoxin in Cord Blood of Jaundiced Neonates. West. Afr. J. Med. 1998, 17, 184–187. [Google Scholar] [PubMed]
- Abdulrazzaq, Y.M.; Osman, N.; Ibrahim, A. Fetal Exposure to Aflatoxins in the United Arab Emirates. Ann. Trop. Paediatr. 2002, 22, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Abdulrazzaq, Y.M.; Osman, N.; Yousif, Z.M.; Trad, O. Morbidity in Neonates of Mothers Who Have Ingested Aflatoxins. Ann. Trop. Paediatr. 2004, 24, 145–151. [Google Scholar] [CrossRef]
- Turner, P.C.; Collinson, A.C.; Cheung, Y.B.; Gong, Y.; Hall, A.J.; Prentice, A.M.; Wild, C.P. Aflatoxin Exposure in Utero Causes Growth Faltering in Gambian Infants. Int. J. Epidemiol. 2007, 36, 1119–1125. [Google Scholar] [CrossRef]
- Shuaib, F.M.B.; Person, S.D.; Funkhouser, E.; Yatich, N.J.; Stiles, J.K.; Ellis, W.O.; Jiang, Y.; Ehiri, J.E.; Williams, J.H.; Jolly, P.E.; et al. Association between Anemia and Aflatoxin B1 Biomarker Levels among Pregnant Women in Kumasi, Ghana. Am. J. Trop. Med. Hyg. 2010, 83, 1077–1083. [Google Scholar] [CrossRef]
- Sodeinde, O.; Chan, M.C.; Maxwell, S.M.; Familusi, J.B.; Hendrickse, R.G. Neonatal Jaundice, Aflatoxins and Naphthols: Report of a Study in Ibadan, Nigeria. Ann. Trop. Paediatr. 1995, 15, 107–113. [Google Scholar] [CrossRef]
- Ahmed, H.; Hendrickse, R.G.; Maxwell, S.M.; Yakubu, A.M. Neonatal Jaundice with Reference to Aflatoxins: An Aetiological Study in Zaria, Northern Nigeria. Ann. Trop. Paediatr. 1995, 15, 11–20. [Google Scholar] [CrossRef]
- Shuaib, F.M.B.; Jolly, P.E.; Ehiri, J.E.; Yatich, N.; Jiang, Y.; Funkhouser, E.; Person, S.D.; Wilson, C.; Ellis, W.O.; Wang, J.-S.; et al. Association between Birth Outcomes and Aflatoxin B1Biomarker Blood Levels in Pregnant Women in Kumasi, Ghana. Trop. Med. Int. Health 2010, 15, 160–167. [Google Scholar] [CrossRef]
- Kristensen, P.; Irgens, L.M.; Andersen, A.; Bye, A.S.; Sundheim, L. Gestational Age, Birth Weight, and Perinatal Death among Births to Norwegian Farmers, 1967-1991. Am. J. Epidemiol. 1997, 146, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, P.; Andersen, A.; Irgens, L.M. Hormone-Dependent Cancer and Adverse Reproductive Outcomes in Farmers’ Families—Effects of Climatic Conditions Favoring Fungal Growth in Grain. Scand. J. Work Environ. Health 2000, 26, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Nordby, K.-C.; Irgens, L.M.; Kristensen, P. Immunological Exposures in Norwegian Agriculture and Pre-Eclampsia. Paediatr. Perinat. Epidemiol. 2006, 20, 462–470. [Google Scholar] [CrossRef] [PubMed]
- Carlos, R.L.J.; Leticia, I.G.; Efrain, F.S.E.; Miguel, R.A. Aflatoxigenic Feeding and Its Possible Implications after Pregnancy. Biomed. Pharmacol. J. 2015, 7, 183–193. [Google Scholar] [CrossRef]
- Omotayo, O.P.; Omotayo, A.O.; Mwanza, M.; Babalola, O.O. Prevalence of Mycotoxins and Their Consequences on Human Health. Toxicol. Res. 2019, 35, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Missmer, S.A.; Suarez, L.; Felkner, M.; Wang, E.; Merrill, A.H.; Rothman, K.J.; Hendricks, K.A. Exposure to Fumonisins and the Occurrence of Neural Tube Defects along the Texas–Mexico Border. Environ. Health Perspect. 2006, 114, 237–241. [Google Scholar] [CrossRef]
