The Molecular Composition of Peptide Toxins in the Venom of Spider Lycosa coelestis as Revealed by cDNA Library and Transcriptomic Sequencing

 and
and 
{kind=link}
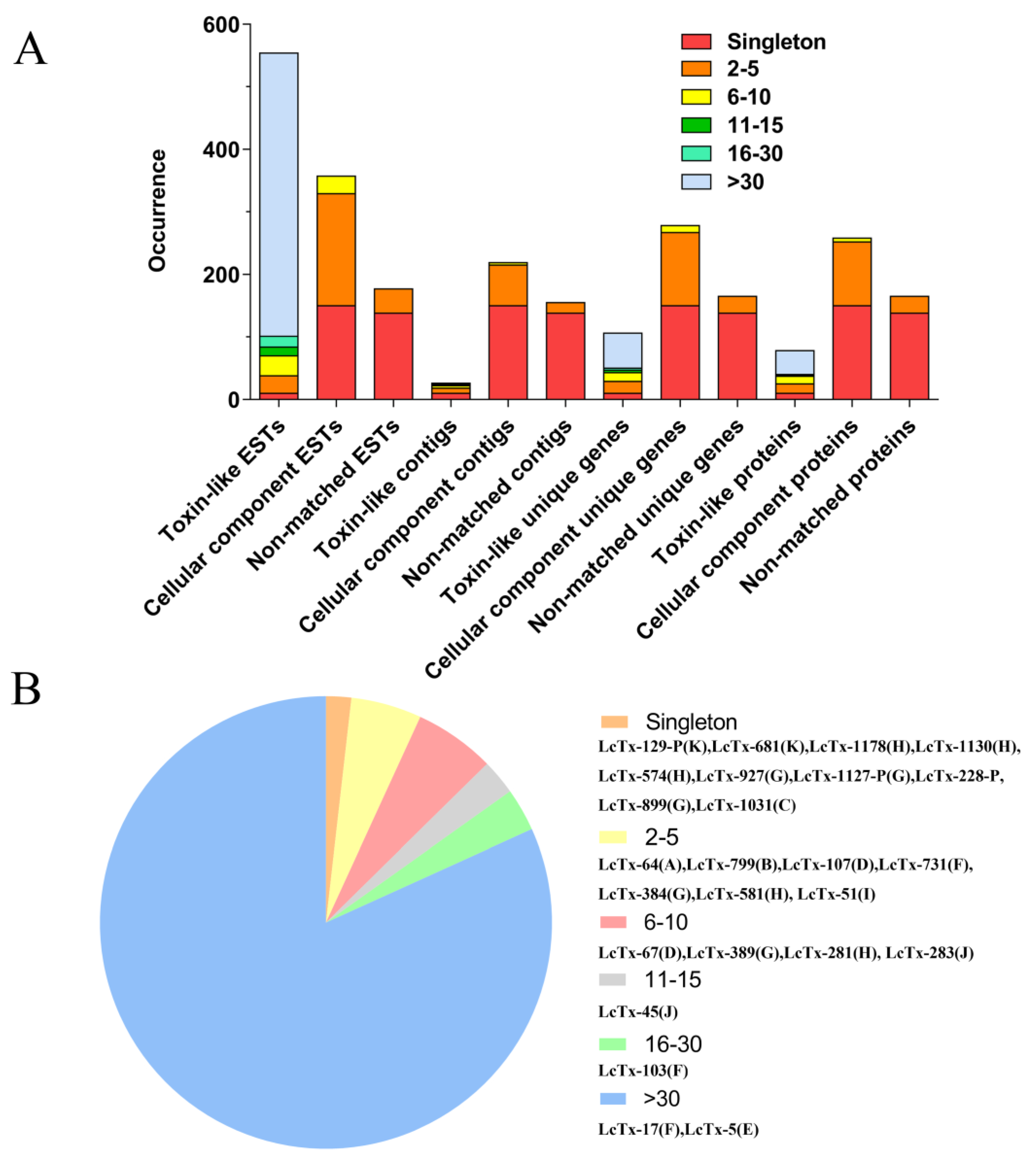{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results and Discussion
2.1. General Features and Annotation of L. coelestis Venom Gland ESTs
2.2. Cluster Analysis of L. coelestis Venom Gland ESTs
2.3. Family Analysis of Putative Toxin Precursors in L. coelestis Venom Gland
2.3.1. Family A
2.3.2. Family B
2.3.3. Family C
2.3.4. Family D
2.3.5. Family E
2.3.6. Family F
2.3.7. Family G
2.3.8. Family H
2.3.9. Family I
2.3.10. Family J
2.3.11. Family K
2.4. RP-HPLC Purification of L. coelestis Venom and Off-Line MALDI-TOF Mass Spectrometry Analysis of the Eluted Fractions
2.5. Activity of L. coelestis Venom against the Voltage-Gated Na+, Ca2+ and K+ Channels in DRG Neurons
3. Conclusions
4. Materials and Methods
4.1. cDNA Library and Transcriptomic Sequencing of the Venom Gland from the Spider L. coelestis
4.2. EST Translation and Annotation
4.3. Cluster Analysis of ESTs and Family Classification of the Toxin-Like Peptides
4.4. Dorsal Root Ganglion (DRG) Neuron Preparation
4.5. RP-HPLC Purification of L. coelestis Venom and MALDI-TOF Analysis of the Eluted Fractions
4.6. Electrophysiology
4.7. Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Casewell, N.R.; Wuster, W.; Vonk, F.J.; Harrison, R.A.; Fry, B.G. Complex cocktails: The evolutionary novelty of venoms. Trends Ecol. Evol. 2013, 28, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.R.; Rikli, H.G. Aspartic Acid Isomerization Characterized by High Definition Mass Spectrometry Significantly Alters the Bioactivity of a Novel Toxin from Poecilotheria. Toxins 2020, 12, 207. [Google Scholar] [CrossRef] [PubMed]
- Hillyard, P. The Book of the Spider: From Arachnophobia to the Love of Spiders; Random House: New York, NY, USA, 1994. [Google Scholar]
- Escoubas, P.; Diochot, S.; Corzo, G. Structure and pharmacology of spider venom neurotoxins. Biochimie 2000, 82, 893–907. [Google Scholar] [CrossRef]
- King, G.F. Venoms as a platform for human drugs: Translating toxins into therapeutics. Expert Opin. Biol. Ther. 2011, 11, 1469–1484. [Google Scholar] [CrossRef]
- Luo, J.; Ding, Y.; Peng, Z.; Chen, K.; Zhang, X.; Xiao, T.; Chen, J. Molecular diversity and evolutionary trends of cysteine-rich peptides from the venom glands of Chinese spider Heteropoda venatoria. Sci. Rep. 2021, 11, 3211. [Google Scholar] [CrossRef] [PubMed]
