The Sea Anemone Neurotoxins Modulating Sodium Channels: An Insight at Structure and Functional Activity after Four Decades of Investigation


Abstract
1. Introduction
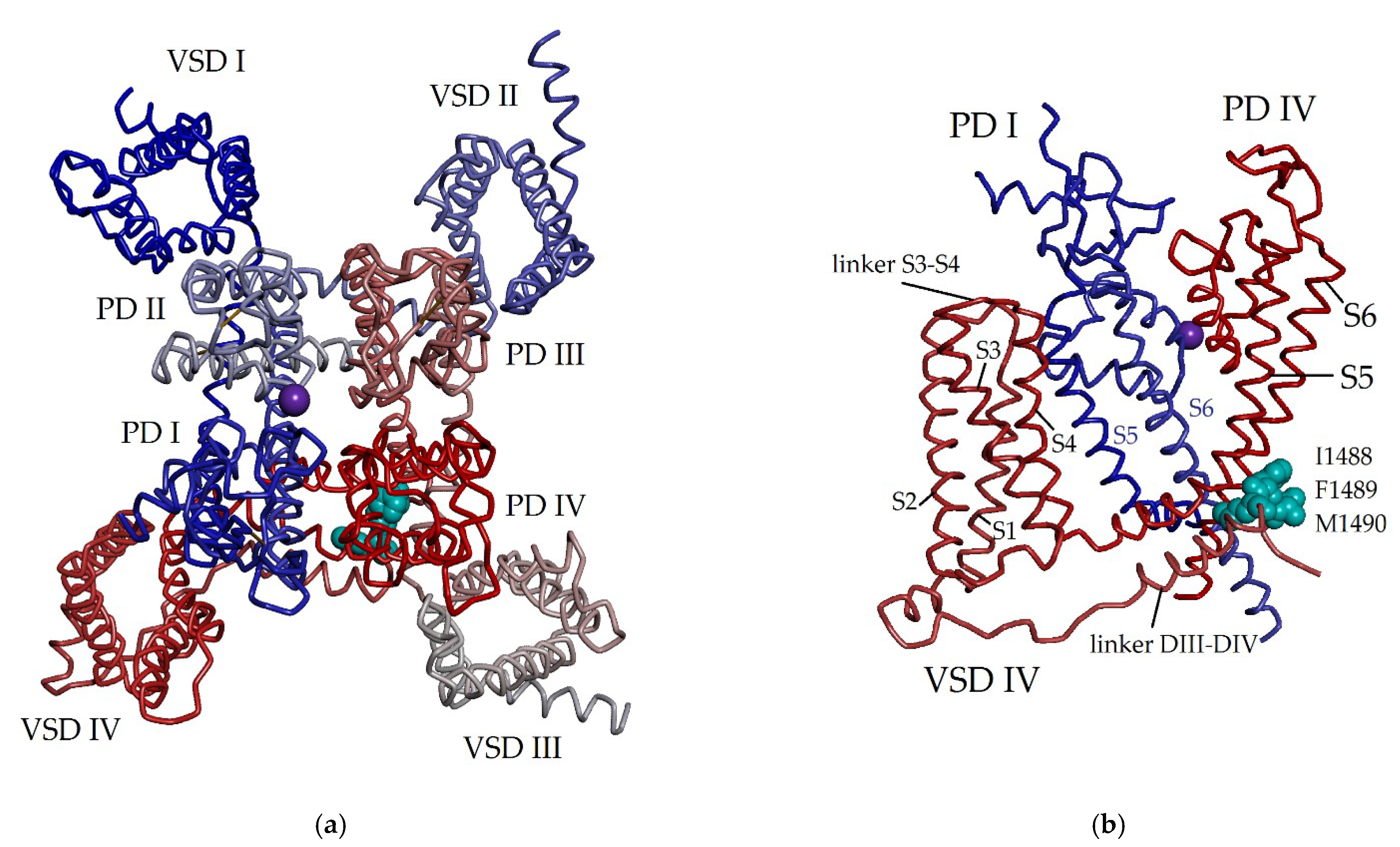
2. Voltage-Gated Sodium Channels as Main Targets of Sea Anemone Neurotoxins
3. Structure-Functional Characteristics of Sea Anemone NaTxs
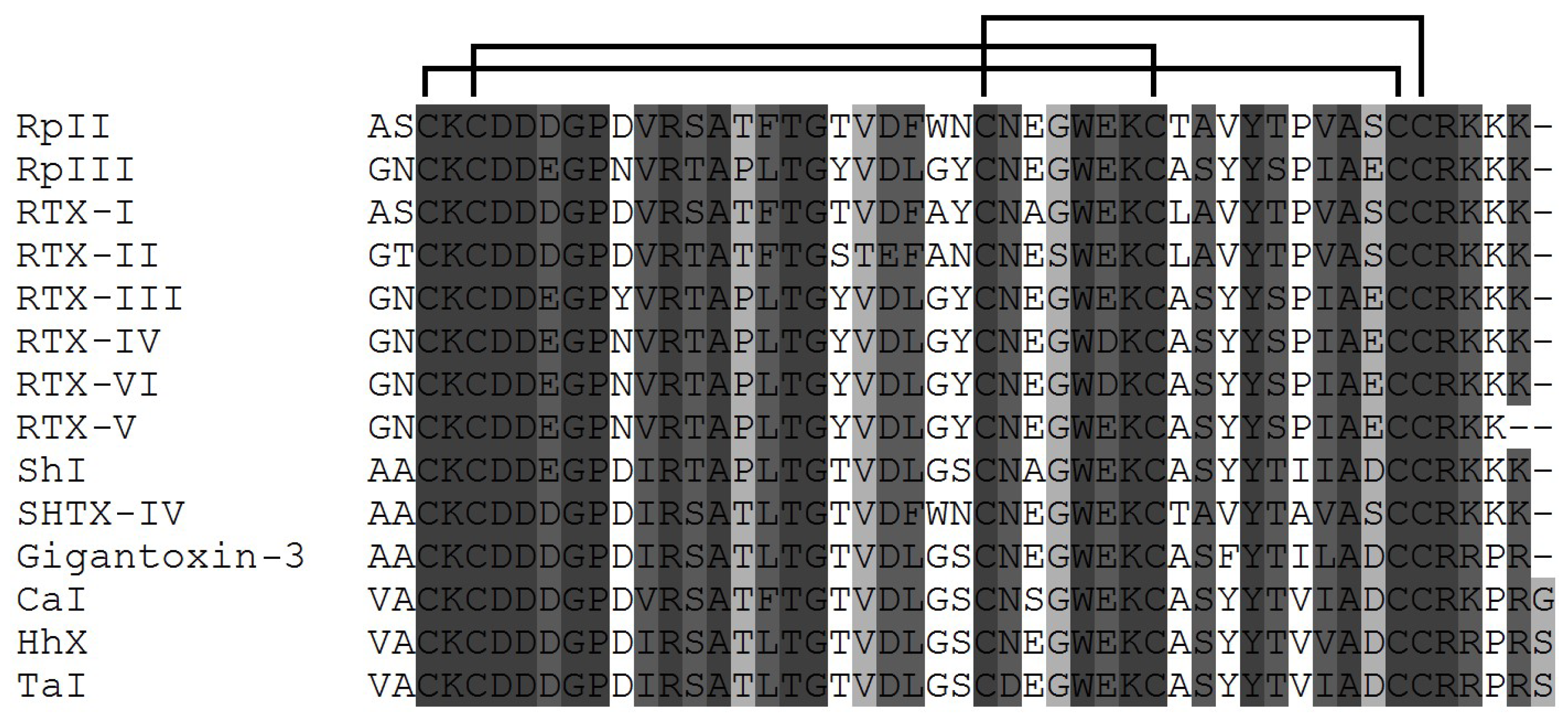
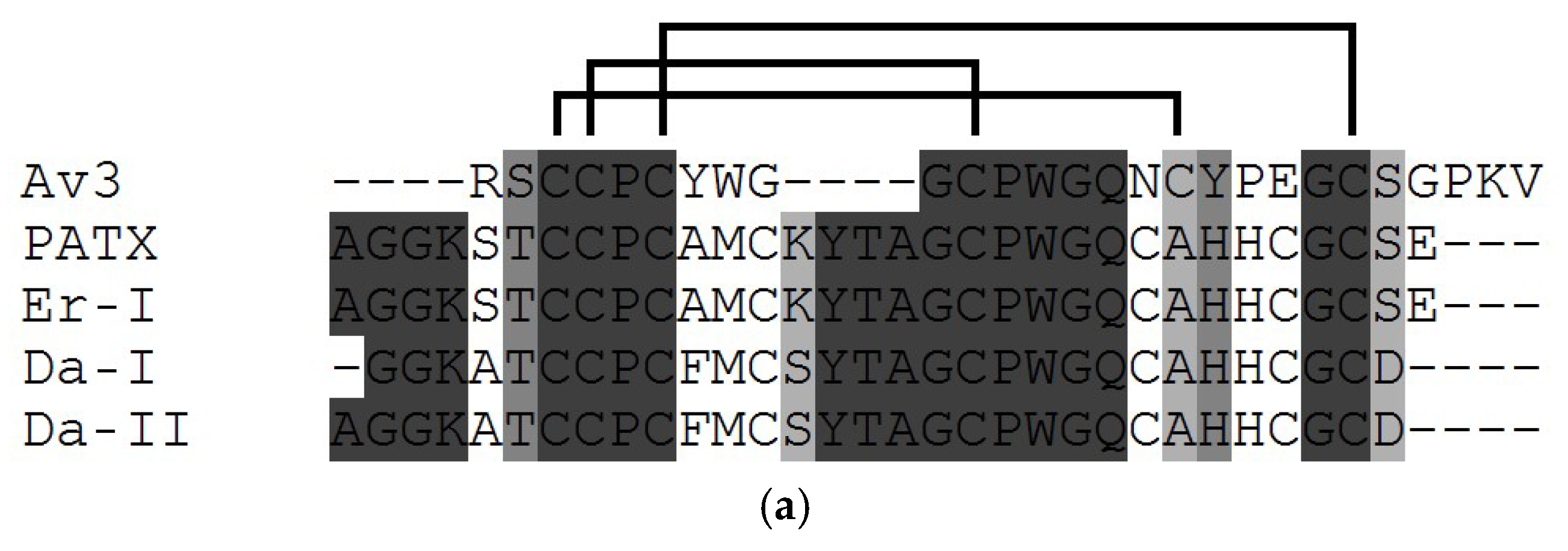
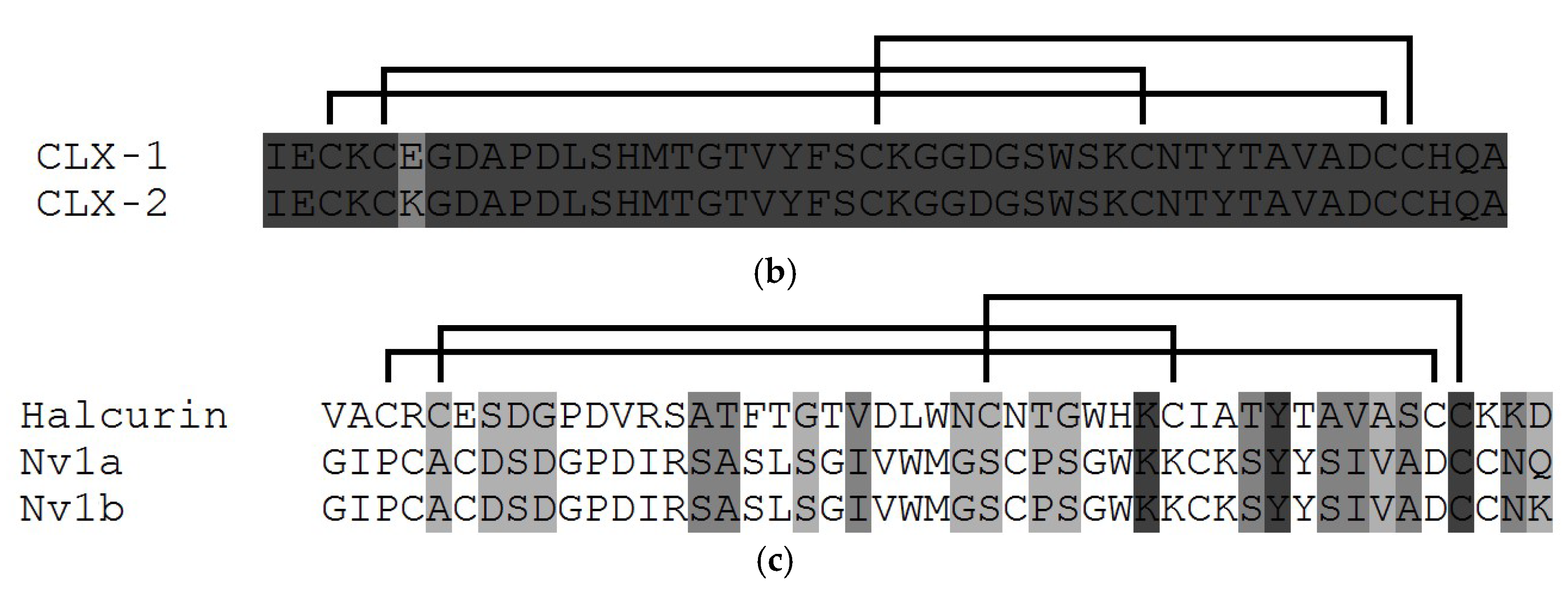
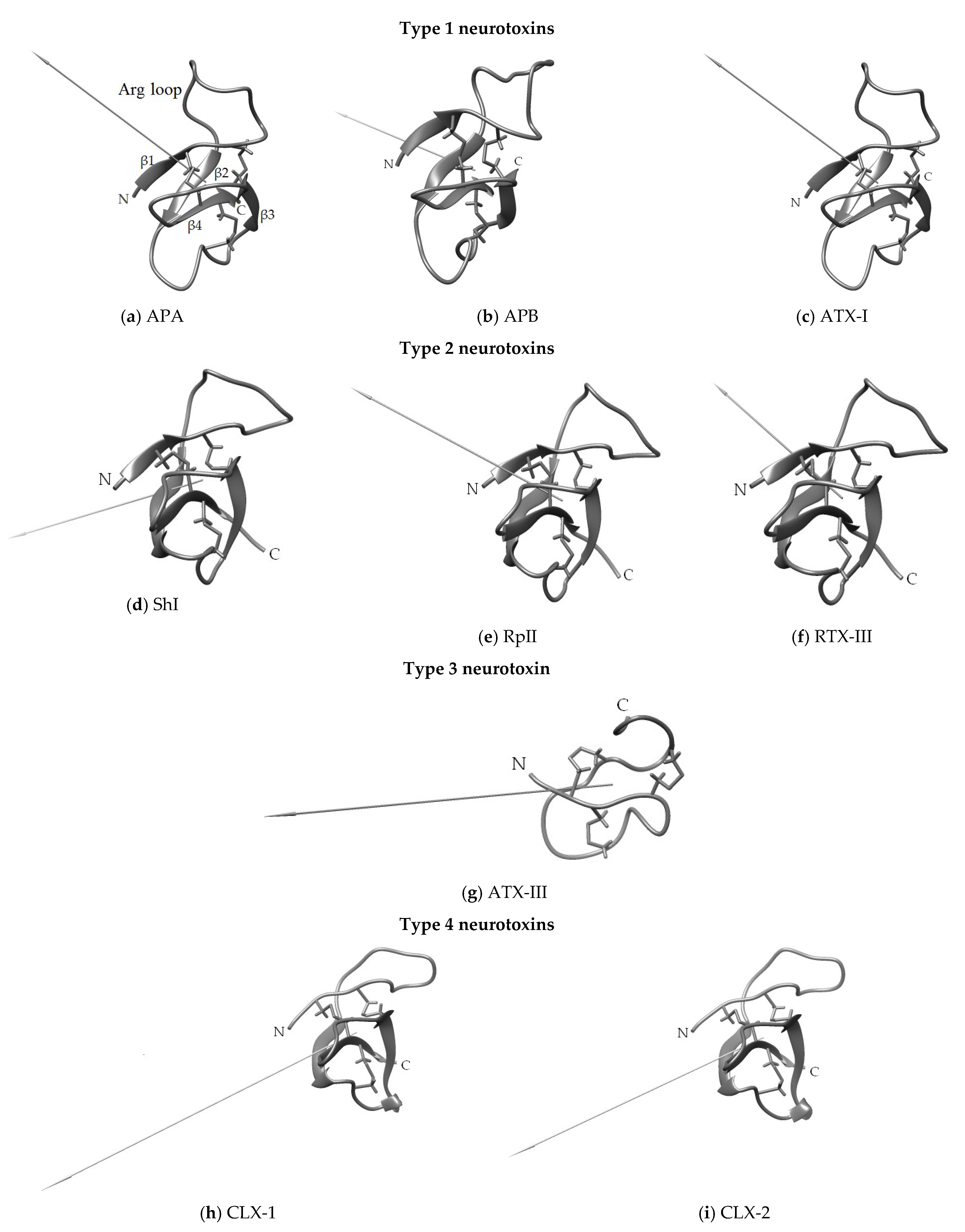
3.1. Structural Types of NaTxs





3.2. Secondary and Spatial Structures of NaTxs

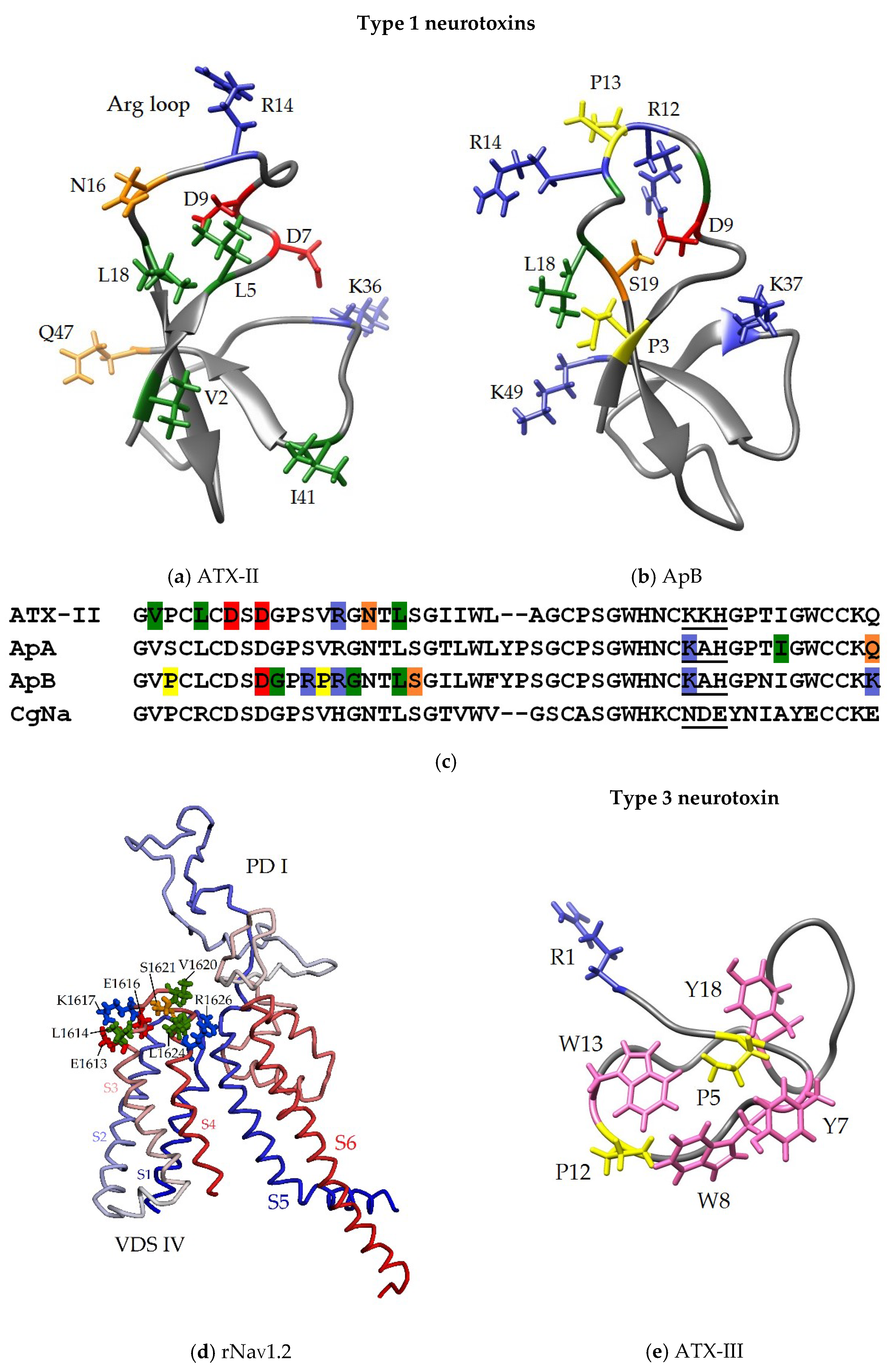
3.3. Some Key Functionally Significant Amino Acid Residues of NaTxs

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
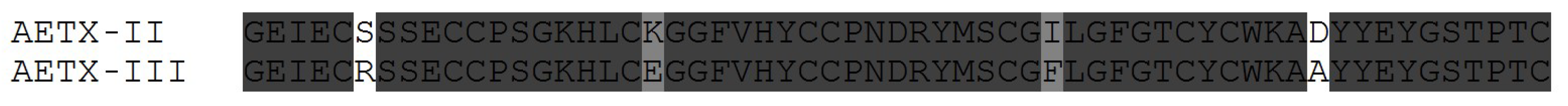{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| See Anemone Species | Neurotoxin | Lethal Dose, (μg/kg) a | Binding to Synaptosomes of Rat Brain, KD (μM) b | Ref. | |
|---|---|---|---|---|---|
| LD50 (Mice) | LD100 (Crabs) | ||||
| A. sulcata | ATX-I ATX-II ATX-III ATX-V | 4000 100 18,000 19.0 | 4.4 3.7 6.7 10.4 | 7.0 0.15 10.0 0.05 | [21,23,26,97,153] |
| A. xanthogrammica | ApA ApB | 66.0174 8.0 | 22.0 78.0 | 0.12 0.035 | [30] [25,26] |
| S. gigantea | Gigantoxin II | 2000 | 14.0 | 10.0 | [26,103] |
| R. paumotensis (=H. magnífica) | RpI RpII RpIII RpIV | 145 4200 53.0 40.0 | 36.0 15.0 10.0 90.0 | 0.9 10.0 0.3 10.0 | [27] [27] [91] [27] |
| S. helianthus | ShI | 20,000 | 0.6 | [31,118] | |
| P. actinostoloides | PaTX | 20,000 | 10.0 | [126,130,134] | |
| H. crispa | RTX-I RTX-II RTX-III RTX-IV RTX-V δ-SHTX-Hcr1f | 3000 1650 25.0 40.0 350 4200 | 3.5 4.0 82.0 4.4 12.0 15.0 | [28] [115] [29] [116] [116] [168] | |

3.4. Toxicity of NaTxs
3.5. Evolution of Sea Anemone Neurotoxins
4. Neurotoxins from Sea Anemone H. crispa (=R. macrodactylus)
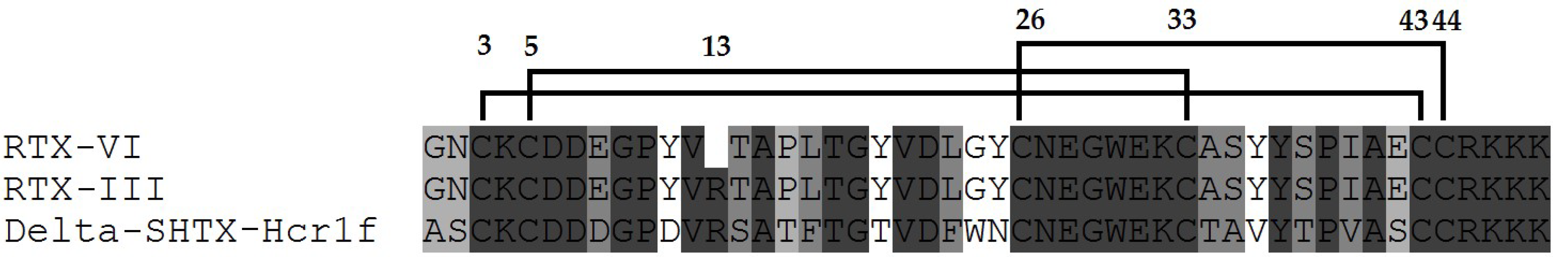
4.1. Isolation, Modifications, Amino Asid Sequences
4.2. New Double-Stranded Type 2 Heteractis Neurotoxin
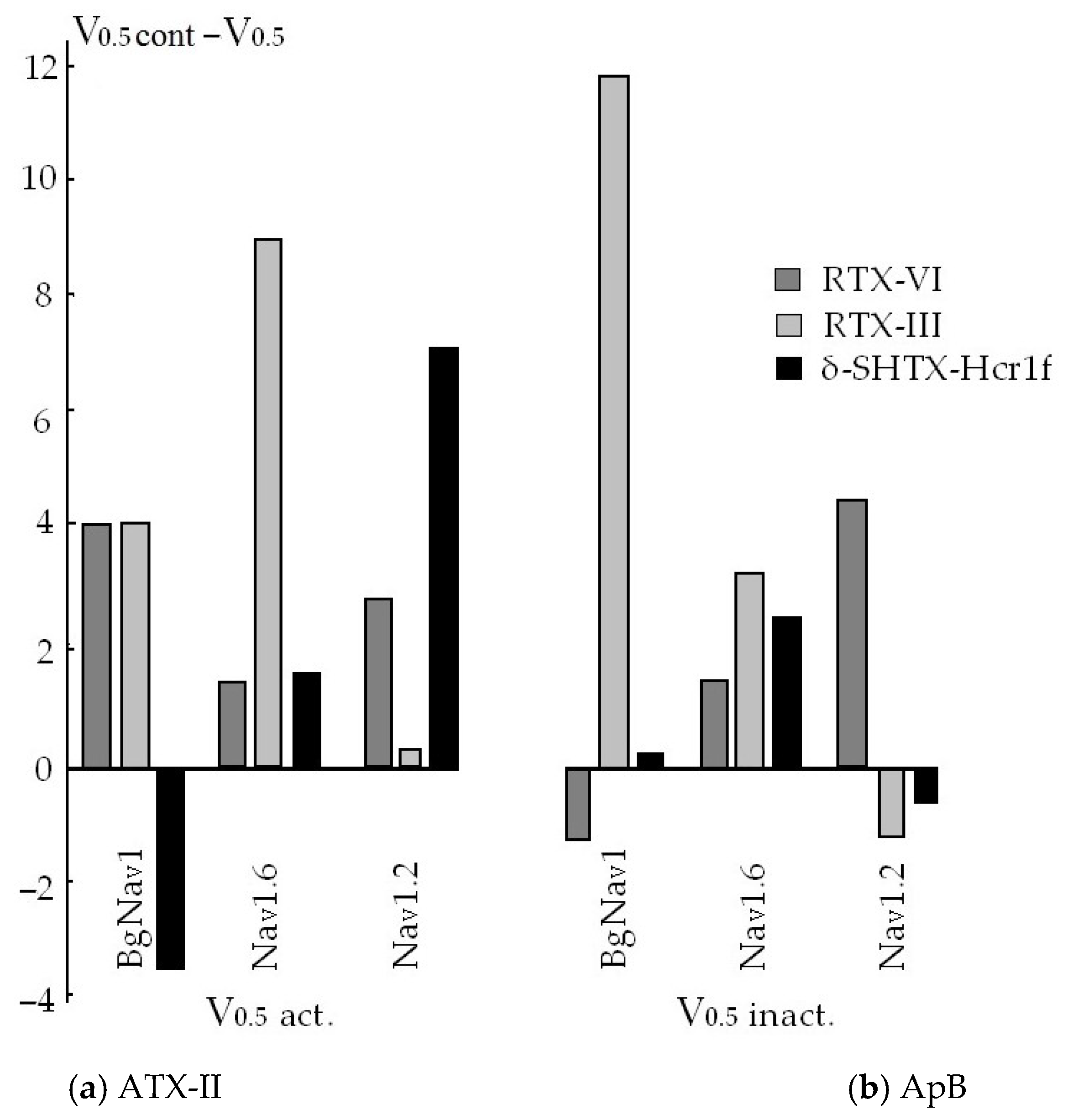
5. Interaction of NaTxs with NaV Subtypes Tested Electrophysiologically and by Molecular Docking
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pennington, M.W.; Czerwinski, A.; Norton, R.S. Peptide therapeutics from venom: Current status and potential. Bioorg. Med. Chem. 2018, 26, 2738–2758. [Google Scholar] [CrossRef]
- Honma, T.; Shiomi, K. Peptide toxins in sea anemones: Structural and functional aspects. Mar. Biotechnol. 2006, 8, 1–10. [Google Scholar] [CrossRef]
- Shiomi, K. Novel peptide toxins recently isolated from sea anemones. Toxicon 2009, 54, 1112–1118. [Google Scholar] [CrossRef]
- Norton, R.S. Sea anemone venom peptides. In Handbook of Biologically Active Peptides, 2nd ed.; Kastin, A.J., Ed.; Elsevier: San Diego, CA, USA, 2013; pp. 430–436. [Google Scholar] [CrossRef]
- Ashwood, L.M.; Norton, R.S.; Undheim, E.A.B.; Hurwood, D.A.; Prentis, P.J. Characterizing functional venom profiles of Anthozoans and Medusozoans within their ecological context. Mar. Drugs 2020, 18, 202. [Google Scholar] [CrossRef]
- Rocha, J.; Peixe, L.; Gomes, N.C.M. Cnidarians as a source of new marine bioactive compounds—An overview of the last decade and future steps for bioprospecting. Mar. Drugs 2011, 9, 1860–1886. [Google Scholar] [CrossRef]
- Daly, M. Functional and genetic diversity of toxins in sea anemones. In Evolution of Venomous Animals and Their Toxins; Gopalakrishnakone, P., Malhotra, A., Eds.; Springer: Dordrecht, The Netherlands, 2017; pp. 87–104. [Google Scholar]
- Surm, J.M.; Moran, Y. Insights into how development and life-history dynamics shape the evolution of venom. EvoDevo 2021, 12, 1. [Google Scholar] [CrossRef]
- Van-Praët, M. Nutrition of sea anemones. Adv. Mar. Biol. 1985, 22, 65–99. [Google Scholar] [CrossRef]
- Karabulut, A.; McClain, M.; Rubinstein, B.; Sabin, K.Z.; McKinney, S.A.; Gibson, M.C. The architecture and operating mechanism of a cnidarian stinging organelle. Nat. Commun. 2022, 13, 3494. [Google Scholar] [CrossRef]
- Wexler, P.; Fonger, G.C.; White, J.; Weinstein, S. Toxinology: Taxonomy, interpretation, and information resources. Sci. Technol. Libr. 2015, 34, 67–90. [Google Scholar] [CrossRef]
- Zenkert, C.; Takahashi, T.; Diesner, M.-O.; Özbek, S. Morphological and Molecular Analysis of the Nematostella vectensis Cnidom. PLoS ONE 2011, 6, e22725. [Google Scholar] [CrossRef]
- Moran, Y.; Genikhovich, G.; Gordon, D.; Wienkoop, S.; Zenkert, C.; Ozbek, S.; Technau, U.; Gurevitz, M. Neurotoxin localization to ectodermal gland cells uncovers an alternative mechanism of venom delivery in sea anemones. Proc. Biol. Sci. 2012, 279, 1351–1358. [Google Scholar] [CrossRef] [PubMed]
- Minagawa, S.; Sugiyama, M.; Ishida, M.; Nagashima, Y.; Shiomi, K. Kunitz-type protease inhibitors from acrorhagi of three species of sea anemones. Comp. Biochem. Physiol. B. Biochem. Mol. Biol. 2008, 150, 240–245. [Google Scholar] [CrossRef] [PubMed]
- Macrander, J.; Brugler, M.R.; Daly, M. A RNA-seq approach to identify putative toxins from acrorhagi in aggressive and non-aggressive Anthopleura elegantissima polyps. BMC Genom. 2015, 16, 221. [Google Scholar] [CrossRef]
- Macrander, J.; Broe, M.; Daly, M. Tissue-specific venom composition and differential gene expression in sea anemones. Genome Biol. Evol. 2016, 8, 2358–2375. [Google Scholar] [CrossRef] [PubMed]
- Sachkova, M.Y.; Singer, S.A.; Macrander, J.; Reitzel, A.M.; Peigneur, S.; Tytgat, J.; Moran, Y. The birth and death of toxins with distinct functions: A case study in the sea anemone Nematostella. Mol. Biol. Evol. 2019, 36, 2001–2012. [Google Scholar] [CrossRef] [PubMed]
