Intraspecific Diversity and Pathogenicity of Bacillus thuringiensis Isolates from an Emetic Illness

 , , , ,
, , , ,
Abstract
1. Introduction
2. Results and Discussion
2.1. Morphological and Biochemical Characterization of Food-Borne Outbreak Isolates
2.2. Molecular Identification and Typing of Isolates from Emetic Illness
2.2.1. Identification of Isolates Using 16S rRNA Gene Sequence Analysis
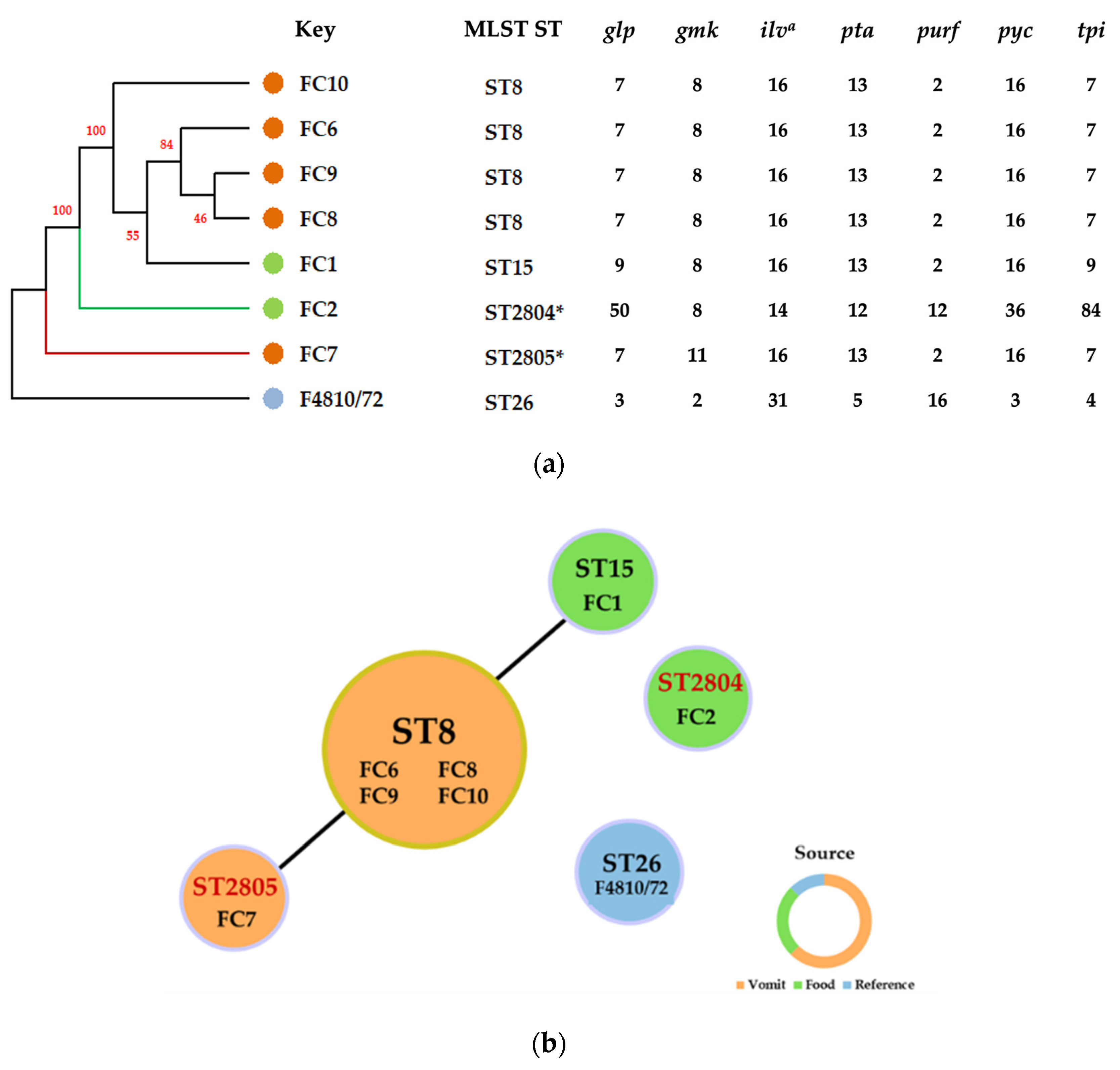
2.2.2. Molecular Typing of Isolates Using Multilocus Sequence Typing (MLST) Analysis
2.3. Detection and Characterization of Genes Associated with Emetic Traits in B. thuringiensis Isolates
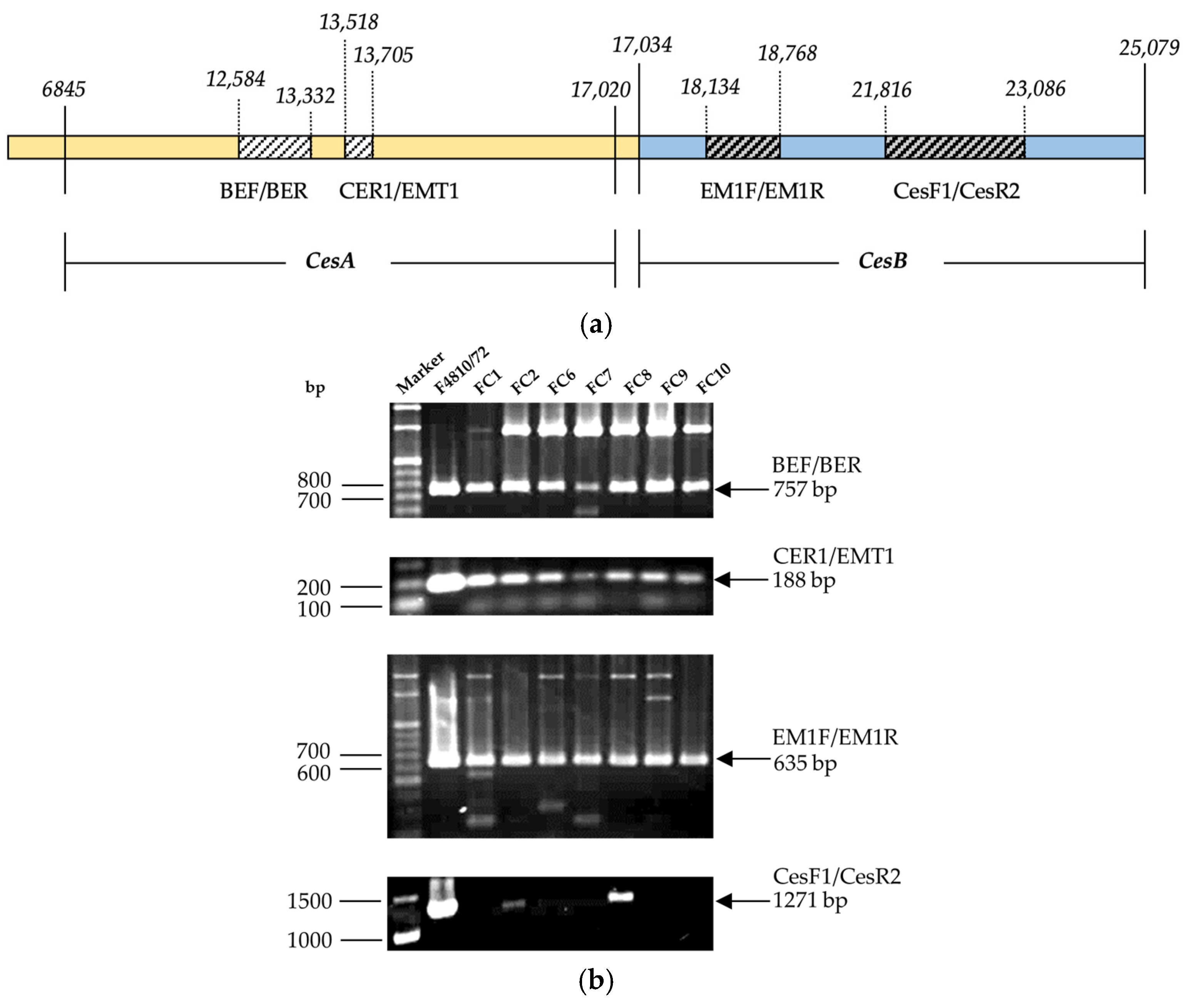
2.3.1. Detection of Non-Ribosomal Peptide Synthetase (NRPS)/Cereulide Synthetase Genes
2.3.2. Sequence Analysis of B. thuringiensis Amplified Gene Fragments
2.4. Cytotoxicity of Emetic B. thuringiensis Isolates to Caco-2 Cells
2.5. Annexin V/PI Apoptosis Detection
3. Conclusions
4. Materials and Methods
4.1. Bacterial Strains/Isolates and Identification
4.2. Identification and Typing of B. thuringiensis Isolates from Emetic Food-Borne Illness
4.2.1. Identification of Isolates Using 16S rRNA Gene Sequencing
4.2.2. Typing of Isolates Using Multilocus Sequence Typing (MLST) Analysis
4.3. Genetic Analysis of B. thuringiensis Isolates
4.3.1. DNA Extraction
4.3.2. Polymerase Chain Reaction (PCR) Amplification of Cereulide Synthetase Gene
4.3.3. DNA Sequencing and Sequence Similarity Analysis
4.4. Cytotoxicity Assay
4.4.1. Preparation of Heat-Treated Supernatants from Bacterial Isolates
4.4.2. Cell Culture
4.4.3. Cytotoxicity Assay
4.4.4. Annexin V/PI Assay
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stenfors Arnesen, L.P.; Fagerlund, A.; Granum, P.E. From soil to gut: Bacillus cereus and its food poisoning toxins. FEMS Microbiol. Rev. 2008, 32, 579–606. [Google Scholar] [CrossRef]
- Ehling-Schulz, M.; Lereclus, D.; Koehler, T.M. The Bacillus cereus group: Bacillus species with pathogenic potential. Microbiol. Spectr. 2019, 7, 1–60. [Google Scholar] [CrossRef]
- McIntyre, L.; Bernard, K.; Beniac, D.; Isaac-Renton, J.L.; Naseby, D.C. Identification of Bacillus cereus group species associated with food poisoning outbreaks in British Columbia, Canada. Appl. Environ. Microbiol. 2008, 74, 7451–7453. [Google Scholar] [CrossRef]