- Pallaghy, P.K.; Norton, R.S.; Nielsen, K.J.; Craik, D.J. A common structural motif incorporating a cystine knot and a triple-stranded beta-sheet in toxic and inhibitory polypeptides. Protein Sci. 1994, 3, 1833–1839. [Google Scholar] [CrossRef] [PubMed]
- Shu, Q.; Lu, S.Y.; Gu, X.C.; Liang, S.P. The structure of spider toxin huwentoxin-II with unique disulfide linkage: Evidence for structural evolution. Protein Sci. 2002, 11, 245–252. [Google Scholar] [CrossRef]
- Wan, H.; Lee, K.S.; Kim, B.Y.; Zou, F.M.; Yoon, H.J.; Je, Y.H.; Li, J.; Jin, B.R. A Spider-Derived Kunitz-Type Serine Protease Inhibitor That Acts as a Plasmin Inhibitor and an Elastase Inhibitor. PLoS ONE 2013, 8, e53343. [Google Scholar] [CrossRef] [PubMed]
- King, G.F.; Gentz, M.C.; Escoubas, P.; Nicholson, G.M. A rational nomenclature for naming peptide toxins from spiders and other venomous animals. Toxicon 2008, 52, 264–276. [Google Scholar] [CrossRef]
- Klint, J.K.; Senff, S.; Rupasinghe, D.B.; Er, S.Y.; Herzig, V.; Nicholson, G.M.; King, G.F. Spider-venom peptides that target voltage-gated sodium channels: Pharmacological tools and potential therapeutic leads. Toxicon 2012, 60, 478–491. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.; Goldstein, S.A.N. Tethered peptide toxins for ion channels. Methods Enzymol. 2021, 654, 203–224. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.; Kennedy, K.; De Blas, G.A.; Orta, G.; Pavarotti, M.A.; Arias, R.J.; de la Vega-Beltrán, J.L.; Li, Q.; Dai, H.; Perozo, E.; et al. Role of human Hv1 channels in sperm capacitation and white blood cell respiratory burst established by a designed peptide inhibitor. Proc. Natl. Acad. Sci. USA 2018, 115, E11847–E11856. [Google Scholar] [CrossRef] [PubMed]
- Windley, M.J.; Herzig, V.; Dziemborowicz, S.A.; Hardy, M.C.; King, G.F.; Nicholson, G.M. Spider-venom peptides as bioinsecticides. Toxins 2012, 4, 191–227. [Google Scholar] [CrossRef]
- Daniel, J.T.; Clark, R.J. G-Protein Coupled Receptors Targeted by Analgesic Venom Peptides. Toxins 2017, 9, 372. [Google Scholar] [CrossRef] [PubMed]
- Van Baelen, A.C.; Robin, P.; Kessler, P.; Maïga, A.; Gilles, N.; Servent, D. Structural and Functional Diversity of Animal Toxins Interacting With GPCRs. Front. Mol. Biosci. 2022, 9, 811365. [Google Scholar] [CrossRef]
- Dutertre, S.; Lewis, R.J. Use of venom peptides to probe ion channel structure and function. J. Biol. Chem. 2010, 285, 13315–13320. [Google Scholar] [CrossRef] [PubMed]
- García-Fernandez, R.; Peigneur, S.; Pons, T.; Alvarez, C.; González, L.; Chávez, M.; Tytgat, J. The Kunitz-Type Protein ShPI-1 Inhibits Serine Proteases and Voltage-Gated Potassium Channels. Toxins 2016, 8, 110. [Google Scholar] [CrossRef]
