Exploring Toxin Genes of Myanmar Russell’s Viper, Daboia siamensis, through De Novo Venom Gland Transcriptomics



Abstract

1. Introduction
2. Results
2.1. De Novo Transcriptome Assembly
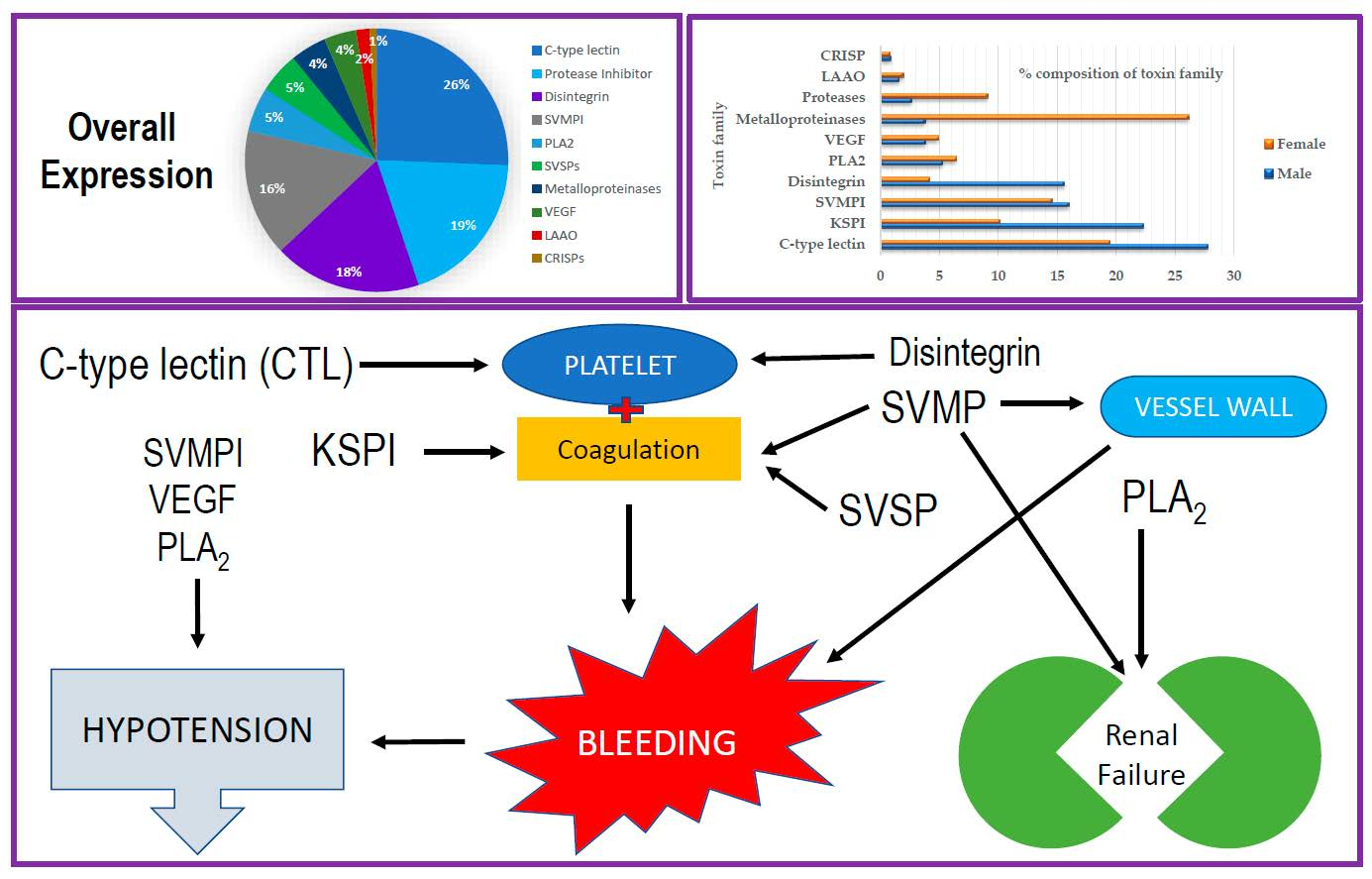
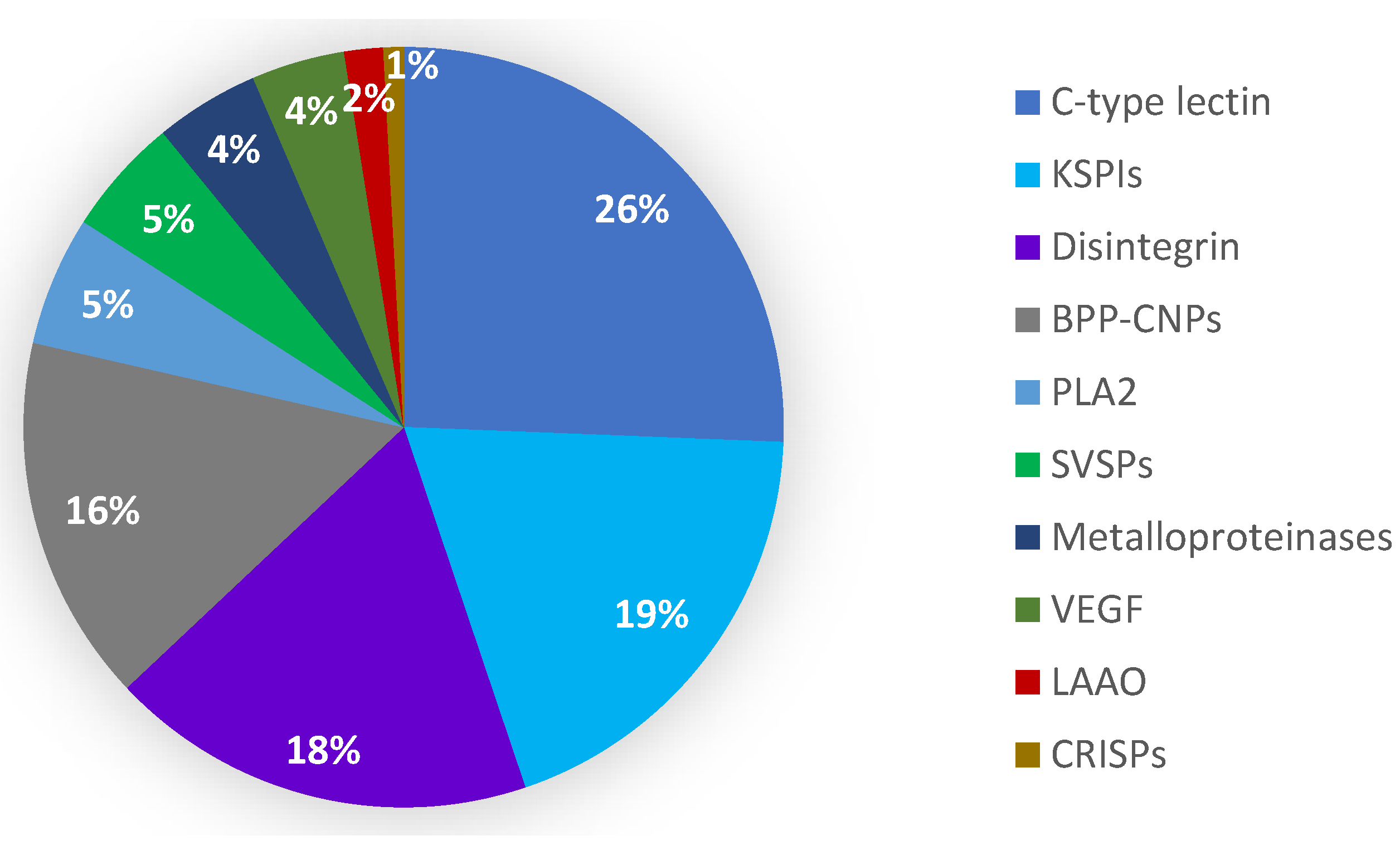
2.2. Categorization of Transcripts and Gene Expression
2.3. Snake Venom C-Type Lectins (CTLs) and Related Proteins
2.4. Kunitz-Type Serine Protease Inhibitors
2.5. RTS-Disintegrins
2.6. Bradykinin-Potentiating Peptides and C-Type Natriuteric Peptide (BPP-CNP) Precursors
2.7. Phospholipase A2 (PLA2)
2.8. Snake Venom Serine Proteases (SVSPs)
2.9. Snake Venom Metalloproteinases (SVMPs)
2.10. Snake Venom Vascular Endothelial Growth Factors (svVEGFs)
2.11. L-Amino Acid Oxidases (LAAOs)
2.12. Cysteine-Rich Venom Proteins (CRISPs)
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Sample Preparation
5.2. RNA Extraction, cDNA Library Construction and Sequencing
5.3. De Novo Transcriptome Assembly and Expression Quantification
5.4. Venom Gland Transcript Classification
5.5. Multiple Sequences Alignment and Gene Tree Reconstruction
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thorpe, R.S.; Pook, C.E.; Malhotra, A. Symptoms of envenoming. Herpetol. J. 2007, 17, 209–218. [Google Scholar]
- Wüster, W. The genus Daboia (Serpentes: Viperidae): Russell’s viper. Hamadryad 1998, 23, 33–40. [Google Scholar]
- Warrell, D.A. Snake venoms in science and clinical medicine 1. Russell’s viper: Biology, venom and treatment of bites. Trans. R. Soc. Trop. Med. Hyg. 1989, 83, 732–740. [Google Scholar] [CrossRef]
- Risch, M.; Georgieva, D.; von Bergen, M.; Jehmlich, N.; Genov, N.; Arni, R.K.; Betzel, C. Snake venomics of the Siamese Russell’s viper (Daboia russelli siamensis)–relation to pharmacological activities. J. Proteom. 2009, 72, 256–269. [Google Scholar] [CrossRef] [PubMed]
- Thomas, R.; Alfred, S.; Zaw, A.; Gentili, S.; Chataway, T.; Peh, C.A.; Hurtado, P.; White, J.; Wiese, M. A demonstration of variation in venom composition of Daboia russelii (Russell’s viper), a significantly important snake of Myanmar, by tandem mass spectrometry. Toxicon 2019, 158, S44–S45. [Google Scholar] [CrossRef]
- Lingam, T.M.C.; Tan, K.Y.; Tan, C.H. Proteomics and antivenom immunoprofiling of Russell’s viper (Daboia siamensis) venoms from Thailand and Indonesia. J. Venom. Anim. Toxins Incl. Trop. Dis. 2020, 26, 1–17. [Google Scholar] [CrossRef]
- Tan, K.Y.; Tan, N.H.; Tan, C.H. Venom proteomics and antivenom neutralization for the Chinese eastern Russell’s viper, Daboia siamensis from Guangxi and Taiwan. Sci. Rep. 2018, 8, 8545. [Google Scholar] [CrossRef]
- Pla, D.; Sanz, L.; Quesada-Bernat, S.; Villalta, M.; Baal, J.; Chowdhury, M.A.W.; León, G.; Gutiérrez, J.M.; Kuch, U.; Calvete, J.J. Phylovenomics of Daboia russelii across the Indian subcontinent. Bioactivities and comparative in vivo neutralization and in vitro third-generation antivenomics of antivenoms against venoms from India, Bangladesh and Sri Lanka. J. Proteom. 2019, 207, 103443. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, A.K.; Kalita, B.; Mackessy, S.P. A proteomic analysis of Pakistan Daboia russelii russelii venom and assessment of potency of Indian polyvalent and monovalent antivenom. J. Proteom. 2016, 144, 73–86. [Google Scholar] [CrossRef]
