1. Introduction
Clostridium perfringens is a kind of anaerobic Gram-positive bacterium that widely exists in the intestinal tissue of humans and animals. It is one of the main pathogenic bacteria that induce necrotizing enteritis and enterotoxamia, which pose a serious threat to animal husbandry production [
1]. The bacterium can ferment sugars such as glucose, lactose, and sucrose, and produce large amounts of gas within tissues, leading to it being named perfringens, which is from the Latin for through (per) and burst (frango). In 1891, during an autopsy on a 38-year-old man, William H. Welch was the first to isolate
Clostridium perfringens and report on its characteristics [
2]. The virulence of
Clostridium perfringens is largely attributable to its ability to produce toxins. At present, 20 types of exotoxins and enzymes produced by
Clostridium perfringens have been identified [
3]. In early investigations,
Clostridium perfringens was divided into five types according to the four toxins produced [
4]. In 1892, Welchii et al. isolated
Clostridium perfringens type A from decayed cadavers, followed by types B (1926), C (1929), D (1932), and E (1943). With further investigation,
Clostridium perfringens enterotoxin (CPE) and necrotizing enterotoxin B (NetB) were found to play important roles in diseases caused by
Clostridium perfringens, so a new classification scheme needed to be developed. In 2018, two new types of
Clostridium perfringens were added, namely, type F and type G, and thus the classification of the bacterium was extended to seven types (A to G) [
5]. At present,
Clostridium perfringens is known to produce α (CPA), β1 (CPB1), ε (ETX), ι (ITX), CPE, Perfringolysin O (PFO), β2 (CPB2), NetB, Kappa, Delta, and other kinds of toxin [
3]. Among them, CPA, CPB, ETX, ITX, CPE, and NetB are the most influential and lethal toxins and can cause tissue, nervous system, and intestinal toxicity.
Studies have shown that
Clostridium perfringens beta-1 toxin (CPB1) is encoded by the plasmid-encoded gene
cpb1. The gene
cpb1 is about 1.0 Kb in length, and encodes CPB1 protein with a molecular weight of approximately 34 kDa [
6]. CPB1 is produced by
Clostridium perfringens types B and C, of which type C is a highly destructive pathogen [
7]. The disease known as Darmbrand [
8] appeared after the World War II, and was determined to be necrotizing enteritis caused by
Clostridia perfringens type C infection; this disease was also an important reason for the high mortality rate of children in Papua New Guinea [
9]. CPB1 is an important virulence factor for hemorrhagic intestinal necrosis induced by
Clostridium perfringens type C [
7], mainly occurring in neonates of many animal species (especially in newborn animals) and human beings [
10].
As a form of programmed cell death, pyroptosis has attracted increasing attention in recent years because of the occurrence and development of diseases, and has become a hotspot in the field of life science [
11]. There are many types of exogenous stimuli that can cause pyroptosis, in the canonical inflammasome activation pathway, [
12] inflammasomes [
13] such as NLR Family Pyrin Domain Containing 3 (NLRP3), which can sense bacteria, viruses, toxins, and ATP stimuli [
14]. These stimuli indirectly activate NLRP3 through K
+ outflow and lead to the binding of NIMA-related kinase 7 to NLRP3, thereby triggering the oligomerization of NLRP3. NLRP3 is then activated by an apoptosis-associated speck-like protein containing a CARD, which in turn activates cysteinyl aspartate specific proteinase 1 (Caspase 1) to produce interleukin (IL)-18 and IL-1β, and shear gasdermin D (GSDMD) to release the GSDMD-N domain, a peptide with the ability to form membrane pores, ultimately resulting in the secretion of activated IL-18 and IL-1β [
15]. This signaling cascade of the NLRP3 and Caspase-1-dependent activation of pyroptosis is called the canonical pathway. Since 1995, a number of studies have found that Caspase 11 in mouse, as well as Caspase 45 in human cells [
16,
17], can induce the death of macrophages in response to a variety of Gram-negative bacterial infections, and the result is similar to the pyroptosis induced by Caspase 1 in morphology. This is called the non-canonical pathway [
18]. Both pyroptosis and apoptosis were found to result in nuclear enrichment and chromosome DNA breakage [
19,
20], and TUNEL and Annexin V staining were positive. Differing from apoptosis, pyroptosis can lead to the formation of membrane pores and the destruction of membrane integrity, resulting in the release of cell contents, an increase in permeability, a release of inflammatory factors, and thus the induction of an inflammatory response.
Currently, accumulating studies have shown that, after infection or injury, K+ outflow activates the NLRP3 inflammasome, which leads to pyroptosis. As a perforated toxin, CPB1 can affect the membrane permeability of THP-1 cells. With the increase in CPB1 concentration and prolonged action, the release of K+ also increases. When the toxin was subcutaneously injected, it caused the rapid exudation of plasma and a significant increase in IL-1β and TNF-α at the injection site. Based on the above findings, we aimed to ascertain whether CPB1 induces pyroptosis by activating the inflammasome of stimulated cells through the Caspase-1-dependent pathway or the Caspase-4/5/11 pathway. We also attempted to investigate whether the release of inflammatory factors induced by CPB1 caused pyroptosis. Therefore, pyroptosis of macrophages induced by CPB1 was explored, and the findings will provide new insights into the pathogenic mechanisms of CPB1.
3. Discussion
Since 1965, extensive studies have been carried out to clarify the significance and mechanisms of programmed cell death. It has now been established that cells can undergo programmed death during development due to pathogen invasion or cell stress and metabolic disorders. Three programmed cell death pathways have been studied in depth in recent years: apoptosis, autophagy, and pyroptosis [
21]. Apoptosis and autophagy do not produce an inflammatory response, while pyroptosis does [
22,
23]. Pyroptosis is a “double-edged sword” within the anti-infection immune processes, related to the inflammatory mechanism of the host. On the one hand, pyroptosis can activate the internal mechanism of cell death, release inflammatory factors, clear pathogens, and prevent infection. On the other hand, excessive Caspase 1 activation can lead to pathological inflammation. Although the onset of inflammation has a protective effect, excessive inflammation is harmful.
Pyroptosis pathogenesis can be divided into two pathways. Pyroptosis mediated by Caspase 1 and activated by inflammasomes is called the canonical pathway, while that mediated by Caspase-4/5 (in humans) or Caspase-11 (in mice) is called the non-canonical pathway. Pyroptosis can lead to the release of cellular contents, included damage-associated molecular patterns (DAMPs) and inflammatory factors (IL-1β and IL-18), causing inflammation [
24].
Clostridium perfringens (
C. perfringens) is a bacterium that exists widely in the environment, and also presents as normal intestinal flora in humans and domestic animals.