- Jouiaei, M.; Sunagar, K.; Gross, A.F.; Scheib, H.; Alewood, P.F.; Moran, Y.; Fry, B.G. Evolution of an ancient venom: Recognition of a novel family of Cnidarian toxins and the common evolutionary origin of sodium and potassium neurotoxins in sea anemone. Mol. Biol. Evol. 2015, 32, 1598–1610. [Google Scholar] [CrossRef] [PubMed]
- Ashwood, L.M.; Mitchell, M.L.; Madio, B.; Hurwood, D.A.; King, G.F.; Undheim, E.A.B.; Norton, R.S.; Prentis, P.J. Tentacle morphological variation coincides with differential expression of toxins in sea anemones. Toxins 2021, 13, 452. [Google Scholar] [CrossRef]
- Prentis, P.J.; Pavasovic, A.; Norton, R.S. Sea anemones: Quiet achievers in the field of peptide toxins. Toxins 2018, 10, 36. [Google Scholar] [CrossRef]
- Béress, L.; Béress, R.; Wunderer, G. Isolation and characterization of three polypeptides with neurotoxicity from Anemonia sulcata. FEBS Lett. 1975, 50, 311–314. [Google Scholar] [CrossRef]
- Béress, L. Biologically active compounds from coelenterates. Pure Appl. Chem. 1982, 54, 1981–1994. [Google Scholar] [CrossRef]
- Wunderer, G.; Fritz, H.; Wachter, E.; Machleidt, W. Amino-acid sequence of a coelenterate toxin: Toxin II from Anemonia sulcata. Eur. J. Biochem. 1976, 68, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Catterall, W.A.; Béress, L. Sea anemone toxin and scorpion toxin share a common receptor site associated with the action potential sodium ionophore. J. Biol. Chem. 1978, 253, 7393–7396. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Hainu, M.; Yasunobu, K.T.; Norton, T.R. Amino acid sequence of the Anthopleura xanthogrammica heart stimulant anthopleurin-A. Biochemistry 1977, 16, 204–208. [Google Scholar] [CrossRef]
- Schweitz, H.; Vincent, J.P.; Barhanin, J.; Frelin, C.; Linden, G.; Hugues, M.; Lazdunski, M. Purification and pharmacological properties of eight sea anemone toxins from Anemonia sulcata, Anthopleura xanthogrammica, Stoichactis giganteus, and Actinodendron plumosum. Biochemistry 1981, 20, 5245–5252. [Google Scholar] [CrossRef]
- Schweitz, H.; Bidard, J.N.; Frelin, C.; Pauron, D.; Vijverberg, H.P.M.; Mahasneh, D.M.; Lazdunski, M.; Vibois, F.; Tsugita, A. Purification, sequence, and pharmacological properties of sea anemone toxins from Radianthus paumotensis. A new class of sea anemone toxins acting on the sodium channel. Biochemistry 1985, 24, 3554–3561. [Google Scholar] [CrossRef]
- Zykova, T.A.; Kozlovskaya, E.P. Amino acid sequence of a neurotoxin from the sea anemone Radianthus macrodactylus. Bioorg. Khim. 1989, 15, 1301–1306. [Google Scholar]
- Zykova, T.A.; Vinokurov, L.M.; Kozlovskaya, E.P.; Elyakov, G.B. Amino acid sequence of neurotoxin III from the sea anemone Radianthus macrodactylus. Bioorg. Khim. 1985, 11, 302–310. [Google Scholar]
- Reimer, N.S.; Yasunobu, C.L.; Yasunobu, K.T.; Norton, T.R. Amino acid sequence of the Anthopleura xanthogrammica heart stimulant, anthopleurin-B. J. Biol. Chem. 1985, 260, 8690–8693. [Google Scholar] [CrossRef]
- Kem, W.R.; Parten, B.; Pennington, M.W.; Price, D.A.; Dunn, B.M. Isolation, characterization, and amino acid sequence of a polypeptide neurotoxin occurring in the sea anemone Stichodactyla helianthus. Biochemistry 1989, 28, 3483–3489. [Google Scholar] [CrossRef]
- Shiomi, K.; Lin, X.-Y.; Nagashima, Y.; Ishida, M. Isolation and amino acid sequence of polypeptide toxins in the Caribbean sea anemone Condylactis passiflora. Fish Sci. 1995, 61, 1016–1021. [Google Scholar] [CrossRef]
- Lin, X.Y.; Ishida, M.; Nagashima, Y.; Shiomi, K. A polypeptide toxin in the sea anemone Actinia equina homologous with other sea anemone sodium cannel toxins: Isolation and amino acid sequence. Toxicon 1996, 34, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Shiomi, K.; Qian, W.H.; Lin, X.Y.; Shimakura, K.; Nagashima, Y.; Ishida, M. Novel polypeptide toxins with crab lethality from the sea anemone Anemonia erythraea. Biochim. Biophys. Acta 1997, 1335, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Schendel, V.; Rash, D.L.; Jenner, A.R.; Undheim, A.B.E. The diversity of venom: The importance of behavior and venom system morphology in understanding its ecology and evolution. Toxins 2019, 11, 666. [Google Scholar] [CrossRef] [PubMed]
- Fry, B.G.; Roelants, K.; Champagne, D.E.; Scheib, H.; Tyndall, J.D.; King, G.F.; Nevalainen, T.J.; Norman, J.A.; Lewis, R.J.; Norton, R.S.; et al. The toxicogenomic multiverse: Convergent recruitment of proteins into animal venoms. Annu. Rev. Genom. Hum. Genet. 2009, 10, 483–511. [Google Scholar] [CrossRef]
- Moran, Y.; Gordon, D.; Gurevitz, M. Sea anemone toxins affecting voltage-gated sodium channels—Molecular and evolutionary features. Toxicon 2009, 54, 1089–1101. [Google Scholar] [CrossRef]
- Cardoso, F.C.; Lewis, R.J. Sodium channels and pain: From toxins to therapies. Br. J. Pharmacol. 2018, 175, 2138–2157. [Google Scholar] [CrossRef]
- Madio, B.; King, G.F.; Undheim, E.A.B. Sea anemone toxins: A structural overview. Mar. Drugs 2019, 17, 325. [Google Scholar] [CrossRef]
- Liao, Q.; Feng, Y.; Yang, B.; Lee, S.M.-Y. Cnidarian peptide neurotoxins: A new source of various ion channel modulators or blockers against central nervous systems disease. Drug Discovery Today 2019, 24, 189–197. [Google Scholar] [CrossRef]
- Diochot, S.; Loret, E.; Bruhn, T.; Beress, L.; Lazdunski, M. APETx1, a new toxin from the sea anemone Anthopleura elegantissima, blocks voltage-gated human ether-ago-go-related gene potassium channels. Mol. Pharmacol. 2003, 64, 59–69. [Google Scholar] [CrossRef]
- Diochot, S.; Baron, A.; Rash, L.D.; Deval, E.; Escoubas, P.; Scarzello, S.; Salinas, M.; Lazdunski, M. A new sea anemone peptide, APETx2, inhibits ASIC3, a major acid-sensitive channel in sensory neurons. EMBO J. 2004, 23, 1516–1525. [Google Scholar] [CrossRef]
