Four PQQ-Dependent Alcohol Dehydrogenases Responsible for the Oxidative Detoxification of Deoxynivalenol in a Novel Bacterium Ketogulonicigenium vulgare D3_3 Originated from the Feces of Tenebrio molitor Larvae

 ,
,
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
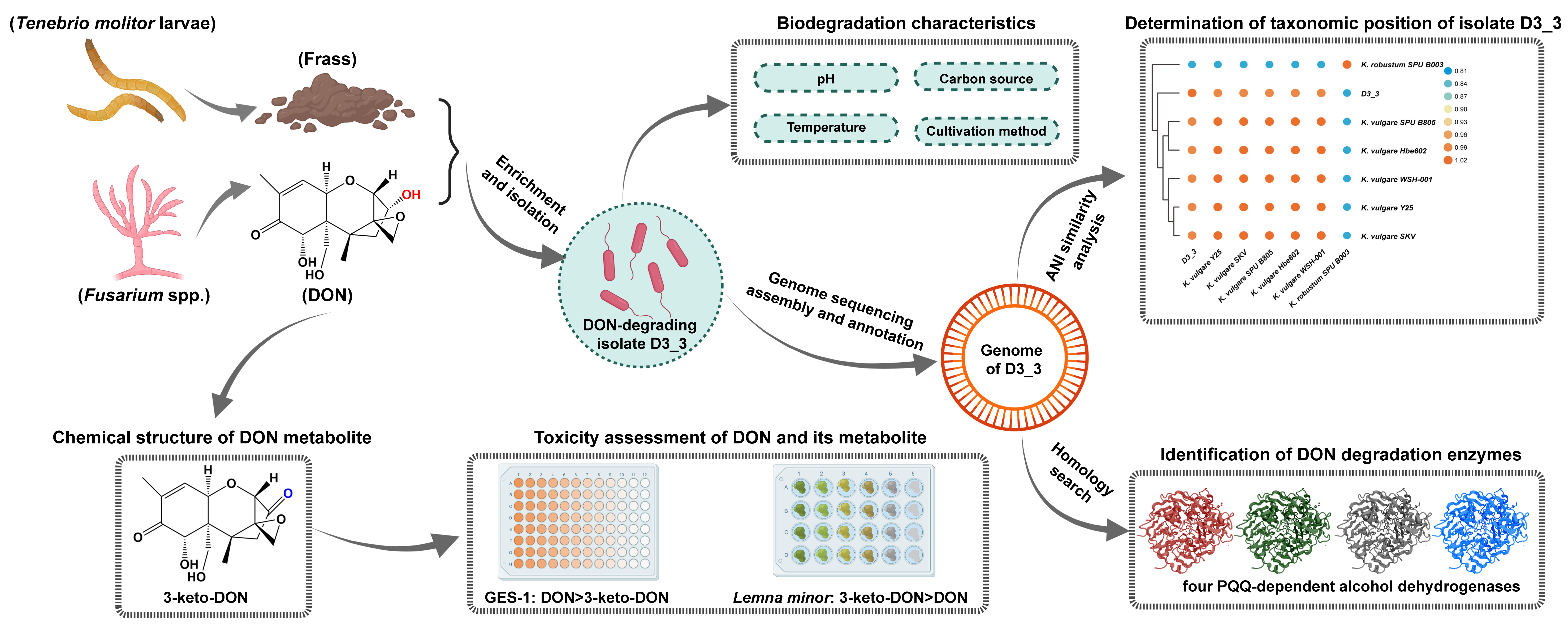
2. Results
2.1. A Potent DON-Oxidizing Strain Ketogulonicigenium Vulgare D3_3 Isolated from Yellow Mealworm Feces
2.2. Effects of Different Growth Factors on the DON-Degrading Activity of Strain D3_3
2.3. Cytotoxicity and Phytotoxicity of 3-Keto-DON
2.3.1. Effect of 3-Keto-DON on the Viability of GES-1 Cells
2.3.2. Effect of 3-Keto-DON on L. Minor
2.4. Four PQQ-Dependent Alcohol Dehydrogenases Responsible for DON Transformation
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Chemicals and Reagents
5.2. Enrichment, Isolation, and Identification of DON-Degrading Microorganisms
5.2.1. Yellow Mealworm Feces Collection and Processing
5.2.2. Enrichment and Isolation Procedures
5.2.3. 16S rRNA-Based Phylogenetic Analysis of DON-Degrading Strain
5.3. Effects of Various Factors on the DON-Degrading Activity of Strain D3_3
5.4. DON and Its Metabolite Analysis
5.4.1. DON Detection Using UPLC Technique
5.4.2. Analysis of DON Degradation Metabolite Using the UPLC-Q-TOF-MS Method
5.5. Toxicity Assay of DON and Its Metabolite
5.5.1. In Vitro Cytotoxicity Assay Using GES-1
5.5.2. Phytotoxicity Assay Using Lemna minor
5.6. Identification of DON-Oxidizing Enzyme in Strain D3_3
5.6.1. Sequencing, Assembly and Annotation of D3_3’ Genome, as well as Scouting Potential Genes Involved in DON Oxidation
5.6.2. Cloning, Expression, and Activity Assay for Potential DON-Oxidizing Enzymes
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kushiro, M. Effects of Milling and Cooking Processes on the Deoxynivalenol Content in Wheat. Int. J. Mol. Sci. 2008, 9, 2127–2145. [Google Scholar] [CrossRef]
- Pestka, J.J. Deoxynivalenol: Toxicity, Mechanisms and Animal Health Risks. Anim. Feed Sci. Technol. 2007, 137, 283–298. [Google Scholar] [CrossRef]
- Pestka, J.J.; Smolinski, A.T. Deoxynivalenol: Toxicology and Potential Effects on Humans. J. Toxicol. Environ. Health Part B 2005, 8, 39–69. [Google Scholar] [CrossRef] [PubMed]
- Del Favero, G.; Janker, L.; Neuditschko, B.; Hohenbichler, J.; Kiss, E.; Woelflingseder, L.; Gerner, C.; Marko, D. Exploring the Dermotoxicity of the Mycotoxin Deoxynivalenol: Combined Morphologic and Proteomic Profiling of Human Epidermal Cells Reveals Alteration of Lipid Biosynthesis Machinery and Membrane Structural Integrity Relevant for Skin Barrier Function. Arch. Toxicol. 2021, 95, 2201–2221. [Google Scholar] [CrossRef] [PubMed]
- Pestka, J.J. Deoxynivalenol: Mechanisms of Action, Human Exposure, and Toxicological Relevance. Arch. Toxicol. 2010, 84, 663–679. [Google Scholar] [CrossRef]