- Smith, L.E.; Prendergast, A.J.; Turner, P.C.; Humphrey, J.H.; Stoltzfus, R.J. Aflatoxin Exposure during Pregnancy, Maternal Anemia, and Adverse Birth Outcomes. Am. J. Trop. Med. Hyg. 2017, 96, 770–776. [Google Scholar] [CrossRef]
- Hagberg, H.; Mallard, C.; Jacobsson, B. Role of Cytokines in Preterm Labour and Brain Injury. BJOG Int. J. Obstet. Gynaecol. 2005, 112, 16–18. [Google Scholar] [CrossRef]
- Wang, Y.; Li, L.; Wang, C.C.; Leung, L.K. Effect of Zeranol on Expression of Apoptotic and Cell Cycle Proteins in Murine Placentae. Toxicology 2013, 314, 148–154. [Google Scholar] [CrossRef]
- Andrews-Trevino, J.Y.; Webb, P.; Shively, G.; Rogers, B.L.; Baral, K.; Davis, D.; Paudel, K.; Pokharel, A.; Shrestha, R.; Wang, J.-S.; et al. Relatively Low Maternal Aflatoxin Exposure Is Associated with Small-For-Gestational-Age but Not with Other Birth Outcomes in a Prospective Birth Cohort Study of Nepalese Infants. J. Nutr. 2019, 149, 1818–1825. [Google Scholar] [CrossRef]
- Andretta, I.; Kipper, M.; Lehnen, C.R.; Lovatto, P.A. Meta-Analysis of the Relationship of Mycotoxins with Biochemical and Hematological Parameters in Broilers. Poult. Sci. 2012, 91, 376–382. [Google Scholar] [CrossRef]
- Yousef, M.I.; Salem, M.H.; Kamel, K.I.; Hassan, G.A.; El-Nouty, F.D. Influence of Ascorbic Acid Supplementation on the Haematological and Clinical Biochemistry Parameters of Male Rabbits Exposed to Aflatoxin B1. J. Environ. Sci. Health Part B 2003, 38, 193–209. [Google Scholar] [CrossRef]
- Eisa, A.; Metwally, A. Effect of Glucomannan on Haematological, Coagulation and Biochemical Parameters in Male Rabbits Fed Aflatoxin-Contaminated Ration. World Mycotoxin J. 2011, 4, 183–188. [Google Scholar] [CrossRef]
- Moodley, D.; Moodley, L.; Reddy, M.F.; Dutton, A.A.; Chuturgoon, J. Fumonisin B 1: An Aetiological Role in Pre-Eclampsia. J. Obstet. Gynaecol. 2001, 21, 599–600. [Google Scholar] [CrossRef] [PubMed]
- Pazaiti, A.; Kontos, M.; Fentiman, I.S. ZEN and the Art of Breast Health Maintenance. Int. J. Clin. Pract. 2011, 66, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, S.V.; Russo, J. Estrogen and Xenoestrogens in Breast Cancer. Toxicol. Pathol. 2009, 38, 110–122. [Google Scholar] [CrossRef]
- Fucic, A.; Gamulin, M.; Ferencic, Z.; Katic, J.; Krayer von Krauss, M.; Bartonova, A.; Merlo, D.F. Environmental Exposure to Xenoestrogens and Oestrogen Related Cancers: Reproductive System, Breast, Lung, Kidney, Pancreas, and Brain. Environ. Health 2012, 11 (Suppl. S1), S8. [Google Scholar] [CrossRef]
- Rogowska, A.; Pomastowski, P.; Sagandykova, G.; Buszewski, B. Zearalenone and Its Metabolites: Effect on Human Health, Metabolism and Neutralisation Methods. Toxicon 2019, 162, 46–56. [Google Scholar] [CrossRef]
- Kuciel-Lisieska, G.; Obremski, K.; Stelmachów, J.; Gajecka, M.; Zielonka, Ł.; Jakimiuk, E.; Gajecki, M. Presence of zearalenone in blood plasma in women with neoplastic lesions in the mammary gland. Bul. Vet. Inst. Pulawy 2008, 52, 671–674. [Google Scholar]
- Belhassen, H.; Jiménez-Díaz, I.; Arrebola, J.P.; Ghali, R.; Ghorbel, H.; Olea, N.; Hedili, A. Zearalenone and Its Metabolites in Urine and Breast Cancer Risk: A Case-Control Study in Tunisia. Chemosphere 2015, 128, 1–6. [Google Scholar] [CrossRef]