- Kalina, R.; Gladkikh, I.; Dmitrenok, P.; Chernikov, O.; Koshelev, S.; Kvetkina, A.; Kozlov, S.; Kozlovskaya, E.; Monastyrnaya, M. New APETx-like peptides from sea anemone Heteractis crispa modulate ASIC1a channels. Peptides 2018, 104, 41–49. [Google Scholar] [CrossRef]
- Kalina, R.S.; Koshelev, S.G.; Zelepuga, E.A.; Kim, N.Y.; Kozlov, S.A.; Kozlovskaya, E.P.; Monastyrnaya, M.M.; Gladkikh, I.N. APETx-like peptides from the sea anemone Heteractis crispa, diverse in their effect on ASIC1a and ASIC3 ion channels. Toxins 2020, 12, 266. [Google Scholar] [CrossRef] [PubMed]
- Schweitz, H.; Bruhn, T.; Guillemare, E.; Moinier, D.; Lancelin, J.M.; Béress, L.; Lazdunski, M. Kalicludines and kaliseptine. Two different classes of sea anemone toxins for voltage sensitive K+ channels. J. Biol. Chem. 1995, 270, 25121–25126. [Google Scholar] [CrossRef] [PubMed]
- Krishnarjuna, B.; MacRaild, C.A.; Sunanda, P.; Morales, R.A.V.; Peigneur, S.; Macrander, J.; Yu, H.H.; Daly, M.; Raghothama, S.; Dhawan, V.; et al. Structure, folding and stability of a minimal homologue from Anemonia sulcata of the sea anemone potassium channel blocker ShK. Peptides 2018, 99, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Castaneda, O.; Sotolongo, V.; Amor, A.M.; Stocklin, R.; Anderson, A.J.; Harvey, A.L.; Engstrom, A.; Wernstedt, C.; Karlsson, E. Characterization of a potassium channel toxin from the Caribbean Sea anemone Stichodactyla helianthus. Toxicon 1995, 33, 603–613. [Google Scholar] [CrossRef]
- Ciccone, R.; Piccialli, I.; Grieco, P.; Merlino, F.; Annunziato, L.; Pannaccione, A. Synthesis and pharmacological evaluation of a novel peptide based on Anemonia sulcata BDS-I toxin as a new KV3.4 inhibitor exerting a neuroprotective effect against amyloid-β peptide. Front. Chem. 2019, 7, 479. [Google Scholar] [CrossRef]
- Valle, A.; Alvarado-Mesen, J.; Lanio, M.E.; Alvarez, C.; Barbosa, J.A.R.G.; Pazos, I.F. The multigene families of actinoporins (part I): Isoforms and genetic structure. Toxicon 2015, 103, 176–187. [Google Scholar] [CrossRef]
- Leychenko, E.; Isaeva, M.; Tkacheva, E.; Zelepuga, E.; Kvetkina, A.; Guzev, K.; Monastyrnaya, M.; Kozlovskaya, E. Multigene Family of Pore-forming Toxins from Sea Anemone Heteractis crispa. Mar. Drugs 2018, 16, 183. [Google Scholar] [CrossRef]
- Palacios-Ortega, J.; Rivera-de-Torre, E.; Gavilanes, J.G.; Slotte, J.P.; Martínez-del-Pozo, Á. Evaluation of different approaches used to study membrane permeabilization by actinoporins on model lipid vesicles. Biochim. Biophys. Acta–Biomembr. 2020, 1862, 183311. [Google Scholar] [CrossRef]
- Andreev, Y.A.; Kozlov, S.A.; Koshelev, S.G.; Ivanova, E.A.; Monastyrnaya, M.M.; Kozlovskaya, E.P.; Grishin, E.V. Analgesic compound from sea anemone Heteractis crispa is the first polypeptide inhibitor of vanilloid receptor 1 (TRPV1). J. Biol. Chem. 2008, 283, 23914–23921. [Google Scholar] [CrossRef]
- Isaeva, M.; Chausova, E.; Zelepuga, E.; Guzev, K.; Tabakmakher, V.; Monastyrnaya, M.; Kozlovskaya, E. A new multigene superfamily of Kunitz-type protease inhibitors from sea anemone Heteractis crispa. Peptides 2012, 34, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Peigneur, S.; Billen, B.; Derua, R.; Waelkens, E.; Debaveye, S.; Béress, L.; Tytgat, J. A bifunctional sea anemone peptide with Kunitz type protease and potassium channel inhibiting properties. Biochem. Pharmacol. 2011, 82, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Monastyrnaya, M.; Peigneur, S.; Zelepuga, E.; Sintsova, O.; Gladkikh, I.; Leychenko, E.; Isaeva, M.; Tytgat, J.; Kozlovskaya, E. Kunitz-type peptide HCRG21 from the sea anemone Heteractis crispa is a full peptide antagonist of the TRPV1 receptor. Mar. Drugs 2016, 14, 229. [Google Scholar] [CrossRef] [PubMed]
- Gladkikh, I.; Peigneur, S.; Sintsova, O.; Pinheiro-Junior, E.L.; Klimovich, A.; Menshov, A.; Kalinovsky, A.; Isaeva, M.; Monastyrnaya, M.; Kozlovskaya, E.; et al. Kunitz-type peptides from the sea anemone Heteractis crispa demonstrate potassium channel blocking and anti-inflammatory activities. Biomedicines 2020, 8, 473. [Google Scholar] [CrossRef] [PubMed]
- Messerli, S.M.; Greenberg, R.M. Cnidarian toxins acting on voltage-gated ion channels. Mar. Drugs 2006, 4, 70–81. [Google Scholar] [CrossRef]
- Norton, R.S. Enhancing the therapeutic potential of peptide toxins. Expert Opin. Drug Discov. 2017, 12, 611–623. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, J.S.; Redaelli, E.; Zaharenko, A.J.; Cassulini, R.R.; Konno, K.; Pimenta, D.C.; Freitas, J.C.; Clare, J.J.; Wanke, E. Binding specificity of sea anemone toxins to Na(v) l.1-1.6 sodium channels—Unexpected contributions from differences in the IV/S3-S4 outer loop. J. Biol. Chem. 2004, 279, 33323–33335. [Google Scholar] [CrossRef]
- Barzilai, M.G.; Kahn, R.; Regev, N.; Gordon, D.; Moran, Y.; Gurevitz, M. The specificity of Av3 sea anemone toxin for arthropods is determined at linker DI/SS2–S6 in the pore module of target sodium channels. Biochem. J. 2014, 463, 271–277. [Google Scholar] [CrossRef]
- Cestèle, S.; Catterall, W.A. Molecular mechanisms of neurotoxin action on voltage-gated sodium channels. Biochimie 2000, 82, 883–892. [Google Scholar] [CrossRef]
- Benzinger, G.R.; Kyle, J.W.; Blumenthal, K.M.; Hanck, D.A. A specific interaction between the cardiac sodium channel and site-3 toxin anthopleurin-B. J. Biol. Chem. 1998, 273, 80–84. [Google Scholar] [CrossRef]
- Marban, E.; Yamagishi, T.; Tomaselli, G.F. Structure and function of voltage-gated sodium channels. J. Physiol. 1998, 508, 647–657. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.J.; Blumenthal, K.M. Site-3 sea anemone toxins: Molecular probes of gating mechanisms in voltage-dependent sodium channels. Toxicon 2007, 49, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Rathmayer, W.; Béress, L. The effect of toxins from Anemonia sulcata (Coelenterata) on neuromuscular transmission and nerve action potentials in the crayfish (Astacus leptodactylus). J. Comp. Physiol. A 1976, 109, 373–382. [Google Scholar] [CrossRef]