- Ceuppens, S.; Boon, N.; Uyttendaele, M. Diversity of Bacillus cereus group strains is reflected in their broad range of pathogenicity and diverse ecological lifestyles. FEMS Microbiol. Ecol. 2013, 84, 433–450. [Google Scholar] [CrossRef]
- Logan, N.A. Bacillus and relatives in food-borne illness. J. Appl. Microbiol. 2012, 112, 417–429. [Google Scholar] [CrossRef]
- Helgason, E.; Økstad, O.A.; Caugant, D.A.; Johansen, H.A.; Fouet, A.; Mock, M.; Hegna, I.; Kolstø, A.B. Bacillus anthracis, Bacillus cereus, and Bacillus thuringiensis—One species on the basis of genetic evidence. Appl. Environ. Microbiol. 2000, 66, 2627–2630. [Google Scholar] [CrossRef]
- Bravo, A.; Likitvivatanavong, S.; Gill, S.S.; Soberón, M. Bacillus thuringiensis: A story of a successful bioinsecticide. Insect Biochem. Mol. Biol. 2011, 41, 423–431. [Google Scholar] [CrossRef]
- Jeong, H.; Jo, S.H.; Hong, C.E.; Park, J.M. Genome sequence of the endophytic bacterium Bacillus thuringiensis strain KB1, a potential biocontrol agent against phytopathogens. Microbiol. Resour. Announce. 2016, 21, e00279-16. [Google Scholar] [CrossRef]
- Valtierra de Luis, D.; Villanueva, M.; Berry, C.; Caballero, P. Potential for Bacillus thuringiensis and other bacterial toxins as biological control agents to combat dipteran pests of medical and agronomic importance. Toxins 2020, 12, 773. [Google Scholar] [CrossRef]
- Sanchis, V.; Bourguet, D. Bacillus thuringiensis: Applications in agriculture and insect resistance management. A review. Agron. Sustain. 2008, 28, 11–20. [Google Scholar] [CrossRef]
- Abbas, M.S.T. Genetically engineered (modified) crops (Bacillus thuringiensis crops) and the world controversy on their safety. Egypt J. Biol. Pest Control 2018, 28, 52. [Google Scholar] [CrossRef]
- Rosenquist, H.; Smidt, L.; Andersen, S.R.; Jensen, G.B.; Wilcks, A. Occurrence and significance of Bacillus cereus and Bacillus thuringiensis in ready-to-eat food. FEMS Microbiol. Lett. 2005, 250, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Frederiksen, K.; Rosenquist, H.; Jørgensen, K.; Wilcks, A. Occurrence of natural Bacillus thuringiensis contaminants and residues of Bacillus thuringiensis-based insecticides on fresh fruits and vegetables. Appl. Environ. Microbiol. 2006, 72, 3435–3440. [Google Scholar] [CrossRef]