- Pillay, D.; Chuturgoon, A.A.; Nevines, E.; Manickum, T.; Deppe, W.; Dutton, M.F. The Quantitative Analysis of Zearalenone and Its Derivatives in Plasma of Patients with Breast and Cervical Cancer. Clin. Chem. Lab. Med. 2002, 40, 946–951. [Google Scholar] [CrossRef]
- Wan, M.L.Y.; Co, V.A.; El-Nezami, H. Endocrine Disrupting Chemicals and Breast Cancer: A Systematic Review of Epidemiological Studies. Crit. Rev. Food Sci. Nutr. 2022, 62, 6549–6576. [Google Scholar] [CrossRef] [PubMed]
- Pajewska, M.; Łojko, M.; Cendrowski, K.; Sawicki, W.; Kowalkowski, T.; Buszewski, B.; Gadzała-Kopciuch, R. The Determination of Zearalenone and Its Major Metabolites in Endometrial Cancer Tissues. Anal. Bioanal. Chem. 2018, 410, 1571–1582. [Google Scholar] [CrossRef] [PubMed]
- Marchese, S.; Polo, A.; Ariano, A.; Velotto, S.; Costantini, S.; Severino, L. Aflatoxin B1 and M1: Biological Properties and Their Involvement in Cancer Development. Toxins 2018, 10, 214. [Google Scholar] [CrossRef] [PubMed]
- Caserta, D.; De Marco, M.P.; Besharat, A.R.; Costanzi, F. Endocrine Disruptors and Endometrial Cancer: Molecular Mechanisms of Action and Clinical Implications, a Systematic Review. Int. J. Mol. Sci. 2022, 23, 2956. [Google Scholar] [CrossRef] [PubMed]
- Eze, U.A.; Okonofua, F.E. High Prevalence of Male Infertility in Africa: Are Mycotoxins to Blame? Afr. J. Reprod. Health 2015, 19, 9–17. [Google Scholar] [PubMed]
- Mally, A.; Solfrizzo, M.; Degen, G.H. Biomonitoring of the mycotoxin Zearalenone: Current state-of-the art and application to human exposure assessment. Arch. Toxicol. 2016, 90, 1281–1292. [Google Scholar] [CrossRef]
- Al-Jaal, B.A.; Jaganjac, M.; Barcaru, A.; Horvatovich, P.; Latiff, A. Aflatoxin, fumonisin, ochratoxin, zearalenone and deoxynivalenol biomarkers in human biological fluids: A systematic literature review, 2001–2018. Food Chem. Toxicol. 2019, 129, 211–228. [Google Scholar] [CrossRef]
- Fleck, S.C.; Churchwell, M.I.; Doerge, D.R.; Teeguarden, J.G. Urine and serum biomonitoring of exposure to environmental estrogens II: Soy isoflavones and zearalenone in pregnant women. Food Chem. Toxicol. 2016, 95, 19–27. [Google Scholar] [CrossRef]
- Mirocha, C.J.; Pathre, S.V.; Robison, T.S. Comparative Metabolism of Zearalenone and Transmission into Bovine Milk. Food Cosmet. Toxicol. 1981, 19, 25–30. [Google Scholar] [CrossRef]
- Warth, B.; Sulyok, M.; Berthiller, F.; Schuhmacher, R.; Krska, R. New Insights into the Human Metabolism of the Fusarium Mycotoxins Deoxynivalenol and Zearalenone. Toxicol. Lett. 2013, 220, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Ferrigo, D.; Raiola, A.; Causin, R. Fusarium Toxins in Cereals: Occurrence, Legislation, Factors Promoting the Appearance and Their Management. Molecules 2016, 21, 627. [Google Scholar] [CrossRef] [PubMed]
- McKenzie, K.S.; Sarr, A.B.; Mayura, K.; Bailey, R.H.; Miller, D.R.; Rogers, T.D.; Norred, W.P.; Voss, K.A.; Plattner, R.D.; Kubena, L.F.; et al. Oxidative Degradation and Detoxification of Mycotoxins Using a Novel Source of Ozone. Food Chem. Toxicol. 1997, 35, 807–820. [Google Scholar] [CrossRef]