- Ravens, U.; Schöllhorn, E. The effects of a toxin (ATX II) from the sea anemone Anemonia sulcata on the electrical and mechanical activity of the denervated hemidiaphragm of the rat. Toxicon 1983, 21, 131–142. [Google Scholar] [CrossRef]
- Catterall, W.A.; Wisedchaisri, G.; Zheng, N. The chemical basis for electrical signaling. Nat. Chem. Biol. 2017, 13, 455–463. [Google Scholar] [CrossRef]
- Shen, H.; Zhou, Q.; Pan, X.; Li, Z.; Wu, J.; Yan, N. Structure of a eukaryotic voltage-gated sodium channel at near-atomic resolution. Science 2017, 355, eaal4326. [Google Scholar] [CrossRef]
- Goldin, A.L.; Barchi, R.L.; Caldwell, J.H.; Hofmann, F.; Howe, J.R.; Hunter, J.C.; Kallen, R.G.; Mandel, G.; Meisler, M.H.; Netter, Y.B.; et al. Nomenclature of voltage-gated sodium channels. Neuron 2000, 28, 365–368. [Google Scholar] [CrossRef]
- Goldin, A.L. Resurgence of sodium channel research. Annu. Rev. Physiol. 2001, 63, 871–894. [Google Scholar] [CrossRef]
- Mariottini, G.L.; Pane, L. The role of Cnidaria in drug discovery. A review on CNS implications and new perspectives. Recent Pat. CNS Drug Discov. 2013, 8, 110–122. [Google Scholar] [CrossRef]
- Wang, J.; Ou, S.; Wang, Y. Distribution and function of voltage-gated sodium channels in the nervous system. Channels 2017, 11, 534–554. [Google Scholar] [CrossRef]
- Li, Z.-M.; Chen, L.-X.; Li, H. Voltage-gated sodium channels and blockers: An overview and where will they go? Curr. Med. Sci. 2019, 39, 863–873. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.; Shi, H.; Tonggu, L.; Gamal El-Din, T.M.; Lenaeus, M.J.; Zhao, Y.; Yoshioka, C.; Zheng, N.; Catterall, W.A. Structure of the cardiac sodium channel. Cell 2020, 180, 122–134. [Google Scholar] [CrossRef] [PubMed]
- Warashina, A.; Jiang, Z.Y.; Ogura, T. Potential-dependent action of Anemonia sulcata toxins III and IV on sodium channels in crayfish giant axons. Pflug. Arch. 1988, 411, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Vincent, J.P.; Balerna, M.; Barhanin, J.; Fosset, M.; Lazdunski, M. Binding of sea anemone toxin to receptor sites associated with gating system of sodium channel in synaptic nerve endings in vitro. Proc. Natl. Acad. Sci. USA 1980, 77, 1646–1650. [Google Scholar] [CrossRef] [PubMed]
- Stevens, M.; Peigneur, S.; Tytgat, J. Neurotoxins and their binding areas on voltage-gated sodium channels. Front. Pharmacol. 2011, 2, 71. [Google Scholar] [CrossRef] [PubMed]
- Rogers, J.C.; Qu, Y.; Tanada, T.N.; Scheuer, T.; Catterall, W.A. Molecular determinants of high affinity binding of alpha-scorpion toxin and sea anemone toxin in the S3–S4 extracellular loop in domain IV of the Na+ channel alpha subunit. J. Biol. Chem. 1996, 271, 15950–15962. [Google Scholar] [CrossRef]
- Oelstrom, K.; Goldschen-Ohm1, M.P.; Holmgren, M.; Chanda, B. Evolutionarily conserved intracellular gate of voltage-dependent sodium channels. Nat. Commun. 2014, 5, 3420. [Google Scholar] [CrossRef]
- Bosmans, F.; Tytgat, J. Sea anemone venom as a source of insecticidal peptides acting on voltage-gated Na+ channels. Toxicon 2007, 49, 550–560. [Google Scholar] [CrossRef]
- Wanke, E.; Zaharenko, A.J.; Redaelli, E.; Schiavon, E. Actions of sea anemone type 1 neurotoxins on voltage-gated sodium channel isoforms. Toxicon 2009, 54, 1102–1111. [Google Scholar] [CrossRef]
- Hartung, K.; Rathmayer, W. Anemonia sulcata toxins modify activation and inactivation of Na+ currents in a crayfish neurone. Pflugers Arch. 1985, 404, 119–125. [Google Scholar] [CrossRef]
- Ständker, L.; Béress, L.; Garateix, A.; Christ, T.; Ravens, U.; Salceda, E.; Soto, E.; John, H.; Forssmann, W.-G.; Aneiros, A. A new toxin from the sea anemone Condylactis gigantea with effect on sodium channel inactivation. Toxicon 2006, 48, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Sorokina, Z.A.; Chizhmakov, I.V.; Elyakov, G.B.; Kozlovskaya, E.P.; Vozhzhova, E.I. Investigation of inactivation mechanism of fast sodium channels using neurotoxin from the sea anemone Radianthus macrodactylus and various chemical reagents. Physiol. J. 1984, 30, 571–579. [Google Scholar]
- Salgado, V.L.; Kem, W.R. Actions of three structurally distinct sea anemone toxins on crustacean and insect sodium channels. Toxicon 1992, 30, 1365–1381. [Google Scholar] [CrossRef] [PubMed]
- Schwiening, C.J. A brief historical perspective: Hodgkin and Huxley. J. Physiol. 2012, 590, 2571–2575. [Google Scholar] [CrossRef] [PubMed]
- Niklas, B.; Jankowska, M.; Gordon, D.; Béress, L.; Stankiewicz, M.; Nowak, W. Interactions of sea anemone toxins with insect sodium channel-insights from electrophysiology and molecular docking studies. Molecules 2021, 26, 1302. [Google Scholar] [CrossRef]
- Nicosia, A.; Mikov, A.; Cammarata, M.; Colombo, P.; Andreev, Y.; Kozlov, S.; Cuttitta, A. The Anemonia viridis venom: Coupling biochemical purification and RNA-seq for translational research. Mar. Drugs 2018, 16, 407. [Google Scholar] [CrossRef]
- Ramírez-Carreto, S.; Vera-Estrella, R.; Portillo-Bobadilla, T.; Licea-Navarro, A.; Bernaldez-Sarabia, J.; Rudiño-Piñera, E.; Verleyen, J.J.; Rodríguez, E.; Rodríguez-Almazán, C. Transcriptomic and proteomic analysis of the tentacles and mucus of Anthopleura dowii Verrill, 1869. Mar. Drugs 2019, 17, 436. [Google Scholar] [CrossRef]
- Lazcano-Pérez, F.; Hernández-Guzmán, U.; Sánchez-Rodríguez, J.; Arreguín-Espinosa, R. Cnidarian neurotoxic peptides affecting central nervous system targets. Cent. Nerv. Syst. Agents Med. Chem. 2016, 16, 173–182. [Google Scholar] [CrossRef]