- Rotter, B.A. Toxicology of Deoxynivalenol (vomitoxin). J. Toxicol. Environ. Health 1996, 48, 1–34. [Google Scholar] [CrossRef]
- Sobrova, P.; Adam, V.; Vasatkova, A.; Beklova, M.; Zeman, L.; Kizek, R. Deoxynivalenol and Its Toxicity. Interdiscip. Toxicol. 2010, 3, 94–99. [Google Scholar] [CrossRef]
- Murata, H.; Yamaguchi, D.; Nagai, A.; Shimada, N. Reduction of Deoxynivalenol Contaminating Corn Silage by Short-Term Ultraviolet Irradiation: A Pilot Study. J. Vet. Med. Sci. 2011, 73, 1059–1060. [Google Scholar] [CrossRef]
- Park, B.J.; Takatori, K.; Sugita-Konishi, Y.; Kim, I.-H.; Lee, M.-H.; Han, D.-W.; Chung, K.-H.; Hyun, S.O.; Park, J.-C. Degradation of Mycotoxins Using Microwave-Induced Argon Plasma at Atmospheric Pressure. Surf. Coat. Technol. 2007, 201, 5733–5737. [Google Scholar] [CrossRef]
- Savi, G.D.; Piacentini, K.C.; Scussel, V.M. Ozone Treatment Efficiency in Aspergillus and Penicillium Growth Inhi- bition and Mycotoxin Degradation of Stored Wheat Grains (Triticum aestivum L.): Ozone Treatment Efficiency on Fungi and Mycotoxins. J. Food Process Pres. 2015, 39, 940–948. [Google Scholar] [CrossRef]
- Wang, L.; Luo, Y.; Luo, X.; Wang, R.; Li, Y.; Li, Y.; Shao, H.; Chen, Z. Effect of Deoxynivalenol Detoxification by Ozone Treatment in Wheat Grains. Food Control 2016, 66, 137–144. [Google Scholar] [CrossRef]
- Zhang, C.; Zhuang, K.; Chen, L.; Ding, W.; Chen, X.; Shao, L.; Wang, G.; Wang, Y. Reducing Deoxynivalenol Con- tent in Wheat by a Combination of Gravity Separation and Milling and Characterization of the Flours Produced. J. Cereal Sci. 2022, 104, 103372. [Google Scholar] [CrossRef]
- Matumba, L.; Van Poucke, C.; Njumbe Ediage, E.; Jacobs, B.; De Saeger, S. Effectiveness of Hand Sorting, Flotation/Washing, Dehulling and Combinations Thereof on the Decontamination of Mycotoxin-Contaminated White Maize. Food Addit. Contam. Part A 2015, 32, 960–969. [Google Scholar] [CrossRef]
- Trenholm, H.L.; Charmley, L.L.; Prelusky, D.B.; Warner, R.M. Washing Procedures Using Water or Sodium Carbonate Solutions for the Decontamination of Three Cereals Contaminated with Deoxynivalenol and Zearalenone. J. Agric. Food. Chem. 1992, 40, 2147–2151. [Google Scholar] [CrossRef]
- Yumbe-Guevara, B.E.; Imoto, T.; Yoshizawa, T. Effects of Heating Procedures on Deoxynivalenol, Nivalenol and Zearalenone Levels in Naturally Contaminated Barley and Wheat. Food Addit. Contam. 2003, 20, 1132–1140. [Google Scholar] [CrossRef] [PubMed]
- Sundstøl Eriksen, G.; Pettersson, H.; Lundh, T. Comparative Cytotoxicity of Deoxynivalenol, Nivalenol, Their Acetylated Derivatives and de-Epoxy Metabolites. Food Chem. Toxicol. 2004, 42, 619–624. [Google Scholar] [CrossRef]
- Shima, J.; Takase, S.; Takahashi, Y.; Iwai, Y.; Fujimoto, H.; Yamazaki, M.; Ochi, K. Novel Detoxification of the Trichothecene Mycotoxin Deoxynivalenol by a Soil Bacterium Isolated by Enrichment Culture. Appl. Environ. Microb. 1997, 63, 3825–3830. [Google Scholar] [CrossRef]