- Jovanovic, J.; Ornelis, V.F.M.; Madder, A.; Rajkovic, A. Bacillus cereus food intoxication and toxicoinfection. Compr. Rev. Food Sci. Food Saf. 2021, 20, 3719–3761. [Google Scholar] [CrossRef]
- Kim, J.B.; Choi, O.K.; Kwon, S.M.; Cho, S.H.; Park, B.J.; Jin, N.Y.; Yu, Y.M.; Oh, D.H. Prevalence and toxin characteristics of Bacillus thuringiensis isolated from organic vegetables. J. Microbiol. Biotechnol. 2017, 27, 1449–1456. [Google Scholar] [CrossRef]
- Johler, S.; Kalbhenn, E.M.; Heini, N.; Brodmann, P.; Gautsch, S.; Bağcioğlu, M.; Contzen, M.; Stephan, R.; Ehling-Schulz, M. Enterotoxin production of Bacillus thuringiensis isolates from biopesticides, foods, and outbreaks. Front. Microbiol. 2018, 9, 1915. [Google Scholar] [CrossRef]
- Jackson, S.G.; Goodbrand, R.B.; Ahmed, R.; Kasatiya, S. Bacillus cereus and Bacillus thuringiensis isolated in a gastroenteritis outbreak investigation. Lett. Appl. Microbiol. 1995, 21, 103–105. [Google Scholar] [CrossRef]
- Bonis, M.; Felten, A.; Pairaud, S.; Dijoux, A.; Maladen, V.; Mallet, L.; Radomski, N.; Duboisset, A.; Arar, C.; Sarda, X.; et al. Comparative phenotypic, genotypic, and genomic analyses of Bacillus thuringiensis associated with food-borne outbreaks in France. PLoS ONE 2021, 19, 16. [Google Scholar] [CrossRef]
- Schwenk, V.; Riegg, J.; Lacroix, M.; Märtlbauer, E.; Jessberger, N. Enteropathogenic potential of Bacillus thuringiensis isolates from soil, animals, food and biopesticides. Foods 2020, 9, 1484. [Google Scholar] [CrossRef]
- Mahler, H.; Pasi, A.; Kramer, J.M.; Schulte, P.; Scoging, A.C.; Bär, W.; Krähenbühl, S. Fulminant liver failure in association with the emetic toxin of Bacillus cereus. N. Engl. J. Med. 1997, 336, 1142–1148. [Google Scholar] [CrossRef]
- Dierick, K.; Coillie, E.V.; Swiecicka, I.; Meyfroidt, G.; Devlieger, H.; Meulemans, A.; Hoedemaekers, G.; Fourie, L.; Heyndrickx, M.; Mahillon, J. Fatal family outbreak of Bacillus cereus-associated food poisoning. J. Clin. Microbiol. 2005, 43, 4277–4279. [Google Scholar] [CrossRef] [PubMed]