- Jiang, L.; Deng, M.; Duan, Z.; Tang, X.; Liang, S. Molecular cloning, bioinformatics analysis and functional characterization of HWTX-XI toxin superfamily from the spider Ornithoctonus huwena. Peptides 2014, 54, 9–18. [Google Scholar] [CrossRef]
- Saez, N.J.; Senff, S.; Jensen, J.E.; Er, S.Y.; Herzig, V.; Rash, L.D.; King, G.F. Spider-Venom Peptides as Therapeutics. Toxins 2010, 2, 2851–2871. [Google Scholar] [CrossRef]
- Tang, C.; Zhou, X.; Nguyen, P.T.; Zhang, Y.; Hu, Z.; Zhang, C.; Yarov-Yarovoy, V.; DeCaen, P.G.; Liang, S.; Liu, Z. A novel tarantula toxin stabilizes the deactivated voltage sensor of bacterial sodium channel. FASEB J. 2017, 31, 3167–3178. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.; Zhou, X.; Zhang, Y.; Xiao, Z.; Hu, Z.; Zhang, C.; Huang, Y.; Chen, B.; Liu, Z.; Liang, S. Synergetic action of domain II and IV underlies persistent current generation in Nav1.3 as revealed by a tarantula toxin. Sci. Rep. 2015, 5, 9241. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Zhang, Y.; Hu, W.; Xu, D.; Tao, H.; Yang, X.; Li, Y.; Jiang, L.; Liang, S. Molecular diversification of peptide toxins from the tarantula Haplopelma hainanum (Ornithoctonus hainana) venom based on transcriptomic, peptidomic, and genomic analyses. J. Proteome Res. 2010, 9, 2550–2564. [Google Scholar] [CrossRef] [PubMed]
- Kuhn-Nentwig, L.; Langenegger, N.; Heller, M.; Koua, D.; Nentwig, W. The Dual Prey-Inactivation Strategy of Spiders—In-Depth Venomic Analysis of Cupiennius salei. Toxins 2019, 11, 167. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, J.; Tang, X.; Wang, F.; Jiang, L.; Xiong, X.; Wang, M.; Rong, M.; Liu, Z.; Liang, S. Transcriptome analysis of the venom glands of the Chinese wolf spider Lycosa singoriensis. Zoology 2009, 113, 10–18. [Google Scholar] [CrossRef]
- Fisyunov, A.; Pluzhnikov, K.; Molyavka, A.; Grishin, E.; Lozovaya, N.; Krishtal, O. Novel spider toxin slows down the activation kinetics of P-type Ca2+ channels in Purkinje neurons of rat. Toxicology 2005, 207, 129–136. [Google Scholar] [CrossRef]
- Norton, R.S.; Pallaghy, P.K. The cystine knot structure of ion channel toxins and related polypeptides. Toxicon 1998, 36, 1573–1583. [Google Scholar] [CrossRef]
- Wong, E.S.; Hardy, M.C.; Wood, D.; Bailey, T.; King, G.F. SVM-based prediction of propeptide cleavage sites in spider toxins identifies toxin innovation in an Australian tarantula. PLoS ONE 2013, 8, e66279. [Google Scholar] [CrossRef]