- Dalié, D.K.D.; Deschamps, A.M.; Richard-Forget, F. Lactic acid bacteria—Potential for control of mould growth and mycotoxins: A review. Food Control 2010, 21, 370–380. [Google Scholar] [CrossRef]
- Vinderola, G.; Ritieni, A. Role of probiotics against mycotoxins and their deleterious effects. J. Food Res. 2014, 4, 10–21. [Google Scholar] [CrossRef]
- Abdolmaleki, K.; Javanmardi, F.; Gavahian, M.; Phimolsiripol, Y.; Ruksiriwanich, W.; Mir, S.A. Emerging technologies in combination with probiotics for aflatoxins removal: An updated review. Int. J. Food Sci. 2022, 57, 5712–5721. [Google Scholar] [CrossRef]
- Sangsila, A.; Faucet-Marquis, V.; Pfohl-Leszkowicz, A.; Itsaranuwat, P. Detoxification of zearalenone by Lactobacillus pentosus strains. Food Control 2016, 62, 187–192. [Google Scholar] [CrossRef]
- Harkai, P.; Szabó, I.; Cserháti, M.; Krifaton, C.; Risa, A.; Radó, J.; Balázs, A.; Berta, K.; Kriszt, B. Biodegradation of aflatoxin-B1, and zearalenone by Streptomyces sp. collection. Int. Biodeterior. Biodegrad. 2016, 108, 48–56. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, C.; Zhang, D.; Zhao, M.; Peng, M.; Guo, P.; Cui, Z. Microbial degradation of zearalenone by a novel microbial consortium, NZDC-6, and its application on contaminated corncob by semisolid fermentation. J. Agric. Food Chem. 2020, 68, 1634–1644. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, C.; Zhang, D.; Zhao, M.; Zheng, D.; Lyu, Y.; Cheng, W.; Guo, P.; Cui, Z. Effective degradation of aflatoxin B1, using a novel thermophilic microbial consortium TADC7. Bioresour. Technol. 2017, 224, 166–173. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, C.; Zhang, D.; Zhao, M.; Zheng, D.; Peng, M.; Cheng, W.; Guo, P.; Cui, Z. Simultaneous degradation of aflatoxin B1, and zearalenone by a microbial consortium. Toxicon 2018, 146, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Wu, H.; Tang, Y.; Qiu, L. Cloning, expression of a peroxiredoxin gene from Acinetobacter sp. SM04, and characterization of its recombinant protein for zearalenone detoxification. Microbiol. Res. 2012, 167, 121–126. [Google Scholar] [CrossRef]
- Rawal, S.; Yip, S.S.M.; Coulombe, R.A. Cloning, expression and functional characterization of cytochrome P450, 3A37, from turkey liver with high aflatoxin B1, epoxidation activity. Chem. Res. Toxicol. 2010, 23, 1322–1329. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Branà, M.T.; Haidukowski, M.; Gallo, A.; Zhang, Q.; Logrieco, A.F. Potential of Trichoderma spp. for biocontrol of aflatoxin-producing Aspergillus flavus. Toxins 2022, 14, 86. [Google Scholar] [CrossRef]
- Gamal, M.; Abou Zaid, M.; Abou Mourad, I.K.; Abd El Kareem, H.; Gomaa, O.M. Trichoderma viride bioactive peptaibol induces apoptosis in Aspergillus niger in-fecting tilapia in fish farms. Aquaculture 2022, 547, 737474. [Google Scholar] [CrossRef]
- Woo, S.L.; Ruocco, M.; Vinale, F.; Nigro, M.; Marra, R.; Lombardi, N. Trichoderma-based products and their widespread use in agriculture. Open Mycol. J. 2014, 8, 71–126. [Google Scholar] [CrossRef]
- Sun, S.; Zhao, R.; Xie, Y.; Liu, Y. Photocatalytic degradation of aflatoxin B1 by activated carbon supported TiO2 catalyst. Food Control 2019, 100, 183–188. [Google Scholar] [CrossRef]
