The Mechanism of Houttuynia cordata Embryotoxicity Was Explored in Combination with an Experimental Model and Network Pharmacology

 ,
,
Abstract
1. Introduction
2. Results
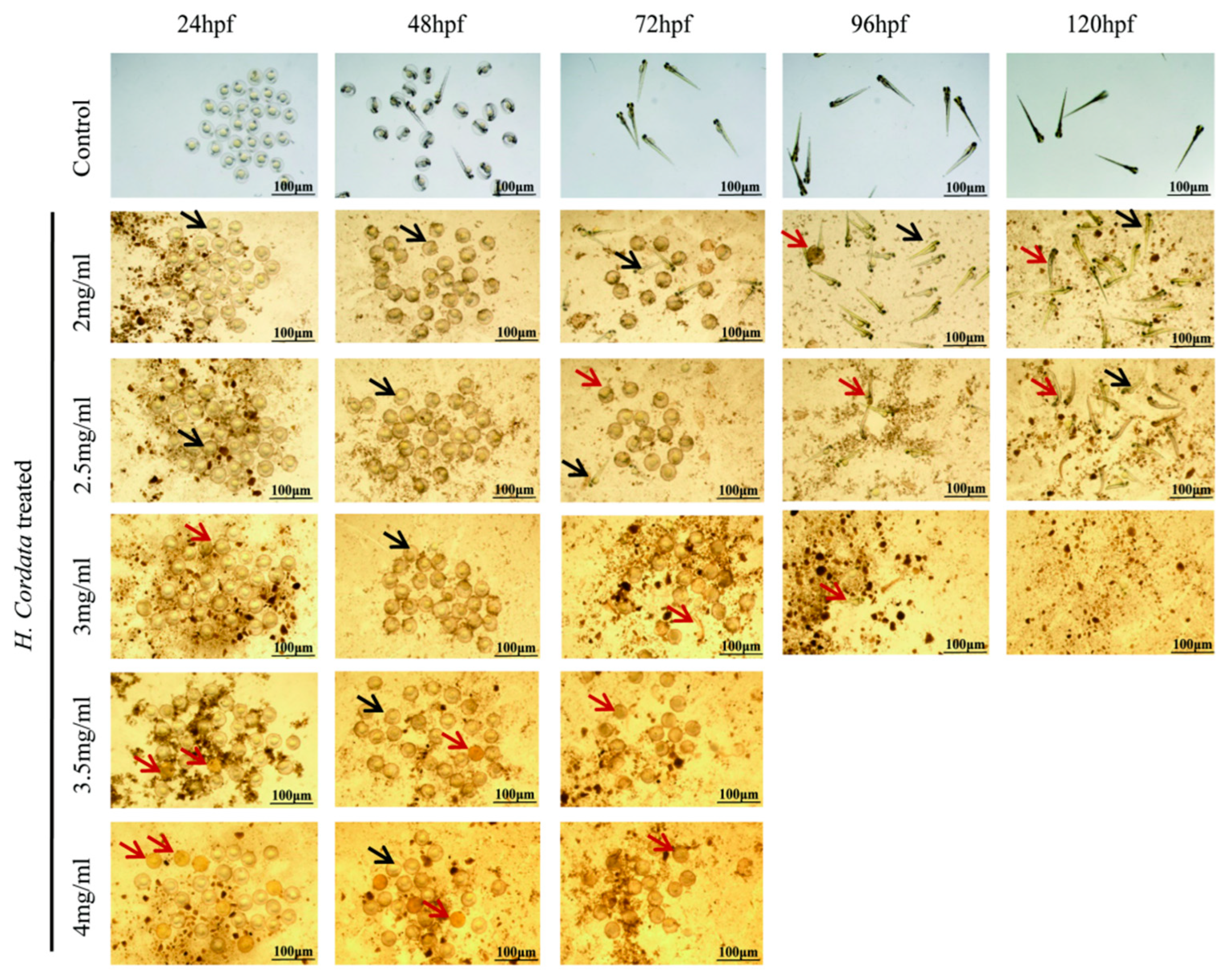
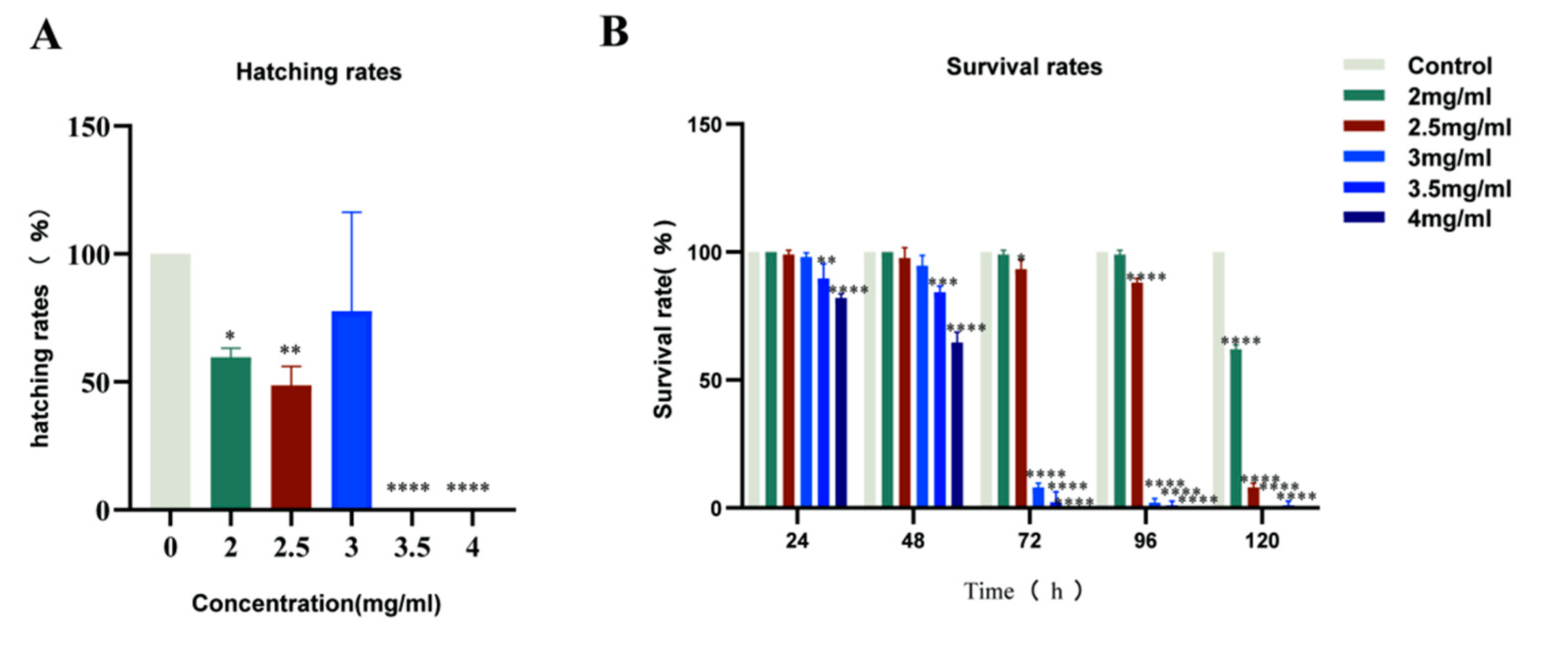
2.1. Zebrafish Mortality Effects
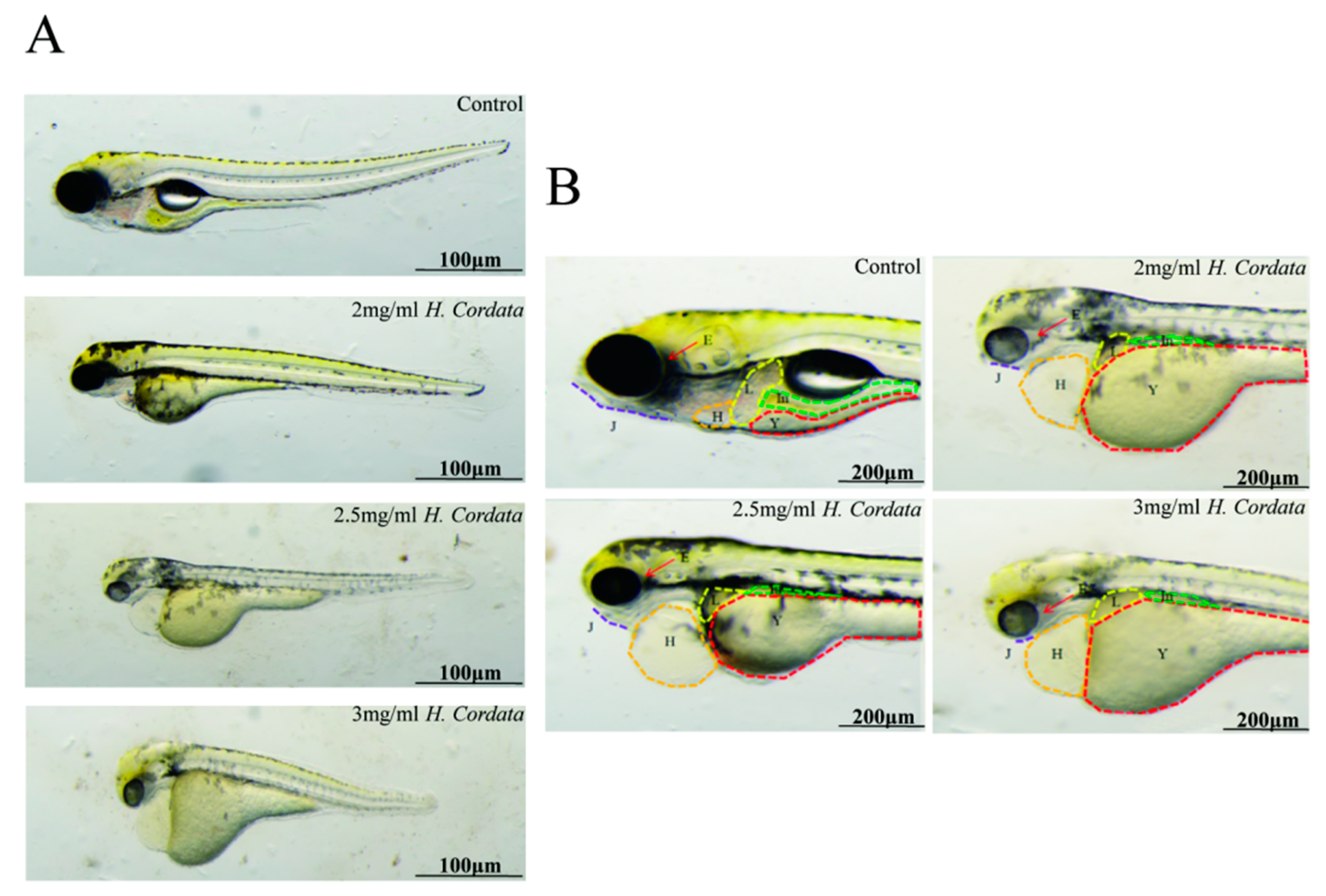
2.2. Zebrafish Hatching Rates and Morphological Deformities
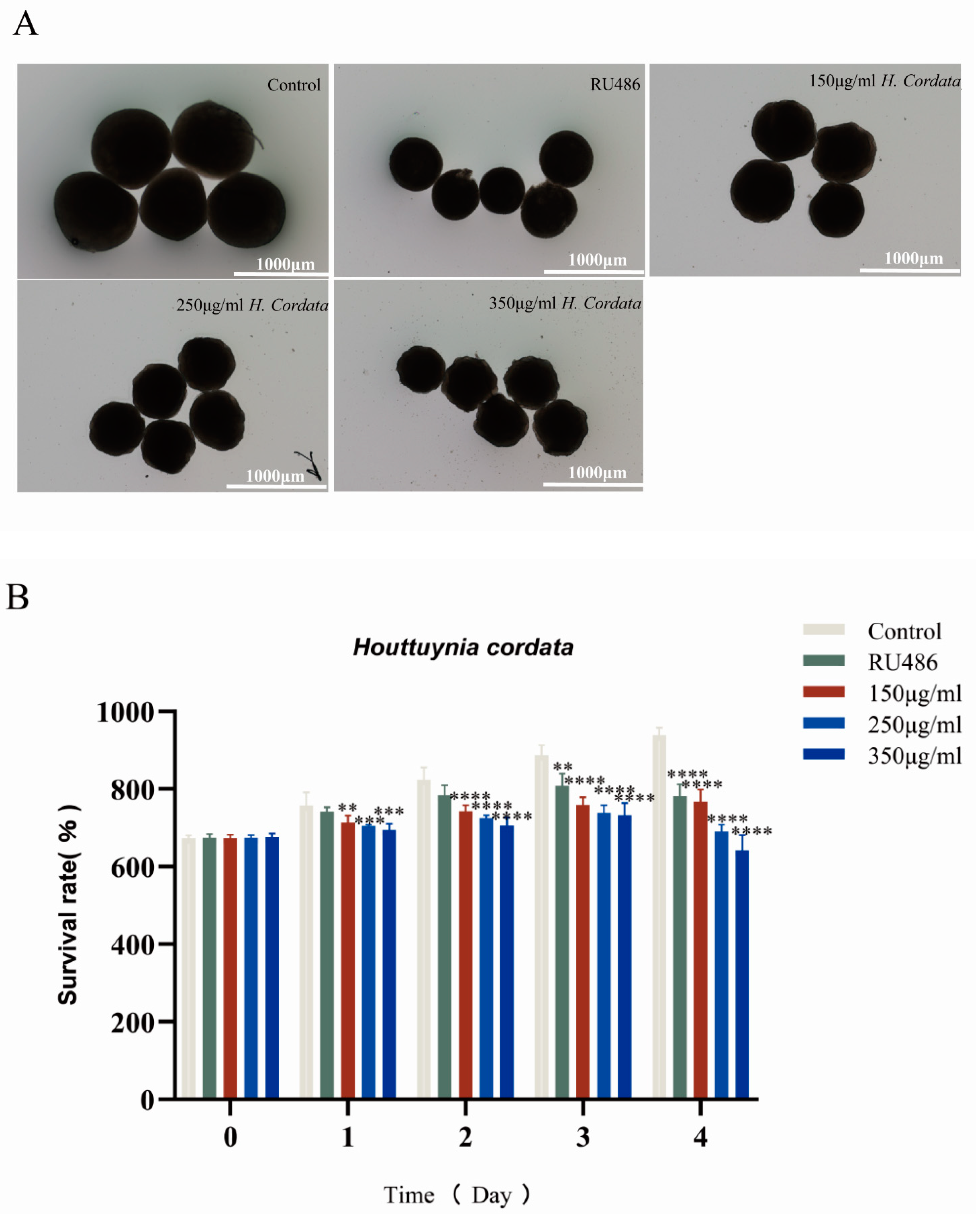
2.3. Embryoid Body Morphogenesis Is Impacted by H. cordata
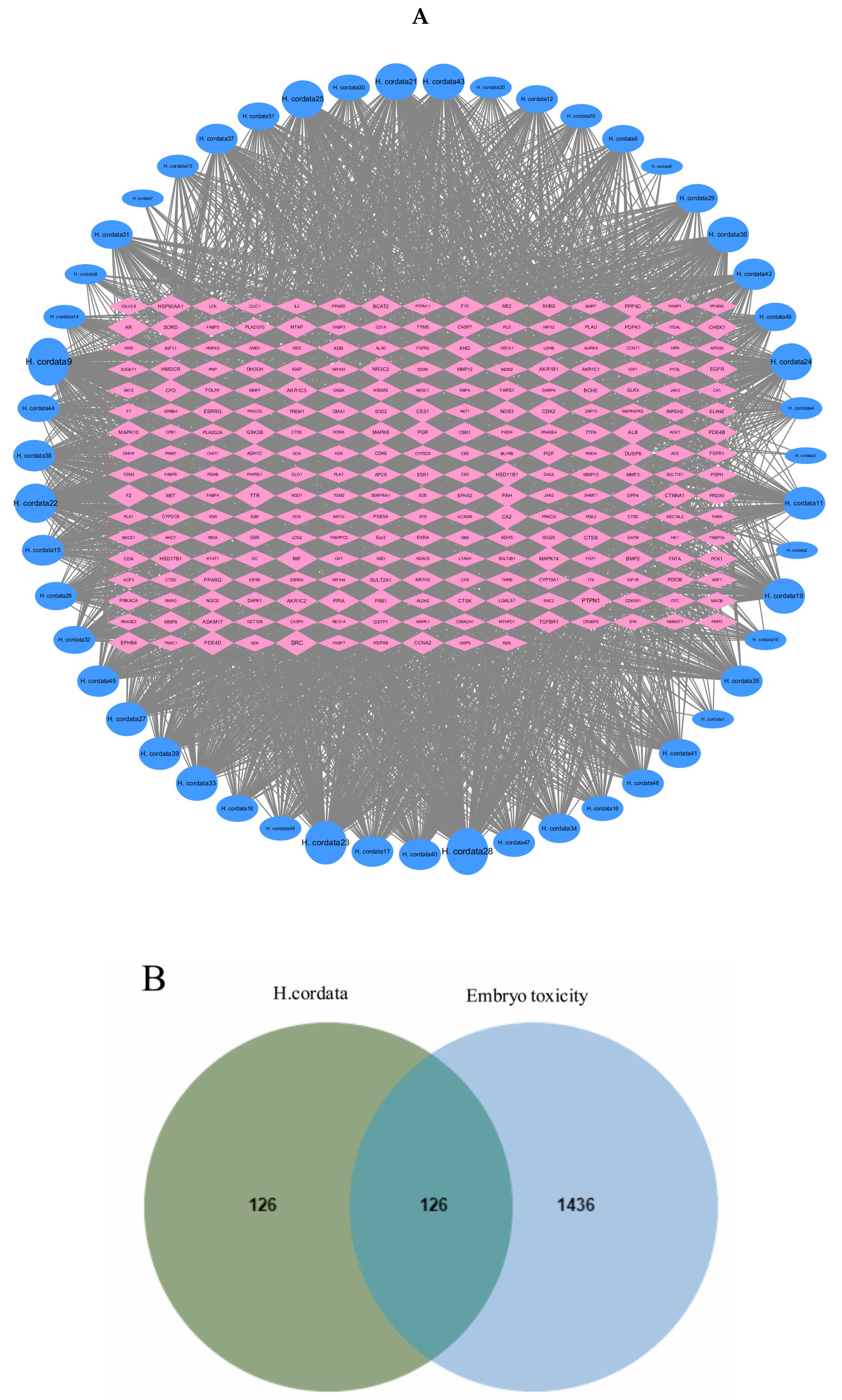
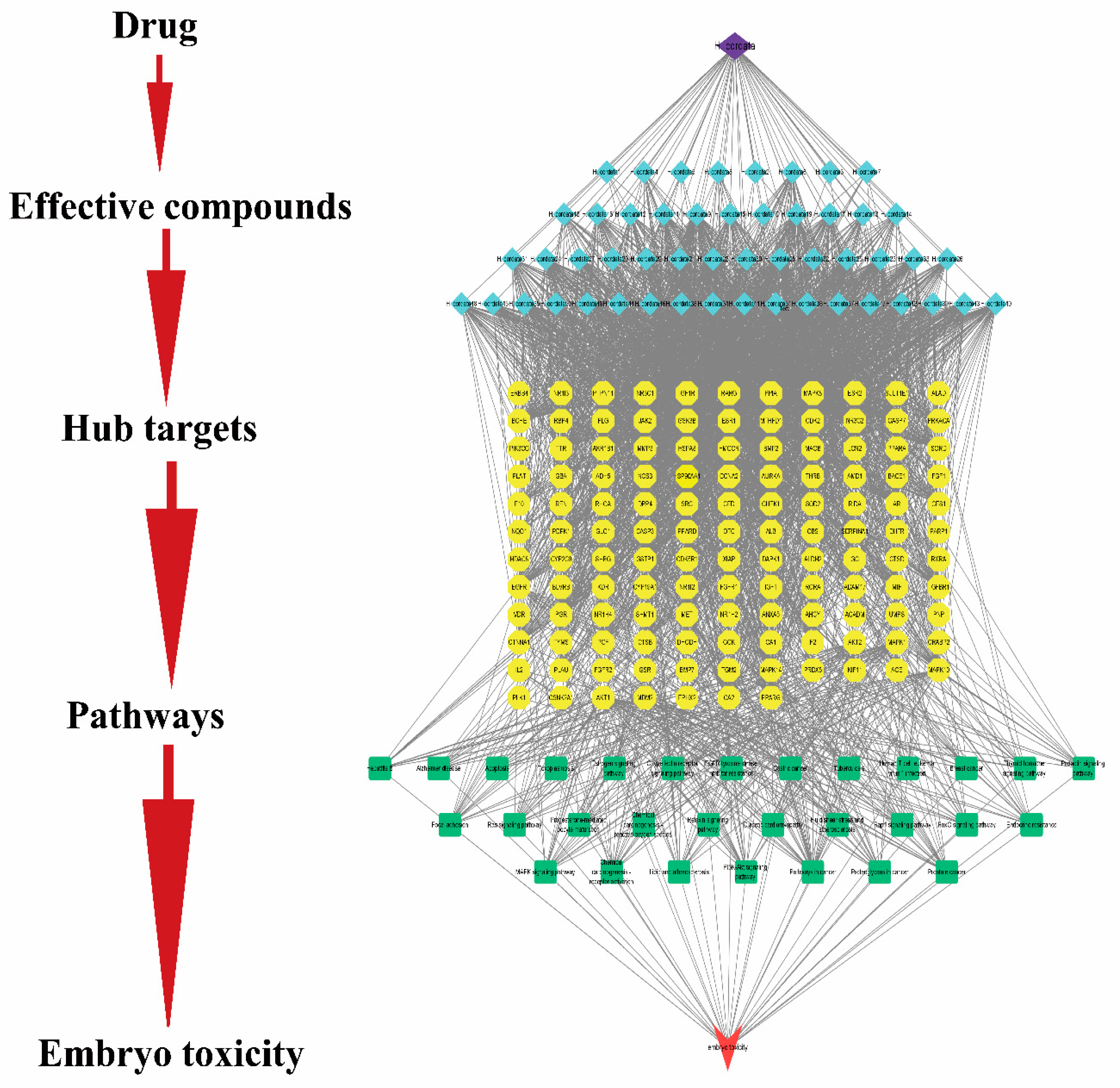
2.4. Active Compounds and Targets of H. cordata
2.5. Gene Targets in Embryotoxicity
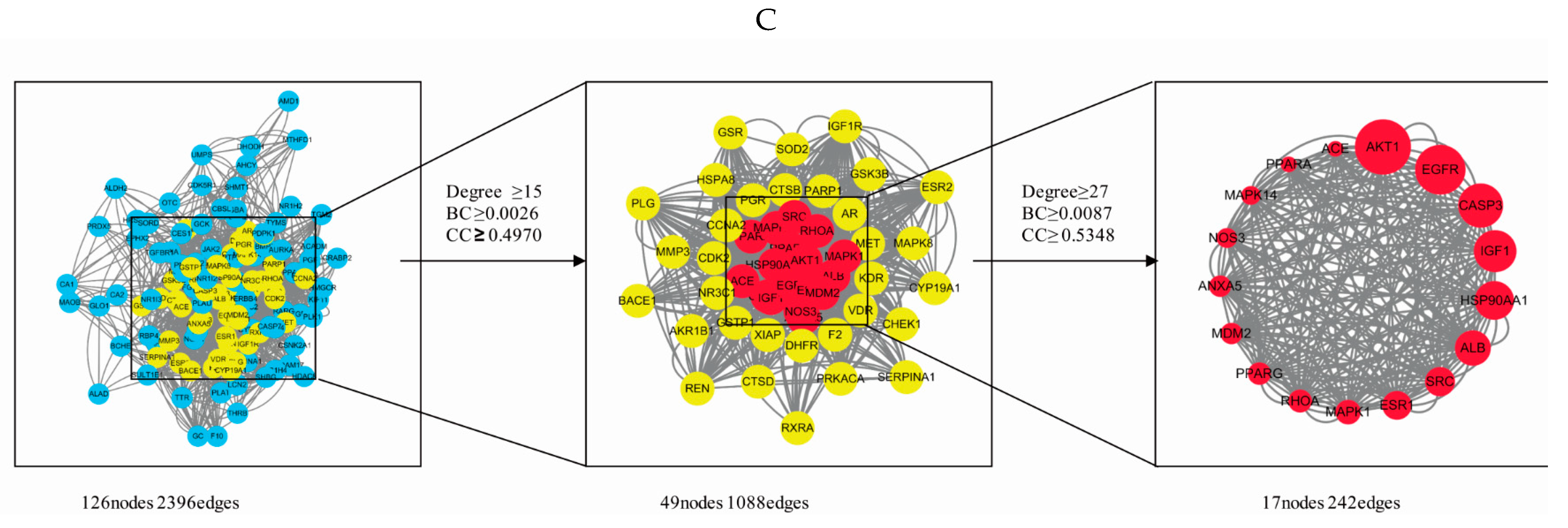
2.6. PPI Network Analysis
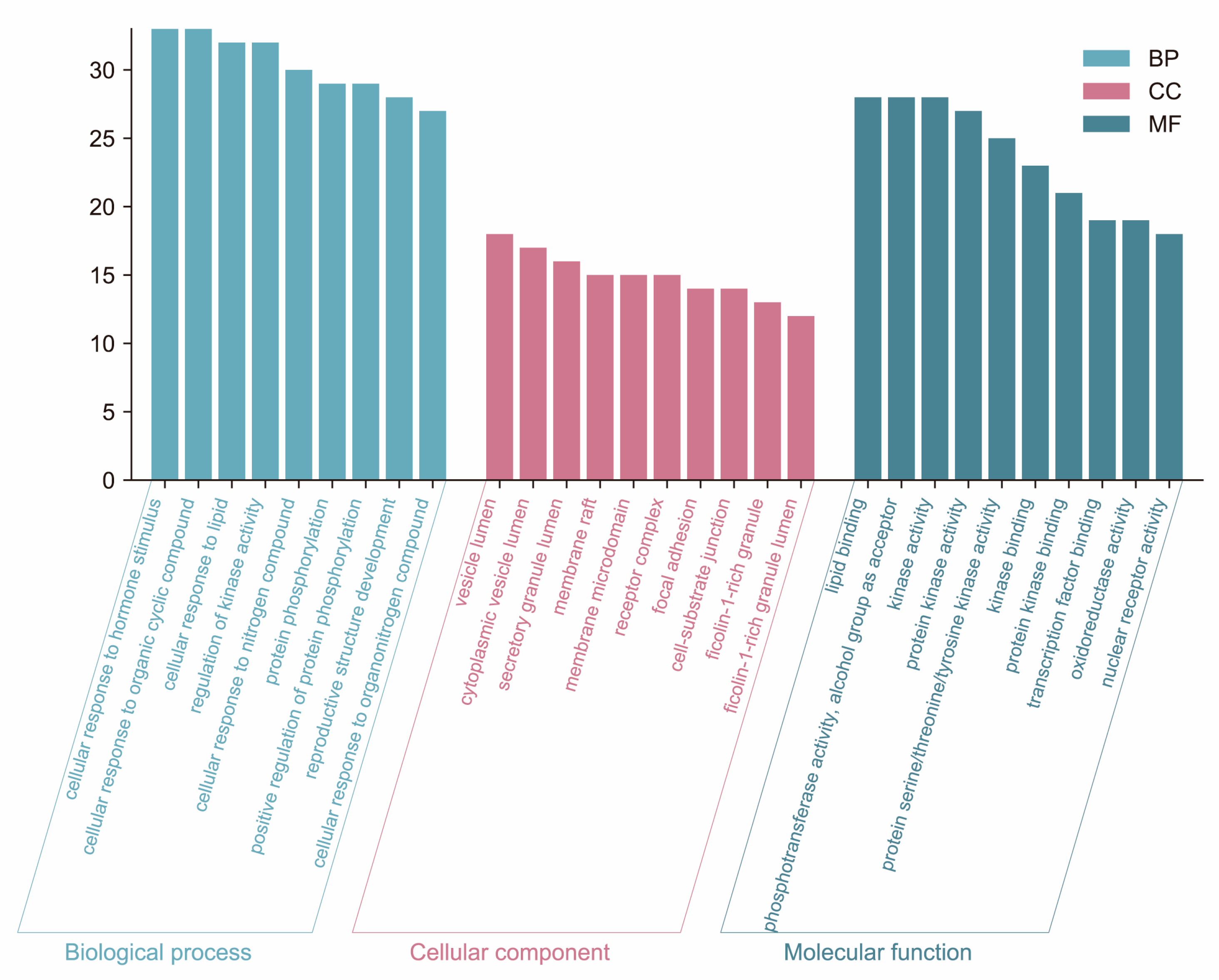
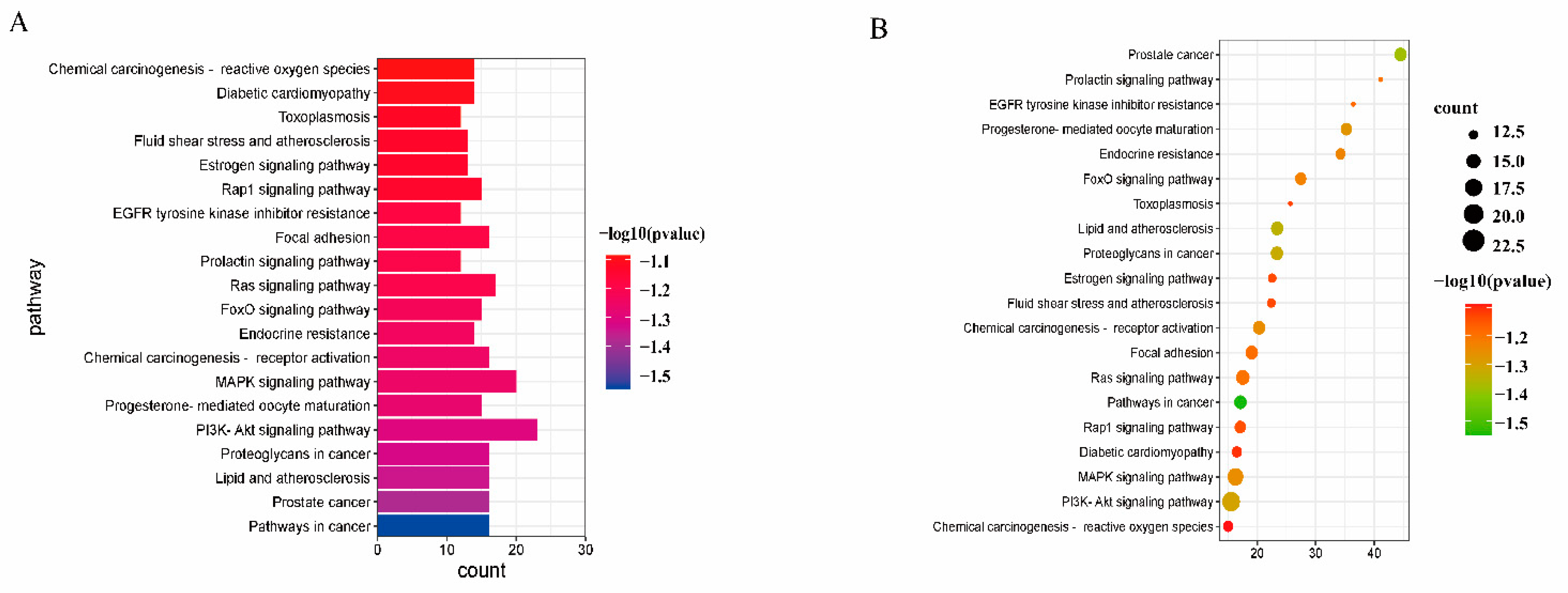
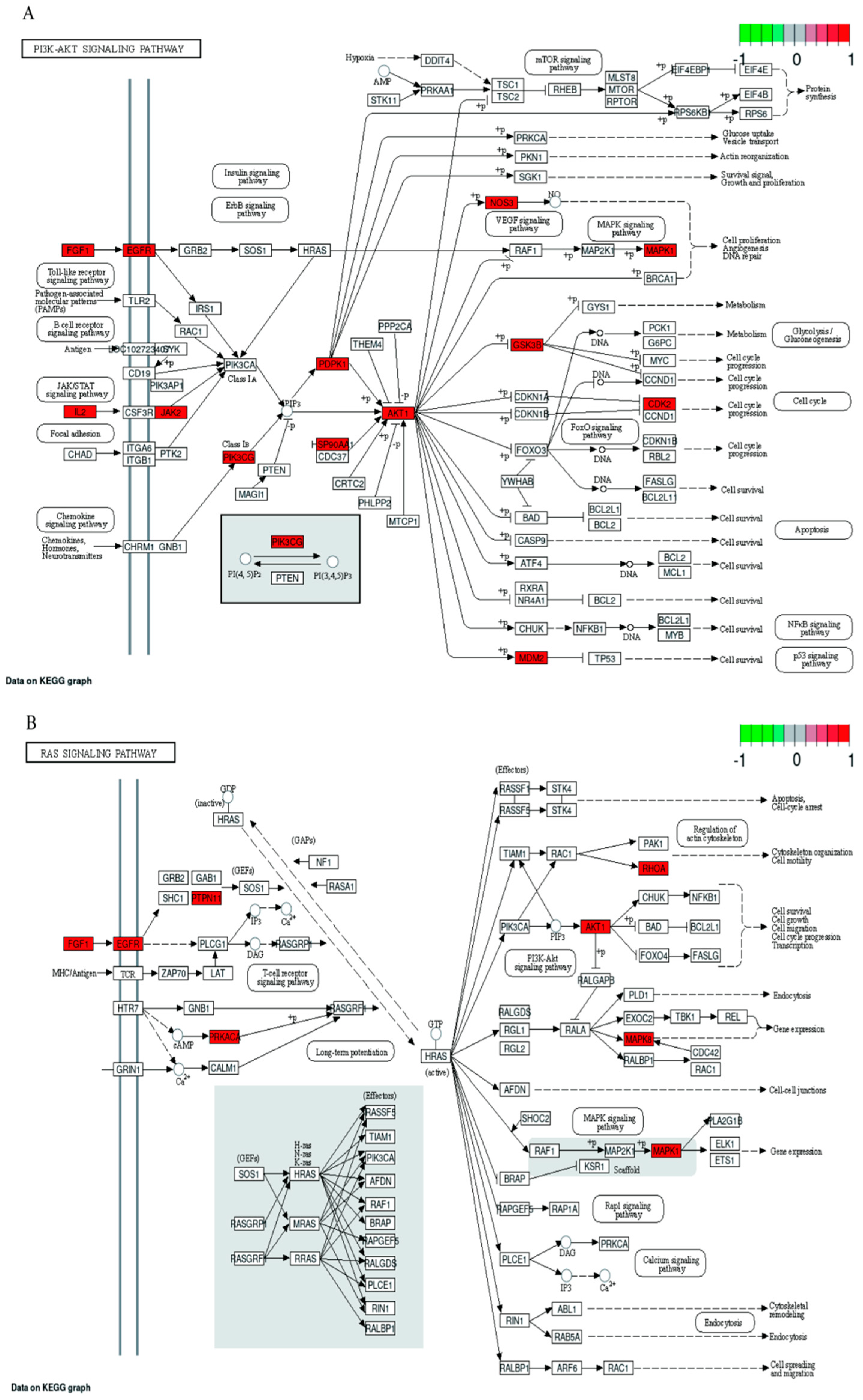
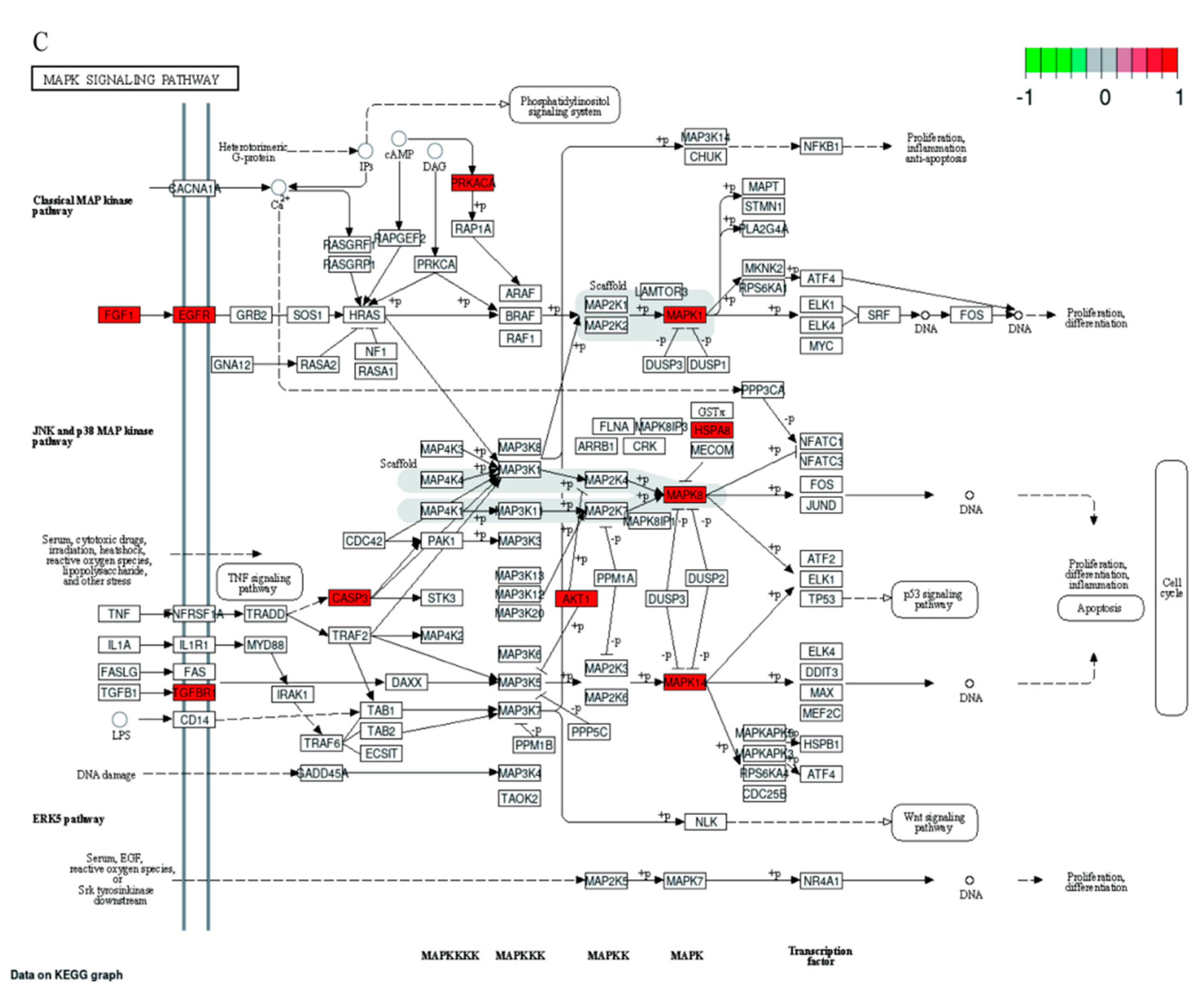
2.7. GO and KEGG Pathway Enrichment Analyses
2.8. Network Construction
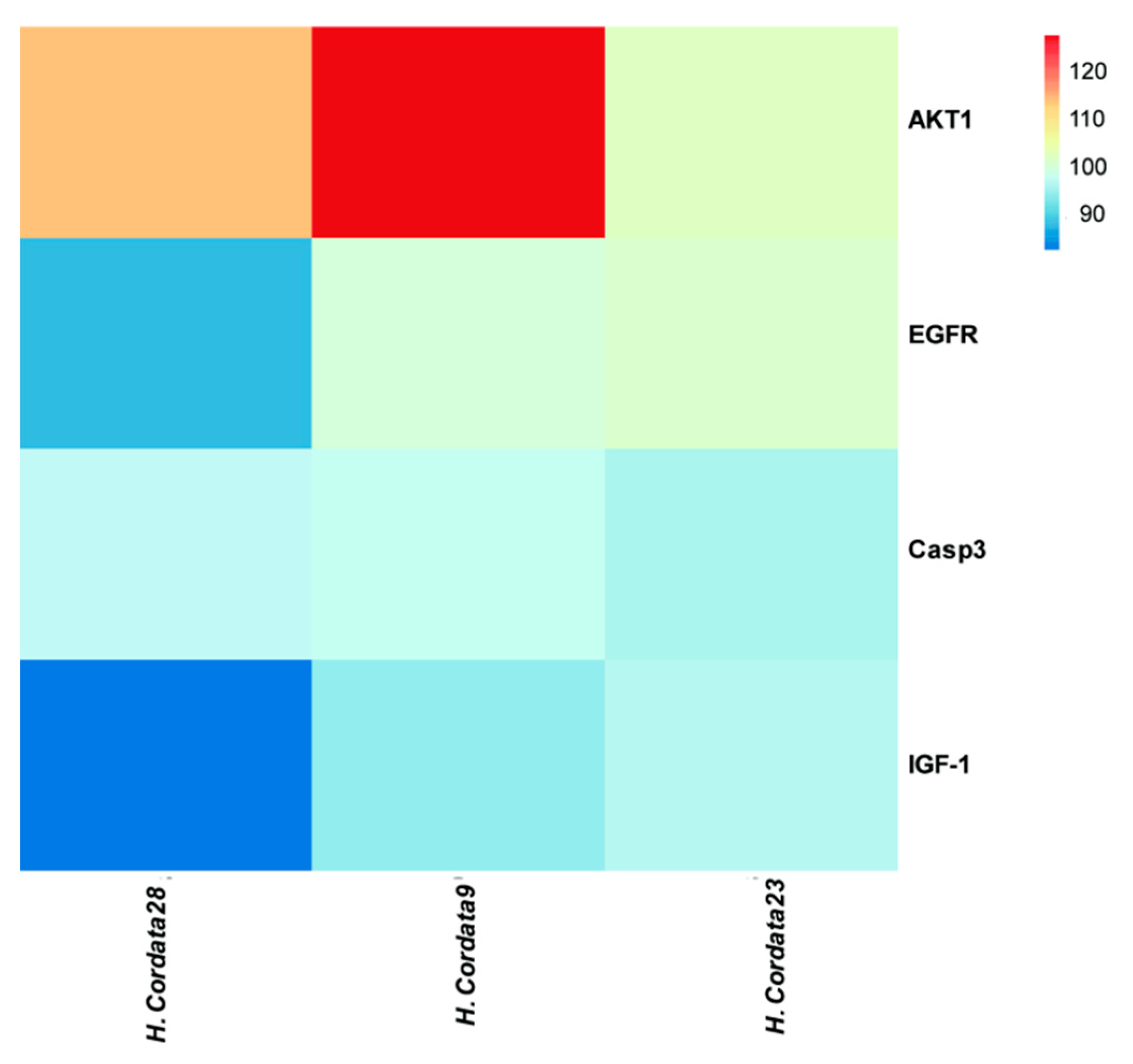
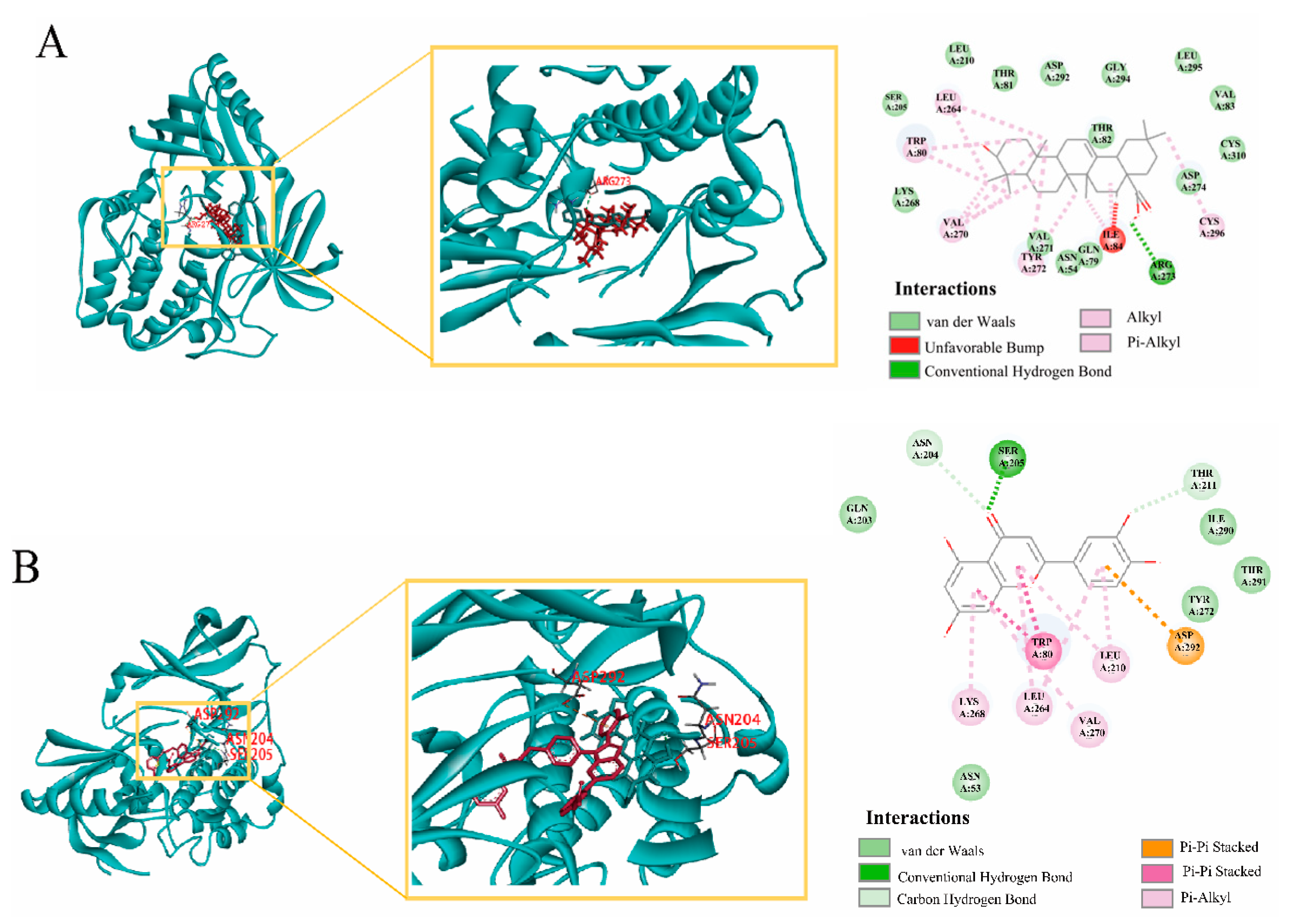
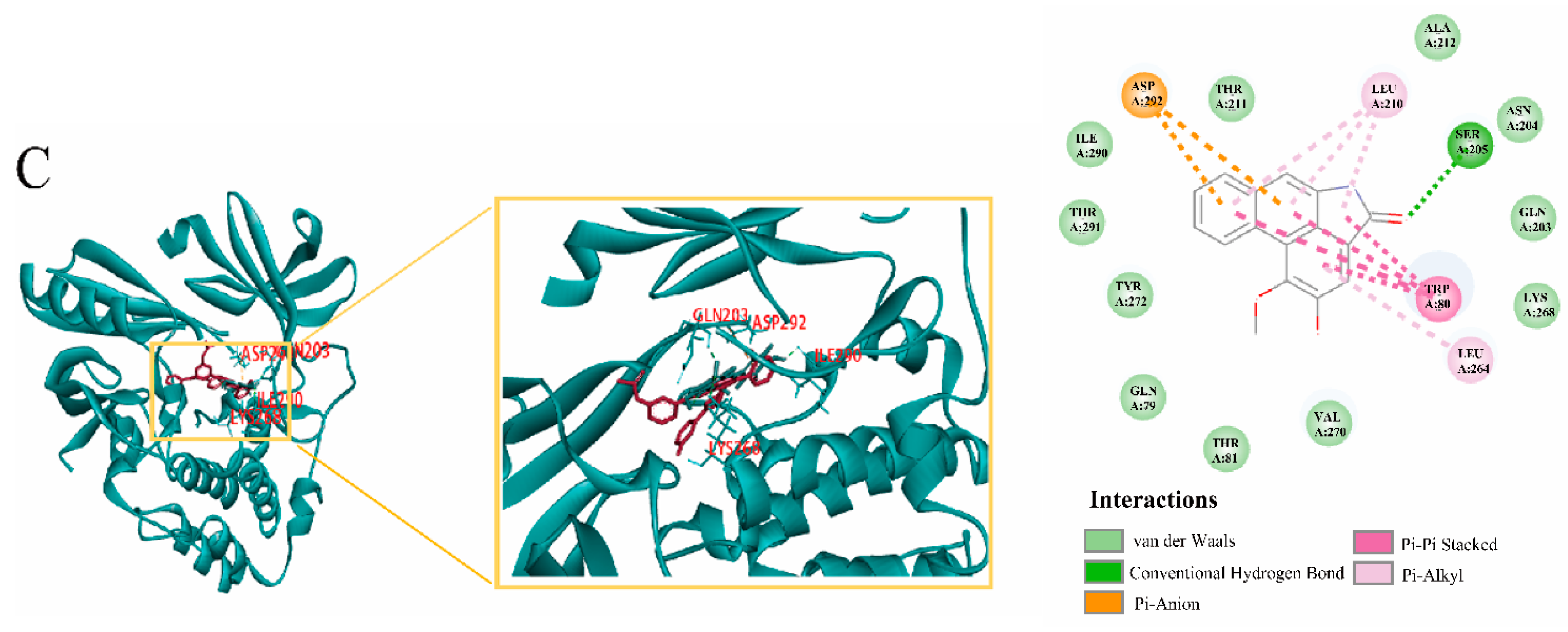
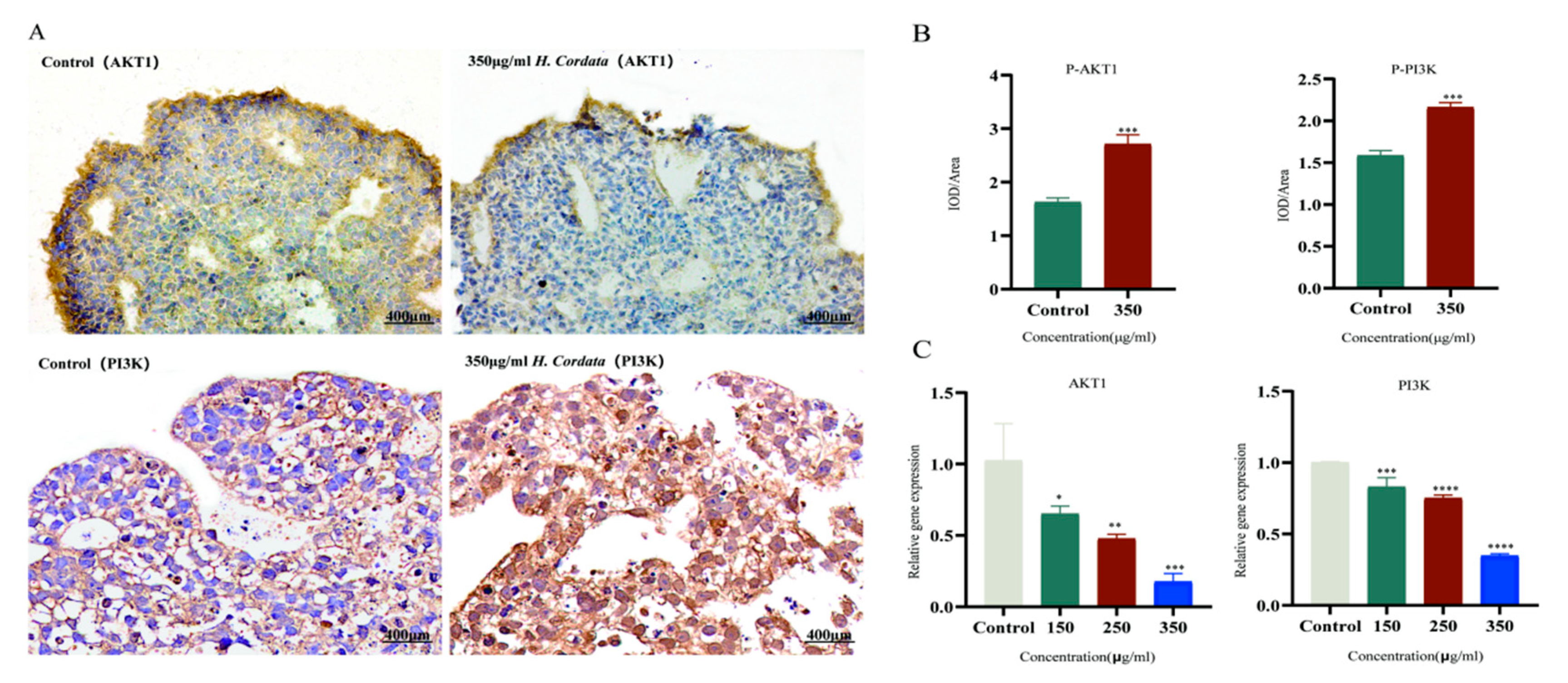
2.9. Molecular Docking Verification, Immunohistochemical and qRT-PCR
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Embryo Toxicity Test of Zebrafish
5.1.1. Test Compounds
5.1.2. Zebrafish Maintenance and Embryo Collection
5.1.3. Exposure of Embryos to Test Compounds
5.1.4. Acute Toxicity Assessment
5.2. Embryoid Body (EBs) Assay
5.2.1. Cell Culture and Embryoid Body Generation
5.2.2. Exposure of the Embryoid Body to Test Compounds
5.3. Exposure of the Embryoid Body to Test Compounds
5.4. Network Pharmacology
5.4.1. Bioactive Ingredients and Targets Screening of H. cordata
5.4.2. Toxicity Targets Acquisition of Embryotoxicity and Filtering Intersecting Targets
5.4.3. Protein-Protein Interaction (PPI) Network Construction
5.4.4. Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathway Enrichment Analyses
5.4.5. Network Construction
5.4.6. Molecular Docking
5.4.7. Immunohistochemistry
