Vitamin D Supplementation Impacts Calcium and Phosphorus Metabolism in Piglets Fed a Diet Contaminated with Deoxynivalenol and Challenged with Lipopolysaccharides


Abstract
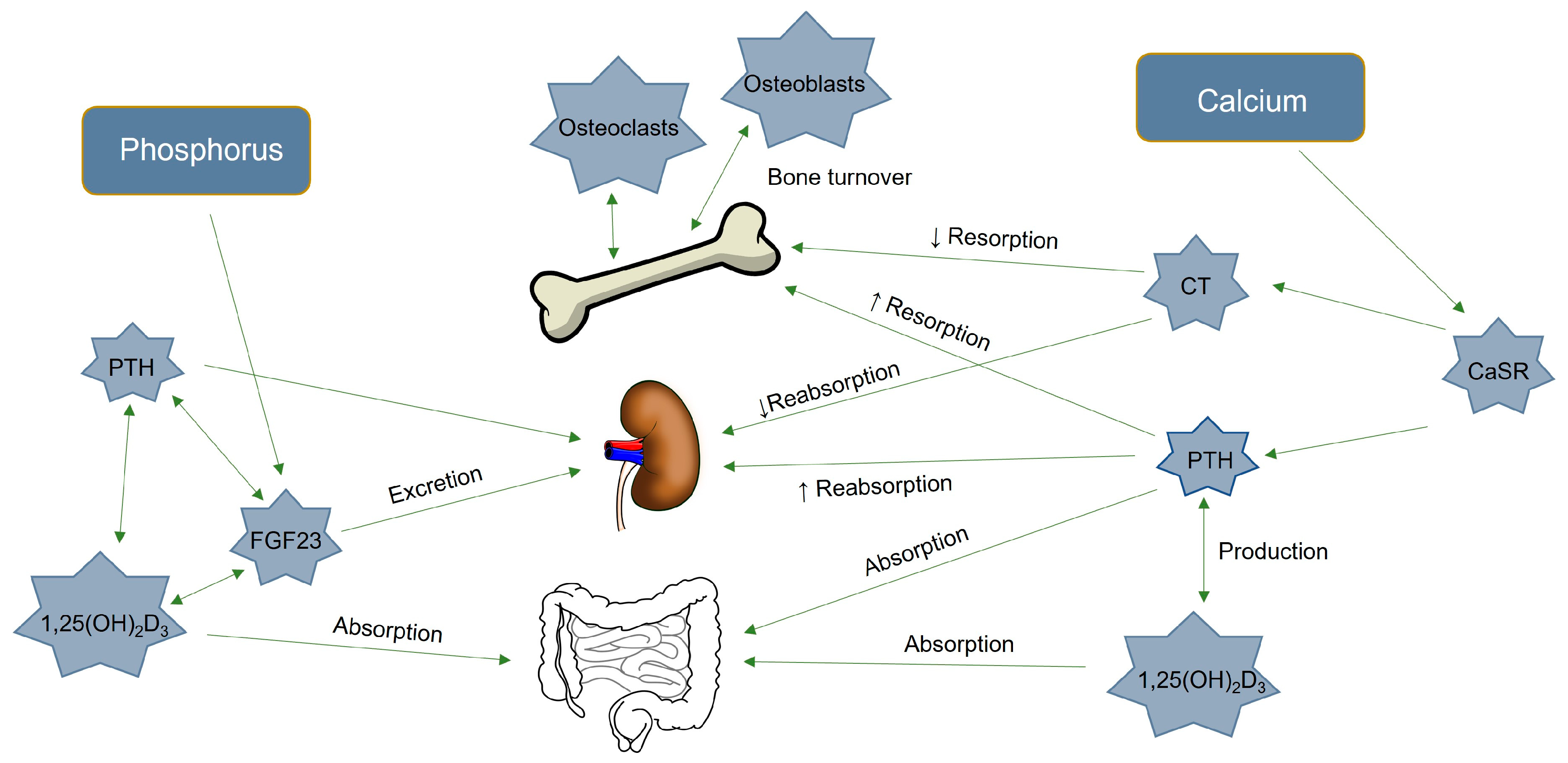
1. Introduction
2. Results
2.1. Growth Performance, Bone Mineralization
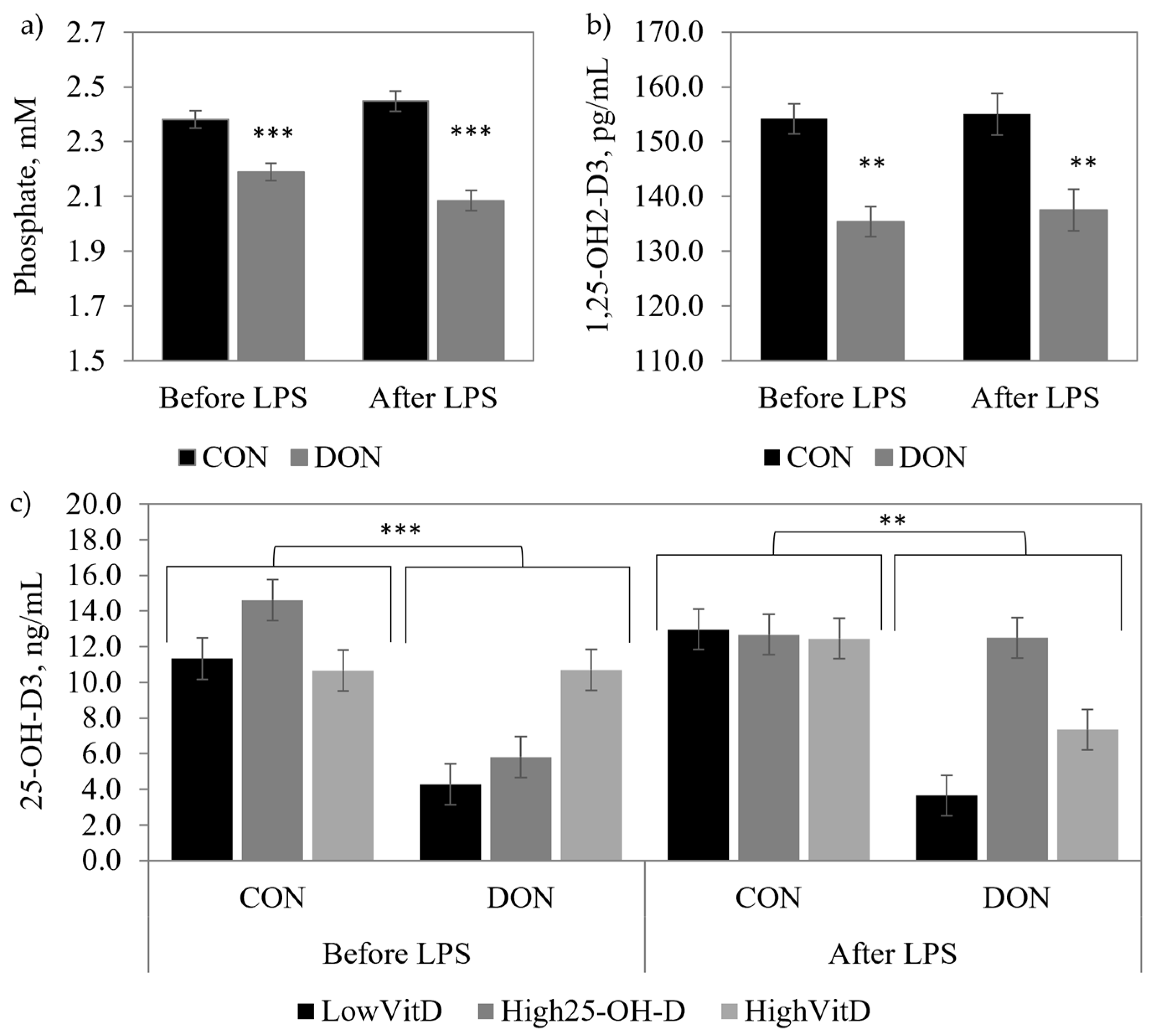
2.2. Blood Parameters
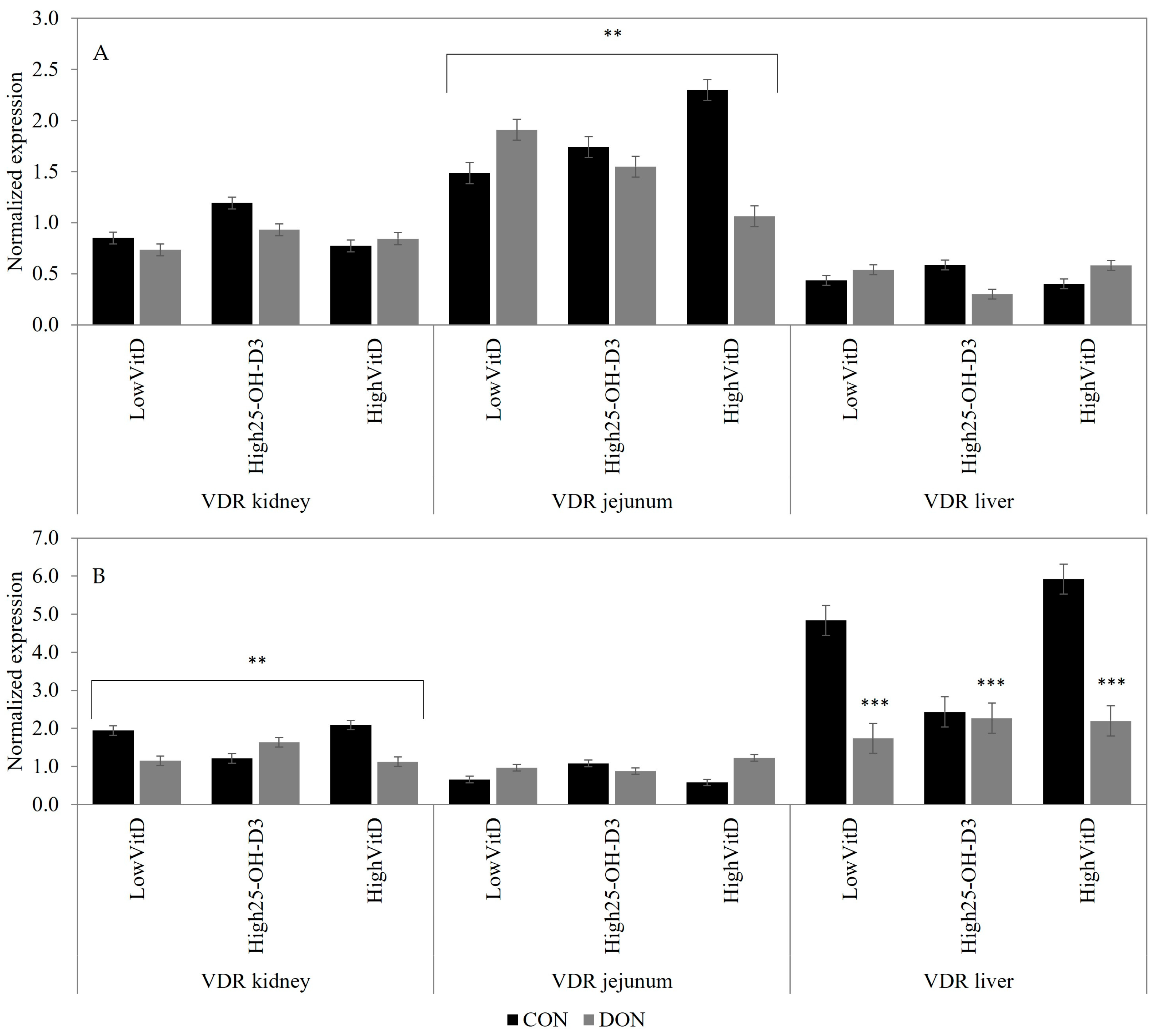
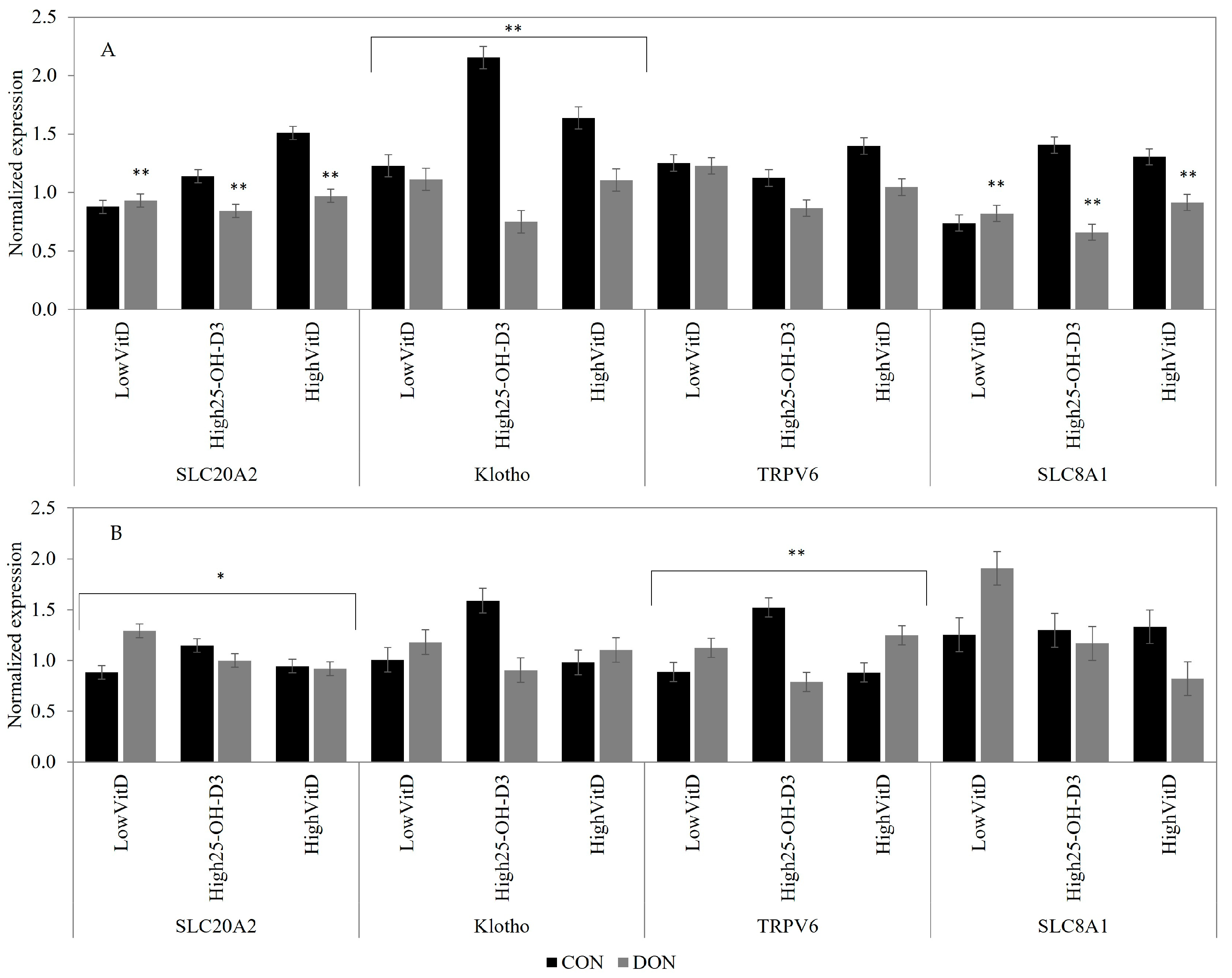
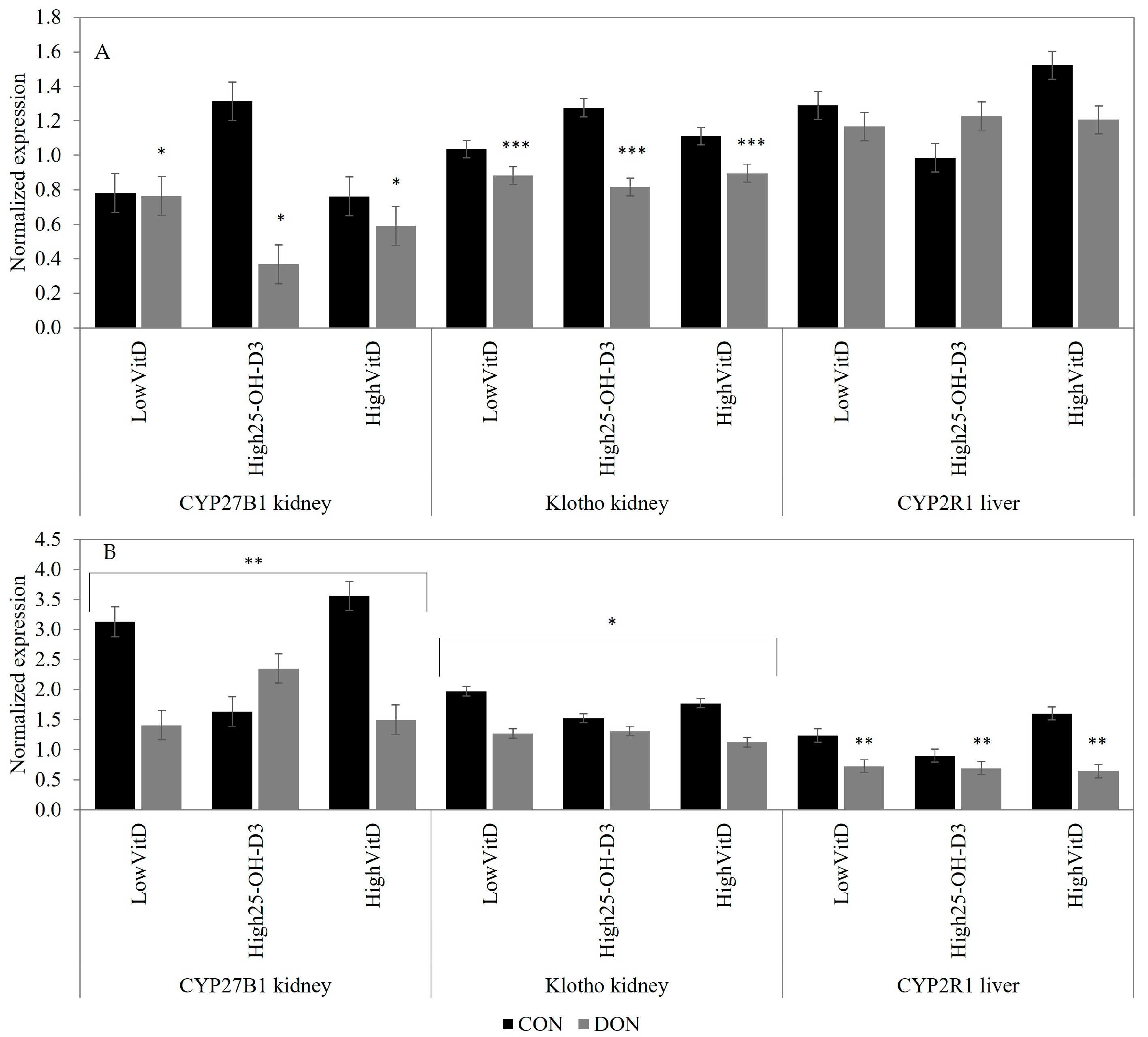
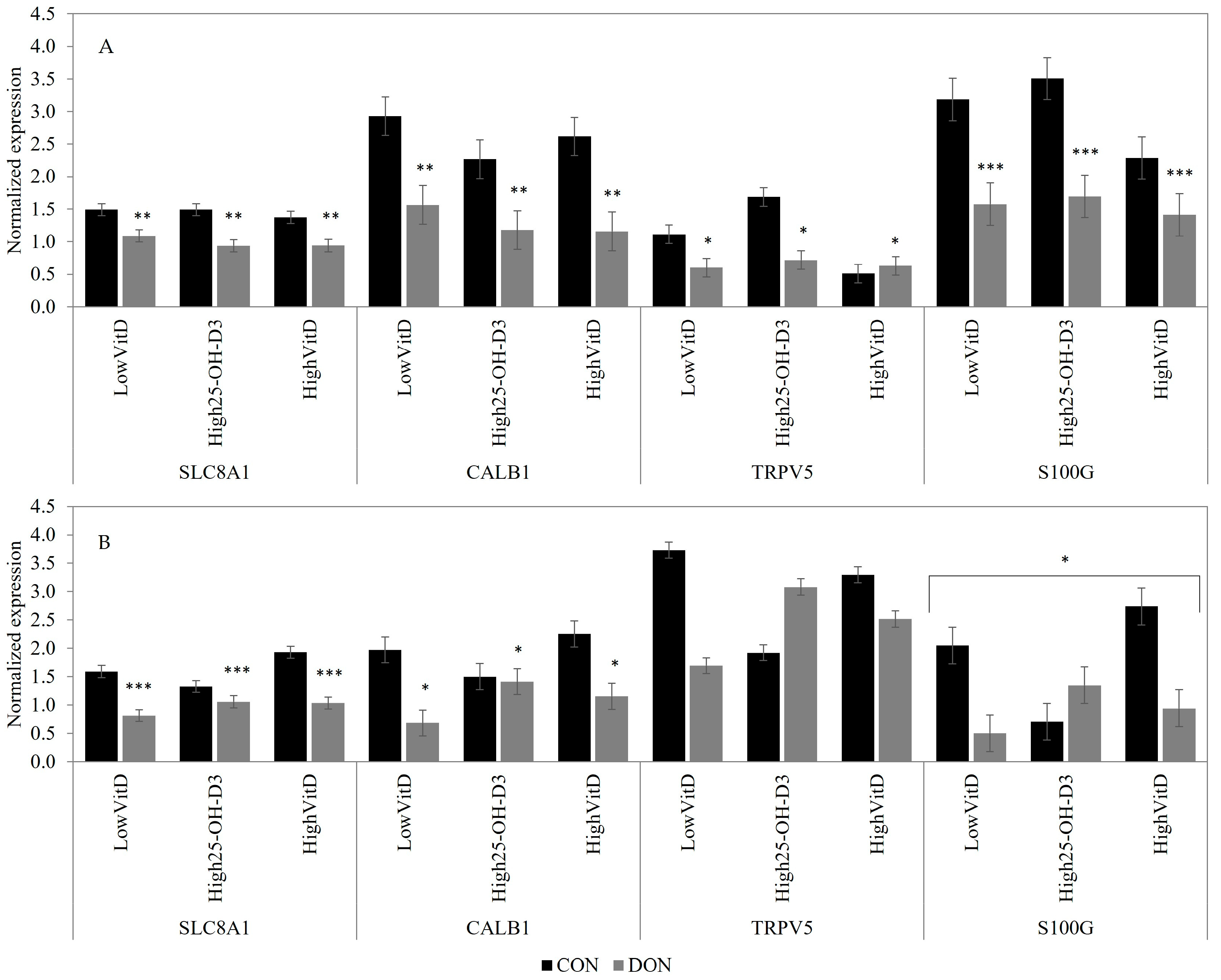
2.3. Gene Expression
3. Discussion
3.1. Impact of DON Contamination and Vitamin D Supplementation
3.2. Impact of DON and Vitamin D Supplement under LPS Challenge
4. Conclusions
5. Materials and Methods
5.1. Animals and Feeding Trial
5.2. Laboratory Analysis
5.2.1. Blood Analysis
5.2.2. Gene Expression Analysis
5.3. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control a | Control | Control | DON | DON | DON | |
|---|---|---|---|---|---|---|
| +HighVitD | +High25OHD | +HighVitD | +High25OHD | |||