- Faraji, H.; Yazdi, F.T.; Razmi, N. The influence of ultraviolet radiation on aflatoxin producing Aspergillus species’ isolated from Iranian rice. Toxicol. Rep. 2022, 9, 1528–1536. [Google Scholar] [CrossRef]
- Calado, T.; Abrunhosa, L.; Cabo Verde, S.; Alté, L.; Venâncio, A.; Fernández-Cruz, M.L. Effect of Gamma-Radiation on Zearalenone-Degradation, Cytotoxicity and Estrogenicity. Foods 2020, 9, 1687. [Google Scholar] [CrossRef]
- Kalagatur, N.K.; Kamasani, J.R.; Mudili, V. Assessment of detoxification efficacy of irradiation on zearalenone mycotoxin in various fruit juices by response surface methodology and elucidation of its in-vitro toxicity. Front. Microbiol. 2018, 9, 2937. [Google Scholar] [CrossRef]
- Shen, M.H.; Singh, R.K. Effective UV wavelength range for increasing aflatoxins reduction and decreasing oil deterioration in contaminated peanuts. Food Res. Int. 2022, 154, 111016. [Google Scholar] [CrossRef]
- Horky, P.; Skalickova, S.; Baholet, D.; Skladanka, J. Nanoparticles as a Solution for Eliminating the Risk of Mycotoxins. Nanomaterials 2018, 8, 727. [Google Scholar] [CrossRef] [PubMed]
- Duishemambet Kyzy, A.; Kocyigit, Y.; Ardag, A.H. Aflatoxin B1 bioremoval by fungal cells immobilised on magnetic nanoparticles. J. Environ. Anal. Chem. 2022, 102, 1–14. [Google Scholar] [CrossRef]
- Lu, D.; Tang, S.; Li, Y.; Cong, Z.; Zhang, X.; Wu, S. Magnetic-propelled Janus yeast cell robots functionalized with metal-organic frameworks for mycotoxin decontamination. Micromachines 2021, 12, 797. [Google Scholar] [CrossRef]
- Malhotra, N.; Lee, J.S.; Liman, R.A.D.; Ruallo, J.M.S.; Villaflores, O.B.; Ger, T.R. Potential toxicity of iron oxide magnetic nanoparticles: A review. Molecules 2020, 25, 3159. [Google Scholar] [CrossRef]
- Mandal, R.; Singh, A.; and Pratap, S.A. Recent developments in cold plasma decontamination technology in the food industry. Trends Food Sci. Technol. 2018, 80, 93–103. [Google Scholar] [CrossRef]
- Hojnik, N.; Modic, M.; Walsh, J.L.; Žigon, D.; Javornik, U.; Plavec, J.; Zegura, B.; Filipic, M.; Cvelbar, U. Unravelling the pathways of air plasma induced aflatoxin B1 degradation and detoxification. J. Hazard. Mater. 2021, 403, 123593. [Google Scholar] [CrossRef]
- Wu, Y.; Cheng, J.H.; Sun, D.W. Blocking and degradation of aflatoxins by cold plasma treatments: Applications and mechanisms. Trends Food Sci. Technol. 2021, 109, 647–661. [Google Scholar] [CrossRef]
- Zhang, X.; Li, G.; Wu, D.; Liu, J.; Wu, Y. Recent advances on emerging nanomaterials for controlling the mycotoxin contamination: From detection to elimination. Food Front. 2020, 1, 360–381. [Google Scholar] [CrossRef]
- Guo, Y.; Zhao, L.; Ma, Q.; Ji, C. Novel strategies for degradation of aflatoxins in food and feed: A review. Food Res. Int. 2021, 140, 109878. [Google Scholar] [CrossRef]
- Ma, F.; Cai, X.; Mao, J.; Yu, L.; Li, P. Adsorptive removal of aflatoxin B1 from vegetable oils via novel adsorbents derived from a metal-organic framework. J. Hazard. Mater. 2021, 412, 125170. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Gómez, E.O.; García-Rosales, G.; Longoria-Gándara, L.C.; Gómez-Vilchis, J.C. Obtention of biochar-Ca nanoparticles using citrus tangerina: A morphological, surface and study remotion of aflatoxin AFB1. J. Hazard. Mater. 2022, 424, 127339. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Wu, X.; Wu, C.; Hou, F.; Wu, L.; Huang, H. Metal-organic frameworks with peroxidase-like activity for efficient removal of aflatoxin B1. Food Chem. 2022, 378, 132037. [Google Scholar] [CrossRef]