- Casewell, N.R.; Huttley, G.A.; Wüster, W. Dynamic evolution of venom proteins in squamate reptiles. Nat. Commun. 2012, 3, 1066. [Google Scholar] [CrossRef] [PubMed]
- Calvete, J.J.; Sanz, L.; Angulo, Y.; Lomonte, B.; Gutiérrez, J.M. Venoms, venomics, antivenomics. FEBS Lett. 2009, 583, 1736–1743. [Google Scholar] [CrossRef]
- Zelanis, A.; Andrade-Silva, D.; Rocha, M.M.; Furtado, M.F.; Serrano, S.M.T.; Junqueira-de-Azevedo, I.L.M.; Ho, P.L. A transcriptomic view of the proteome variability of newborn and adult Bothrops jararaca snake venoms. PLoS Negl. Trop. Dis. 2012, 6, e1554. [Google Scholar] [CrossRef] [PubMed]
- Barlow, A.; Pook, C.E.; Harrison, R.A.; Wüster, W. Coevolution of diet and prey-specific venom activity supports the role of selection in snake venom evolution. Proc. R. Soc. B Biol. Sci. 2009, 276, 2443–2449. [Google Scholar] [CrossRef] [PubMed]
- Menezes, M.C.; Furtado, M.F.; Travaglia-Cardoso, S.R.; Camargo, A.C.M.; Serrano, S.M.T. Sex-based individual variation of snake venom proteome among eighteen Bothrops jararaca siblings. Toxicon 2006, 47, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Mirtschin, P.J.; Shine, R.; Nias, T.J.; Dunstan, N.L.; Hough, B.J.; Mirtschin, M. Influences on venom yield in Australian tigersnakes (Notechis scutatus) and brownsnakes (Pseudonaja textilis: Elapidae, Serpentes). Toxicon 2002, 40, 1581–1592. [Google Scholar] [CrossRef]
- Brahma, R.K.; McCleary, R.J.R.; Kini, R.M.; Doley, R. Venom gland transcriptomics for identifying, cataloging, and characterizing venom proteins in snakes. Toxicon 2015, 93, 1–10. [Google Scholar] [CrossRef]
- Schwartz, T.S.; Tae, H.; Yang, Y.; Mockaitis, K.; Van Hemert, J.L.; Proulx, S.R.; Choi, J.H.; Bronikowski, A.M. A garter snake transcriptome: Pyrosequencing, de novo assembly, and sex-specific differences. BMC Genom. 2010, 11, 694. [Google Scholar] [CrossRef]
- Cardoso, K.C.; Da Silva, M.J.; Costa, G.G.L.; Torres, T.T.; Del Bem, L.E.V.; Vidal, R.O.; Menossi, M.; Hyslop, S. A transcriptomic analysis of gene expression in the venom gland of the snake Bothrops alternatus (urutu). BMC Genom. 2010, 11, 605. [Google Scholar] [CrossRef]
- Tan, C.H.; Tan, K.Y.; Fung, S.Y.; Tan, N.H. Venom-gland transcriptome and venom proteome of the Malaysian king cobra (Ophiophagus hannah). BMC Genom. 2015, 16, 687. [Google Scholar] [CrossRef]
- Ducancel, F.; Durban, J.; Verdenaud, M. Transcriptomics and venomics: Implications for medicinal chemistry. Future Med. Chem. 2014, 6, 1629–1643. [Google Scholar] [CrossRef]
- Barua, A.; Mikheyev, A.S. Toxin expression in snake venom evolves rapidly with constant shifts in evolutionary rates. Proc. R. Soc. B Biol. Sci. 2020, 287, 20200613. [Google Scholar] [CrossRef]
- Faisal, T.; Tan, K.Y.; Sim, S.M.; Quraishi, N.; Tan, N.H.; Tan, C.H. Proteomics, functional characterizatioin and antivenom neutralization of the venom of Pakistani Russell’s viper (Daboia russelii) from the wild. J. Proteom. 2018, 183, 1–3. [Google Scholar] [CrossRef]