| Ingredient (%) | ||||||
| Control wheat | 50.00 | 50.00 | 50.00 | |||
| Contaminated wheat | 50.00 | 50.00 | 50.00 | |||
| Corn | 14.06 | 14.06 | 14.06 | 14.06 | 14.06 | 14.06 |
| Soybean meal | 20.29 | 20.29 | 20.29 | 20.29 | 20.29 | 20.29 |
| Hamlet Soy Protein (HP300) | 3.03 | 3.03 | 3.03 | 3.03 | 3.03 | 3.03 |
| Choice white fat | 2.05 | 2.05 | 2.05 | 2.05 | 2.05 | 2.05 |
| Whey powder | 7.00 | 7.00 | 7.00 | 7.00 | 7.00 | 7.00 |
| Monocalcium phosphate | 0.88 | 0.88 | 0.88 | 0.88 | 0.88 | 0.88 |
| Limestone | 1.16 | 1.16 | 1.16 | 1.16 | 1.16 | 1.16 |
| Vitamin and mineral premix y,z | 0.50 | 0.50 | 0.50 | 0.50 | 0.50 | 0.50 |
| Salt | 0.30 | 0.30 | 0.30 | 0.30 | 0.30 | 0.30 |
| Lysine-HCl | 0.45 | 0.45 | 0.45 | 0.45 | 0.45 | 0.45 |
| DL-Methionine | 0.13 | 0.13 | 0.13 | 0.13 | 0.13 | 0.13 |
| L-Threonine | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 |
| Calculated composition (%) | ||||||
| Calcium | 0.85 | 0.85 | 0.85 | 0.85 | 0.85 | 0.85 |