- He, J.W.; Bondy, G.S.; Zhou, T.; Caldwell, D.; Boland, G.J.; Scott, P.M. Toxicology of 3-Epi-Deoxynivalenol, a De oxynivalenol-Transformation Product by Devosia mutans 17-2-E-8. Food Chem. Toxicol. 2015, 84, 250–259. [Google Scholar] [CrossRef] [PubMed]
- Mayer, E.; Novak, B.; Springler, A.; Schwartz-Zimmermann, H.E.; Nagl, V.; Reisinger, N.; Hessenberger, S.; Schatzmayr, G. Effects of Deoxynivalenol (DON) and Its Microbial Biotransformation Product Deepoxy-Deoxynivalenol (DOM-1) on a Trout, Pig, Mouse, and Human Cell Line. Mycotoxin Res. 2017, 33, 297–308. [Google Scholar] [CrossRef]
- Ikunaga, Y.; Sato, I.; Grond, S.; Numaziri, N.; Yoshida, S.; Yamaya, H.; Hiradate, S.; Hasegawa, M.; Toshima, H.; Koitabashi, M.; et al. Nocardioides Sp. Strain WSN05-2, Isolated from a Wheat Field, Degrades Deoxynivalenol, Producing the Novel Intermediate 3-Epi-Deoxynivalenol. Appl. Microbiol. Biotechnol. 2011, 89, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Carere, J.; Hassan, Y.I.; Lepp, D.; Zhou, T. The Identification of DepB: An Enzyme Responsible for the Final De- toxification Step in the Deoxynivalenol Epimerization Pathway in Devosia mutans 17-2-E-8. Front. Microbiol. 2018, 9, 1573. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, H.H.; Zhao, C.; Han, Y.T.; Liu, Y.C.; Zhang, X.L. Isolation and Characterization of a Novel De- oxynivalenol-Transforming Strain Paradevosia shaoguanensis DDB001 from Wheat Field Soil. Lett. Appl. Microbiol. 2017, 65, 414–422. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhang, H.; Qin, X.; Wang, X.; Wang, Y.; Bin, Y.; Xie, X.; Zheng, F.; Luo, H. Biodegradation of Deoxyni- valenol by Nocardioides sp. ZHH-013: 3-Keto-Deoxynivalenol and 3-Epi-Deoxynivalenol as Intermediate Products. Front. Microbiol. 2021, 12, 658421. [Google Scholar] [CrossRef] [PubMed]
- Carere, J.; Hassan, Y.I.; Lepp, D.; Zhou, T. The Enzymatic Detoxification of the Mycotoxin Deoxynivalenol: Iden- tification of DepA from the DON Epimerization Pathway. Microb. Biotechnol. 2018, 11, 1106–1111. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Wang, Y.; Ji, F.; Xu, L.; Yu, M.; Shi, J.; Xu, J. Biodegradation of Deoxynivalenol and Its Derivatives by Devosia insulae A16. Food Chem. 2019, 276, 436–442. [Google Scholar] [CrossRef]
- Zhang, J.; Qin, X.; Guo, Y.; Zhang, Q.; Ma, Q.; Ji, C.; Zhao, L. Enzymatic Degradation of Deoxynivalenol by a Novel Bacterium, Pelagibacterium halotolerans ANSP101. Food Chem. Toxicol. 2020, 140, 111276. [Google Scholar] [CrossRef]
- Grau, T.; Vilcinskas, A.; Joop, G. Sustainable Farming of the Mealworm Tenebrio molitor for the Production of Food and Feed. Z. Für Nat. C 2017, 72, 337–349. [Google Scholar] [CrossRef]
- Janković-Tomanić, M.; Petković, B.; Todorović, D.; Vranković, J.; Perić-Mataruga, V. Physiological and Behavioral Effects of the Mycotoxin Deoxynivalenol in Tenebrio molitor Larvae. J. Stored Prod. Res. 2019, 83, 236–242. [Google Scholar] [CrossRef]