5.4.8. RNA Isolation and Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tian, Y.; Ding, Y.; Shao, B.; Yang, J.; Wu, J. Interaction between homologous functional food Astragali Radix and intestinal flora. Zhongguo Zhong Yao Za Zhi 2020, 45, 2486–2492. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Wang, S.; Zhang, T.; Sun, Y.; Han, L.; Banahene, P.O.; Wang, Q. Predicting the potential toxicity of 26 components in Cassiae semen using in silico and in vitro approaches. Curr. Res. Toxicol. 2021, 2, 237–245. [Google Scholar] [CrossRef]
- Jang, H.; Roh, S.; Jeong, E.; Kim, B.; Sunwoo, M. Ginkgotoxin Induced Seizure Caused by Vitamin B6 Deficiency. J. Epilepsy Res. 2015, 5, 104–106. [Google Scholar] [CrossRef] [PubMed]
- Galanjuk, S.; Zuhr, E.; Donmez, A.; Bartsch, D.; Kurian, L.; Tigges, J.; Fritsche, E. The Human Induced Pluripotent Stem Cell Test as an Alternative Method for Embryotoxicity Testing. Int. J. Mol. Sci. 2022, 23, 3295. [Google Scholar] [CrossRef]
- Wu, Z.; Deng, X.; Hu, Q.; Xiao, X.; Jiang, J.; Ma, X.; Wu, M. Houttuynia cordata Thunb: An Ethnopharmacological Review. Front. Pharmacol. 2021, 12, 714694. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Sha, X.; Luo, Y.; Chen, J.; Li, X.; Wang, J.; Cao, G.; Peng, X. Acute and subacute toxicity evaluation of Houttuynia cordata ethanol extract and plasma metabolic profiling analysis in both male and female rats. J. Appl. Toxicol. JAT 2021, 41, 2068–2082. [Google Scholar] [CrossRef]
- Pezzuto, J.M.; Swanson, S.M.; Mar, W.; Che, C.T.; Cordell, G.A.; Fong, H.H. Evaluation of the mutagenic and cytostatic potential of aristolochic acid (3,4-methylenedioxy-8-methoxy-10-nitrophenanthrene-1-carboxylic acid) and several of its derivatives. Mutat. Res. 1988, 206, 447–454. [Google Scholar] [CrossRef]
- Kim, J.M.; Hwang, I.H.; Jang, I.S.; Kim, M.; Bang, I.S.; Park, S.J.; Chung, Y.J.; Joo, J.C.; Lee, M.G. Houttuynia cordata Thunb Promotes Activation of HIF-1A-FOXO3 and MEF2A Pathways to Induce Apoptosis in Human HepG2 Hepatocellular Carcinoma Cells. Integr. Cancer Ther. 2017, 16, 360–372. [Google Scholar] [CrossRef]
- Miyata, M.; Koyama, T.; Yazawa, K. Water extract of Houttuynia cordata Thunb. Leaves exerts anti-obesity effects by inhibiting fatty acid and glycerol absorption. J. Nutr. Sci. Vitaminol 2010, 56, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.; Koppula, S. Hepatoprotective Effect of Houttuynia cordata Thunb Extract against Carbon Tetrachloride-induced Hepatic Damage in Mice. Indian J. Pharm. Sci. 2014, 76, 267–273. [Google Scholar] [PubMed]