| Phosphorus digestible | 0.42 | 0.42 | 0.42 | 0.42 | 0.42 | 0.42 |
| Analyzed composition | ||||||
| Calcium, % | 0.94 | 0.84 | 0.94 | 1.05 | 1.20 | 1.27 |
| Phosphorus, % | 0.65 | 0.58 | 0.64 | 0.60 | 0.76 | 0.61 |
| Vitamin D, IU/kg | 226 | 1802 | 209 | 170 | 2444 | 232 |
| 25-OH-D3 | --- | --- | 2156 | --- | --- | 2864 |
| Deoxynivalenol, mg/kg | 0.11 | 0.10 | 0.18 | 5.16 | 4.69 | 4.98 |
| Gene | Primer Sequence (5′-3′) | Product Size, bp | Genebank Accession No. |
|---|---|---|---|
| Vitamin D | |||
| CYP2R1 | (F) TTCATCCCTTCTTGACTCCAAC | 201 | XM_003480731 |
| (R) TTTATCTGTCACCTGTCACCAC | |||
| VDR | (F) AGGCTTCTTCAGACGGAGCAT | 143 | NM_001097414.1 |
| (R) ACTCCTTCATCATGCCGATGT | |||
| CYP27B1 | (F) TGGGCTCTCTATGAACTCTCTC | 157 | DQ295065.1 |
| (R) GTCTTAGCACTTCCTTGACCAC | |||
| Phosphorus | |||
| Klotho | (F) ACGCGGAACATGACGTACAG | 121 | XM_013989399.1 |
| (R) CCTGCAAGGCGATGGAGAT | |||
| SLC20A2 | (F) GTGCACCTGCTCTTCCACTTC | 106 | XM_005657658.1 |
| (R) ACAAAGCTACCAGAGGACCAATG | |||