- Kozlov, S.; Malyavka, A.; McCutchen, B.; Lu, A.; Schepers, E.; Herrmann, R.; Grishin, E. A novel strategy for the identification of toxinlike structures in spider venom. Proteins Struct. Funct. Bioinform. 2005, 59, 131–140. [Google Scholar] [CrossRef]
- Cardoso, F.; Pacífico, L.; Carvalho, D.; Victória, J.; Neves, A.; Chávez-Olórtegui, C.; Gomez, M.; Kalapothakis, E. Molecular cloning and characterization of Phoneutria nigriventer toxins active on calcium channels. Toxicon 2003, 41, 755–763. [Google Scholar] [CrossRef]
- Touchard, A.; Dauvois, M.; Arguel, M.J.; Petitclerc, F.; Leblanc, M.; Dejean, A.; Orivel, J.; Nicholson, G.M.; Escoubas, P. Elucidation of the unexplored biodiversity of ant venom peptidomes via MALDI–TOF mass spectrometry and its application for chemotaxonomy. J. Proteom. 2014, 105, 217–231. [Google Scholar] [CrossRef] [PubMed]
- Schaffrath, S.; Predel, R. A simple protocol for venom peptide barcoding in scorpions. EuPA Open Proteom. 2014, 3, 239–245. [Google Scholar] [CrossRef]
- Escoubas, P.; Quinton, L.; Nicholson, G.M. Venomics: Unravelling the complexity of animal venoms with mass spectrometry. J. Mass Spectrom. 2008, 43, 279–295. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Wu, X.; Zhang, P.; Duan, Z.; Zhou, X.; Chen, M.; Farooq, A.; Liang, S.; Liu, Z. Peptide-rich venom from the spider Heteropoda venatoria potently inhibits insect voltage-gated sodium channels. Toxicon 2017, 125, 44–49. [Google Scholar] [CrossRef]
- Hu, Z.; Zhou, X.; Chen, J.; Tang, C.; Xiao, Z.; Ying, D.; Liu, Z.; Liang, S. The venom of the spider Selenocosmia jiafu contains various neurotoxins acting on voltage-gated ion channels in rat dorsal root ganglion neurons. Toxins 2014, 6, 988–1001. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, F.; Li, D.; Xu, S.; He, J.; Yu, H.; Li, J.; Liu, Z.; Liang, S. The venom of the fishing spider Dolomedes sulfurous contains various neurotoxins acting on voltage-activated ion channels in rat dorsal root ganglion neurons. Toxicon 2013, 65, 68–75. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, J.; Liu, Z.; Wang, M.; Wang, J.; Lu, S.; Zhu, L.; Zeng, X.; Liang, S. Effects of the venom of the spider Ornithoctonus hainana on neonatal rat ventricular myocytes cellular and ionic electrophysiology. Toxicon 2014, 87, 104–112. [Google Scholar] [CrossRef]
- Berta, T.; Qadri, Y.; Tan, P.H.; Ji, R.R. Targeting dorsal root ganglia and primary sensory neurons for the treatment of chronic pain. Expert Opin. Ther. Targets 2017, 21, 695–703. [Google Scholar] [CrossRef]