- Ren, X.; Sui, X.; Yin, J. The effect of Houttuynia cordata injection on pseudorabies herpesvirus (PrV) infection in vitro. Pharm. Biol. 2011, 49, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.Q.; Marikawa, Y. Embryoid body test with morphological and molecular endpoints implicates potential developmental toxicity of trans-resveratrol. Toxicol. Appl. Pharmacol. 2018, 355, 211–225. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Yang, F.; Zhan, H.; Liu, B.; Cai, J.; Luo, Y.; Zhou, X. Houttuynia cordata Extract Ameliorates Bladder Damage and Improves Bladder Symptoms via Anti-Inflammatory Effect in Rats with Interstitial Cystitis. Evid.-Based Complement. Altern. Med. 2020, 2020, 9026901. [Google Scholar] [CrossRef] [PubMed]
- Li, G.Z.; Chai, O.H.; Lee, M.S.; Han, E.H.; Kim, H.T.; Song, C.H. Inhibitory effects of Houttuynia cordata water extracts on anaphylactic reaction and mast cell activation. Biol. Pharm. Bull. 2005, 28, 1864–1868. [Google Scholar] [CrossRef] [PubMed]
- Prommaban, A.; Kodchakorn, K.; Kongtawelert, P.; Banjerdpongchai, R. Houttuynia cordata Thunb fraction induces human leukemic Molt-4 cell apoptosis through the endoplasmic reticulum stress pathway. Asian Pac. J. Cancer Prev. 2012, 13, 1977–1981. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Kim, I.S.; Kim, J.H.; Kim, J.S.; Kim, D.H.; Yun, C.Y. Suppressive effects of Houttuynia cordata Thunb (Saururaceae) extract on Th2 immune response. J. Ethnopharmacol. 2008, 117, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Yang, J.; Liu, C.; He, M.; Yan, H. Complete plastome of Houttuynia cordata (Saururaceae), a medicinal and edible plant. Mitochondrial DNA B Resour. 2019, 4, 3208–3209. [Google Scholar] [CrossRef]
- Hopkins, A.L. Network pharmacology. Nat. Biotechnol. 2007, 25, 1110–1111. [Google Scholar] [CrossRef]
- Westerhoff, H.V. Network-based pharmacology through systems biology. Drug Discov. Today Technol. 2015, 15, 15–16. [Google Scholar] [CrossRef]