| Calcium | |||
| S100G | (F) GAAGGAGGAGCTGAAGCAACTG | 139 | NM_214140.2 |
| (R) CACTAACACCTGGAATTCTTCAAAAC | |||
| CALB-1 | (F) TGGATCAGTATGGGCAAAGAGA | 133 | NM_001130226.1 |
| (R) GTCTTCATGAATTCCTCACAGGACTT | |||
| TRPV5 | (F) GCTGCGAGTACGTCGCTATGT | 86 | XM_003484001.2 |
| (R) CAGAGGGCTGTTTCTCAGAGAGA | |||
| SLC8A1 | (F) AATTGCTAGAGCTACTGTGTATTTCG | 136 | FJ268730.1 |
| (R) ATTGGGCTTCTTTATGGTTATTTCT | |||
| TRPV6 | (F) TGGGTGTCCCAAAGTCCAAG | 95 | XM_013985575.1 |
| (R) ACTCCCTCCTCCTCCCAAAT | |||
| Reference | |||
| GAPDH | (F) CCCCAACGTGTCGGTTGT | 91 | XM_021091114.1 |
| (R) CTCGGACGCCTGCTTCAC | |||
| β-Actin | (F) CATCACCATCGGCAACGA | 128 | XM_003357928.4 |
| (R) GGATGTCGACGTCGCACTT | |||
| HPRT | (F) TTGTGGTAGGCTATGCCCTTGACT | 117 | NM_001032376 |
| (R) CTCAACTTGAACTCTCCTCTTAGG |
References
- 5 Key Areas to Tackle High Swine Feed Costs. Available online: https://porkcheckoff.org/news/5-key-areas-to-tackle-high-swine-feed-costs/ (accessed on 15 August 2022).