C. perfringens can infect animals and humans by the ingestion of spores or through changes in the gut flora, followed by excessive growth of bacteria and toxin production, which causes enterotoxemia, necrotic hemorrhagic enteritis, and enterocolitis [
25].
C. perfringens beta1 toxin (CPB1) is a lethal toxin secreted by types B and C. It is able to form membrane pores on susceptible cells, leading to cell distension and lysis. As a perforating toxin, CPB1 can affect the membrane permeability of THP-1 cells. With an increase in its concentration and the prolongation of the action time, the release of K+ in the cells will also increase. Interestingly, several studies have reported that the K+ efflux or Ca2+ flux from cells can induce NLRP3 inflammasome activation and assembly. It is well known that the assembly of the NLRP3 activates Caspase 1, and Caspase 1 activation leads to cytokine secretion. In some circumstances, it results in pyroptosis. Therefore, we speculate that CPB1 toxin maybe induce pyroptosis by causing K+ efflux from cells.
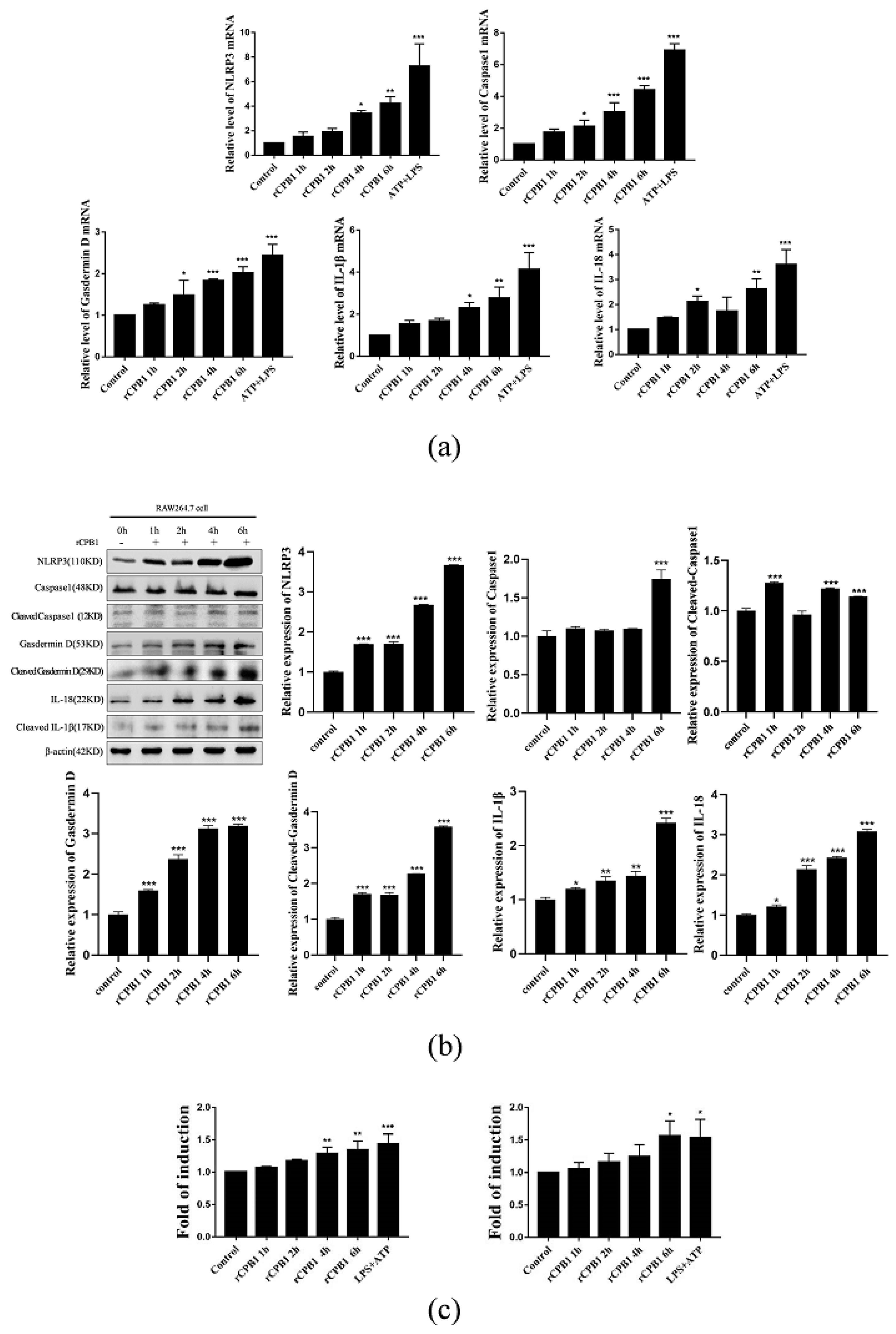
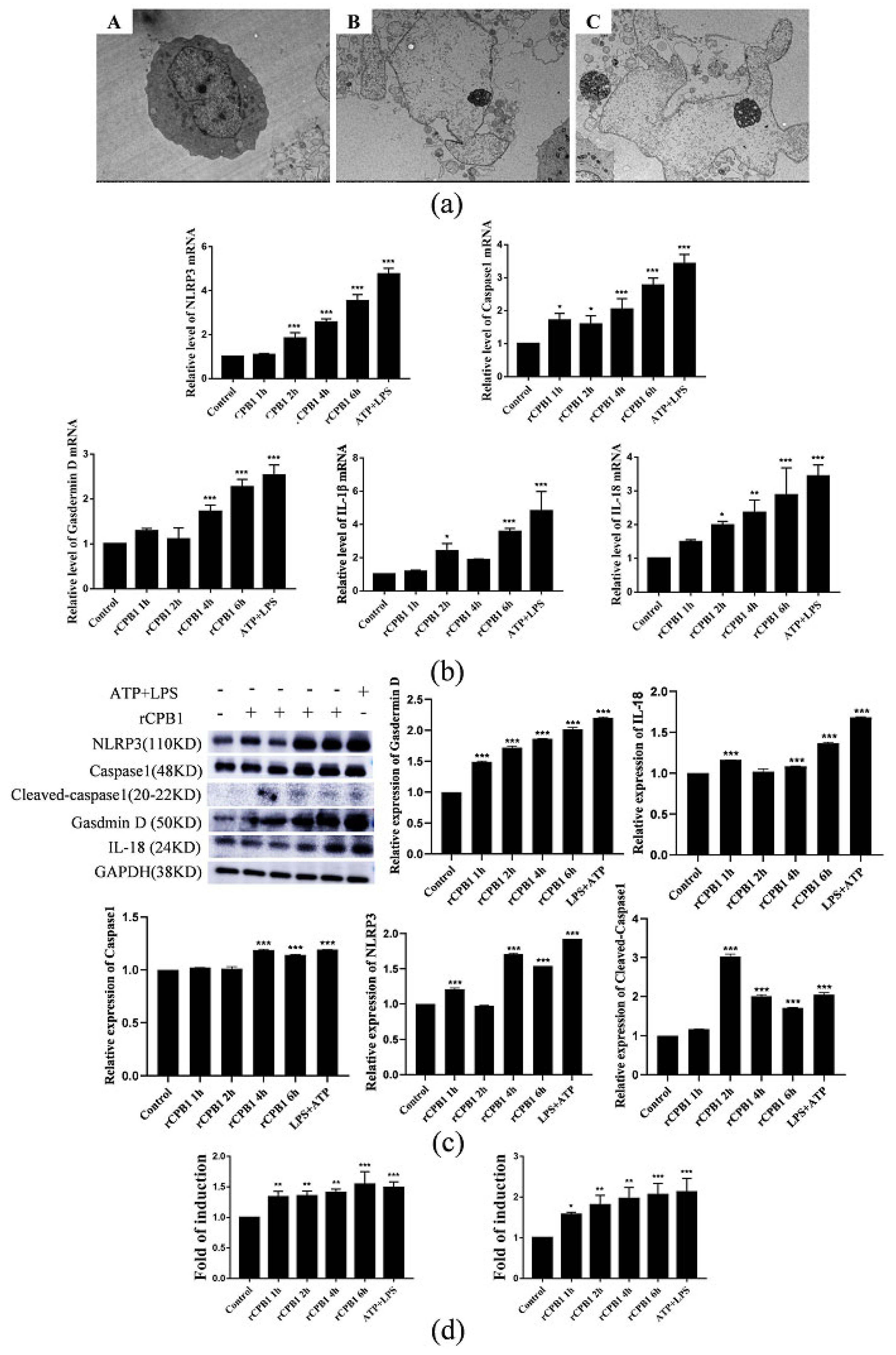
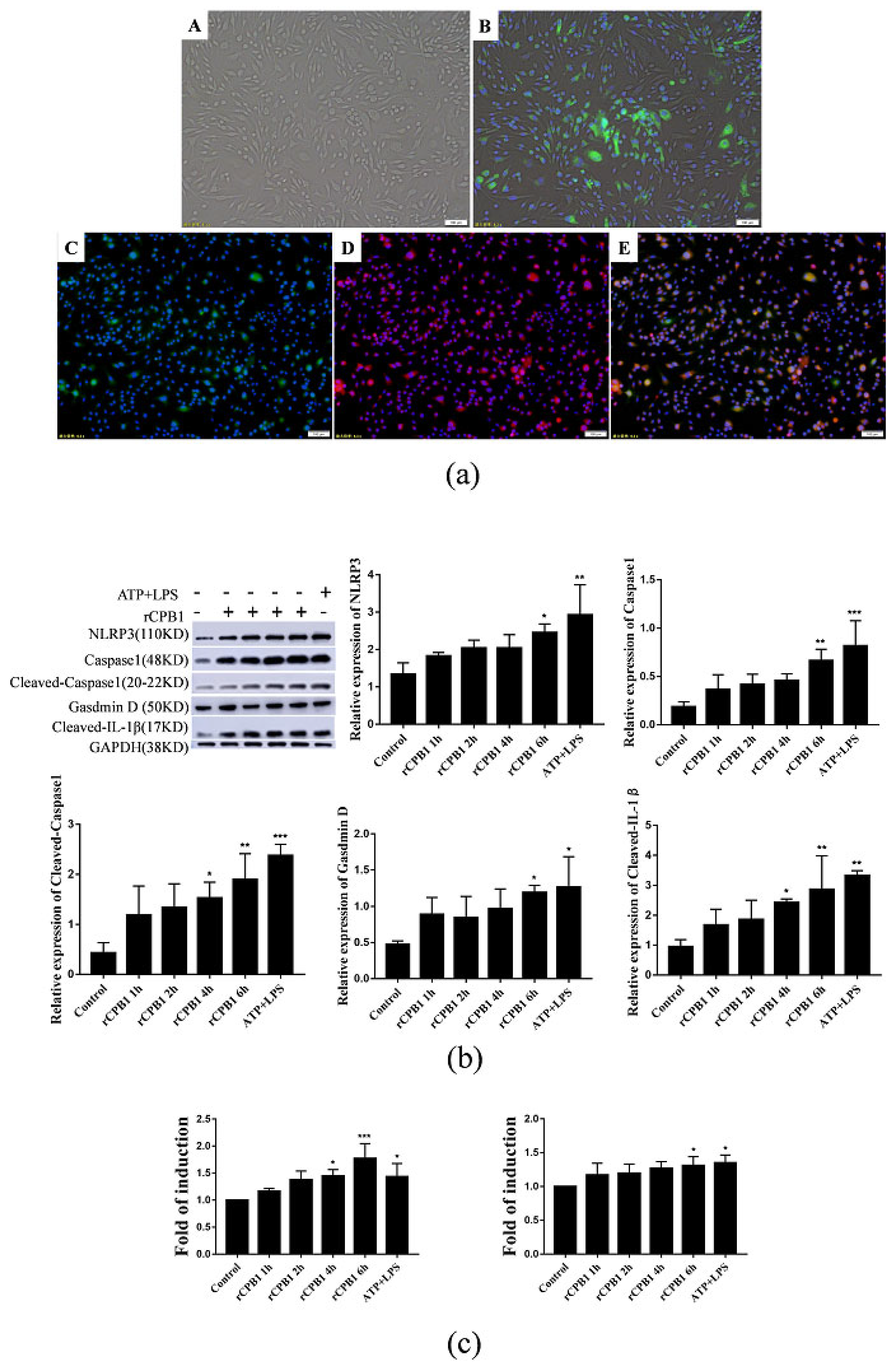
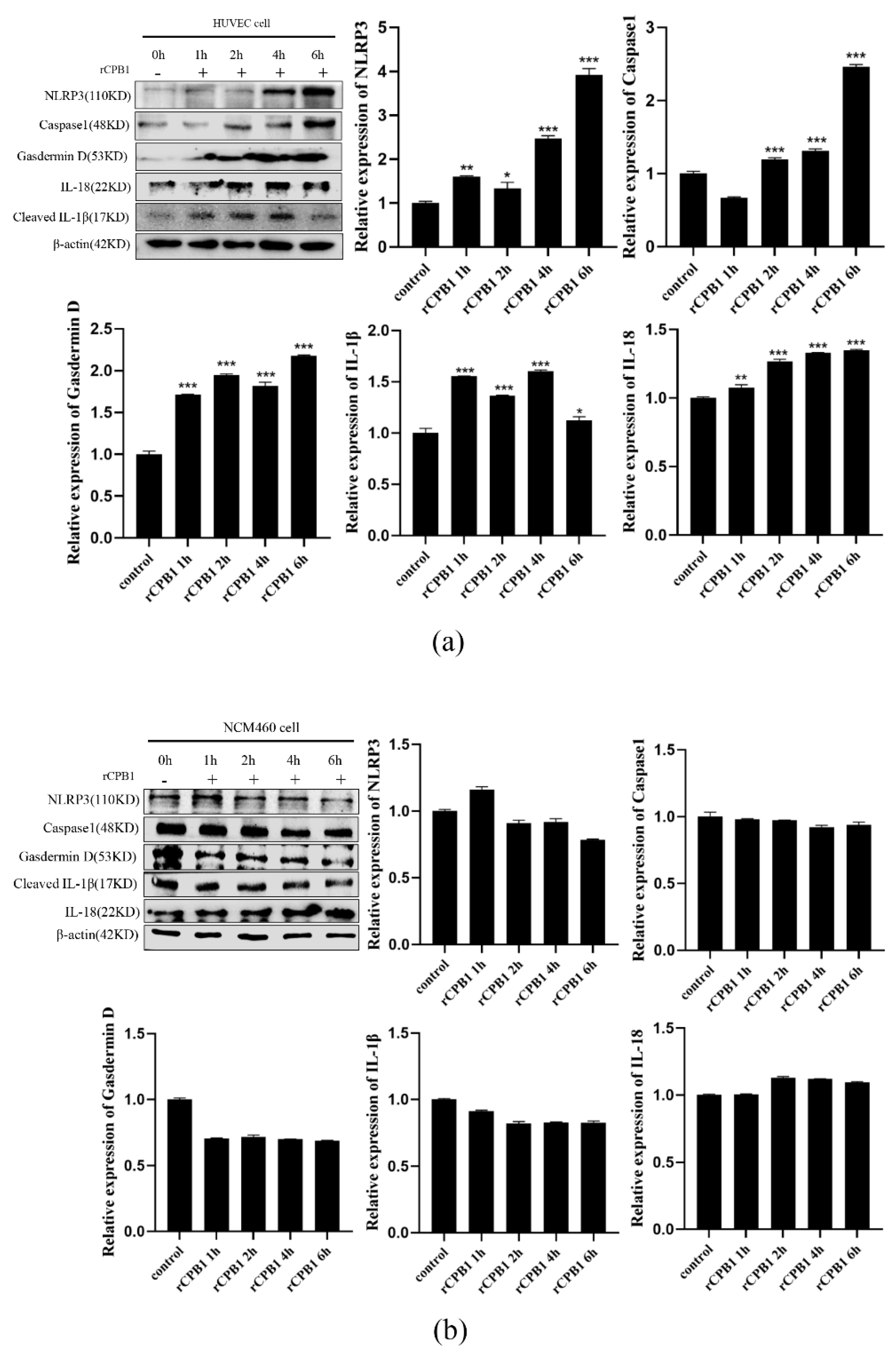