- Naranjo, M.; Denayer, S.; Botteldoorn, N.; Delbrassinne, L.; Veys, J.; Waegenaere, J.; Sirtaine, N.; Driesen, R.B.; Sipido, K.R.; Mahillon, J.; et al. Sudden death of a young adult associated with Bacillus cereus food poisoning. J. Clin. Microbiol. 2011, 49, 4379–4381. [Google Scholar] [CrossRef] [PubMed]
- Rodrigo, D.; Rosell, C.M.; Martinez, A. Risk of Bacillus cereus in relation to rice and derivatives. Foods 2021, 10, 302. [Google Scholar] [CrossRef]
- Messelhäusser, U.; Frenzel, E.; Blöchinger, C.; Zucker, R.; Kämpf, P.; Ehling-Schulz, M. Emetic Bacillus cereus are more volatile than thought: Recent food-borne outbreaks and prevalence studies in Bavaria (2007–2013). Biomed. Res. Int. 2014, 2014, 465603. [Google Scholar] [CrossRef]
- Logan, N.A.; Berkeley, R.C. Identification of Bacillus strains using the API system. J. Gen. Microbiol. 1984, 130, 1871–1882. [Google Scholar] [CrossRef]
- Ehling-Schulz, M.; Svensson, B.; Guinebretiere, M.H.; Lindbäck, T.; Andersson, M.; Schulz, A.; Fricker, M.; Christiansson, A.; Granum, P.E.; Märtlbauer, E.; et al. Emetic toxin formation of Bacillus cereus is restricted to a single evolutionary lineage of closely related strains. Microbiology 2005, 151, 183–197. [Google Scholar] [CrossRef]
- Yang, Y.; Foster, J.T.; Yi, M.; Zhan, L.; Zhang, Y.; Zhou, B.; Jiang, J.; Mei, L. Phenotypic homogeneity of emetic Bacillus cereus isolates in China. Lett. Appl. Microbiol. 2021, 73, 646–651. [Google Scholar] [CrossRef] [PubMed]
- Thorsen, L.; Hansen, B.M.; Nielsen, K.F.; Hendriksen, N.B.; Phipps, R.K.; Budde, B.B. Characterization of emetic Bacillus weihenstephanensis, a new cereulide-producing bacterium. Appl. Environ. Microbiol. 2006, 72, 5118–5121. [Google Scholar] [CrossRef] [PubMed]
- de Bock, T.; Zhao, X.; Jacxsens, L.; Devlieghere, F.; Rajkovic, A.; Spanoghe, P.; Höfte, M.; Uyttendaele, M. Evaluation of B. thuringiensis-based biopesticides in the primary production of fresh produce as a food safety hazard and risk. Food Control 2021, 130, 108390. [Google Scholar] [CrossRef]
- Yang, Y.; Yu, X.; Zhan, L.; Chen, J.; Zhang, Y.; Zhang, J.; Chen, H.; Zhang, Z.; Zhang, Y.; Lu, Y.; et al. Multilocus sequence type profiles of Bacillus cereus isolate from infant formula in China. Food Microbiol. 2017, 62, 46–50. [Google Scholar] [CrossRef]