- Metrione, R.M.; Schweitz, H.; Walsh, K.A. The amino acid sequence of toxin RpIII from the sea anemone, Radianthus paumotensis. FEBS Lett. 1987, 218, 59–62. [Google Scholar] [CrossRef]
- Al-Sabi, A.; McArthur, J.; Ostroumov, V.; French, R.J. Marine toxins that target voltage-gated sodium channels. Mar. Drugs 2006, 4, 157–192. [Google Scholar] [CrossRef]
- Alsen, C. Biological significance of peptides from Anemonia sulcata. Fed. Proc. 1983, 42, 101–108. [Google Scholar] [PubMed]
- Chahine, M.; Plante, E.; Kallen, R.G.J. Sea anemone toxin (ATX II) modulation of heart and skeletal muscle sodium channel alpha-subunits expressed in tsA201 cells. Membr. Biol. 1996, 152, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Moran, Y.; Weinberger, H.; Sullivan, J.C.; Reitzel, A.M.; Finnerty, J.R.; Gurevitz, M. Concerted evolution of sea anemone neurotoxin genes is revealed through analysis of the Nematostella vectensis genome. Mol. Biol. Evol. 2008, 25, 737–747. [Google Scholar] [CrossRef] [PubMed]
- Wunderer, G.; Eulitz, M. Amino-acid sequence of toxin I from Anemonia sulcata. Eur. J. Biochem. 1978, 89, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Scheffler, J.J.; Tsugita, A.; Linden, G.; Schweitz, H.; Lazdunski, M. The amino acid sequence of toxin V from Anemonia sulcata. Biochem. Biophys. Res. Commun. 1982, 107, 272–278. [Google Scholar] [CrossRef]
- Moran, Y.; Weinberger, H.; Lazarus, N.; Gur, M.; Kahn, R.; Gordon, D.; Gurevitz, M. Fusion and retrotransposition events in the evolution of the sea anemone Anemonia viridis neurotoxin genes. J. Mol. Evol. 2009, 69, 115–124. [Google Scholar] [CrossRef]
- Kelso, G.J.; Blumenthal, K.M. Identification and characterization of novel sodium channel toxins from the sea anemone Anthopleura xanthogrammica. Toxicon 1998, 36, 41–51. [Google Scholar] [CrossRef]
- Honma, T.; Hasegawa, Y.; Ishida, M.; Nagai, H.; Nagashima, Y.; Shiomi, K. Isolation and molecular cloning of novel peptide toxins from the sea anemone Antheopsis maculata. Toxicon 2005, 45, 33–41. [Google Scholar] [CrossRef]
- Shiomi, K.; Lin, X.-Y.; Nagashima, Y.; Ishima, M. Isolation and amino acid sequence of a polypeptide toxin from the sea anemone Radianthus crispus. Fish. Sci. 1996, 62, 629–633. [Google Scholar] [CrossRef]
- Salceda, E.; Perez-Castells, J.; Lopez-Mendez, B.; Garateix, A.; Salazar, H.; Lopez, O.; Anerous, A.; Standker, L.; Béress, L.; Forssman, W.-G.; et al. CgNa, a type I toxin from the giant Caribbean sea anemone Condylactis gigantea shows structural similarities to both type I and II toxins, as well as distinctive structural and functional properties (1). Biochem. J. 2007, 406, 67–76. [Google Scholar] [CrossRef]
- Shiomi, K.; Honma, T.; Ide, M.; Nagashima, Y.; Ishida, M.; Chino, M. An epidermal growth factor-like toxin and two sodium channel toxins from the sea anemone Stichodactyla gigantea. Toxicon 2003, 41, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Malpezzi, E.L.A.; de Freitas, J.C.; Muramoto, K.; Kamiya, H. Characterization of peptides in sea anemone venom collected by a novel procedure. Toxicon 1993, 31, 853–864. [Google Scholar] [CrossRef] [PubMed]
- Goudet, C.; Ferrer, T.; Galan, L.; Artiles, A.; Batista, C.V.F.; Possani, L.D.; Alvarez, J.; Aneiros, A.; Tytgat, J. Characterization of two Bunodosoma granulifera toxins active on cardiac sodium channels. Br. J. Pharmacol. 2001, 134, 1195–1206. [Google Scholar] [CrossRef] [PubMed]
- Cunha, R.B.; Santana, A.N.; Amaral, P.C.; Carvalho, M.D.; Carvalho, D.M.; Cavalheiro, E.A.; Maigret, B.; Ricart, C.A.; Cardi, B.A.; Sousa, M.V.; et al. Primary structure, behavioral and electroencephalographic effects of an epileptogenic peptide from the sea anemone Bunodosoma cangicum. Toxicon 2005, 45, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Zaharenko, A.J.; Ferreira, W.A., Jr.; Oliveira, J.S.; Richardson, M.; Pimenta, D.C.; Konno, K.; Portaro, F.C.V.; de Freitas, J.C. Proteomics of the neurotoxic fraction from the sea anemone Bunodosoma cangicum venom: Novel peptides belonging to new classes of toxins. Comp. Biochem. Physiol. Part D Genom. Proteom. 2008, 3, 219–225. [Google Scholar] [CrossRef]
- Zaharenko, A.J.; Ferreira, W.A., Jr.; de Oliveira, J.S.; Konno, K.; Richardson, M.; Schiavon, E.; Wanke, E.; de Freitas, J.C. Revisiting cangitoxin, a sea anemone peptide: Purification and characterization of cangitoxins II and III from the venom of Bunodosoma cangicum. Toxicon 2008, 51, 1303–1307. [Google Scholar] [CrossRef]
- Bruhn, T.; Schaller, C.; Schulze, C.; Sanchez-Rodriguez, J.; Dannmeier, C.; Ravens, U.; Heubach, J.F.; Eckhardt, K.; Schmidtmayer, J.; Schmidt, H.; et al. Isolation and characterization of five neurotoxic and cardiotoxic polypeptides from the sea anemone Anthopleura elegantissima. Toxicon 2001, 39, 693–702. [Google Scholar] [CrossRef]
- Norton, R.S. Structure and structure-function relationships of sea anemone proteins that interact with the sodium channel. Toxicon 1991, 29, 1051–1084. [Google Scholar] [CrossRef]
- Wang, L.; Ou, J.; Peng, L.; Zhong, X.; Du, J.; Liu, Y.; Huang, Y.; Liu, W.; Zhang, Y.; Dong, M.; et al. Functional expression and characterization of four novel neurotoxins from sea anemone Anthopleura sp. Biochem. Biophys. Res. Commun. 2004, 313, 163–170. [Google Scholar] [CrossRef]
- Sunahara, S.; Muramoto, K.; Tenma, K.; Kamiya, H. Amino acid sequence of two sea anemone toxins from Anthopleura fuscoviridis. Toxicon 1987, 25, 211–219. [Google Scholar] [CrossRef]