It has been reported in the literature that CPB1-toxin-induced mouse skin TNF-α and IL-1β levels increased in a dose-dependent manner, along with increased plasma extravasation. In the present study, we found that the expression levels of NLRP3, Caspase 1, Gasdermin D, IL-18, and IL-1β were up-regulated after RAW264.7, THP-1 macrophages, and BMDM were treated with rCPB1. The morphological changes of THP-1 macrophages were observed with an electron microscope, and it was found that the cell membrane ruptured and the cell contents overflowed in cells treated with rCPB1. Similarly, the rCPB1 toxin could cause the HUVEC cells to release inflammatory factors. These phenomena indicate that rCPB1 could induce pyroptosis in macrophage cells and endothelial cells. However, the results of qRT-PCR and Western blotting showed no significant changes in the mRNA and protein-related pyroptosis in NCM460 cells treated with rCPB1 toxin. The ELISA results showed the expression of IL-18 and IL-1β levels, and for the NCM460 cells there was no significant change either. This result suggests that the rCPB1 toxin protein can cause the death of colonic mucosal epithelium, but does not induce pyroptosis in NCM460 cells.
It has been reported that the CPB1 toxin can cause necrotizing enteritis in animals, which can lead to hemorrhagic necrosis of the intestinal mucosa. It is generally considered that the intestinal mucosal epithelium is the main target of CPB1-toxin-induced damage. However, studies have also shown that CPB1 does not cause direct damage to the intestinal epithelial cells of pigs after many relevant studies, although intestinal epithelial cells secrete low levels of IL-1β. A large number of immune cells, including macrophages and monocytes, are distributed in the intestinal epithelium and lamina propria, to compose the complex intestinal immune microenvironment. Several lines of study have found that the CPB1 toxin can induce different cytotoxic responses on different cell lines. The reason for this phenomenon may be related to the receptors on the plasma membrane surface of different cell types. For example, Takehara et al. discovered that the P2X purinoreceptor 7 (P2RX7) may be the receptor with the CPB1 toxin [
26]. The P2RX7 receptor is widely expressed in many types of cells, including intestinal epithelial cells, and whether the CPB1 toxin caused the death of intestinal epithelial cells is directly or indirectly caused by the pore-forming toxin. Combined with the results of this experiment, the expression of pyroptosis-related factors did not occur in intestinal epithelial cells after being stimulated by CPB1 toxin. It is speculated that the death mode of intestinal epithelial cells may not be inflammatory death, such as pyroptosis. The release of inflammatory factors in animals may be caused by immune cells in the lamina propria of the intestinal tissue.