- Woyengo, T.A.; Beltranena, E.; Zijlstra, R.T. Nonruminant Nutrition Symposium: Controlling feed cost by including alternative ingredients into pig diets: A review. J. Anim. Sci. 2014, 92, 1293–1305. [Google Scholar] [CrossRef]
- Pestka, J.J. Deoxynivalenol: Toxicity, mechanisms and animal health risks. Anim. Feed Sci. Technol. 2007, 137, 283–298. [Google Scholar] [CrossRef]
- Döll, S.; Schrickx, J.A.; Dänicke, S.; Fink-Gremmels, J. Interactions of deoxynivalenol and lipopolysaccharides on cytokine excretion and mRNA expression in porcine hepatocytes and Kupffer cell enriched hepatocyte cultures. Toxicol. Lett. 2009, 190, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.; Dwivedi, P.D.; Pandey, H.P.; Das, M. Role of oxidative stress in Deoxynivalenol induced toxicity. Food Chem. Toxicol. 2014, 72, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Sobrova, P.; Adam, V.; Vasatkova, A.; Beklova, M.; Zeman, L.; Kizek, R. Deoxynivalenol and its toxicity. Interdiscip. Toxicol. 2010, 3, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Zhang, L.; Xu, Z.; Liu, X.; Chen, L.; Dai, J.; Karrow, N.A.; Sun, L. Occurrence of Aflatoxin B1, deoxynivalenol and zearalenone in feeds in China during 2018–2020. J. Anim. Sci. Biotechnol. 2021, 12, 74. [Google Scholar] [CrossRef] [PubMed]
- Le Thanh, B.V.; Lessard, M.; Chorfi, Y.; Guay, F. The efficacy of anti-mycotoxin feed additives in preventing the adverse effects of wheat naturally contaminated with Fusarium mycotoxins on performance, intestinal barrier function and nutrient digestibility and retention in weanling pigs. Can. J. Anim. Sci. 2015, 95, 197–209. [Google Scholar] [CrossRef]
- Reddy, K.E.; Lee, W.; Jeong, J.y.; Lee, Y.; Lee, H.-J.; Kim, M.S.; Kim, D.-W.; Yu, D.; Cho, A.; Oh, Y.K.; et al. Effects of deoxynivalenol- and zearalenone-contaminated feed on the gene expression profiles in the kidneys of piglets. Asian-Australas. J. Anim. Sci. 2018, 31, 138–148. [Google Scholar] [CrossRef] [PubMed]
- Serviento, A.M.; Brossard, L.; Renaudeau, D. An acute challenge with a deoxynivalenol-contaminated diet has short- and long-term effects on performance and feeding behavior in finishing pigs. J. Anim. Sci. 2018, 96, 5209–5221. [Google Scholar] [CrossRef]
- Accensi, F.; Pinton, P.; Callu, P.; Abella-Bourges, N.; Guelfi, J.-F.; Grosjean, F.; Oswald, I.P. Ingestion of low doses of deoxynivalenol does not affect hematological, biochemical, or immune responses of piglets1. J. Anim. Sci. 2006, 84, 1935–1942. [Google Scholar] [CrossRef]
- Becker, C.; Reiter, M.; Pfaffl, M.W.; Meyer, H.H.D.; Bauer, J.; Meyer, K.H.D. Expression of immune relevant genes in pigs under the influence of low doses of deoxynivalenol (DON). Mycotoxin Res. 2011, 27, 287–293. [Google Scholar] [CrossRef] [PubMed]
- Le Thanh, B.V.; Lemay, M.; Bastien, A.; Lapointe, J.; Lessard, M.; Chorfi, Y.; Guay, F. The potential effects of antioxidant feed additives in mitigating the adverse effects of corn naturally contaminated with Fusarium mycotoxins on antioxidant systems in the intestinal mucosa, plasma, and liver in weaned pigs. Mycotoxin Res. 2016, 32, 99–116. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Zhang, L.; Mo, Y.; Li, J.; Yang, J.; Wang, J.; Karrow, N.A.; Wu, H.; Sun, L. Ferroptosis is involved in deoxynivalenol-induced intestinal damage in pigs. J. Anim. Sci. Biotechnol. 2023, 14, 29. [Google Scholar] [CrossRef] [PubMed]
- Sassi, F.; Tamone, C.; D’Amelio, P. Vitamin D: Nutrient, Hormone, and Immunomodulator. Nutrients 2018, 10, 1656. [Google Scholar] [CrossRef] [PubMed]
- Christakos, S.; Ajibade, D.V.; Dhawan, P.; Fechner, A.J.; Mady, L.J. Vitamin D: Metabolism. Endocrinol. Metab. Clin. N. Am. 2010, 39, 243–253. [Google Scholar] [CrossRef]
- Combs, G.F. Chapter 6—Vitamin D. In The Vitamins, 4th ed.; Combs, G.F., Ed.; Academic Press: San Diego, CA, USA, 2012; pp. 139–180. [Google Scholar]
- Flynn, A. The role of dietary calcium in bone health. Proc. Nutr. Soc. 2003, 62, 851–858. [Google Scholar] [CrossRef]
- Fukumoto, S. Phosphate metabolism and vitamin D. BoneKEy Rep. 2014, 3, 497. [Google Scholar] [CrossRef]
- Nussey, S.; Whitehead, S.A. The parathyroid glands and vitamin D. In Endocrinology: An Integrated Approach, 1st ed.; BIOS Scientific Publishers: Oxford, UK; London, UK, 2001; pp. 171–215. [Google Scholar]
- Haussler, M.R.; Whitfield, G.K.; Haussler, C.A.; Jurutka, P.W. The role of vitamin D in the FGF23, klotho, and phosphate bone-kidney endocrine axis. Rev. Endocr. Metab. Disord. 2012, 13, 57–69. [Google Scholar] [CrossRef]
- van de Graaf, S.F.J.; Boullart, I.; Hoenderop, J.G.J.; Bindels, R.J.M. Regulation of the epithelial Ca2+ channels TRPV5 and TRPV6 by 1α,25-dihydroxy Vitamin D3 and dietary Ca2+. J. Steroid Biochem. Mol. Biol. 2004, 89–90, 303–308. [Google Scholar] [CrossRef]
- Zou, D.; Wu, W.; He, Y.; Ma, S.; Gao, J. The role of klotho in chronic kidney disease. BMC Nephrol. 2018, 19, 285. [Google Scholar] [CrossRef]
- Sergeev, I.N.; Kravchenko, L.V.; Piliia, N.M.; Batukhanov, A.B.; Sobolev, V.S.; Kuz’mina, E.E.; Iakushina, L.M.; Spirichev, V.B.; Tutel’ian, V.A. The effect of the trichothecene mycotoxin deoxynivalenol (vomitoxin) on calcium homeostasis, vitamin D metabolism and receptors in rats. Vopr. Med. Khim. 1990, 36, 26–29. [Google Scholar] [PubMed]