- Zingales, V.; Taroncher, M.; Martino, P.A.; Ruiz, M.J.; Caloni, F. Climate Change and Effects on Molds and Mycotoxins. Toxins. 2022, 14, 445. [Google Scholar] [CrossRef]
- Battilani, P.; Toscano, P.; Van der Fels-Klerx, H.J.; Moretti, A.; Camardo Leggieri, M.; Brera, C.; Rortais, A.; Goumperis, T.; Robinson, T. Aflatoxin B1 contamination in maize in Europe increases due to climate change. Sci. Rep. 2016, 6, 24328. [Google Scholar] [CrossRef]
- Garcia-Cela, E.; Crespo-Sempere, A.; Ramos, A.J.; Sanchis, V.; Marin, S. Ecophysiological characterization of Aspergillus carbonarius, Aspergillus tubingensis and Aspergillus niger isolated from grapes in Spanish vineyards. Int. J. Food Microbiol. 2014, 173, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Poppenberger, B.; Berthiller, F.; Bachmann, H.; Lucyshyn, D.; Peterbauer, C.; Mitterbauer, R.; Schuhmacher, R.; Krska, R.; Glössl, J.; Adam, G. Heterologous expression of Arabidopsis UDP-glucosyltransferases in Saccharomyces cerevisiae for production of zearalenone-4-O-glucoside. Appl. Environ. Microbiol. 2006, 72, 4404–4410. [Google Scholar] [CrossRef]
- Gratz, S.W. Do Plant-Bound Masked Mycotoxins Contribute to Toxicity? Toxins 2017, 9, 85. [Google Scholar] [CrossRef]
- Knutsen, H.K.; Barregård, L.; Bignami, M.; Brüschweiler, B.; Ceccatelli, S.; Cottrill, B.; Dinovi, M.; Edler, L.; Grasl-Kraupp, B.; Hogstrand, C.; et al. Appropriateness to set a group health-based guidance value for fumonisins and their modified forms. EFSA J. 2018, 16, e05172. [Google Scholar] [CrossRef]
- Sun, Y.; Jiang, J.; Mu, P.; Lin, R.; Wen, J.; Deng, Y. Toxicokinetics and metabolism of deoxynivalenol in animals and humans. Arch. Toxicol. 2022, 10, 2639–2654. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kościelecka, K.; Kuć, A.; Kubik-Machura, D.; Męcik-Kronenberg, T.; Włodarek, J.; Radko, L. Endocrine Effect of Some Mycotoxins on Humans: A Clinical Review of the Ways to Mitigate the Action of Mycotoxins. Toxins 2023, 15, 515. https://doi.org/10.3390/toxins15090515
Kościelecka K, Kuć A, Kubik-Machura D, Męcik-Kronenberg T, Włodarek J, Radko L. Endocrine Effect of Some Mycotoxins on Humans: A Clinical Review of the Ways to Mitigate the Action of Mycotoxins. Toxins. 2023; 15(9):515. https://doi.org/10.3390/toxins15090515
Chicago/Turabian StyleKościelecka, Klaudia, Aleksandra Kuć, Daria Kubik-Machura, Tomasz Męcik-Kronenberg, Jan Włodarek, and Lidia Radko. 2023. "Endocrine Effect of Some Mycotoxins on Humans: A Clinical Review of the Ways to Mitigate the Action of Mycotoxins" Toxins 15, no. 9: 515. https://doi.org/10.3390/toxins15090515
APA StyleKościelecka, K., Kuć, A., Kubik-Machura, D., Męcik-Kronenberg, T., Włodarek, J., & Radko, L. (2023). Endocrine Effect of Some Mycotoxins on Humans: A Clinical Review of the Ways to Mitigate the Action of Mycotoxins. Toxins, 15(9), 515. https://doi.org/10.3390/toxins15090515