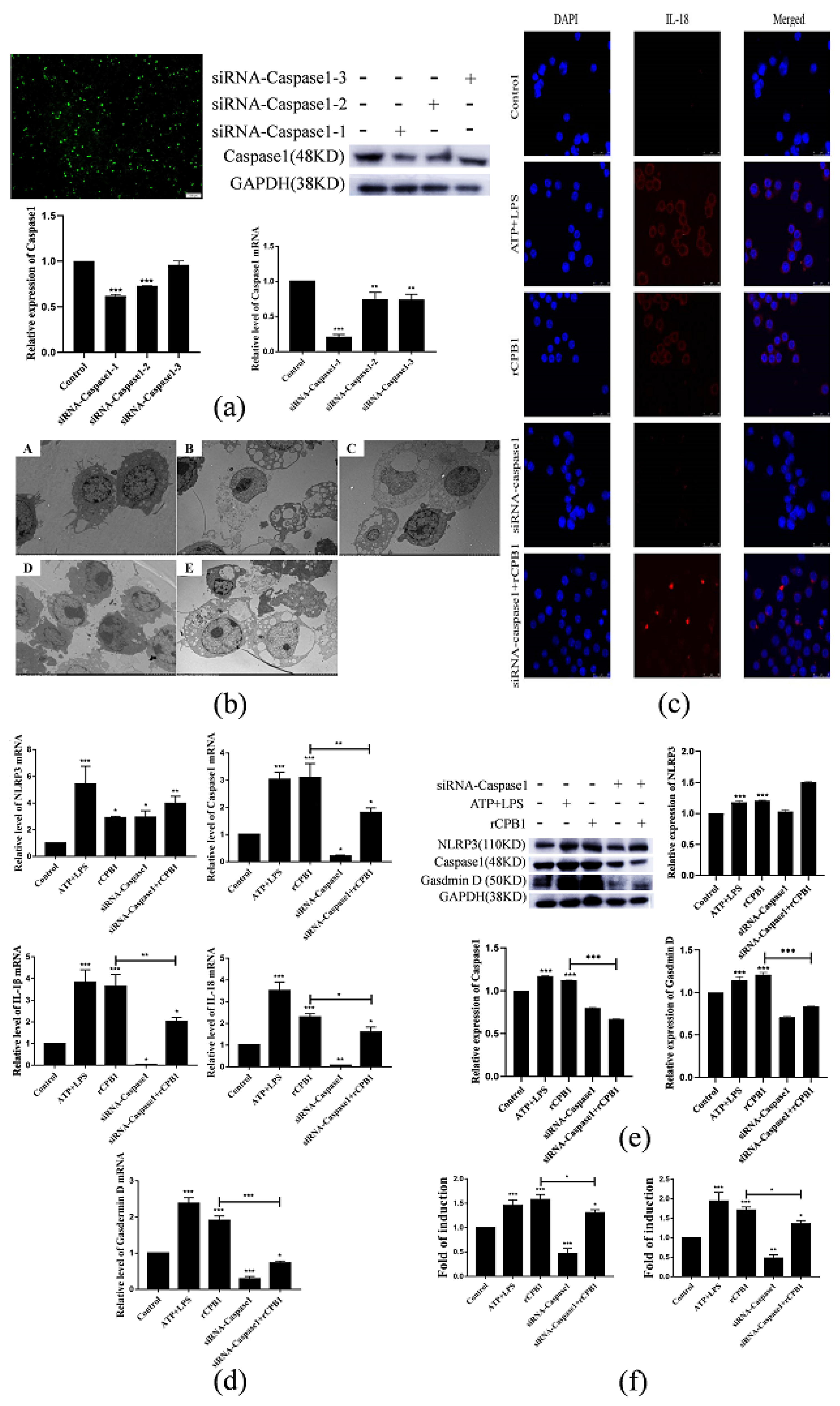
After considering our experimental results, we speculate that rCPB1 can induce the assembly of the NLRP3 inflammasome in macrophages, possibly through a K+ flow mechanism, which then activates Caspase 1. The activation of Caspase 1 in turn leads to the cleavage of GSDMD, and subsequently the formation of channels in the plasma membrane, and the release of the inflammatory cytokines IL-18 and IL-1β. Ultimately, a large number of inflammatory cytokines are produced through the inflammatory cascade, which destroys the intestinal epithelium, leading to diseases such as necrotizing enteritis in the host.
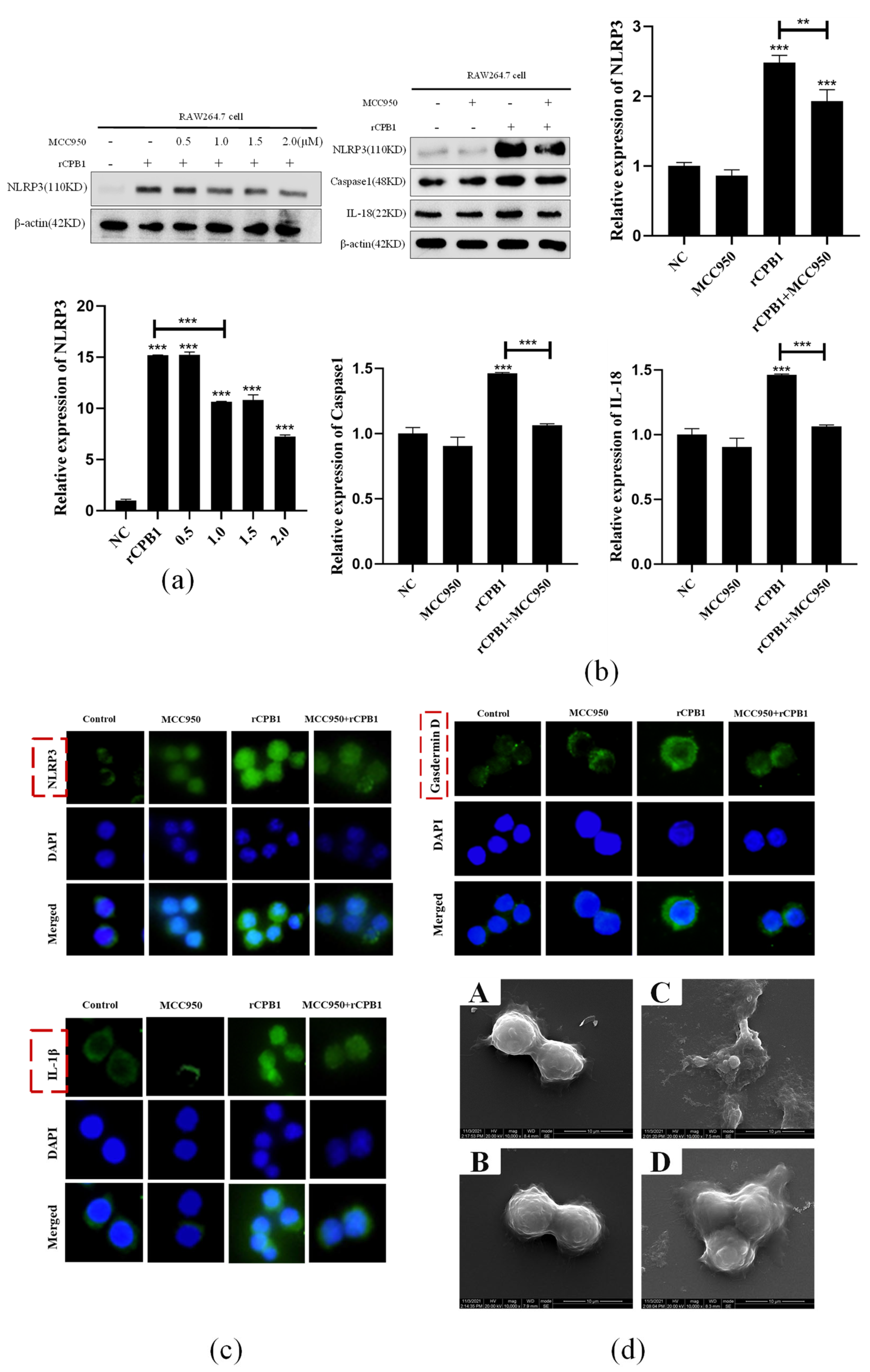
In this experiment, we also found that small-molecule inhibitors MCC950 and small interfering RNA in Caspase 1 could inhibit the occurrence of pyroptosis and alleviate the excessive release of inflammatory factors from macrophages. The NLRP3 inflammasome inhibitor MCC950 has been used in the treatment of some clinical diseases. In the treatment of necrotizing enteritis caused by Clostridium perfringens, whether it can be considered to use drugs that inhibit the excessive release of inflammatory factors by immune cells to avoid the subsequent inflammatory factor storm is still in question.