- Saethang, T.; Somparn, P.; Payungporn, S.; Sriswasdi, S.; Yee, K.T.; Hodge, K.; Knepper, M.A.; Chanhome, L.; Khow, O.; Chaiyabutr, N.; et al. Identification of Daboia siamensis venom using intergrated multi-omics data. Sci. Rep. 2022, 12, 13140. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef]
- Yee, K.T.; Tongsima, S.; Vasieva, O.; Ngamphiw, C.; Wilantho, A.; Wilkinson, M.C.; Somparn, P.; Pisitkun, T.; Rojnuckarin, P. Analysis of snake venom metalloproteinases from Myanmar Russell’s viper transcriptome. Toxicon 2018, 146, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Yee, K.T.; Rojnuckarin, P. Complementary DNA library of Myanmar Russell’s viper (Daboia russelii siamensis) venom gland. Comp. Biochem. Physiol. Part-C Toxicol. Pharmacol. 2020, 227, 108634. [Google Scholar] [CrossRef]
- Holding, M.L.; Margres, M.J.; Mason, A.J.; Parkinson, C.L.; Rokyta, D.R. Evaluating the performance of de novo assembly methods for venom-gland transcriptomics. Toxins 2018, 10, 249. [Google Scholar] [CrossRef] [PubMed]
- Yee, K.T.; Pitts, M.; Tongyoo, P.; Rojnuckarin, P.; Wilkinson, M.C. Snake venom metalloproteinases and their peptide inhibitors from Myanmar Russell’s viper venom. Toxins 2017, 9, 15. [Google Scholar] [CrossRef]
- Yang, Y.; Zhu, Y.; Luo, Y.; Liu, Q.; Hua, X.; Li, J.; Gao, F.; Hofer, J.; Gao, X.; Xiao, L.; et al. Transcriptome analysis of Mesobuthus martensii revealed the differences of their toxins between females and males. Eur. Zool. J. 2022, 89, 1294–1311. [Google Scholar] [CrossRef]
- Lourenço, A.; Zorzella Creste, C.F.; Curtolo de Barros, L.; Delazari dos Santos, L.; Pimenta, D.C.; Barraviera, B.; Ferreira, R.S. Individual venom profiling of Crotalus durissus terrificus specimens from a geographically limited region: Crotamine assessment and captivity evaluation on the biological activities. Toxicon 2013, 69, 75–81. [Google Scholar] [CrossRef]
- Tan, C.H.; Tan, K.Y.; Yap, M.K.K.; Tan, N.H. Venomics of Tropidolaemus wagleri, the sexually dimorphic temple pit viper: Unveiling a deeply conserved atypical toxin arsenal. Sci. Rep. 2017, 7, 43237. [Google Scholar] [CrossRef]
- Amorim, F.G.; Costa, T.R.; Baiwir, D.; De Pauw, E.; Quinton, L.; Sampaio, S.V. Proteopeptidomic, functional and immunoreactivity characterization of Bothrops moojeni snake venom: Influence of snake gender on venom composition. Toxins 2018, 10, 177. [Google Scholar] [CrossRef] [PubMed]
- Freitas-de-Sousa, L.A.; Nachtigall, P.G.; Portes-Junior, J.A.; Holding, M.L.; Nystrom, G.S.; Ellsworth, S.A.; Guimarães, N.C.; Tioyama, E.; Ortiz, F.; Silva, B.R.; et al. Size Matters: An evaluation of the molecular basis of ontogenetic modifications in the composition of Bothrops jararacussu snake venom. Toxins 2020, 12, 791. [Google Scholar] [CrossRef]
- Pe, T.; Aye, B.; Myint, A.-A.; Swe, T.-N.; Warrell, D. Effect of the snake length’s and recent feeding on venom antigenaemia and severity of envenoming in Russell’s viper (Daboia russelii siamensis) bites. In Management of Snakebite and Research; WHO, Regional Office for South-East-Asia: Geneva, Switzerland, 2002; pp. 41–45. [Google Scholar]