- Liu, Y.; Lai, Q.; Du, J.; Shao, Z. Genetic diversity, and population structure of the Bacillus cereus group bacteria from diverse marine environments. Sci. Rep. 2017, 7, 689. [Google Scholar] [CrossRef] [PubMed]
- Carroll, L.M.; Wiedmann, M.; Mukherjee, M.; Nicholas, D.C.; Mingle, L.A.; Dumas, N.B.; Cole, J.A.; Kovac, J. Characterization of emetic and diarrheal Bacillus cereus strains from a 2016 food-borne outbreak using whole-genome sequencing: Addressing the microbiological, epidemiological, and bioinformatic challenges. Front. Microbiol. 2019, 10, 144. [Google Scholar] [CrossRef] [PubMed]
- Hoffmaster, A.R.; Novak, R.T.; Marston, C.K.; Gee, J.E.; Helsel, L.; Pruckler, J.M.; Wilkins, P.P. Genetic diversity of clinical isolates of Bacillus cereus using multilocus sequence typing. BMC Microbiol. 2008, 8, 191. [Google Scholar] [CrossRef]
- Biggel, M.; Jessberger, N.; Kovac, J.; Johler, S. Recent paradigm shifts in the perception of the role of Bacillus thuringiensis in food-borne disease. Food Microbiol. 2022, 105, 104025. [Google Scholar] [CrossRef]
- Raymond, B.; Federici, B.A. In defense of Bacillus thuringiensis, the safest and most successful microbial insecticide available to humanity—A response to EFSA. FEMS Microbiol. Ecol. 2017, 93, fix084. [Google Scholar] [CrossRef]
- Frentzel, H.; Juraschek, K.; Pauly, N.; Kelner-Burgos, Y.; Wichmann-Schauer, H. Indications of biopesticidal Bacillus thuringiensis strains in bell pepper and tomato. Int. J. Food Microbiol. 2020, 321, 108542. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Shu, C.; Soberón, M.; Bravo, A.; Zhang, J. Systematic characterization of Bacillus Genetic Stock Center Bacillus thuringiensis strains using Multi-Locus Sequence Typing. J. Invertebr. Pathol. 2018, 155, 5–13. [Google Scholar] [CrossRef]
- Ehling-Schulz, M.; Fricker, M.; Grallert, H.; Rieck, P.; Wagner, M.; Scherer, S. Cereulide synthetase gene cluster from emetic Bacillus cereus: Structure and location on a mega virulence plasmid related to Bacillus anthracis toxin plasmid pXO1. BMC Microbiol. 2006, 6, 20. [Google Scholar] [CrossRef]