- Saegusa, H.; Kurihara, T.; Zong, S.; Kazuno, A.; Matsuda, Y.; Nonaka, T.; Han, W.; Toriyama, H.; Tanabe, T. Suppression of inflammatory and neuropathic pain symptoms in mice lacking the N-type Ca2+ channel. EMBO J. 2001, 20, 2349–2356. [Google Scholar] [CrossRef]
- McGivern, J.G. Targeting N-type and T-type calcium channels for the treatment of pain. Drug Discov. Today 2006, 11, 245–253. [Google Scholar] [CrossRef]
- Lewis, R.J.; Garcia, M.L. Therapeutic potential of venom peptides. Nat. Rev. Drug Discov. 2003, 2, 790–802. [Google Scholar] [CrossRef] [PubMed]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M.; et al. De novo transcript sequence reconstruction from RNA-seq using the Trinity platform for reference generation and analysis. Nat. Protoc. 2013, 8, 1494–1512. [Google Scholar] [CrossRef] [PubMed]
- Clewley, J.P. Macintosh sequence analysis software. DNAStar’s LaserGene. Mol. Biotechnol. 1995, 3, 221–224. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Francois-Moutal, L.; Wang, Y.; Moutal, A.; Cottier, K.E.; Melemedjian, O.K.; Yang, X.; Wang, Y.; Ju, W.; Largent-Milnes, T.M.; Khanna, M.; et al. A membrane-delimited N-myristoylated CRMP2 peptide aptamer inhibits CaV2.2 trafficking and reverses inflammatory and postoperative pain behaviors. Pain 2015, 156, 1247–1264. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, X.; Chen, Y.; Liu, H.; Kong, X.; Liang, X.; Zhang, Y.; Tang, C.; Liu, Z. The Molecular Composition of Peptide Toxins in the Venom of Spider Lycosa coelestis as Revealed by cDNA Library and Transcriptomic Sequencing. Toxins 2023, 15, 143. https://doi.org/10.3390/toxins15020143
Wu X, Chen Y, Liu H, Kong X, Liang X, Zhang Y, Tang C, Liu Z. The Molecular Composition of Peptide Toxins in the Venom of Spider Lycosa coelestis as Revealed by cDNA Library and Transcriptomic Sequencing. Toxins. 2023; 15(2):143. https://doi.org/10.3390/toxins15020143
Chicago/Turabian StyleWu, Xiangyue, Yan Chen, Hao Liu, Xiangjin Kong, Xinyao Liang, Yu Zhang, Cheng Tang, and Zhonghua Liu. 2023. "The Molecular Composition of Peptide Toxins in the Venom of Spider Lycosa coelestis as Revealed by cDNA Library and Transcriptomic Sequencing" Toxins 15, no. 2: 143. https://doi.org/10.3390/toxins15020143
APA StyleWu, X., Chen, Y., Liu, H., Kong, X., Liang, X., Zhang, Y., Tang, C., & Liu, Z. (2023). The Molecular Composition of Peptide Toxins in the Venom of Spider Lycosa coelestis as Revealed by cDNA Library and Transcriptomic Sequencing. Toxins, 15(2), 143. https://doi.org/10.3390/toxins15020143