5. Materials and Methods
5.1. Bacterial Strains, Plasmids, Cell Lines, Reagents, and Mice
E. coli BL21 (DE3) (pET30a-GFP) with green fluorescence were stored in our laboratory. BL21 (DE3) were purchased from Quanshijin Biotechnology Co., Ltd., Beijing, China. Mouse mononuclear macrophage leukemia cells (RAW264.7), human mononuclear leukemia cells (THP-1), normal colon mucosal epithelial cells (NCM460), and mouse fibroblast cell line L929 were purchased from the National Collection of Authenticated Cell Cultures; human umbilical vein endothelial cells (HUVEC) were donated by Professor Gao Liyang. The pTIG-Trx plasmid was donated by Professor Wang Jinglin. C57BL/6J mice were purchased from Weitong Lihua Experimental Animal Technology Co., Ltd., Beijing, China.
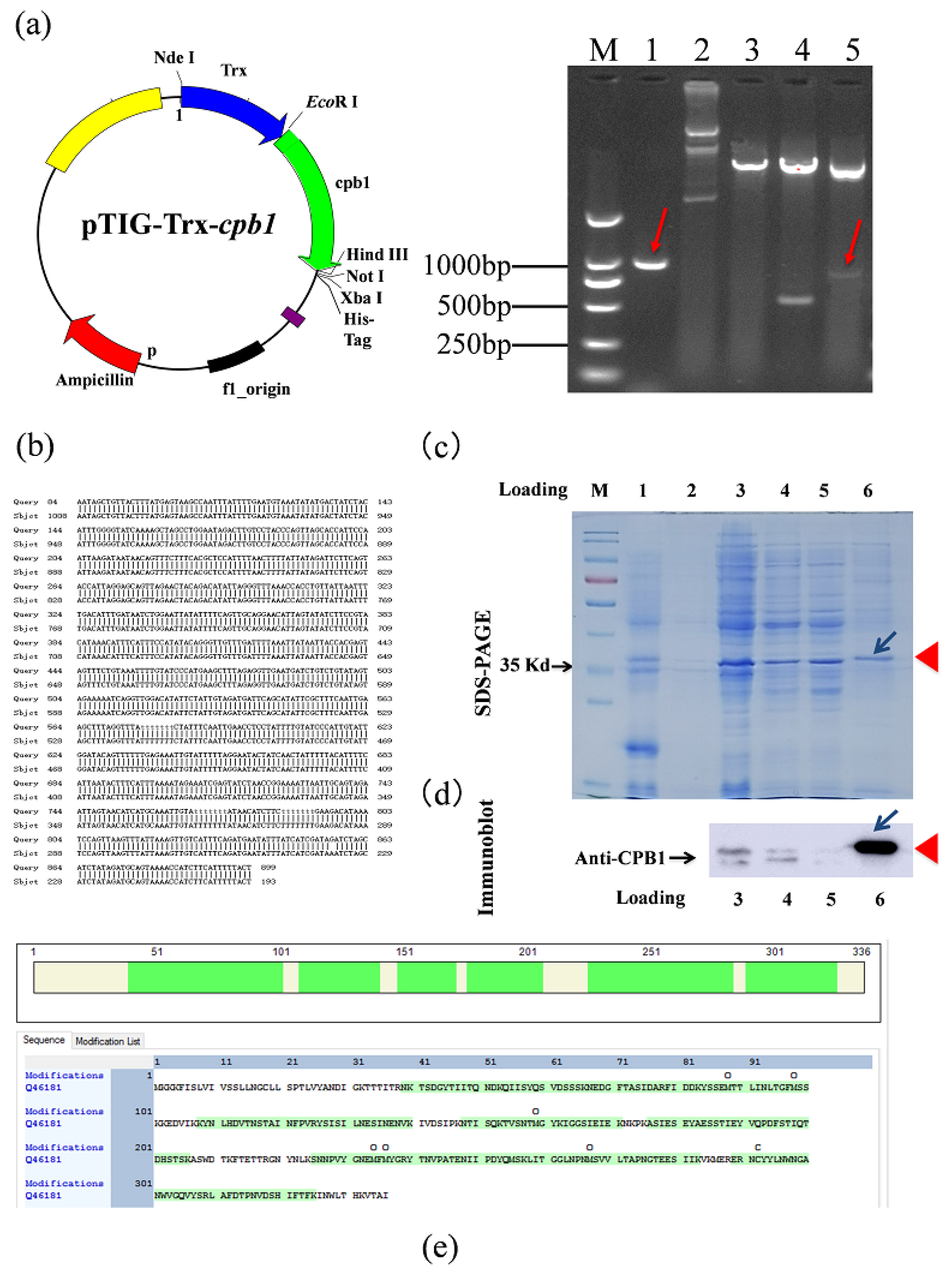
5.2. Constructing the Plasmid That Expresses rCPB1
According to the cpb1 DNA sequence in GenBank (X83275.1), the following primers were designed to amplify the encoding regions of the cpb1 gene: cpb1-F: 5′ CGGAATTCCATATGGATATAGGCAAAACTACTACTAT′ and cpb1-R: 5′ GAATGCGGCCGCAATAGCTGTTACTTTATG′. Enzyme digestion was used to analyze the cpb1 gene loci, and we found two Hind III enzyme loci of approximately 500 bp and 600 bp; therefore, the recombinant plasmid was treated with EcoR I single-enzyme digestion and EcoR I/Hind III and EcoR I/Not I double-enzyme digestion and subjected to agarose gel electrophoresis analysis. The amplified DNA fragment was cloned into the pTIG-Trx vector to construct the pTIG-Trx-cpb1 plasmid for sequencing and transformed into BL21 (DE3) to induce the expression of CPB1.