- Toh, M.; Moffitt, M.C.; Henrichsen, L.; Raftery, M.; Barrow, K.; Cox, J.M.; Marquis, C.P.; Neilan, B.A. Cereulide, the emetic toxin of Bacillus cereus, is putatively a product of nonribosomal peptide synthesis. J. Appl. Microbiol. 2004, 97, 992–1000. [Google Scholar] [CrossRef]
- Horwood, P.F.; Burgess, G.W.; Oakey, H.J. Evidence for non-ribosomal peptide synthetase production of cereulide (the emetic toxin) in Bacillus cereus. FEMS Microbiol. Lett. 2004, 236, 319–324. [Google Scholar] [CrossRef]
- Ehling-Schulz, M.; Fricker, M.; Scherer, S. Identification of emetic toxin producing Bacillus cereus strains by a novel molecular assay. FEMS Microbiol. Lett. 2004, 232, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Ehling-Schulz, M.; Vukov, N.; Schulz, A.; Shaheen, R.; Andersson, M.; Märtlbauer, E.; Scherer, S. Identification and partial characterization of the nonribosomal peptide synthetase gene responsible for cereulide production in emetic Bacillus cereus. Appl. Environ. Microbiol. 2005, 71, 105–113. [Google Scholar] [CrossRef]
- Kim, J.B.; Kim, J.M.; Park, Y.B.; Han, J.A.; Lee, S.H.; Kwak, H.S.; Hwang, I.G.; Yoon, M.H.; Lee, J.B.; Oh, D.H. Evaluation of various PCR assays for the detection of emetic toxin producing Bacillus cereus. J. Microbiol. Biotechnol. 2010, 20, 1107–1113. Available online: https://pubmed.ncbi.nlm.nih.gov/20668404/ (accessed on 1 December 2022). [PubMed]
- Mei, X.; Xu, K.; Yang, L.; Yuan, Z.; Mahillon, J.; Hu, X. The genetic diversity of cereulide biosynthesis gene cluster indicates a composite transposon Tnces in emetic Bacillus weihenstephanensis. BMC Microbiol. 2014, 14, 149. [Google Scholar] [CrossRef]
- Ehling-Schulz, M.; Frenzel, E.; Gohar, M. Food-bacteria interplay: Pathometabolism of emetic Bacillus cereus. Front. Microbiol. 2015, 6, 704. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, T.; Itoh, Y.; Hayashi, S.; Sadakari, K.; Kamikado, H.; Honjoh, K.; Kobayashi, H. Bacillus cereus crs1, crs2 Genes for Cereulide Synthetase 1, Cereulide Synthetase 2, Complete cds. Genbank Nucleotide Database Accession no. AB 248763.2, National Center for Biotechnology Information, U.S. National Library of Medicine, U.S.A. Available online: http://www.ncbi.nlm.nih.gov/nuccore/AB248763.2 (accessed on 1 December 2022).