- Clemetson, K.J.; Morita, T.; Manjunatha Kini, R. Scientific and standardization committee communications: Classification and nomenclature of snake venom C-type lectins and related proteins. J. Thromb. Haemost. 2009, 7, 360. [Google Scholar] [CrossRef] [PubMed]
- Clemetson, K.J. Snaclecs (snake C-type lectins) that inhibit or activate platelets by binding to receptors. Toxicon 2010, 56, 1236–1246. [Google Scholar] [CrossRef] [PubMed]
- Morita, T. Structures and functions of snake venom CLPs (C-type lectin-like proteins) with anticoagulant-, procoagulant-, and platelet-modulating activities. Toxicon 2005, 45, 1099–1114. [Google Scholar] [CrossRef]
- Teixeira, C.; Fernandes, C.M.; Leiguez, E.; Chudzinski-Tavassi, A.M. Inflammation Induced by platelet-activating Viperid snake venoms: Perspectives on thromboinflammation. Front. Immunol. 2019, 10, 2082. [Google Scholar] [CrossRef]
- Zhong, S.R.; Jin, Y.; Wu, J.B.; Chen, R.Q.; Jia, Y.H.; Wang, W.Y.; Xiong, Y.L.; Zhang, Y. Characterization and molecular cloning of dabocetin, a potent antiplatelet C-type lectin-like protein from Daboia russellii siamensis venom. Toxicon 2006, 47, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Zhong, S.; Jin, Y.; Li, D.; Wang, W.; Xiong, Y. Molecular cloning and sequence analyses of cDNAs encoding seven C-type lectin-like protein subunits from Daboia russellii siamensis. Zool. Res. 2005, 26, 337–343. [Google Scholar]
- Jebali, J.; Bazaa, A.; Sarray, S.; Benhaj, K.; Karboul, A.; El Ayeb, M.; Marrakchi, N.; Gargouri, A. C-type lectin protein isoforms of Macrovipera lebetina: cDNA cloning and genetic diversity. Toxicon 2009, 53, 228–237. [Google Scholar] [CrossRef]
- Inagaki, H. Snake venom protease inhibitors: Enhanced identification, expanding biological function, and promising future. In Snake Venoms; Inagaki, H., Vogel, C.-W., Mukherjee, A.K., Rahmy, T.R., Gopalakrishnakone, P., Eds.; Springer: Dordrecht, The Netherlands, 2015; pp. 1–26. [Google Scholar] [CrossRef]
- Thakur, R.; Mukherjee, A.K. Pathophysiological significance and therapeutic applications of snake venom protease inhibitors. Toxicon 2017, 131, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Vasconcelos, A.A.; Estrada, J.C.; David, V.; Wermelinger, L.S.; Almeida, F.C.L.; Zingali, R.B. Structure-function relationship of the disintegrin family: Sequence signature and integrin interaction. Front. Mol. Biosci. 2021, 8, 783301. [Google Scholar] [CrossRef] [PubMed]
- Juárez, P.; Wagstaff, S.C.; Sanz, L.; Harrison, R.A.; Calvete, J.J. Molecular cloning of Echis ocellatus disintegrins reveals non-venom-secreted proteins and a pathway for the evolution of ocellatusin. J. Mol. Evol. 2005, 63, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Sanz-Soler, R.; Sanz, L.; Calvette, J.J. Distribution of RPTNL genes across Reptilia: Hypothesized role for RPTLN in the evolution of SVMPs. Integr. Comp. Biol. 2016, 56, 989–1003. [Google Scholar] [CrossRef]
- Sanz-Soler, R.; Lorente, C.; Company, B.; Sanz, L.; Juarez, P.; Perez, A.; Zhang, Y.; Jin, Y.; Chen, R.; Eble, J.A.; et al. Recombinant expression of mutants of the Frankenstein disintegrin, RTS-ocellatusin. Evidence for the independent origin of RGD and KTS/RTS disintegins. Toxicon 2012, 60, 665–675. [Google Scholar] [CrossRef]