- van Losenoord, W.; Krause, J.; Parker-Nance, S.; Krause, R.; Stoychev, S.; Frost, C.L. Purification and biochemical characterization of a putative sodium channel agonist secreted from the South African Knobbly sea anemone Bunodosoma capense. Toxicon 2019, 168, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Nesher, N.; Shapira, E.; Sher, D.; Moran, Y.; Tsveyer, L.; Turchetti-Maia, A.L.; Horowitz, M.; Hochner, B.; Zlotkin, E. AdE-1, a new inotropic Na+ channel toxin AdE-1 from Aiptasia diaphana, is similar to, yet distinct from, known anemone Na+ channel toxins. Biochem. J. 2013, 451, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Zykova, T.A.; Kozlovskaya, E.P.; Elyakov, G.B. Amino acid sequence of neurotoxin II from the sea anemone Radianthus macrodactylus. Bioorg. Khim. 1988, 14, 878–882. [Google Scholar] [PubMed]
- Zykova, T.A.; Kozlovskaya, E.P.; Elyakov, G.B. Amino acid sequence of neurotoxins IV and V from the sea anemone Radianthus macrodactylus. Bioorg. Khim. 1988, 14, 1489–1494. [Google Scholar] [PubMed]
- Honma, T.; Kawahata, S.; Ishida, M.; Nagai, H.; Nagashima, Y.; Shiomi, K. Novel peptide toxins from the sea anemone Stichodactyla haddoni. Peptides 2008, 29, 536–544. [Google Scholar] [CrossRef]
- Wilcox, G.R.; Fogh, R.H.; Norton, R.S. Refined structure in solution of the sea anemone neurotoxin ShI. J. Biol. Chem. 1993, 268, 24707–24719. [Google Scholar] [CrossRef]
- Maeda, M.; Honma, T.; Shiomi, K. Isolation and cDNA cloning of type 2 sodium channel peptide toxins from three species of sea anemones (Cryptodendrum adhaesivum, Heterodactyla hemprichii and Thalassianthus aster) belonging to the family Thalassianthidae. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2010, 157, 389–393. [Google Scholar] [CrossRef]
- Catterall, W.A. Neurotoxins as allosteric modifiers of voltage-sensitive sodium channels. Adv. Cytopharmacol. 1979, 3, 305–316. [Google Scholar]
- Alsen, C.; Harris, J.B.; Tessereus, I. Mechanical and electrophysiological effects of sea anemone (Anemonia sulcata) toxins on rat innervated and denervated skeletal muscles. Brit. J. Pharmacol. 1981, 74, 61–71. [Google Scholar] [CrossRef][Green Version]
- Moran, Y.; Kahn, R.; Cohen, L.; Gur, M.; Karbat, I.; Gordon, D.; Gurevitz, M. Molecular analysis of the sea anemone toxin Av3 reveals selectivity to insects and demonstrates the heterogeneity of receptor site-3 on voltage-gated Na+ channels. Biochem. J. 2007, 406, 41–48. [Google Scholar] [CrossRef]
- Hanck, D.A.; Sheets, M.F. Site-3 toxins and cardiac sodium channels. Toxicon 2007, 49, 181–193. [Google Scholar] [CrossRef] [PubMed]
- Martinez, G.; Kopeyan, C.; Schweitz, H.; Lazdunski, M. Toxin III from Anemonia sulcata: Primary structure. FEBS Lett. 1977, 84, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Béress, L.; Wunderer, G.; Wachter, E. Amino acid sequence of toxin III from Anemonia sulcata. Hoppe Seylers Z. Physiol. Chem. 1977, 358, 985–988. [Google Scholar] [CrossRef] [PubMed]
- Nishida, S.; Fujita, S.; Warashina, A.; Satake, M.; Tamiya, N. Amino acid sequence of a sea anemone toxin from Parasicyonis actinostoloides. Eur. J. Biochem. 1985, 150, 171–173. [Google Scholar] [CrossRef]
- Honma, T.; Iso, T.; Ishida, M.; Nagashima, Y.; Shiomi, K. Occurrence of type 3 sodium channel peptide toxins in two species of sea anemones (Dofleinia armata and Entacmaea ramsayi). Toxicon 2003, 41, 637–639. [Google Scholar] [CrossRef]
- Norton, R.S.; Cross, K.; Braach-Maksvytis, V.; Wachter, E. 1H-NMR study of the solution properties and secondary structure of neurotoxin III from the sea anemone Anemonia sulcata. Biochem. J. 1993, 293, 545–551. [Google Scholar] [CrossRef][Green Version]
- Manoleras, N.; Norton, R.S. Three-dimensional structure in solution of neurotoxin III from the sea anemone Anemonia sulcata. Biochemistry 1994, 33, 11051–11061. [Google Scholar] [CrossRef]
- Nishida, S.; Fujita, S.; Warashina, A. Parasicyonis toxin: Effect on crayfish giant axon. Comp. Biochem. Physiol. C. Comp. Pharmacol. 1980, 67C, 71–74. [Google Scholar] [CrossRef]
- Warashina, A.; Ogura, T.; Fujita, S. Binding properties of sea anemone toxins to sodium channels in the crayfish giant axon. Comp. Biochem. Physiol. C. Comp. Pharmacol. Toxicol. 1988, 90, 351–359. [Google Scholar]
- Cariello, L.; de Santis, A.; Fiore, F.; Piccoli, R.; Spagnuolo, A.; Zanetti, L.; Parente, A. Calitoxin, a neurotoxic peptide from the sea anemone Calliactis parasitica: Amino acid sequence and electrophysiological properties. Biochemistry 1989, 28, 2484–2489. [Google Scholar] [CrossRef]
- Spagnuolo, A.; Zanetti, L.; Cariello, L.; Piccoli, R. Isolation and characterization of two genes encoding calitoxins, neurotoxic peptides from Calliactis parasitica (Cnidaria). Gene 1994, 138, 187–191. [Google Scholar] [CrossRef] [PubMed]
- Kono, A.; Honma, T.; Shiomi, K. Important amino acid residues for the crab toxicity of PaTX, a type 3 sodium channel peptide toxin from the sea anemone Entacmaea actinostoloides. Fish Sci. 2015, 81, 187–192. [Google Scholar] [CrossRef]
- Ishida, M.; Yokoyama, A.; Shimakura, K.; Nagashima, Y.; Shiomi, K. Halcurin, a polypeptide toxin from the sea anemone Halcurias sp. with a structural resemblance to type 1 and 2 toxins. Toxicon 1997, 35, 537–544. [Google Scholar] [CrossRef] [PubMed]
- Moran, Y.; Gurevitz, M. When positive selection of neurotoxin genes is missing. The riddle of the sea anemone Nematostella vectensis. FEBS J. 2006, 273, 3886–3892. [Google Scholar] [CrossRef] [PubMed]
- Moran, Y.; Weinberger, H.; Reitzel, A.M.; Sullivan, J.C.; Kahn, R.; Gordon, D.; Finnerty, J.R.; Gurevitz, M. Intron retention as a posttranscriptional regulatory mechanism of neurotoxin expression at early life stages of the starlet anemone Nematostella vectensis. J. Mol. Biol. 2008, 380, 437–443. [Google Scholar] [CrossRef]
- UniProt. B1NWS1 NA227_NEMVE. Available online: https://www.uniprot.org/uniprot/B1NWS1 (accessed on 27 October 2022).