5.3. Induction of the Expression of rCPB1
BL21(DE3) pTIG-Trx-cpb1 were isolated and cultured in LB medium with 50 μg/mL of ampicillin and 0.5 mM isopropyl β-D-1-thiogalactopyranoside (IPTG) in a 20 °C incubator with shaking overnight. Then, the bacterial protein samples were analyzed for expression of the rCPB1 on 12% SDS-PAGE.
5.4. Purification and Assay of rCPB1
Given that the rCPB1 expressed from the pTIG-Trx vector had low solubility, it could be purified using inclusion body renaturation. In brief, the IPTG-induced BL21 (DE3) pTIG-Trx-cpb1 were harvested with centrifugation (12,000 rpm, 10 min). The pellet was resuspended with PBS and ultrasonically broken. Then, the mixture was separated by centrifugation at 12,000 rpm for 30 min. The pellet was resuspended, respectively, in eluent I (50 mM Tris-Cl [pH 8.0], 10 mM EDTA, 5% glycerin, 0.1% TritonX-100, 100 mM β-mercaptoethanol), eluent II (eluent I with 0.3 M urea), or eluent III (eluent I with 0.6 M urea). To the bacteria obtained from the 1 L culture, 40 mL washing solution was added; centrifugation at 12,000 rpm for 15 min was conducted after each wash. The pellet was resuspended in denaturation solution (50 mM Tris-Cl [pH 8.0], 5% glycerin, 8 M urea) with ultrasound for 10 min. At this point, insoluble matter was significantly reduced. To bacteria obtained from 1 L culture, 20 mL of denaturation solution was added; the mixture was centrifuged at 12,000 rpm for 20 min. The target protein was now dissolved in the supernatant. We used a 3.5-kDa dialysis bag for renaturation of the supernatant in a low-temperature refrigerator. The process was as follows: addition of refolding solution I (50 mM Tris-Cl [pH 8.0], 5% glycerol, 6 M urea), dialysis for 2 h; addition of refolding solution II (50 mM Tris-Cl [pH 8.0], 5% glycerol, 4 M urea), dialysis for 3 h; addition of refolding solution III (50 mM Tris-Cl [pH 8.0], 5% glycerol, 2 M urea), dialysis for 2 h; and addition of refolding solution IV (50 mM Tris-Cl [pH 8.0], 5% glycerol) for 2 h or overnight. After renaturation, the solution was slightly turbid, but no obvious floccules appeared. The solution was centrifuged at 12,000 rpm for 20 min, and the supernatant containing the target protein was decanted. The endotoxin of the rCPB1 proteins was removed using a ToxinEraser™ Kit (GenScript, Piscataway, NJ, USA). The protein fractions were determined with 12% SDS-PAGE and an immunoblotting assay using a CPB1 antibody (Gene Tex GTX41667). Immune complexes were detected using the automatic chemiluminescence imager (GE Amersham Imager 600).
5.5. Cell Culture and Treatment
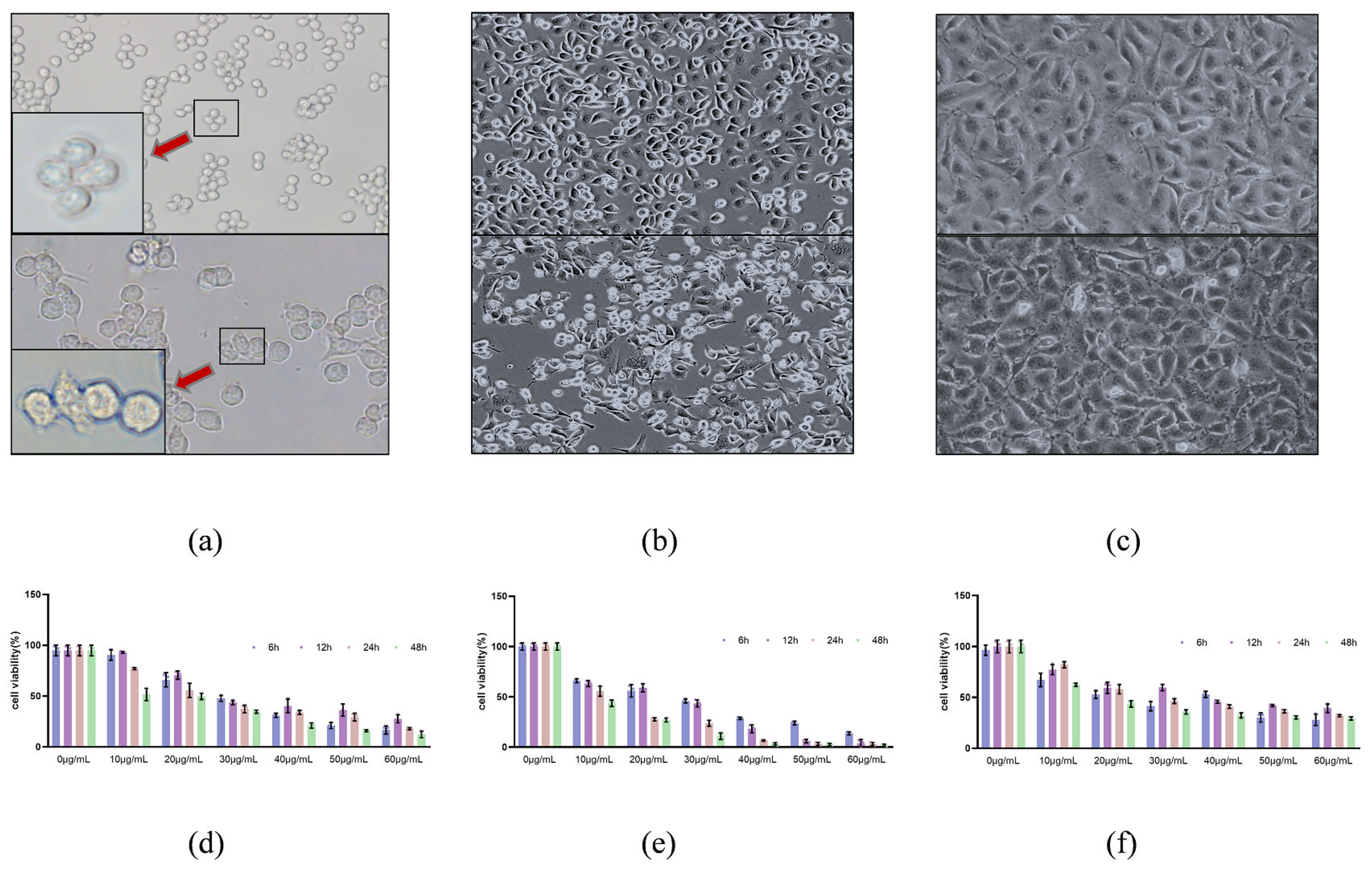
All cells were cultured in an incubator (37 °C, 5% CO2). RAW264.7 and HUVEC cells were cultured in DMEM medium. THP-1 and NCM460 cells were cultured in RPMI 1640 medium. Complete medium was composed of basal medium and 10% fetal bovine serum. Additionally, the cells in good condition were transferred to a six-well plate at a density of 1 × 106 cells/mL and cultured overnight at 37 °C. The dose of rCPB1 that caused 50% cell death was added to the cells for treatment for 1 h, 2 h, 4 h, or 6 h. In the positive control group, 5 mM ATP (Sigma-Aldrich, St. Louis, MO, USA) was added for treatment: 0.5 h upon 1 μg/mL LPS treatment 5.5 h. In the inhibitor group, different doses of MCC950 were added for 1 h before rCPB1 treatment.