- Zhang, R.; Zhu, X.; Bai, H.; Ning, K. Network Pharmacology Databases for Traditional Chinese Medicine: Review and Assessment. Front. Pharmacol. 2019, 10, 123. [Google Scholar] [CrossRef]
- Sobanska, M.; Scholz, S.; Nyman, A.M.; Cesnaitis, R.; Gutierrez Alonso, S.; Kluver, N.; Kuhne, R.; Tyle, H.; de Knecht, J.; Dang, Z.; et al. Applicability of the fish embryo acute toxicity (FET) test (OECD 236) in the regulatory context of Registration, Evaluation, Authorisation, and Restriction of Chemicals (REACH). Environ. Toxicol. Chem. 2018, 37, 657–670. [Google Scholar] [CrossRef]
- Kim, H.; Jeong, H.; Hong, S.; Oh, M. Houttuyniae Herba Attenuates Kainic Acid-Induced Neurotoxicity via Calcium Response Modulation in the Mouse Hippocampus. Planta Med. 2015, 81, 1697–1704. [Google Scholar] [CrossRef]
- Shen, Y.; Cheng, M.; Liu, X.; Zhu, D.; Gao, J. Sodium Houttuyfonate Inhibits Bleomycin Induced Pulmonary Fibrosis in Mice. Front. Pharmacol. 2021, 12, 596492. [Google Scholar] [CrossRef]
- Colborn, T.; vom Saal, F.; Soto, A. Developmental effects of endocrine-disrupting chemicals in wildlife and humans. Environ. Health Perspect. 1993, 101, 378–384. [Google Scholar] [CrossRef]
- Liu, J.; Lu, Y.F.; Wu, Q.; Xu, S.F.; Shi, F.G.; Klaassen, C.D. Oleanolic acid reprograms the liver to protect against hepatotoxicants, but is hepatotoxic at high doses. Liver Int. Off. J. Int. Assoc. Study Liver 2019, 39, 427–439. [Google Scholar] [CrossRef]
- Kong, D.; Xing, L.; Liu, R.; Jiang, J.; Wang, W.; Shang, L.; Wei, X.; Hao, W. Individual and combined developmental toxicity assessment of bisphenol A and genistein using the embryonic stem cell test in vitro. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2013, 60, 497–505. [Google Scholar] [CrossRef] [PubMed]
- Evain-Brion, D.; Alsat, E. Epidermal growth factor receptor and human fetoplacental development. J. Pediatr. Endocrinol. 1994, 7, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Gong, Y. LncRNA-TCL6 promotes early abortion and inhibits placenta implantation via the EGFR pathway. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 7105–7112. [Google Scholar] [CrossRef]