- Ochoa Sanabria, C.; Hogan, N.; Madder, K.; Gillott, C.; Blakley, B.; Reaney, M.; Beattie, A.; Buchanan, F. Yellow Mealworm Larvae (Tenebrio molitor) Fed Mycotoxin-Contaminated Wheat—A Possible Safe, Sustainable Protein Source for Animal Feed? Toxins 2019, 11, 282. [Google Scholar] [CrossRef]
- Van Broekhoven, S.; Gutierrez, J.M.; De Rijk, T.C.; De Nijs, W.C.M.; Van Loon, J.J.A. Degradation and Excretion of the Fusarium Toxin Deoxynivalenol by an Edible Insect, the Yellow Mealworm (Tenebrio molitor L.). World Mycotoxin J. 2017, 10, 163–169. [Google Scholar] [CrossRef]
- Jain, C.; Rodriguez-R, L.M.; Phillippy, A.M.; Konstantinidis, K.T.; Aluru, S. High Throughput ANI Analysis of 90K Prokaryotic Genomes Reveals Clear Species Boundaries. Nat. Commun. 2018, 9, 5114. [Google Scholar] [CrossRef]
- He, W.-J.; Zhang, L.; Yi, S.-Y.; Tang, X.-L.; Yuan, Q.-S.; Guo, M.-W.; Wu, A.-B.; Qu, B.; Li, H.-P.; Liao, Y.-C. An Aldo-Keto Reductase Is Responsible for Fusarium Toxin-Degrading Activity in a Soil Sphingomonas Strain. Sci. Rep. 2017, 7, 9549. [Google Scholar] [CrossRef]
- He, W.-J.; Shi, M.-M.; Yang, P.; Huang, T.; Zhao, Y.; Wu, A.-B.; Dong, W.-B.; Li, H.-P.; Zhang, J.-B.; Liao, Y.-C. A Quinone-Dependent Dehydrogenase and Two NADPH-Dependent Aldo/Keto Reductases Detoxify Deoxynivalenol in Wheat via Epimerization in a Devosia Strain. Food Chem. 2020, 321, 126703. [Google Scholar] [CrossRef]
- Fuchs, E.; Binder, E.M.; Heidler, D.; Krska, R. Structural Characterization of Metabolites after the Microbial Deg- radation of Type A Trichothecenes by the Bacterial Strain BBSH 797. Food Addit. Contam. 2002, 19, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Mu, P.; Wen, J.; Sun, Y.; Chen, Q.; Deng, Y. Detoxification of Trichothecene Mycotoxins by a Novel Bacterium, Eggerthella sp. DII-9. Food Chem. Toxicol. 2018, 112, 310–319. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Mu, P.; Zhu, X.; Chen, X.; Tang, S.; Wu, Y.; Miao, X.; Wang, X.; Wen, J.; Deng, Y. Dual Function of a Novel Bacterium, Slackia sp. D-G6: Detoxifying Deoxynivalenol and Producing the Natural Estrogen Analogue, Equol. Toxins 2020, 12, 85. [Google Scholar] [CrossRef] [PubMed]
- He, J.W.; Hassan, Y.I.; Perilla, N.; Li, X.-Z.; Boland, G.J.; Zhou, T. Bacterial Epimerization as a Route for Deoxyni- valenol Detoxification: The Influence of Growth and Environmental Conditions. Front. Microbiol. 2016, 7, 572. [Google Scholar] [CrossRef] [PubMed]
- Ito, M.; Sato, I.; Koitabashi, M.; Yoshida, S.; Imai, M.; Tsushima, S. A Novel Actinomycete Derived from Wheat Heads Degrades Deoxynivalenol in the Grain of Wheat and Barley Affected by Fusarium Head Blight. Appl. Microbiol. Biotechnol. 2012, 96, 1059–1070. [Google Scholar] [CrossRef]