- Calvete, J.J. Structure-function correlations of snake venom disintegrins. Curr. Pharm. Des. 2005, 11, 829–835. [Google Scholar] [CrossRef] [PubMed]
- Wagstaff, S.C.; Favreau, P.; Cheneval, O.; Laing, G.D.; Wilkinson, M.C.; Miller, R.L.; Stöcklin, R.; Harrison, R.A. Molecular characterisation of endogenous snake venom metalloproteinase inhibitors. Biochem. Biophys. Res. Commun. 2008, 365, 650–656. [Google Scholar] [CrossRef] [PubMed]
- Munawar, A.; Ali, S.A.; Akrem, A.; Betzel, C. Snake venom peptides: Tools of biodiscovery. Toxins 2018, 10, 474. [Google Scholar] [CrossRef] [PubMed]
- Averin, A.S.; Utkin, Y.N. Cardiovascular Effects of Snake Toxins: Cardiotoxicity and Cardioprotection. Acta Nat. 2021, 13, 4–14. [Google Scholar] [CrossRef]
- Frangieh, J.; Rima, M.; Fajloun, Z.; Henrion, D.; Sabatier, J.M.; Legros, C.; Mattei, C. Snake venom components: Tools and cures to target cardiovascular diseases. Molecules 2021, 26, 2223. [Google Scholar] [CrossRef]
- Coronado, M.A.; de Moraes, F.R.; Ullah, A.; Masood, R.; Mariutti, R.; Brognaro, H.; Georgieva, D.; Murakami, M.T.; Betzel, C.; Arni, R.K. Three-dimensional structures and mechanism of snake venom serine proteinases, metalloproteinases, and phospholipase A2s. In Venom Genomics and Proteomics; Gopalakrishnakone, P., Calvete, J., Eds.; Springer: Dordrecht, The Netherlands, 2016; pp. 239–267. [Google Scholar] [CrossRef]
- Chaisakul, J.; Khow, O.; Wiwatwarayos, K.; Rusmili, M.R.A.; Prasert, W.; Othman, I.; Abidin, S.A.Z.; Charoenpitakchai, M.; Hodgson, W.C.; Chanhome, L.; et al. A biochemical and pharmacological characterization of phospholipase A2 and metalloproteinase fractions from Eastern Russell’s viper (Daboia siamensis) venom: Two major components associated with acute kidney injury. Toxins 2021, 13, 521. [Google Scholar] [CrossRef]
- Tsai, I.-H.; Tasi, H.-Y.; Wang, Y.-M.; Pe, T.; Warrel, D.A. Venom phospholipases of Russell’s vipers from Myanmar and eastern India-Cloning, characterizarion and phylogeographic analysis. Biochim. Biophys. Acta. 2007, 1774, 1020–1028. [Google Scholar] [CrossRef]
- Thwin, M.-M.; Gopalakrishnakone, P.; Yuen, R.; Tan, C.H. A major lethal factor of the venom of Burmese Russell’s viper (Daboia russellil siamensis): Isolation, n-terminal sequencing and biological activities of daboiatoxin. Toxicon 1995, 33, 63–76. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, W.; Gao, R.; Lu, Q.; Wang, W.; Xiong, Y. Structure-function features and effects on blood coagulation of snake venom serine proteases. Zool. Res. 1998, 19, 97–106. [Google Scholar]
- Kang, T.S.; Georgieva, D.; Genov, N.; Murakami, M.T.; Sinha, M.; Kumar, R.P.; Kaur, P.; Kumar, S.; Dey, S.; Sharma, S.; et al. Enzymatic toxins from snake venom: Structural characterization and mechanism of catalysis. FEBS J. 2011, 278, 4544–4576. [Google Scholar] [CrossRef]
- Serrano, S.M.T.; Maroun, R.C. Snake venom serine proteinases: Sequence homology vs. substrate specificity, a paradox to be solved. Toxicon 2005, 45, 1115–1132. [Google Scholar] [CrossRef]