- Nieminen, T.; Rintaluoma, N.; Andersson, M.; Taimisto, A.-M.; Ali-Vehmas, T.; Seppälä, A.; Priha, O.; Salkinoja-Salonen, M. Toxinogenic Bacillus pumilus and Bacillus licheniformis from mastitic milk. Vet. Microbiol. 2007, 124, 329–339. [Google Scholar] [CrossRef]
- Parvathi, A.; Krishna, K.; Jose, J.; Joseph, N.; Nair, S. Biochemical and molecular characterization of Bacillus pumilus isolated from coastal environment in Cochin, India. Braz. J. Microbiol. 2009, 40, 269–275. Available online: https://www.ncbi.nlm.nih.gov/pubmed/24031357 (accessed on 1 December 2022). [CrossRef]
- Agata, N.; Ohta, M.; Mori, M. Production of an emetic toxin, Cereuide, is associated with a specific class of Bacillus cereus. Curr. Microbiol. 1996, 33, 67–69. [Google Scholar] [CrossRef] [PubMed]
- Hoton, F.M.; Andrup, L.; Swiecicka, I.; Mahillon, J. The cereulide genetic determinants of emetic Bacillus cereus are plasmid-borne. Microbiology 2005, 151, 2121–2124. [Google Scholar] [CrossRef]
- Isobe, M.; Ishikawa, T.; Suwan, S.; Agata, N.; Ohta, M. Synthesis and activity of cereulide, a cyclic dodecadepsipeptide ionophore as emetic toxin from Bacillus cereus. Bioorg. Med. Chem. 1995, 5, 2855–2858. [Google Scholar] [CrossRef]
- Alonzo, D.A.; Magarvey, N.A.; Schmeing, T.M. Characterization of cereulide synthetase, a toxin-producing macromolecular machine. PLoS ONE 2015, 10, e0128569. [Google Scholar] [CrossRef] [PubMed]
- Ramarao, N.; Tran, S.L.; Marin, M.; Vidic, J. Advanced methods for detection of Bacillus cereus and its pathogenic factors. Sensors 2020, 20, 2667. [Google Scholar] [CrossRef] [PubMed]
- Hughes, S.; Bartholomew, B.; Hardy, J.; Kramer, J. Potential application of a HEp-2 cell assay in the investigation of Bacillus cereus emetic-syndrome food poisoning. FEMS Microbiol. Lett. 1988, 52, 7–11. [Google Scholar] [CrossRef]
- Beattie, S.A.; Williams, A. Detection of toxigenic strains of Bacillus cereus and other Bacillus spp. with an improved cytotoxicity assay. Lett. Appl. Microbiol. 1999, 28, 221–225. [Google Scholar] [CrossRef]
- Finlay, W.J.; Logan, N.A.; Sutherland, A.D. Semiautomated metabolic staining assay for Bacillus cereus emetic toxin. Appl. Environ. Microbiol. 1999, 65, 1811–1812. [Google Scholar] [CrossRef] [PubMed]
- Virtanen, S.M.; Roivainen, M.; Andersson, M.A.; Ylipaasto, P.; Hoornstra, D.; Mikkola, R.; Salkinoja-Salonen, M.S. In vitro toxicity of cereulide on porcine pancreatic langerhans islets. Toxicon 2008, 51, 1029–1037. [Google Scholar] [CrossRef]
- Hoornstra, D.; Andersson, M.A.; Teplova, V.V.; Mikkola, R.; Uotila, L.M.; Andersson, L.C.; Roivainen, M.; Gahmberg, C.G.; Salkinoja-Salonen, M.S. Potato crop as a source of emetic Bacillus cereus and cereulide-induced mammalian cell toxicity. Appl. Environ. Microbiol. 2013, 79, 3534–3543. [Google Scholar] [CrossRef]
- Griffiths, R.I.; Whiteley, A.S.; O’Donnell, A.G.; Bailey, M.J. Rapid method for coextraction of DNA and RNA from natural environments for analysis of ribosomal DNA and rRNA-based microbial community composition. Appl. Environ. Microbiol. 2000, 66, 5488–5491. [Google Scholar] [CrossRef]
- Rajoka, M.S.R.; Mehwish, H.M.; Siddiq, M.; Haobin, Z.; Zhu, J.; Yan, L.; Shao, D.; Xu, X.; Shi, J. Identification, characterization, and probiotic potential of Lactobacillus rhamnosus isolated from human milk. LWT-Food Sci. Technol. 2017, 84, 271–280. [Google Scholar] [CrossRef]
- Lane, D.J. 16S/23S rRNA Sequencing. In Nucleic Acid Techniques in Bacterial Systematic; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley and Sons: New York, NY, USA, 1991; pp. 115–175. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Window 95/98/NT. In Nucleic Acids Symposium Series; Oxford University Press: London, UK, 1999; Volume 41, pp. 95–98. [Google Scholar] [CrossRef]