- Ito, M.; Sato, I.; Ishizaka, M.; Yoshida, S.; Koitabashi, M.; Yoshida, S.; Tsushima, S. Bacterial Cytochrome P450 System Catabolizing the Fusarium Toxin Deoxynivalenol. Appl. Environ. Microb. 2013, 79, 1619–1628. [Google Scholar] [CrossRef]
- Jia, R.; Cao, L.; Liu, W.; Shen, Z. Detoxification of Deoxynivalenol by Bacillus subtilis ASAG 216 and Characteri- zation the Degradation Process. Eur. Food Res. Technol. 2021, 247, 67–76. [Google Scholar] [CrossRef]
- Li, B.; Duan, J.; Ren, J.; Francis, F.; Li, G. Isolation and Characterization of Two New Deoxynivalenol-Degrading Strains, Bacillus sp. HN117 and Bacillus sp. N22. Toxins 2022, 14, 781. [Google Scholar] [CrossRef] [PubMed]
- Qu, R.; Jiang, C.; Wu, W.; Pang, B.; Lei, S.; Lian, Z.; Shao, D.; Jin, M.; Shi, J. Conversion of DON to 3- Epi -DON in Vitro and Toxicity Reduction of DON in Vivo by Lactobacillus rhamnosus. Food Funct. 2019, 10, 2785–2796. [Google Scholar] [CrossRef] [PubMed]
- Sato, I.; Ito, M.; Ishizaka, M.; Ikunaga, Y.; Sato, Y.; Yoshida, S.; Koitabashi, M.; Tsushima, S. Thirteen Novel Deox- ynivalenol-Degrading Bacteria Are Classified within Two Genera with Distinct Degradation Mechanisms. FEMS Microbiol. Lett. 2012, 327, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Hou, Q.; Guo, Q.; Zhang, J.; Sun, Y.; Wei, H.; Shen, L. Isolation and Characterization of a Deoxynivalenol-Degrading Bacterium Bacillus licheniformis YB9 with the Capability of Modulating Intestinal Microbial Flora of Mice. Toxins 2020, 12, 184. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, D.L.; Guo, B.Y. Biodegradation of Deoxynivalenol by Two-Member Bacterial Consortia Isolated from Tenebrio molitor Faeces. Int. Biodeter. Biodegr. 2023. in preparation. [Google Scholar]
- Urbance, J.W.; Bratina, B.J.; Stoddard, S.F.; Schmidt, T.M. Taxonomic Characterization of Ketogulonigenium vulgare Gen. Nov., Sp. Nov. and Ketogulonigenium robustum sp. Nov., Which Oxidize L-Sorbose to 2-Keto-L-Gulonic Acid. Int. J. Syst. Evol. Micr. 2001, 51, 1059–1070. [Google Scholar] [CrossRef]
- Sugisawa, T.; Miyazaki, T.; Hoshino, T. Microbial Production of L-Ascorbic Acid from D-Sorbitol, L-Sorbose, L-Gulose, and L-Sorbosone by Ketogulonicigenium vulgare DSM 4025. Biosci. Biotechnol. Biochem. 2005, 69, 659–662. [Google Scholar] [CrossRef][Green Version]
- Zhang, J.; Zhou, J.; Liu, J.; Chen, K.; Liu, L.; Chen, J. Development of Chemically Defined Media Supporting High Cell Density Growth of Ketogulonicigenium vulgare and Bacillus megaterium. Bioresour. Technol. 2011, 102, 4807–4814. [Google Scholar] [CrossRef]
- He, J.W. Detoxification of Deoxynivalenol by a Soil Bacterium Devosia mutans 17-2-E-8. Ph.D. Thesis, University of Guelph, Guelph, ON, Canada, 2015. [Google Scholar]
- Yang, Y.; Yu, S.; Tan, Y.; Liu, N.; Wu, A. Individual and Combined Cytotoxic Effects of Co-Occurring Deoxyniva- lenol Family Mycotoxins on Human Gastric Epithelial Cells. Toxins 2017, 9, 96. [Google Scholar] [CrossRef]