PMA (Sigma-Aldrich, St. Louis, MO, USA), LPS (Sigma-Aldrich, St. Louis, MO, USA) and MCC950 (MCE, Monmouth Junction, NJ, USA) powders were dissolved in DMSO (Sigma-Aldrich, St. Louis, MO, USA) to achieve a stock solution, and kept in aliquot at −20 °C.
5.6. Cytotoxicity Assay
RAW264.7, NCM460, and HUVEC cells were used to evaluate the activity of the rCPB1. Cells were seeded into 96-well plates at a density of 8 × 104 cells/mL per well for overnight culture, and six duplicate wells were set up in each group. The medium was changed for fresh medium, and different concentrations of toxin proteins were used for further incubation for different processing times. Then, CCK-8 solution was added to each well. After 2 h, the absorbance value was measured at 450 nm using a luciferase plate analyzer and the data were recorded.
5.7. Preparation and Phagocytosis of BMDM
Mice with 6–9 weeks of cervical dislocation were disinfected and the femurs removed. The bone marrow of the femurs was forced out with PBS until the wash was colorless. The supernatant was discarded after centrifugation (800 rpm, 5 min). The pellet was resuspended in RBC lysis buffer at 20s, followed by the same amount of DMEM complete medium, and centrifuged. The cells were resuspended in complete medium containing 20% L929 culture supernatant, and the solution was transferred to the six-well plate at a density of 1 × 106 cells/mL. After culturing for 7 days, the induced E. coli BL21(DE3) (pET30a-GFP) were added to a final concentration of 5 × 107 cells/mL and cultured at 37 °C for 2 h. The phagocytosis effect was analyzed and the images were captured.
5.8. Transfection
After being treated with rCPB1 with different times, the cells were washed 2–3 times with PBS. Lipofectamine™ iMAX transfection reagent (Thermo, Waltham, MA, USA) performed cell transfection through the concentration of siRNAs, according to the manufacturer’s recommendations.
The sequence for siRNAs is shown below:
siRNA-Caspase1-1: 5′-GGTATCCAGGAGGGAATATGT-3′,
siRNA-Caspase1-2: 5′-GCCCAAGGTGATCATTATTCA-3′,
siRNA-Caspase1-3: 5′-GGCATTAAGAAGGCCCATATA-3′.
5.9. Real-Time Fluorescence Quantitative PCR
After being treated with rCPB1 for different times, the cells were washed 2–3 times with PBS and lysed with Trizol reagent (Ambion, Austin, TX, USA), so that total RNAs were extracted. The RNA concentration was measured using a Nanodrop 8000 nucleic acid protein analyzer, and the measured data were recorded. Then, complementary DNA was prepared by using cDNA Synthesis Kits (Thermo, Waltham, MA, USA). SYBR Green (QIAGEN, DUS, Hilden, Germany)-based quantitative PCR was performed by using the QuantStudio™ Detection System (Thermo, Waltham, MA, USA), according to the manufacturer’s recommendations. It counted according to 2
−ΔΔCT methods. Additionally, primers used in the qPCR assays are shown in
Supplementary Table S1.
5.10. Enzyme-Linked Immunosorbent Assay
After being treated with rCPB1 for different times, culture media were collected and centrifuged to remove any cell debris or suspended cells. IL-1β and IL-18 released in the culture supernatant were measured by using a human or mouse ELISA kit (Lianshuo Biological Technology, Shanghai, China), according to the manufacturer’s recommendations.
5.11. Immunofluorescent Staining
After being treated with rCPB1 for different times, the cells were washed with PBS and fixed with 4% paraformaldehyde (RT, 20 min), then permeabilized with 0.5% Triton X-100 (RT, 20 min). Cells were blocked with 3% bovine serum albumin (RT, 1 h) and then incubated with primary antibodies at 4 °C overnight. After being washed, the cells were incubated with the secondary antibody (RT, 1 h) and mounted for fluorescence with DAPI (Thermo, Waltham, MA, USA).
Among them, antibodies against NLRP3, Caspase 1, Actin, IL-18 and Cleaved IL-1β were purchased from Cell Signaling Technology, Danvers, MA, USA. Additionally, antibodies against Gasdermin D, GAPDH and HRP goat anti-mouse or goat anti-rabbit were purchased from Proteintech, Wuhan, China. The FITC-labeled CD11C antibody and Texas Red-labeled CD16 antibody were purchased from Thermo, Waltham, MA, USA.
5.12. Transmission Electron Microscope Observation
After being treated with rCPB1 for different times, the cells were gently collected in a centrifuge tube, centrifuged (800 rpm, 5 min), and the supernatant discarded. They were then washed twice with PBS and mixed with 2% glutaraldehyde fixative solution for 2 h at 4 °C; the solution was changed after 2 h, and this was repeated 3 times. The cells were then fixed with 1% hungry acid for 2 h, then rinsed with PBS for 5 min three times. They were treated at 4 °C with 30% ethanol, 50% ethanol, and 70% ethanol successively for 10 min, and treated at room temperature with 80% ethanol, 90% ethanol, and 100% ethanol for 10 min. Then, epoxy propane was added for 15 min and the process was repeated. Incomplete embedding solution and epoxy propane at ratios of 1:1 and 2:1 were added for 1 h, respectively, followed by overnight treatment with incomplete embedding solution, and incubation at 35 °C, 42 °C, and 60 °C for 6 h, 12 h, and 48h, respectively. The embedded cells were cut into slices, placed on slides, and observed using transmission electron microscopy.
5.13. Statistical Analysis
All data were obtained after at least three independent experiments. One-way ANOVA in GraphPad Prism 7.0 software was used for statistical analysis, and statistical evaluation of the differences was performed with a t-test for two groups. A difference was considered to be statistically significant at a p value of <0.05. Data are presented as the mean ± standard deviation (SD).

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