- Li, Y.; Wang, Z.; Yan, X.; Chen, M.; Bao, J.; Wu, G.; Ge, Z.; Zhou, D.; Wang, Y.; Li, R. IC-4, a new irreversible EGFR inhibitor, exhibits prominent anti-tumor and anti-angiogenesis activities. Cancer Lett. 2013, 340, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Ao, R.; Liang, Y.; Liu, X.; Tan, K.; Wang, X.; Liu, D.; Zhang, T.; Sun, G.; Xie, J. Stavudine exposure results in developmental abnormalities by causing DNA damage, inhibiting cell proliferation and inducing apoptosis in mouse embryos. Toxicology 2020, 439, 152443. [Google Scholar] [CrossRef]
- Liu, M.; Chen, H.; Dai, H.; Zhou, L.; Wang, Y.; Xin, X.; Chen, C.; Li, Z.; Ge, R. Effects of bis(2-butoxyethyl) phthalate exposure in utero on the development of fetal Leydig cells in rats. Toxicol. Lett. 2021, 351, 65–77. [Google Scholar] [CrossRef] [PubMed]
- Pantaleon, M.; Kaye, P. IGF-I and insulin regulate glucose transport in mouse blastocysts via IGF-I receptor. Mol. Reprod. Dev. 1996, 44, 71–76. [Google Scholar] [CrossRef]
- Harvey, M.; Kaye, P. Mediation of the actions of insulin and insulin-like growth factor-1 on preimplantation mouse embryos in vitro. Mol. Reprod. Dev. 1992, 33, 270–275. [Google Scholar] [CrossRef]
- Liu, Z.; Shangguan, Y.; Zhu, P.; Sultan, Y.; Feng, Y.; Li, X.; Ma, J. Developmental toxicity of glyphosate on embryo-larval zebrafish (Danio rerio). Ecotoxicol. Environ. Saf. 2022, 236, 113493. [Google Scholar] [CrossRef]
- Ye, Y.; Jiang, S.; Du, T.; Ding, M.; Hou, M.; Mi, C.; Liang, T.; Zhong, H.; Xie, J.; Xu, W.; et al. Environmental Pollutant Benzo[a]pyrene Upregulated Long Non-coding RNA HZ07 Inhibits Trophoblast Cell Migration by Inactivating PI3K/AKT/MMP2 Signaling Pathway in Recurrent Pregnancy Loss. Reprod. Sci. 2021, 28, 3085–3093. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Meng, X.; Feng, C.; Xiao, J.; Zhao, X.; Xiong, B.; Feng, J. Benzophenone-3 induced abnormal development of enteric nervous system in zebrafish through MAPK/ERK signaling pathway. Chemosphere 2021, 280, 130670. [Google Scholar] [CrossRef]
- Barroso, R.; Osuamkpe, C.; Nagamani, M.; Yallampalli, C. Nitric oxide inhibits development of embryos and implantation in mice. Mol. Hum. Reprod. 1998, 4, 503–507. [Google Scholar] [CrossRef] [PubMed]