- McCue, M.; Reichert, J.L.; Crenshaw, T.D. Impact of dietary vitamin D3 supplements in nursery diets on subsequent growth and bone responses of pigs during an immune challenge. J. Anim. Sci. 2019, 97, 4895–4903. [Google Scholar] [CrossRef]
- Witschi, A.-K.M.; Liesegang, A.; Gebert, S.; Weber, G.M.; Wenk, C. Effect of source and quantity of dietary vitamin D in maternal and creep diets on bone metabolism and growth in piglets1. J. Anim. Sci. 2011, 89, 1844–1852. [Google Scholar] [CrossRef] [PubMed]
- Burild, A.; Lauridsen, C.; Faqir, N.; Sommer, H.M.; Jakobsen, J. Vitamin D3 and 25-hydroxyvitamin D3 in pork and their relationship to vitamin D status in pigs. J. Nutr. Sci. 2016, 5, e3. [Google Scholar] [CrossRef] [PubMed]
- Madsen, P.A.; Etheve, S.; Heegaard, P.M.H.; Skovgaard, K.; Mary, A.L.; Litta, G.; Lauridsen, C. Influence of vitamin D metabolites on vitamin D status, immunity and gut health of piglets. Vet. Immunol. Immunopathol. 2023, 257, 110557. [Google Scholar] [CrossRef]
- Pinton, P.; Braicu, C.; Nougayrede, J.-P.; Laffitte, J.; Taranu, I.; Oswald, I.P. Deoxynivalenol Impairs Porcine Intestinal Barrier Function and Decreases the Protein Expression of Claudin-4 through a Mitogen-Activated Protein Kinase-Dependent Mechanism. J. Nutr. 2010, 140, 1956–1962. [Google Scholar] [CrossRef]
- Klunker, L.R.; Kahlert, S.; Panther, P.; Diesing, A.K.; Reinhardt, N.; Brosig, B.; Kersten, S.; Dänicke, S.; Rothkötter, H.J.; Kluess, J.W. Deoxynivalenol and E.coli lipopolysaccharide alter epithelial proliferation and spatial distribution of apical junction proteins along the small intestinal axis. J. Anim. Sci. 2013, 91, 276–285. [Google Scholar] [CrossRef]
- Kullik, K.; Brosig, B.; Kersten, S.; Valenta, H.; Diesing, A.K.; Panther, P.; Reinhardt, N.; Kluess, J.; Rothkötter, H.J.; Breves, G.; et al. Interactions between the Fusarium toxin deoxynivalenol and lipopolysaccharides on the in vivo protein synthesis of acute phase proteins, cytokines and metabolic activity of peripheral blood mononuclear cells in pigs. Food Chem. Toxicol. 2013, 57, 11–20. [Google Scholar] [CrossRef]
- Dänicke, S.; Brosig, B.; Kahlert, S.; Panther, P.; Reinhardt, N.; Diesing, A.-K.; Kluess, J.; Kersten, S.; Valenta, H.; Rothkötter, H.-J. The plasma clearance of the Fusarium toxin deoxynivalenol (DON) is decreased in endotoxemic pigs. Food Chem. Toxicol. 2012, 50, 4405–4411. [Google Scholar] [CrossRef]
- Renner, L.; Kahlert, S.; Tesch, T.; Bannert, E.; Frahm, J.; Barta-Böszörményi, A.; Kluess, J.; Kersten, S.; Schönfeld, P.; Rothkötter, H.-J.; et al. Chronic DON exposure and acute LPS challenge: Effects on porcine liver morphology and function. Mycotoxin Res. 2017, 33, 207–218. [Google Scholar] [CrossRef]
- Dänicke, S.; Valenta, H.; Ganter, M.; Brosig, B.; Kersten, S.; Diesing, A.-K.; Kahlert, S.; Panther, P.; Kluess, J.; Rothkötter, H.-J. Lipopolysaccharides (LPS) modulate the metabolism of deoxynivalenol (DON) in the pig. Mycotoxin Res. 2014, 30, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Islam, Z.; Pestka, J.J. LPS priming potentiates and prolongs proinflammatory cytokine response to the trichothecene deoxynivalenol in the mouse. Toxicol. Appl. Pharmacol. 2006, 211, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Lessard, M.; Savard, C.; Deschene, K.; Lauzon, K.; Pinilla, V.A.; Gagnon, C.A.; Lapointe, J.; Guay, F.; Chorfi, Y. Impact of deoxynivalenol (DON) contaminated feed on intestinal integrity and immune response in swine. Food Chem. Toxicol. 2015, 80, 7–16. [Google Scholar] [CrossRef]
- Andretta, I.; Kipper, M.; Lehnen, C.R.; Hauschild, L.; Vale, M.M.; Lovatto, P.A. Meta-analytical study of productive and nutritional interactions of mycotoxins in growing pigs. Animal 2012, 6, 1476–1482. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Bates, M.A.; Bursian, S.J.; Flannery, B.; Zhou, H.-R.; Link, J.E.; Zhang, H.; Pestka, J.J. Peptide YY3–36 and 5-Hydroxytryptamine Mediate Emesis Induction by Trichothecene Deoxynivalenol (Vomitoxin). Toxicol. Sci. 2013, 133, 186–195. [Google Scholar] [CrossRef]
- Wang, X.; Chen, X.; Cao, L.; Zhu, L.; Zhang, Y.; Chu, X.; Zhu, D.; Rahman, S.u.; Peng, C.; Feng, S.; et al. Mechanism of deoxynivalenol-induced neurotoxicity in weaned piglets is linked to lipid peroxidation, dampened neurotransmitter levels, and interference with calcium signaling. Ecotoxicol. Environ. Saf. 2020, 194, 110382. [Google Scholar] [CrossRef]
- Wu, W.; Zhou, H.-R.; Bursian, S.J.; Link, J.E.; Pestka, J.J. Calcium-Sensing Receptor and Transient Receptor Ankyrin-1 Mediate Emesis Induction by Deoxynivalenol (Vomitoxin). Toxicol. Sci. 2016, 155, 32–42. [Google Scholar] [CrossRef]
- Sims, N.A.; Martin, T.J. Coupling the activities of bone formation and resorption: A multitude of signals within the basic multicellular unit. BoneKEy Rep. 2014, 3, 481. [Google Scholar] [CrossRef]
- Goltzman, D. Functions of vitamin D in bone. Histochem. Cell Biol. 2018, 149, 305–312. [Google Scholar] [CrossRef]