- Francisco, A.P.; Bugalho, M.; Ramirez, M.; Carriço, J.A. Global optimal eBURST analysis of multilocus typing data using a graphic matroid approach. BMC Bioinform. 2009, 10, 152. [Google Scholar] [CrossRef]
- Nascimento, M.; Sousa, A.; Ramirez, M.; Francisco, A.P.; Carriço, J.A.; Vaz, C. PHYLOViZ 2.0: Providing scalable data integration and visualization for multiple phylogenetic inference methods. J. Bioinform. 2017, 33, 128–129. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M.A. Simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Ausubel, F.M.; Brent, R.; Kingston, R.E.; Moore, D.D.; Seidman, J.G.; Smith, J.A.; Struhl, K. Purification and concentration of DNA from aqueous solution. In Short Protocols in Molecular Biology; John Wiley & Sons, Inc.: New York, NY, USA, 1995. [Google Scholar]
- Yang, I.C.; Shih, D.Y.C.; Huang, T.P.; Huang, Y.P.; Wang, J.Y.; Pan, T.M. Establishment of a novel multiplex PCR assay and detection of toxigenic strains of the species in the Bacillus cereus group. J. Food Prot. 2005, 68, 2123–2130. [Google Scholar] [CrossRef]
- Xu, M.; McCanna, D.J.; Sivak, J.G. Use of the viability reagent PrestoBlue in comparison with alamarBlue and MTT to assess the viability of human corneal epithelial cells. J. Pharmacol. Toxicol. Methods 2015, 71, 1–7. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Accession No. (FC Isolate) | Closest Relative | Accession No. (Closest Relative) | Percent Identity | Percent Coverage |
|---|---|---|---|---|---|
| FC1 | ON351569 | Bacillus thuringiensis HER1410 | CP050183.1 | 100.00 | 93 |
| FC2 | ON351570 | Bacillus thuringiensis GCU1 | MN590524.1 | 99.93 | 97 |
| Bacillus thuringiensis 41(7) il | KY323326.1 | 100.00 | 97 | ||
| FC6 | ON351571 | Bacillus thuringiensis HER1410 | CP050183.1 | 99.93 | 99 |
| FC7 | ON351572 | Bacillus thuringiensis HER1410 | CP050183.1 | 100.00 | 98 |
| FC8 | ON351573 | Bacillus thuringiensis HER1410 | CP050183.1 | 100.00 | 99 |
| FC9 | ON351574 | Bacillus thuringiensis HER1410 | CP050183.1 | 100.00 | 97 |
| FC10 | ON351575 | Bacillus thuringiensis LAA3 | OM585510.1 | 100.00 | 95 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pheepakpraw, J.; Kaewkod, T.; Konkit, M.; Krongdang, S.; Jantakee, K.; Praphruet, R.; Bovonsombut, S.; Panya, A.; Tragoolpua, Y.; Logan, N.A.; et al. Intraspecific Diversity and Pathogenicity of Bacillus thuringiensis Isolates from an Emetic Illness. Toxins 2023, 15, 89. https://doi.org/10.3390/toxins15020089
Pheepakpraw J, Kaewkod T, Konkit M, Krongdang S, Jantakee K, Praphruet R, Bovonsombut S, Panya A, Tragoolpua Y, Logan NA, et al. Intraspecific Diversity and Pathogenicity of Bacillus thuringiensis Isolates from an Emetic Illness. Toxins. 2023; 15(2):89. https://doi.org/10.3390/toxins15020089
Chicago/Turabian StylePheepakpraw, Jintana, Thida Kaewkod, Maytiya Konkit, Sasiprapa Krongdang, Kanyaluck Jantakee, Rueankaew Praphruet, Sakunnee Bovonsombut, Aussara Panya, Yingmanee Tragoolpua, Niall A. Logan, and et al. 2023. "Intraspecific Diversity and Pathogenicity of Bacillus thuringiensis Isolates from an Emetic Illness" Toxins 15, no. 2: 89. https://doi.org/10.3390/toxins15020089
APA StylePheepakpraw, J., Kaewkod, T., Konkit, M., Krongdang, S., Jantakee, K., Praphruet, R., Bovonsombut, S., Panya, A., Tragoolpua, Y., Logan, N. A., & Chitov, T. (2023). Intraspecific Diversity and Pathogenicity of Bacillus thuringiensis Isolates from an Emetic Illness. Toxins, 15(2), 89. https://doi.org/10.3390/toxins15020089