- Tshering, G.; Plengsuriyakarn, T.; Na-Bangchang, K.; Pimtong, W. Embryotoxicity evaluation of atractylodin and beta-eudesmol using the zebrafish model. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2021, 239, 108869. [Google Scholar] [CrossRef]
- Wang, H.; Meng, Z.; Liu, F.; Zhou, L.; Su, M.; Meng, Y.; Zhang, S.; Liao, X.; Cao, Z.; Lu, H. Characterization of boscalid-induced oxidative stress and neurodevelopmental toxicity in zebrafish embryos. Chemosphere 2020, 238, 124753. [Google Scholar] [CrossRef]
- Duan, M.; Zhang, J.; Liu, J.; Qian, L.; Chen, X.; Zhao, F.; Zhao, W.; Zhong, Z.; Yang, Y.; Wang, C. Toxic effects of broflanilide exposure on development of zebrafish (Danio rerio) embryos and its potential cardiotoxicity mechanism. Environ. Pollut. 2021, 286, 117481. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.S. Construction of an abortion toxicity drug screening model based on induced pluripotent stem cells. Acta Pharm. Sin. 2021, 56, 503–510. [Google Scholar] [CrossRef]
- Yuan, C.J.; Marikawa, Y. Developmental toxicity assessment of common excipients using a stem cell-based in vitro morphogenesis model. Food Chem. Toxicol. 2017, 109, 376–385. [Google Scholar] [CrossRef]
- Kang, H.Y.; Choi, Y.K.; Jo, N.R.; Lee, J.H.; Ahn, C.; Ahn, I.Y.; Kim, T.S.; Kim, K.S.; Choi, K.C.; Lee, J.K.; et al. Advanced developmental toxicity test method based on embryoid body’s area. Reprod. Toxicol. 2017, 72, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.-W.; Deng, Y.-L.; Zhang, X.; Liu, M.-M.; Liu, Y.; Xie, Y.-T.; Wan, Q.; Huang, M.; Zhang, T.; Xi, J.-H.; et al. Study on the mechanism of treating COVID-19 with Shenqi Wan based on network pharmacology. Drug Dev. Ind. Pharm. 2021, 47, 1279–1289. [Google Scholar] [CrossRef] [PubMed]
- Hou, F.; Liu, Y.; Cheng, Y.; Zhang, N.; Yan, W.; Zhang, F. Exploring the Mechanism of Scutellaria baicalensis Georgi Efficacy against Oral Squamous Cell Carcinoma Based on Network Pharmacology and Molecular Docking Analysis. Evid.-Based Complement. Altern. Med. 2021, 2021, 5597586. [Google Scholar] [CrossRef]