- Takeda, K.; Matsumura, H.; Ishida, T.; Samejima, M.; Ohno, H.; Yoshida, M.; Igarashi, K.; Nakamura, N. Charac- terization of a Novel PQQ-Dependent Quinohemoprotein Pyranose Dehydrogenase from Coprinopsis cinerea Classified into Auxiliary Activities Family 12 in Carbohydrate-Active Enzymes. PLoS ONE 2015, 10, e0115722. [Google Scholar] [CrossRef]
- Good, N.M.; Vu, H.N.; Suriano, C.J.; Subuyuj, G.A.; Skovran, E.; Martinez-Gomez, N.C. Pyrroloquinoline Quinone Ethanol Dehydrogenase in Methylobacterium extorquens AM1 Extends Lanthanide-Dependent Metabolism to Multicarbon Substrates. J. Bacteriol. 2016, 198, 3109–3118. [Google Scholar] [CrossRef]
- Chattopadhyay, A.; Förster-Fromme, K.; Jendrossek, D. PQQ-Dependent Alcohol Dehydrogenase (QEDH) of Pseudomonas Aeruginosa Is Involved in Catabolism of Acyclic Terpenes. J. Basic Microb. 2010, 50, 119–124. [Google Scholar] [CrossRef]
- Sarmiento-Pavía, P.D.; Sosa-Torres, M.E. Bioinorganic Insights of the PQQ-Dependent Alcohol Dehydrogenases. J. Biol. Inorg. Chem. 2021, 26, 177–203. [Google Scholar] [CrossRef] [PubMed]
- Toyama, H.; Mathews, F.S.; Adachi, O.; Matsushita, K. Quinohemoprotein Alcohol Dehydrogenases: Structure, Function, and Physiology. Arch. Biochem. Biophys. 2004, 428, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Yan, R.; Li, Y.; Lu, Z.; Bie, X.; Zhao, H.; Lu, F.; Chen, M. Structure–Function Analysis of a Quinone-Dependent Dehydrogenase Capable of Deoxynivalenol Detoxification. J. Agric. Food. Chem. 2022, 70, 6764–6774. [Google Scholar] [CrossRef] [PubMed]
- Keltjens, J.T.; Pol, A.; Reimann, J.; Op den Camp, H.J.M. PQQ-Dependent Methanol Dehydrogenases: Rare-Earth Elements Make a Difference. Appl. Microbiol. Biotechnol. 2014, 98, 6163–6183. [Google Scholar] [CrossRef]
- Toyama, H.; Fujii, A.; Matsushita, K.; Shinagawa, E.; Ameyama, M.; Adachi, O. Three Distinct Quinoprotein Al- cohol Dehydrogenases Are Expressed When Pseudomonas putida Is Grown on Different Alcohols. J. Bacteriol. 1995, 177, 2442–2450. [Google Scholar] [CrossRef][Green Version]
- Studier, F.W. Protein Production by Auto-Induction in High-Density Shaking Cultures. Protein Expr. Purif. 2005, 41, 207–234. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Di Veroli, G.Y.; Fornari, C.; Goldlust, I.; Mills, G.; Koh, S.B.; Bramhall, J.L.; Richards, F.M.; Jodrell, D.I. An Auto mated Fitting Procedure and Software for Dose-Response Curves with Multiphasic Features. Sci. Rep. 2015, 5, 14701. [Google Scholar] [CrossRef]