- Babenko, V.V.; Mikov, A.N.; Manuvera, V.A.; Anikanov, N.A.; Kovalchuk, S.I.; Andreev, Y.A.; Logashina, Y.A.; Kornilov, D.A.; Manolov, A.I.; Sanamyan, N.P.; et al. Identification of unusual peptides with new Cys frameworks in the venom of the cold-water sea anemone Cnidopus japonicus. Sci. Rep. 2017, 7, 14534. [Google Scholar] [CrossRef]
- Liu, P.; Jo, S.; Bean, B.P. Modulation of neuronal sodium channels by the sea anemone peptide BDS-I. J. Neurophysiol. 2012, 107, 3155–3167. [Google Scholar] [CrossRef]
- Oliveira, J.S.; Zaharenko, A.J.; Ferreira, W.A., Jr.; Konno, K.; Shida, C.S.; Richardson, M.; Lúcio, A.D.; Beirão, P.S. BcIV, a new paralyzing peptide obtained from the venom of the sea anemone Bunodosoma caissarum. A comparison with the Na+ channel toxin BcIII. Biochim. Biophys. Acta 2006, 1764, 1592–1600. [Google Scholar] [CrossRef]
- Diochot, S.; Schweitz, H.; Béress, L.; Lazdunski, M. Sea anemone peptides with a specific blocking activity against the fast inactivating potassium channel Kv3.4. J. Biol. Chem. 1998, 273, 6744–6749. [Google Scholar] [CrossRef]
- Torda, A.E.; Mabbutt, B.C.; van Gunsteren, W.F.; Norton, R.S. Backbone folding of the polypeptide cardiac stimulant anthopleurin-A determined by nuclear magnetic resonance, distance geometry and molecular dynamics. FEBS Lett. 1988, 239, 266–270. [Google Scholar] [CrossRef]
- Monks, S.A.; Pallaghy, P.K.; Scanlon, M.J.; Norton, R.S. Solution structure of the cardiostimulant polypeptide anthopleurin-B and comparison with anthopleurin-A. Structure 1995, 3, 791–803. [Google Scholar] [CrossRef]
- Widmer, H.; Billeter, M.; Wüthrich, K. Three-dimensional structure of the neurotoxin ATX Ia from Anemonia sulcata in aqueous solution determined by nuclear magnetic resonance spectroscopy. Proteins 1989, 6, 357–371. [Google Scholar] [CrossRef]
- Fogh, R.H.; Kem, W.R.; Norton, R.S. Solution structure of neurotoxin I from the sea anemone Stichodactyla helianthus. A nuclear magnetic resonance, distance geometry, and restrained molecular dynamics study. J. Biol. Chem. 1990, 265, 13016–13028. [Google Scholar] [CrossRef] [PubMed]
- Seibert, A.L.; Liu, J.; Hanck, D.A.; Blumenthal, K.M. Arg-14 loop of site 3 anemone toxins: effects of glycine replacement on toxin affinity. Biochemistry 2003, 42, 14515–14521. [Google Scholar] [CrossRef] [PubMed]
- Mouhat, S.; Jouirou, B.; Mosbah, A.; De Waard, M.; Sabatier, J.-M. Diversity of folds in animal toxins acting on ion channels. Biochem. J. 2004, 378, 717–726. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera? A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef]
- Hanck, D.A.; Sheets, M.F. Modification of inactivation in cardiac sodium channels: Ionic current studies with Anthopleurin-A toxin. J. Gen. Physiol. 1995, 106, 601–616. [Google Scholar] [CrossRef]
- Béress, L.; Ritter, R.; Ravens, U. The influence of the rate of electrical stimulation on the effects of the Anemonia sulcata toxin ATX II in guinea pig papillary muscle. Eur. J. Pharmacol. 1982, 79, 265–272. [Google Scholar] [CrossRef]
- Sunagar, K.; Morgenstern, D.; Reitzel, A.M.; Moran, Y. Ecological venomics: How genomics, transcriptomics and proteomics can shed new light on the ecology and evolution of venom. J. Proteom. 2016, 135, 62–72. [Google Scholar] [CrossRef]
- Isenberg, G.; Ravens, U. The effects of the Anemonia sulcata toxin (ATX II) on membrane currents of isolated mammalian myocytes. J. Physiol. 1984, 357, 127–149. [Google Scholar] [CrossRef]
- Spencer, C.I. Actions of ATX-II and other gating-modifiers on Na+ currents in HEK-293 cells expressing WT and deltaKPQ hNav1.5 Na+ channels. Toxicon 2009, 53, 78–89. [Google Scholar] [CrossRef] [PubMed]
- Zaharenko, A.J.; Schiavona, E.; Ferreira, W.A., Jr.; Lecchi, M.; de Freitas, J.C.; Richardson, M.; Wanke, E. Characterization of selectivity and pharmacophores of type 1 sea anemone toxins by screening seven Nav sodium channel isoforms. Peptides 2012, 34, 158–167. [Google Scholar] [CrossRef] [PubMed]
- Billen, B.; Debaveye, S.; Béress, L.; Garateix, A.; Tytgat, J. Phyla- and subtype-selectivity of CgNa, a Na+ channel toxin from the venom of the Giant Caribbean sea anemone Condylactis gigantea. Front. Pharmacol. 2010, 1, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, M.J.; Blumenthal, K.M. Cloning and expression of wild-type and mutant forms of the cardiotonic polypeptide anthopleurin B. J. Biol. Chem. 1992, 267, 13958–13963. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, M.J.; Blumenthal, K.M. Importance of the unique cationic residues arginine 12 and lysine 49 in the activity of the cardiotonic polypeptide anthopleurin-B. J. Biol. Chem. 1994, 269, 254–259. [Google Scholar] [CrossRef] [PubMed]
- Dias-Kadambi, B.L.; Drum, C.L.; Hanck, D.A.; Blumenthal, K.M. Leucine 18, a hydrophobic residue essential for high affinity binding of anthopleurin B to the voltage-sensitive sodium channel. J. Biol. Chem. 1996, 271, 9422–9428. [Google Scholar] [CrossRef] [PubMed]
- Mahnir, V.M.; Kozlovskaya, E.P.; Elyakov, G.B. Modification of arginine in sea anemone toxin RTX-III from Radianthus macrodactylus. Toxicon 1989, 27, 1075–1084. [Google Scholar] [CrossRef]
- Mahnir, V.M.; Kozlovskaya, E.P. Structure-toxicity relationships of neurotoxin RTX-III from the sea anemone Radianthus macrodactylus: Modification of amino groups. Toxicon 1991, 29, 819–826. [Google Scholar] [CrossRef]
- Barhanin, J.; Hugues, M.; Schweitz, H.; Vincent, J.P.; Lazdunski, M. Structure-function relationships of sea anemone toxin II from Anemonia sulcata. J. Biol. Chem. 1981, 256, 5764–5769. [Google Scholar] [CrossRef]
- Moran, Y.; Cohen, L.; Kahn, R.; Karbat, I.; Gordon, D.; Gurevitz, M. Expression and mutagenesis of the sea anemone toxin Av2 reveals key amino acid residues important for activity on voltage-gated sodium channels. Biochemistry 2006, 45, 8864–8873. [Google Scholar] [CrossRef]
- Khera, P.K.; Blumenthal, K.M. Role of the cationic residues arginine 14 and lysine 48 in the function of the cardiotonic polypeptide anthopleurin B. J. Biol. Chem. 1994, 269, 921–925. [Google Scholar] [PubMed]
- Khera, P.K.; Benzinger, G.R.; Lipkind, G.; Drum, C.L.; Hanck, D.A.; Blumenthal, K.M. Multiple cationic residues of anthopleurin B that determine high affinity and channel isoform discrimination. Biochemistry 1995, 34, 8533–8541. [Google Scholar] [CrossRef] [PubMed]
- Norton, R.S. Structures of sea anemone toxins. Toxicon 2009, 54, 1075–1088. [Google Scholar] [CrossRef] [PubMed]
- Blumenthal, K.M.; Seibert, A.L. Voltage-gated sodium channel toxins: Poisons, probes, and future promise. Cell Biochem. Biophys. 2003, 38, 215–238. [Google Scholar] [CrossRef]
- Kalina, R.S.; Peigneur, S.; Zelepuga, E.A.; Dmitrenok, P.S.; Kvetkina, A.N.; Kim, N.Y.; Leychenko, E.V.; Tytgat, J.; Kozlovskaya, E.P.; Monastyrnaya, M.M.; et al. New insights into the type ii toxins from the sea anemone Heteractis crispa. Toxins 2020, 12, 44. [Google Scholar] [CrossRef]
- Zhu, Q.; Du, Y.; Nomura, Y.; Gao, R.; Cang, Z.; Wei, G.W.; Gordon, D.; Gurevitz, M.; Groome, J.; Dong, K. Charge substitutions at the voltage-sensing module of domain III enhance actions of site-3 and site-4 toxins on an insect sodium channel. Insect Biochem. Mol. Biol. 2021, 137, 103625. [Google Scholar] [CrossRef]
- Kelso, G.J.; Drum, C.L.; Hanck, D.A.; Blumenthal, K.M. Role of Pro-13 in directing high-affinity binding of Anthopleurin B to the voltage-sensitive sodium channel. Biochemistry 1996, 25, 14157–14164. [Google Scholar] [CrossRef]
- Khera, P.K.; Blumenthal, K.M. Importance of highly conserved anionic residues and electrostatic interactions in the activity and structure of the cardiotonic polypeptide anthopleurin B. Biochemistry 1996, 35, 3503–3507. [Google Scholar] [CrossRef]
- Xiao, Y.; Blumenthal, K.; Cummins, T.R. Gating-pore currents demonstrate selective and specific modulation of individual sodium channel voltage-sensors by biological toxins. Mol. Pharmacol. 2014, 86, 159–167. [Google Scholar] [CrossRef]
- Ishikawa, Y.; Onadera, K.; Takeuchi, A. Purification and effest of the neurotoxin from sea anemone Parasicyonis actinostoloides. J. Neurochem. 1979, 33, 69–72. [Google Scholar] [CrossRef]