- Thakur, R.; Mukherjee, A.K. A brief appraisal on Russell’s viper venom (Daboia russelii russelii) proteinases. In Snake Venoms; Inagaki, H., Vogel, C.W., Mukherjee, A., Rahmy, T., Eds.; Springer: Dordrecht, The Netherlands, 2017; pp. 123–144. [Google Scholar] [CrossRef]
- Gâz Florea, Ş.A.; Gâz Florea, A.; Kelemen, H.; Muntean, D.-L. Snake Venom Metalloproteinases. Acta Medica Marisiensis 2016, 62, 106–111. [Google Scholar] [CrossRef]
- Takeda, S.; Takeya, H.; Iwanaga, S. Snake venom metalloproteinases: Structure, function and relevance to the mammalian ADAM/ADAMTS family proteins. Biochim. Biophys. Acta Proteins Proteom. 2012, 1824, 164–176. [Google Scholar] [CrossRef] [PubMed]
- Chellapandi, P. Structural, functional and therapeutic aspects of snake venom metalloproteinases. Mini. Rev. Org. Chem. 2014, 11, 28–44. [Google Scholar] [CrossRef]
- Takeda, S.; Igarashi, T.; Mori, H. Crystal structure of RVV-X: An example of evolutionary gain of specificity by ADAM proteinases. FEBS Lett. 2007, 581, 5859–5864. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.S.; Chen, J.M.; Lin, C.W.; Khoo, K.H.; Tsai, I.H. New insights into the functions and N-glycan structures of factor X activator from Russell’s viper venom. FEBS J. 2008, 275, 3944–3958. [Google Scholar] [CrossRef] [PubMed]
- Suntravat, M.; Yusuksawad, M.; Sereemaspun, A.; Pérez, J.C.; Nuchprayoon, I. Effect of purified Russell’s viper venom-factor X activator (RVV-X) on renal hemodynamics, renal functions, and coagulopathy in rats. Toxicon 2011, 58, 230–238. [Google Scholar] [CrossRef]
- Chen, H.S.; Tsai, H.Y.; Wang, Y.M.; Tsai, I.H. P-III hemorrhagic metalloproteinases from Russell’s viper venom: Cloning, characterization, phylogenetic and functional site analyses. Biochimie 2008, 90, 1486–1498. [Google Scholar] [CrossRef] [PubMed]
- Takeda, S. ADAM and ADAMTS family proteins and snake venom metalloproteinases: A structural overview. Toxins 2016, 8, 155. [Google Scholar] [CrossRef]
- Hikita, A.; Tanaka, N.; Yamane, S.; Ikeda, Y.; Furukawa, H.; Tohma, S.; Suzuki, R.; Tanaka, S.; Mitomi, H.; Fukui, N. Involvement of a disintegrin and metalloproteinase 10 and 17 in shedding of tumor necrosis factor-α. Biochem. Cell Biol. 2009, 87, 581–593. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, Y.; Matsunaga, Y.; Tokunaga, Y.; Obayashi, S.; Saito, M.; Morita, T. Snake venom vascular endothelial growth factors (VEGF-Fs) exclusively vary their structures and functions among species. J. Biol. Chem. 2009, 284, 9885–9891. [Google Scholar] [CrossRef]
- Ferreira, I.G.; Pucca, M.B.; de Oliveira, I.S.; Cerni, F.A.; de Cássia da Silva Jacob, B.; Arantes, E.C. Snake venom vascular endothelial growth factors (svVEGFs): Unravelling their molecular structure, functions, and research potential. Cytokine Growth Factor Rev. 2021, 60, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, Y.; Takani, K.; Atoda, H.; Morita, T. Snake venom vascular endothelial growth factors (VEGFs) exhibit potent activity through their specific recognition of KDR (VEGF receptor 2). J. Biol. Chem. 2003, 278, 51985–51988. [Google Scholar] [CrossRef]