- Zhong, Z.; Guo, X.; Zheng, Y. Network Pharmacology-Based and Molecular Docking Analysis of Resveratrol’s Pharmacological Effects on Type I Endometrial Cancer. Anti-Cancer Agents Med. Chem. 2022, 22, 1933–1944. [Google Scholar] [CrossRef]
- Gao, Q.; Tian, D.; Han, Z.; Lin, J.; Chang, Z.; Zhang, D.; Ma, D. Network Pharmacology and Molecular Docking Analysis on Molecular Targets and Mechanisms of Buyang Huanwu Decoction in the Treatment of Ischemic Stroke. Evid.-Based Complement. Altern. Med. Ecam 2021, 2021, 8815447. [Google Scholar] [CrossRef]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Compounds |
|---|---|
| H. cordata1 | strigol |
| H. cordata2 | Sesamin |
| H. cordata3 | apigenin |
| H. cordata4 | Scutellarein |
| H. cordata5 | Genistein |
| H. cordata6 | pilloin |
| H. cordata7 | Lauric acid |
| H. cordata8 | Methyl ferulate |
| H. cordata9 | Luteolin |
| H. cordata10 | piperolactam B |
| H. cordata11 | piperolactam C |
| H. cordata12 | piperolactam D |
| H. cordata13 | 4-amino-N-(2-phenylethyl)benzamide |
| H. cordata14 | asimilobine |
| H. cordata15 | Norisoboldine |
| H. cordata16 | laetanine |
| H. cordata17 | Liriodenine |
| H. cordata18 | N-Methylasimilobine |
| H. cordata19 | perlolyrine |
| H. cordata20 | Vomifoliol |
| H. cordata21 | acantrifoside E |
| H. cordata22 | Epigensexinmethyl ether |
| H. cordata23 | Aristolochia lactam AII |
| H. cordata24 | strigone |
| H. cordata25 | Kaempferol |
| H. cordata26 | sorgomol |
| H. cordata27 | 5-deoxystrigol |
| H. cordata28 | oleanolic acid |
| H. cordata29 | Phenyl β-D-glucopyranoside |
| H. cordata30 | 3-hydroxy-1,2-dimethoxy-5-methyl-5H-dibenzoindol-4-one |
| H. cordata31 | 3-methoxy-5-methyl-5H-benzodioxolo-benzoindol-4-one |
| H. cordata32 | 1,2,3,4,5-pentamethoxy-dibenzo-quinolin-7-one |
| H. cordata33 | CARBOSTYRIL,4-HYDROXY-1-METHYL-3-[[P-(PHENYLAZO)PHENYL]AZO]-(7CI) |
| H. cordata34 | 7-oxodehydroasimilobine |
| H. cordata35 | lysicamine |
| H. cordata36 | atherospermidine |
| H. cordata37 | 3-methoxy-6H-benzodioxolo-benzoquinoline-4,5-dione |
| H. cordata38 | 3-methoxy-6-methyl-6H-benzodioxolo-benzoquinoline-4,5-dione |
| H. cordata | ouregidione |
| H. cordata40 | cepharadioneB |
| H. cordata41 | cepharadione A |