- Suzuki, Y.; Landowski, C.P.; Hediger, M.A. Mechanisms and Regulation of Epithelial Ca2+ Absorption in Health and Disease. Annu. Rev. Physiol. 2008, 70, 257–271. [Google Scholar] [CrossRef]
- Pu, F.; Chen, N.; Xue, S. Calcium intake, calcium homeostasis and health. Food Sci. Hum. Wellness 2016, 5, 8–16. [Google Scholar] [CrossRef]
- Khananshvili, D. The SLC8 gene family of sodium-calcium exchangers (NCX)—Structure, function, and regulation in health and disease. Mol. Asp. Med. 2013, 34, 220–235. [Google Scholar] [CrossRef] [PubMed]
- Payros, D.; Alassane-Kpembi, I.; Pierron, A.; Loiseau, N.; Pinton, P.; Oswald, I.P. Toxicology of deoxynivalenol and its acetylated and modified forms. Arch. Toxicol. 2016, 90, 2931–2957. [Google Scholar] [CrossRef] [PubMed]
- Clinkenbeard, E.L.; White, K.E. Systemic Control of Bone Homeostasis by FGF23 Signaling. Curr. Mol. Biol. Rep. 2016, 2, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Bian, A.; Xing, C.; Hu, M.C. Alpha Klotho and phosphate homeostasis. J. Endocrinol. Investig. 2014, 37, 1121–1126. [Google Scholar] [CrossRef]
- Forster, I.C.; Hernando, N.; Biber, J.; Murer, H. Phosphate transporters of the SLC20 and SLC34 families. Mol. Aspects Med. 2013, 34, 386–395. [Google Scholar] [CrossRef]
- Lütke-Dörhoff, M.; Schulz, J.; Westendarp, H.; Visscher, C.; Wilkens, M.R. Dietary supplementation of 25-hydroxycholecalciferol as an alternative to cholecalciferol in swine diets: A review. J. Anim. Physiol. Anim. Nutr. 2022, 106, 1288–1305. [Google Scholar] [CrossRef]
- O’Doherty, J.V.; Gahan, D.A.; O’Shea, C.; Callan, J.J.; Pierce, K.M. Effects of phytase and 25-hydroxyvitamin D3 inclusions on the performance, mineral balance and bone parameters of grower–finisher pigs fed low-phosphorus diets. Animal 2010, 4, 1634–1640. [Google Scholar] [CrossRef]
- Regassa, A.; Adhikari, R.; Nyachoti, C.M.; Kim, W.K. Effects of 25-(OH)D3 on fecal Ca and P excretion, bone mineralization, Ca and P transporter mRNA expression and performance in growing female pigs. J. Environ. Sci. Health B 2015, 50, 293–299. [Google Scholar] [CrossRef]
- Upadhaya, S.D.; Chung, T.K.; Jung, Y.J.; Kim, I.H. Dietary 25(OH)D3 supplementation to gestating and lactating sows and their progeny affects growth performance, carcass characteristics, blood profiles and myogenic regulatory factor-related gene expression in wean-finish pigs. Anim. Biosci. 2022, 35, 461–474. [Google Scholar] [CrossRef]
- Fujita, H.; Sugimoto, K.; Inatomi, S.; Maeda, T.; Osanai, M.; Uchiyama, Y.; Yamamoto, Y.; Wada, T.; Kojima, T.; Yokozaki, H.; et al. Tight Junction Proteins Claudin-2 and -12 Are Critical for Vitamin D-dependent Ca2+ Absorption between Enterocytes. Mol. Biol. Cell 2008, 19, 1912–1921. [Google Scholar] [CrossRef] [PubMed]
- Jorde, R.; Grimnes, G. Increased calcium intake is associated lower serum 25-hydroxyvitamin D levels in subjects with adequate vitamin D intake: A population-based observational study. BMC Nutr. 2020, 6, 49. [Google Scholar] [CrossRef] [PubMed]
- Christakos, S.; Lieben, L.; Masuyama, R.; Carmeliet, G. Vitamin D endocrine system and the intestine. BoneKEy Rep. 2014, 3, 496. [Google Scholar] [CrossRef] [PubMed]
- Stanek, C.; Reinhardt, N.; Diesing, A.-K.; Nossol, C.; Kahlert, S.; Panther, P.; Kluess, J.; Rothkötter, H.-J.; Kuester, D.; Brosig, B.; et al. A chronic oral exposure of pigs with deoxynivalenol partially prevents the acute effects of lipopolysaccharides on hepatic histopathology and blood clinical chemistry. Toxicol. Lett. 2012, 215, 193–200. [Google Scholar] [CrossRef] [PubMed]
- González-Vega, J.C.; Stein, H.H. Calcium Digestibility and Metabolism in Pigs. Asian-Australas. J. Anim. Sci. 2014, 27, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Holowaychuk, M.K.; Birkenheuer, A.J.; Li, J.; Marr, H.; Boll, A.; Nordone, S.K. Hypocalcemia and Hypovitaminosis D in Dogs with Induced Endotoxemia. J. Vet. Intern. Med. 2012, 26, 244–251. [Google Scholar] [CrossRef]
- Carlstedt, F.; Eriksson, M.; Kiiski, R.; Larsson, A.; Lind, L. Hypocalcemia during porcine endotoxemic shock: Effects of calcium administration. Crit. Care Med. 2000, 28, 2909–2914. [Google Scholar] [CrossRef]
- Xu, S.; Chen, Y.-H.; Tan, Z.-X.; Xie, D.-D.; Zhang, C.; Xia, M.-Z.; Wang, H.; Zhao, H.; Xu, D.-X.; Yu, D.-X. Vitamin D3 pretreatment alleviates renal oxidative stress in lipopolysaccharide-induced acute kidney injury. J. Steroid Biochem. Mol. Biol. 2015, 152, 133–141. [Google Scholar] [CrossRef]
- Zúñiga, S.; Firrincieli, D.; Housset, C.; Chignard, N. Vitamin D and the vitamin D receptor in liver pathophysiology. Clin. Res. Hepatol. Gastroenterol. 2011, 35, 295–302. [Google Scholar] [CrossRef]
- Dong, B.; Zhou, Y.; Wang, W.; Scott, J.; Kim, K.; Sun, Z.; Guo, Q.; Lu, Y.; Gonzales, N.M.; Wu, H.; et al. Vitamin D Receptor Activation in Liver Macrophages Ameliorates Hepatic Inflammation, Steatosis, and Insulin Resistance in Mice. Hepatology 2020, 71, 1559–1574. [Google Scholar] [CrossRef]
- National Research Council. Nutrient Requirements of Swine: Eleventh Revised Edition; The National Academies Press: Washington, DC, USA, 2012; p. 420.