- Tokunaga, Y.; Yamazaki, Y.; Morita, T. Specific distribution of VEGF-F in Viperinae snake venoms: Isolation and characterization of a VEGF-F from the venom of Daboia russelli siamensis. Arch. Biochem. Biophys. 2005, 439, 241–247. [Google Scholar] [CrossRef]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef] [PubMed]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed]
- Macrander, J.; Panda, J.; Janies, D.; Daly, M.; Reitzel, A.M. Venomix: A simple bioinformatic pipeline for identifying and characterizing toxin gene candidates from transcriptomic data. Peer J. 2018, 6, e5361. [Google Scholar] [CrossRef]
- Jungo, F.; Bairoch, A.; Prot, T. the toxin protein annotation program of the Swiss-Prot protein knowledgebase. Toxicon 2005, 45, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M.; et al. De novo transcript sequence reconstruction from RNA-Seq: Reference generation and analysis with Trinity. Nat Protoc. 2013, 8, 8. [Google Scholar] [CrossRef]
- Gacesa, R.; Barlow, D.J.; Long, P.F. Machine learning can differentiate venom toxins from other proteins having non-toxic physiological functions. PeerJ Comput. Sci. 2016, 2, e90. [Google Scholar] [CrossRef]
- UniProt Consortium. UniProt: The universal protein knowledgebase in 2021. Nucleic Acids Res. 2021, 49, D480–D489. [Google Scholar] [CrossRef] [PubMed]
- Madeira, F.; Pearce, M.; Tivey, A.R.N.; Basutkar, P.; Lee, J.; Edbali, O.; Madhusoodanan, N.; Kolesnikov, A.; Lopez, R. Search and sequence analysis tools services from EMBL-EBI in 2022. Nucleic Acids Res. 2022, 50, W276–W279. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Bembom, O.; Ivanek, R. seqLogo: Sequence logos for DNA sequence alignments. R pakage version 1.64.0. Available online: https://bioconductor.org/packages/release/bioc/html/seqLogo.html (accessed on 26 March 2022).
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trinity Assembly Parameters | Male | Female |
|---|---|---|
| Total raw reads | 82,251,232 | 79,578,046 |
| Total clean reads | 78,647,670 | 74,444,200 |
| Total clean nucleotides (nt) | 7,752,565,867 | 7,235,655,822 |
| Q20 percentage | 98.93% | 98.55% |
| GC percentage | 46.26% | 53.43% |
| Contigs created | 88,325 | 50,858 |
| Total assembled bases | 75,163,976 | 39,200,694 |
| Mean length (nt) | 850.99 | 770.79 |
| N50 | 1870 | 1379 |
| Unigenes/transcripts assembled (only longest isoform per gene) | 77,668 | 46,884 |
| No. | Contig ID | Annotation | Accession No. | Species | Amino Acid | TPM Value |
|---|---|---|---|---|---|---|
| 1 | c17840_g2_i1_M_FL | Kunitz-type serine protease inhibitor 1 | Q2ES50 | Daboia russelii | 84 | 109,755.22 |
| c11654_g1_i1_F_FL | 16,745.60 | |||||
| 2 | c6152_g1_i1_M_FL | Serine protease inhibitor 1 | A0A6G5ZVV7 | Vipera anatolica senliki | 148 | 1.59 |
| c6152_g1_i2_M_FL | 0.76 | |||||
| 3 | c3688_g1_i1_M_FL | Serine protease inhibitor 4 | A0A6G5ZUI5 | Vipera anatolica senliki | 514 | 26.46 |
| c5062_g2_i1_F_FL | 9.09 | |||||
| 4 | c12297_g1_i1_M_FL | Tissue factor pathway inhibitor | U3TBN9 | Protobothrops flavoviridis | 303 | 2.45 |
| 5 | c1956_g1_i1_M_PL | Serine protease inhibitor 3 | A0A6G5ZW53 | Vipera anatolica senliki | 103 | 1.48 |