| H. cordata42 | 1,2,3-trimethoxy-4H,6H-dibenzoquinolin-5-one |
| H. cordata43 | 1,2,3-trimethoxy-6-methyl-4H,6H-dibenzoquinolin-5-one |
| H. cordata44 | 1,2-dimethoxy-3-hydroxy-5-oxonoraporphine |
| H. cordata45 | 1,2,3-trimethoxy-3-hydroxy-5-oxonoraporphine |
| H. cordata46 | sauristolactam |
| H. cordata47 | cepharanone B |
| H. cordata48 | Aristolactam A |
| H. cordata49 | Aristolactam B |
| H. cordata50 | AristolactamBⅡ |
| H. cordata51 | Isoramanone |
| Core Components | Key Targets | Docking Score |
|---|---|---|
| oleanolic acid | AKT1 | 113.486 |
| Luteolin | AKT1 | 126.616 |
| Aristolochia lactam AII | AKT1 | 102.669 |
| oleanolic acid | EGFR | 89.5131 |
| Luteolin | EGFR | 100.632 |
| Aristolochia lactam AII | EGFR | 101.622 |
| oleanolic acid | CASP3 | 97.9379 |
| Luteolin | CASP3 | 98.2926 |
| Aristolochia lactam AII | CASP3 | 96.8157 |
| oleanolic acid | IGF-1 | 84.2265 |
| Luteolin | IGF-1 | 95.4801 |
| Aristolochia lactam AII | IGF-1 | 97.4921 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Yang, G.; Yang, C.; Shi, Z.; Ru, Y.; Shen, N.; Xiao, C.; Wang, Y.; Gao, Y. The Mechanism of Houttuynia cordata Embryotoxicity Was Explored in Combination with an Experimental Model and Network Pharmacology. Toxins 2023, 15, 73. https://doi.org/10.3390/toxins15010073
Liu Y, Yang G, Yang C, Shi Z, Ru Y, Shen N, Xiao C, Wang Y, Gao Y. The Mechanism of Houttuynia cordata Embryotoxicity Was Explored in Combination with an Experimental Model and Network Pharmacology. Toxins. 2023; 15(1):73. https://doi.org/10.3390/toxins15010073
Chicago/Turabian StyleLiu, Yufu, Guodong Yang, Chunqi Yang, Zhuo Shi, Yi Ru, Ningning Shen, Chengrong Xiao, Yuguang Wang, and Yue Gao. 2023. "The Mechanism of Houttuynia cordata Embryotoxicity Was Explored in Combination with an Experimental Model and Network Pharmacology" Toxins 15, no. 1: 73. https://doi.org/10.3390/toxins15010073
APA StyleLiu, Y., Yang, G., Yang, C., Shi, Z., Ru, Y., Shen, N., Xiao, C., Wang, Y., & Gao, Y. (2023). The Mechanism of Houttuynia cordata Embryotoxicity Was Explored in Combination with an Experimental Model and Network Pharmacology. Toxins, 15(1), 73. https://doi.org/10.3390/toxins15010073