- Isabel, B.; Rey, A.; López, B. Optimum vitamin nutrition in pigs. In Optimum Vitamin Nutrition—In the Production of Quality Animal Foods; DSM Nutritional Products Limited; 5M Publishing: Sheffield, UK, 2012; pp. 243–308. [Google Scholar]
- Kim, B.G.; Lindemann, M.D. A spreadsheet method for experimental animal allotment. J. Anim. Sci. 2007, 85, 112. [Google Scholar]






| Deoxynivalenol | Vitamin D | p Value | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Parameters | CON | DON | Low VitD | High25-OH-D3 | High VitD | SEM | SD | DON | VitD |
| ADG, g/d | 555.4 | 358.1 | 447.2 | 459.1 | 466.4 | 19.4 | 116.6 | 0.001 | 0.66 |
| ADFI, g/d | 759.8 | 493.5 | 611.0 | 619.6 | 649.3 | 29.3 | 156.3 | 0.001 | 0.51 |
| BMC/BW (%) | 1.28 | 1.41 | 1.34 | 1.35 | 1.34 | 0.02 | 0.10 | 0.001 | 0.93 |
| BMC, g | 253.5 | 220.3 | 234.1 | 234.9 | 241.4 | 10.1 | 40.3 | 0.001 | 0.71 |
| G:F | 0.73 | 0.73 | 0.73 | 0.74 | 0.72 | 0.01 | 0.05 | 0.77 | 0.29 |
| Initial body weight, kg | 7.79 | 7.97 | 7.96 | 7.69 | 7.98 | 0.30 | 1.09 | 0.43 | 0.50 |
| Final body weight, kg | 19.8 | 15.7 | 17.6 | 17.6 | 18.0 | 0.61 | 3.26 | 0.001 | 0.72 |
| Item | DON, ng/mL | DOM-1, ng/mL | Calcium, mM | Magnesium, μM | |||||
|---|---|---|---|---|---|---|---|---|---|
| Treatments | |||||||||
| Deoxynivalenol | Vitamin D | Before LPS | After LPS | Before LPS | After LPS | Before LPS | After LPS | Before LPS | After LPS |
| CON | LowVitD | 0.21 | 1.50 | 0.00 | 0.00 | 3.02 | 2.66 | 559.53 | 594.30 |
| CON | High25-OH-D3 | 2.31 | 0.59 | 0.00 | 0.00 | 2.91 | 2.83 | 569.41 | 679.71 |
| CON | HighVitD | 0.90 | 0.63 | 0.00 | 0.00 | 2.99 | 2.68 | 660.50 | 662.89 |
| DON | LowVitD | 23.55 | 18.31 | 4.68 | 4.94 | 2.76 | 2.64 | 521.11 | 630.23 |
| DON | High25-OH-D3 | 23.16 | 19.07 | 3.33 | 4.09 | 2.94 | 2.64 | 589.56 | 504.17 |
| DON | HighVitD | 30.62 | 20.28 | 4.24 | 5.27 | 2.90 | 2.42 | 601.80 | 613.01 |
| SEM | 2.05 | 1.96 | 0.23 | 0.31 | 0.12 | 0.17 | 56.78 | 60.02 | |
| SD | 13.42 | 11.45 | 2.27 | 2.47 | 0.39 | 0.45 | 150.8 | 165.1 | |
| Mixed Procedure | n = 32 | n = 32 | n = 33 | n = 33 | n = 33 | n = 33 | n = 33 | n = 33 | |
| DON | <0.001 | <0.001 | <0.001 | <0.001 | NS | NS | NS | NS | |
| VitD | NS | NS | 0.09 | NS | NS | NS | NS | NS | |
| DON × VitD | NS | NS | 0.09 | NS | NS | NS | NS | NS | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sauvé, B.; Chorfi, Y.; Montminy, M.-P.L.; Guay, F. Vitamin D Supplementation Impacts Calcium and Phosphorus Metabolism in Piglets Fed a Diet Contaminated with Deoxynivalenol and Challenged with Lipopolysaccharides. Toxins 2023, 15, 394. https://doi.org/10.3390/toxins15060394
Sauvé B, Chorfi Y, Montminy M-PL, Guay F. Vitamin D Supplementation Impacts Calcium and Phosphorus Metabolism in Piglets Fed a Diet Contaminated with Deoxynivalenol and Challenged with Lipopolysaccharides. Toxins. 2023; 15(6):394. https://doi.org/10.3390/toxins15060394
Chicago/Turabian StyleSauvé, Béatrice, Younes Chorfi, Marie-Pierre Létourneau Montminy, and Frédéric Guay. 2023. "Vitamin D Supplementation Impacts Calcium and Phosphorus Metabolism in Piglets Fed a Diet Contaminated with Deoxynivalenol and Challenged with Lipopolysaccharides" Toxins 15, no. 6: 394. https://doi.org/10.3390/toxins15060394
APA StyleSauvé, B., Chorfi, Y., Montminy, M.-P. L., & Guay, F. (2023). Vitamin D Supplementation Impacts Calcium and Phosphorus Metabolism in Piglets Fed a Diet Contaminated with Deoxynivalenol and Challenged with Lipopolysaccharides. Toxins, 15(6), 394. https://doi.org/10.3390/toxins15060394