| 6 | c12495_g1_i1_M_PL | Serine protease inhibitor 6 | A0A6G5ZUI2 | Vipera anatolica senliki | 272 | 56.61 |
| c12495_g1_i2_M_PL | 50.11 | |||||
| c12495_g1_i3_M_PL | 74.49 | |||||
| c2247_g1_i2_F_PL | 51.7 | |||||
| c2247_g1_i1_F_PL | 33.08 | |||||
| 7 | c15981_g1_i1_M_PL | kunitz/BPTI-like toxin | A0A6P9AX37 | Protobothrops guttatus | 139 | 47.25 |
| 8 | c11558_g3_i1_F_PL | kunitz/BPTI-like toxin | XP_015682430.1 | Protobothrops mucrosquamatus | 139 | 25.45 |
| No. | Contig ID | Annotation | Accession No. | Species | Amino Acid | TPM Value |
|---|---|---|---|---|---|---|
| 1 | c20548_g3_i4_M_FL | Factor V activator RVV-Vγ | P18965 | Daboia siamensis | 259 | 5495.65 |
| 2 | c11820_g1_i3_F_PL | Thrombin-like_enzyme_TLBm (Factor V activator RVV-Vγ) | P18965 | D. siamensis | 251 | 2205.37 |
| 3 | c11820_g1_i1_F_PL | Thrombin-like_enzyme_TLBm (Factor V activator RVV-Vγ) | P18965 | D. siamensis | 241 | 1794.30 |
| 4 | c11820_g1_i4_F_PL | Thrombin-like_enzyme_TLBm (Factor V activator RVV-Vγ) | P18965 | D. siamensis | 251 | 1355.44 |
| 5 | c38983_g1_i1_F_PL | α-fibrinogenase-like | E5L0E3 | D. siamensis | 105 | 3768.67 |
| 6 | c20548_g3_i1_M_PL | α-fibrinogenase-like | E5L0E3 | D. siamensis | 131 | 3196.39 |
| 7 | c20548_g3_i3_M_PL | α-fibrinogenase-like | E5L0E3 | D. siamensis | 155 | 952.41 |
| 8 | c11820_g1_i9_F_PL | Thrombin-like_enzyme_catroxobin-1 (β-fibrinogenase-like) | E5L0E4 | D. siamensis | 126 | 981.63 |
| 9 | c20548_g3_i5_M_FL | Serine protease VLSP-1 | E0Y418 | Microvipera lebetina | 265 | 3027.55 |
| 10 | c11820_g1_i8_F_PL | Thrombin-like_enzyme_TLBm (Serine protease VLSP-1) | E0Y418 | M. lebetina | 242 | 1400.97 |
| 11 | c11820_g1_i6_F_PL | Thrombin-like_enzyme_TLBm (Serine protease VLSP-1) | E0Y418 | M. lebetina | 189 | 816.73 |
| 12 | c11820_g1_i2_F_PL | Thrombin-like_enzyme_TLBm (Serine protease VLSP-1) | E0Y418 | M. lebetina | 126 | 448.44 |
| 13 | c11820_g1_i5_F_PL | Thrombin-like_enzyme_TLBm (Serine protease VLSP-3) | E0Y420 | M. lebetina | 134 | 1318.30 |
| 14 | c11820_g1_i7_F_PL | Thrombin-like_enzyme_catroxobin-1 (β-fibrinogenase-like) | E5L0E4 | D. siamensis | 104 | 919.53 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yee, K.T.; Macrander, J.; Vasieva, O.; Rojnuckarin, P. Exploring Toxin Genes of Myanmar Russell’s Viper, Daboia siamensis, through De Novo Venom Gland Transcriptomics. Toxins 2023, 15, 309. https://doi.org/10.3390/toxins15050309
Yee KT, Macrander J, Vasieva O, Rojnuckarin P. Exploring Toxin Genes of Myanmar Russell’s Viper, Daboia siamensis, through De Novo Venom Gland Transcriptomics. Toxins. 2023; 15(5):309. https://doi.org/10.3390/toxins15050309
Chicago/Turabian StyleYee, Khin Than, Jason Macrander, Olga Vasieva, and Ponlapat Rojnuckarin. 2023. "Exploring Toxin Genes of Myanmar Russell’s Viper, Daboia siamensis, through De Novo Venom Gland Transcriptomics" Toxins 15, no. 5: 309. https://doi.org/10.3390/toxins15050309
APA StyleYee, K. T., Macrander, J., Vasieva, O., & Rojnuckarin, P. (2023). Exploring Toxin Genes of Myanmar Russell’s Viper, Daboia siamensis, through De Novo Venom Gland Transcriptomics. Toxins, 15(5), 309. https://doi.org/10.3390/toxins15050309