- Megateli, S.; Dosnon-Olette, R.; Trotel-Aziz, P.; Geffard, A.; Semsari, S.; Couderchet, M. Simultaneous Effects of Two Fungicides (Copper and Dimethomorph) on Their Phytoremediation Using Lemna Minor. Ecotoxicology 2013, 22, 683–692. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, S. Complete Genome Sequence of Paradevosia shaoguanensis Type Strain J5-3, Obtained Using Nanopore and Illumina Sequencing Technologies. Microbiol. Resour. Ann. 2021, 10, e00099-21. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Devel- oped for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Gough, J.; Karplus, K.; Hughey, R.; Chothia, C. Assignment of Homology to Genome Sequences Using a Library of Hidden Markov Models That Represent All Proteins of Known Structure. J. Mol. Biol. 2001, 313, 903–919. [Google Scholar] [CrossRef]
- Pandurangan, A.P.; Stahlhacke, J.; Oates, M.E.; Smithers, B.; Gough, J. The SUPERFAMILY 2.0 Database: A Sig- nificant Proteome Update and a New Webserver. Nucleic Acids Res. 2019, 47, D490–D494. [Google Scholar] [CrossRef]
- Almagro Armenteros, J.J.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; Von Heijne, G.; Nielsen, H. SignalP 5.0 Improves Signal Peptide Predictions Using Deep Neural Networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef]
- Madeira, F.; Pearce, M.; Tivey, A.R.N.; Basutkar, P.; Lee, J.; Edbali, O.; Madhusoodanan, N.; Kolesnikov, A.; Lopez, R. Search and Sequence Analysis Tools Services from EMBL-EBI in 2022. Nucleic Acids Res. 2022, 50, W276–W279. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Li, K.; Li, J.; Wang, T.; Gu, L.; Xun, L. T5 Exonuclease-Dependent Assembly Offers a Low-Cost Method for Efficient Cloning and Site-Directed Mutagenesis. Nucleic Acids Res. 2019, 47, e15. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Zhao, D.; Zhang, W.; Wang, S.; Wu, Y.; Wang, S.; Yang, Y.; Guo, B. Four PQQ-Dependent Alcohol Dehydrogenases Responsible for the Oxidative Detoxification of Deoxynivalenol in a Novel Bacterium Ketogulonicigenium vulgare D3_3 Originated from the Feces of Tenebrio molitor Larvae. Toxins 2023, 15, 367. https://doi.org/10.3390/toxins15060367
Wang Y, Zhao D, Zhang W, Wang S, Wu Y, Wang S, Yang Y, Guo B. Four PQQ-Dependent Alcohol Dehydrogenases Responsible for the Oxidative Detoxification of Deoxynivalenol in a Novel Bacterium Ketogulonicigenium vulgare D3_3 Originated from the Feces of Tenebrio molitor Larvae. Toxins. 2023; 15(6):367. https://doi.org/10.3390/toxins15060367
Chicago/Turabian StyleWang, Yang, Donglei Zhao, Wei Zhang, Songshan Wang, Yu Wu, Songxue Wang, Yongtan Yang, and Baoyuan Guo. 2023. "Four PQQ-Dependent Alcohol Dehydrogenases Responsible for the Oxidative Detoxification of Deoxynivalenol in a Novel Bacterium Ketogulonicigenium vulgare D3_3 Originated from the Feces of Tenebrio molitor Larvae" Toxins 15, no. 6: 367. https://doi.org/10.3390/toxins15060367
APA StyleWang, Y., Zhao, D., Zhang, W., Wang, S., Wu, Y., Wang, S., Yang, Y., & Guo, B. (2023). Four PQQ-Dependent Alcohol Dehydrogenases Responsible for the Oxidative Detoxification of Deoxynivalenol in a Novel Bacterium Ketogulonicigenium vulgare D3_3 Originated from the Feces of Tenebrio molitor Larvae. Toxins, 15(6), 367. https://doi.org/10.3390/toxins15060367