- Baydan, L.V.; Kozlovskaya, E.P.; Elyakov, G.B.; Tishkin, S.M.; Shuba, M.F.; Elyakov, G.B. Effect of anemonotoxin on neuromuscular transmission in skeletal and smooth muscles. Dokl. Acad. Sci. USSR 1981, 259, 1000–1002. [Google Scholar]
- Wemmer, D.E.; Kumar, N.V.; Metrione, R.M.; Lazdunski, M.; Drobny, G.; Kallenbach, N.R. NMR analysis and sequence of toxin II from the sea anemone Radianthus paumotensis. Biochemistry 1986, 25, 6842–6849. [Google Scholar] [CrossRef] [PubMed]
- Moran, Y.; Praher, D.; Schlesinger, A.; Ayalon, A.; Tal, Y.; Technau, U. Analysis of soluble protein contents from the nematocysts of a model sea anemone sheds light on venom evolution. Mar. Biotechnol. 2013, 15, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Sunagar, K.; Undheim, E.A.B.; Chan, A.H.C.; Koludarov, I.; Muñoz-Gómez, S.A.; Antunes, A.; Fry, B.G. Evolution stings: The origin and diversification of scorpion toxin peptide scaffolds. Toxins 2013, 5, 2456–2487. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.; Lin, S.; Huang, H.; Hsiao, M. Genetic organization of α-bungarotoxins from Bungarus multicinctus (Taiwan banded krait): Evidence showing that the production of α-bungarotoxin isotoxins is not derived from edited mRNAs. Nucleic Acids Res. 1999, 27, 3970–3975. [Google Scholar] [CrossRef] [PubMed]
- Tamiya, T.; Ohno, S.; Nishimura, E.; Fujimi, T.J.; Tsuchiya, T. Complete nucleotide sequences of cDNAs encoding long chain alpha-neurotoxins from sea krait, Laticauda semifasciata. Toxicon 1999, 37, 181–185. [Google Scholar] [CrossRef]
- Ohno, M.; Menez, R.; Ogawa, T.; Danse, J.M.; Shimohigashi, Y.; Fromen, C.; Ducancel, F.; Zinn-Justin, S.; Le Du, M.H.; Boulain, J.C.; et al. Molecular evolution of snake toxins: Is the functional diversity of snake toxins associated with a mechanism of accelerated evolution? Prog. Nucleic Acid Res. Mol. Biol. 1998, 59, 307–364. [Google Scholar] [CrossRef]
- Puillandre, N.; Koua, D.; Favreau, P.; Olivera, B.M.; Stocklin, R. Molecular phylogeny, classification and evolution of conopeptides. J. Mol. Evol. 2012, 74, 297–309. [Google Scholar] [CrossRef]
- Smith, E.G.; Surm, J.M.; Macrander, J.; Simhi, A.; Amir, G.; Sachkova, M.Y.; Lewandowska, M.; Reitzel, A.M.; Moran, Y. Dominant toxin hypothesis: Unravelling the venom phenotype across micro and macroevolution. BioRxiv 2022. [Google Scholar] [CrossRef]
- Hinds, M.G.; Norton, R.S. Sequential 1H-NMR Assignments of Neurotoxin III from Sea anemone Heteractis macrodactylus and structural comparison with related toxins. J. Protein Chem. 1993, 12, 371–378. [Google Scholar] [CrossRef]
- Nabiullin, A.A.; Odinokov, S.E.; Vozhzhova, E.I.; Kozlovskaya, E.P.; Elyakov, G.B. Study of the conformational stability of toxin I from the sea anemone Radianthus macrodactylus. Bioorg. Khim. 1982, 8, 1644–1648. [Google Scholar]
- Odinokov, S.E.; Nabiullin, A.A.; Kozlovskaya, E.P.; Elyakov, G.B. Structure-function relationship of polypeptide toxins: Modifying gating mechanism of sodium channel. Pure Appl. Chem. 1989, 61, 497–500. [Google Scholar] [CrossRef]
- Kryshtal, O.A.; Osipchuk, I.V.; Elyakov, G.B.; Kozlovskaya, E.P. Effect of toxin from the sea anemone Homostichantus duerdemi on the sodium inwart current in mammalian neurons. Neurofiziologiia 1982, 14, 402–409. [Google Scholar]
- Sorokina, Z.A.; Chizhmakov, I.V.; Kozlovskaya, E.P.; Vozhzhova, E.V.; Elyakov, G.B. Positive cooperativity of tetrodotoxin binding by sodium channels of neurons in the spinal ganglia of rats, induced by the anemonotoxin RTX-III. Dokl. Acad. Sci. USSR 1985, 282, 433–436. [Google Scholar]
- Mahnir, V.M.; Kozlovskaya, E.P. Modification of the RTX-III neurotoxin from the sea anemone Radianthus macrodactylus with acetic anhydride. Bioorg. Khim. 1989, 15, 465–470. [Google Scholar]
- Mahnir, V.M.; Kozlovskaya, E.P. Modification of a neurotoxin from the sea anemone Radianthus macrodactylus. Bioorg. Khim. 1990, 16, 643–648. [Google Scholar]
- Mahnir, V.M.; Kozlovskaya, E.P.; Elyakov, G.B. Modification of carboxyl groups in sea anemone toxin RTX-III from Radianthus macrodactylus. Toxicon 1990, 28, 1255–1263. [Google Scholar] [CrossRef]
- Mahnir, V.M.; Kozlovskaya, E.P. An unusual result of the modification of anemonotoxin RTX-III with malonic aldehyde. Chem. Nat. Comp. 1989, 3, 383–387. [Google Scholar]
- Kuhn-Nentwig, L.; Schaller, J.; Schürch, S.; Nentwig, W. Venom of Cupiennius salei (Ctenidae). In Spider Venoms; Gopalakrishnakone, P., Corzo, G.A., de Lima, M.E., Diego-García, E., Eds.; Springer: Berlin/Heidelberg, Germany, 2016; pp. 47–70. [Google Scholar]
- Israel, M.R.; Tay, B.; Deuis, J.R.; Vetter, I. Sodium channels and venom peptide pharmacology. Adv. Pharmacol. 2017, 79, 67–116. [Google Scholar] [CrossRef]
- Roger, S.; Gillet, L.; Le Guennec, J.Y.; Besson, P. Voltage-gated sodium channels and cancer: Is excitability their primary role? Front. Pharmacol. 2015, 6, 1–22. [Google Scholar] [CrossRef]




| Modified Residue(s) | Reagent | LD50 Modif./LD50 Nativ (Mice) |
|---|---|---|
| Gly1 (α-NH2) Lys4 Lys (C-terminal) | [3H]-acetic anhydride | 12 2 2 |
| Arg13 | malonic aldehyde phenylglyoxal 1,2-cyclohexanedione | 4–5 |
| COOH | [3H]-glycine methyl ether/EDC [14C]-trimethyloxonium tetrafluoroborate | 1–2 |
| Trp30 | 2-hydroxy-5-nitrobenzyl bromide | 1 |
| S-S bonds | 2-mercaptoethanol/iodoacetamide | >100 |
| NaTx | Voltage-Gated Sodium Channel Subtype | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| NaV1.1 | NaV1.2 | NaV1.3 | NaV1.4 | NaV1.5 | NaV1.6 | NaV1.7 | NaV1.8 | BgNaV1 | VdNaV1 | |
| δ-SHTX Hcr1f | + | + | - | - | - | + | n.d. | - | + | + |
| RTX-III | - | - | + | - | - | + | n.d. | - | + | + |
| RTX-VI | - | + | - | - | - | + | n.d. | - | + | + |
| BcIII | + | + | + | + | + | + | n.d. | n.d. | n.d. | n.d. |
| AFT-II | + | + | + | + | + | + | n.d. | n.d. | n.d. | n.d. |
| ATX-II | + | + | + | + | + | + | n.d. | n.d. | n.d. | n.d. |
| CGTX-II | - | - | - | - | + | + | - | n.d. | n.d. | n.d. |
| δ-AITX-Bcg1a | + | + | + | + | + | + | - | n.d. | n.d. | n.d. |
| δ-AITX-Bcg1b | - | - | - | - | - | - | - | n.d. | n.d. | n.d. |
| BgII | n.d. | + | n.d. | + | + | n.d. | n.d. | - | n.d. | n.d. |
| BgIII | n.d. | + | n.d. | + | + | n.d. | n.d. | - | n.d. | n.d. |
| CgNa | - | - | + | - | + | + | - | n.d. | n.d. | n.d. |
| Nv1 | - | - | - | - | - | - | n.d. | - | + | n.d. |
| Nv4/Nv5 | - | - | - | - | - | - | n.d. | - | - | n.d. |
| Toxin | EC50 nM | |||||||
|---|---|---|---|---|---|---|---|---|
| NaV1.1 | NaV1.2 | NaV1.3 | NaV1.4 | NaV1.5 | NaV1.6 | NaV1.7 | BgNaV1 | |
| SHTX-Hcr1f | 79.5 | 183.5 | 226.1 | |||||
| RTX-III | – | 381.8 | 978.1 | |||||
| RTX-VI | 120.9 | 282.3 | 760.5 | |||||
| ATX-II | 6.01 | 7.88 | 759.22 | 109.49 | 49.05 | ~180 | 1800 | |
| AFT-II | 390.55 | ~2000 | 459.36 | 30.62 | 62.5 | ~300 | 5800 | |
| BcIII | ~300 | 1449.17 | 1458.42 | 820.84 | 307 | ~900 | 5700 | |
| CGTX-II | 165 | >1000 | >1000 | >1000 | 105 | 133 | ||
| BgII | 300 | 1000 | 500 | |||||
| BgIII | 18,900 | 9800 | 5100 | |||||
| δ-AITX-Bcg1a | 453 | 440 | 740 | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monastyrnaya, M.M.; Kalina, R.S.; Kozlovskaya, E.P. The Sea Anemone Neurotoxins Modulating Sodium Channels: An Insight at Structure and Functional Activity after Four Decades of Investigation. Toxins 2023, 15, 8. https://doi.org/10.3390/toxins15010008
Monastyrnaya MM, Kalina RS, Kozlovskaya EP. The Sea Anemone Neurotoxins Modulating Sodium Channels: An Insight at Structure and Functional Activity after Four Decades of Investigation. Toxins. 2023; 15(1):8. https://doi.org/10.3390/toxins15010008
Chicago/Turabian StyleMonastyrnaya, Margarita Mikhailovna, Rimma Sergeevna Kalina, and Emma Pavlovna Kozlovskaya. 2023. "The Sea Anemone Neurotoxins Modulating Sodium Channels: An Insight at Structure and Functional Activity after Four Decades of Investigation" Toxins 15, no. 1: 8. https://doi.org/10.3390/toxins15010008
APA StyleMonastyrnaya, M. M., Kalina, R. S., & Kozlovskaya, E. P. (2023). The Sea Anemone Neurotoxins Modulating Sodium Channels: An Insight at Structure and Functional Activity after Four Decades of Investigation. Toxins, 15(1), 8. https://doi.org/10.3390/toxins15010008